Characterization of Cetacean Proline-Rich Antimicrobial Peptides Displaying Activity against ESKAPE Pathogens

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Identification of New Cetacean PrAMPs
2.2. Antibacterial Activity—Membrane Directed or Internal?
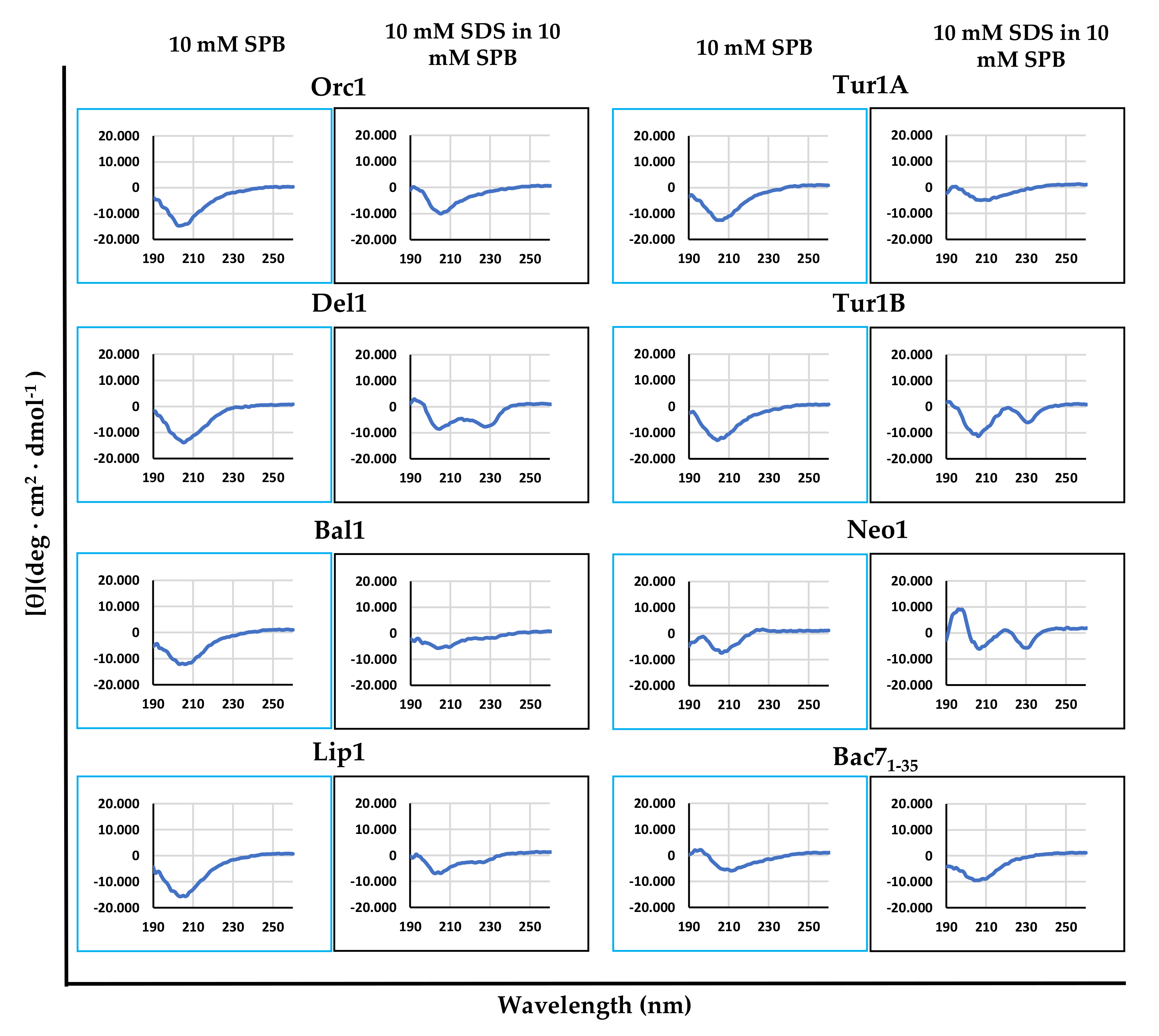
2.3. Dependence of cePrAMP Conformation on the Environment
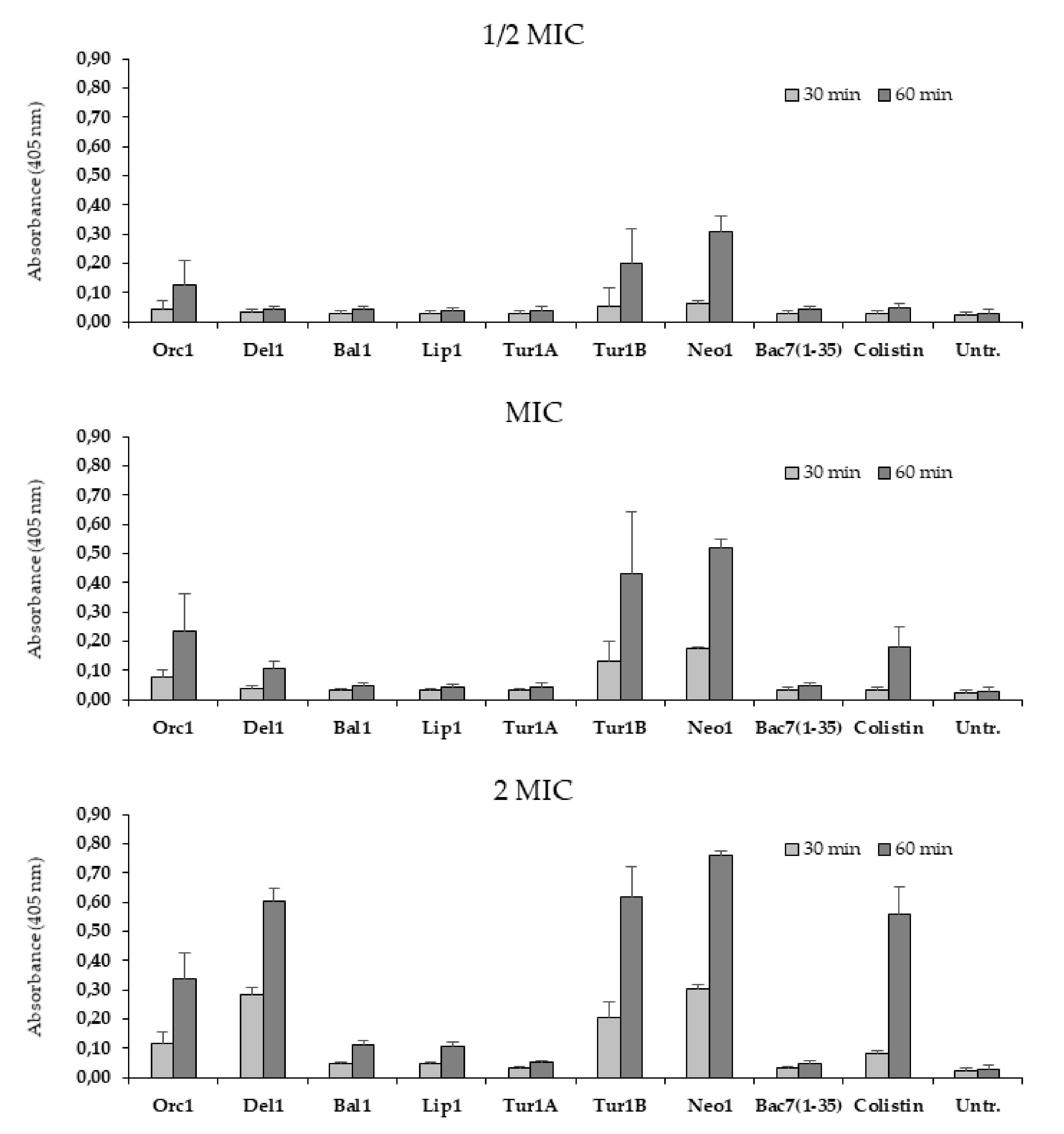
2.4. Membrane Permeabilization
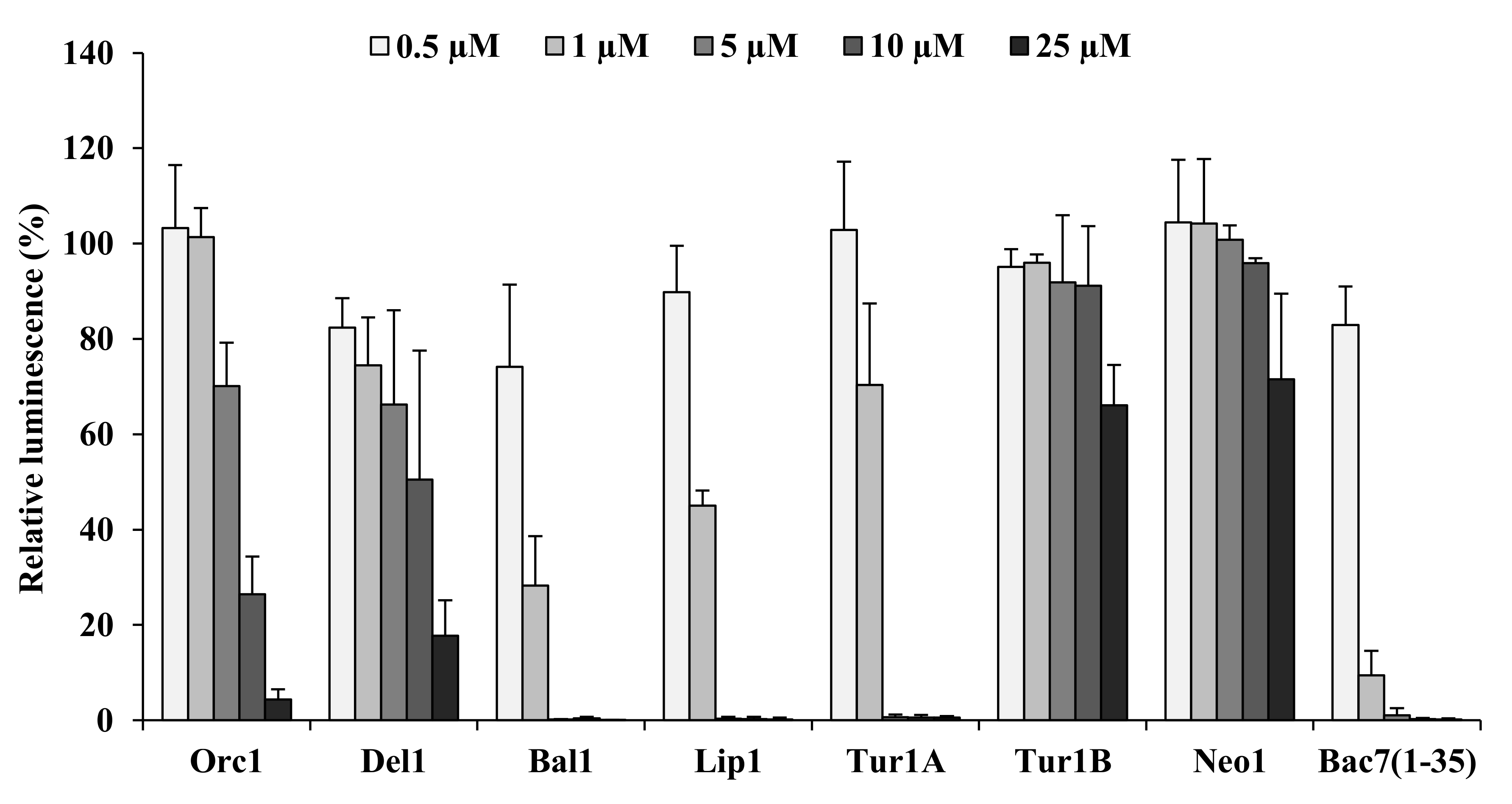
2.5. Inhibition of Bacterial Protein Synthesis
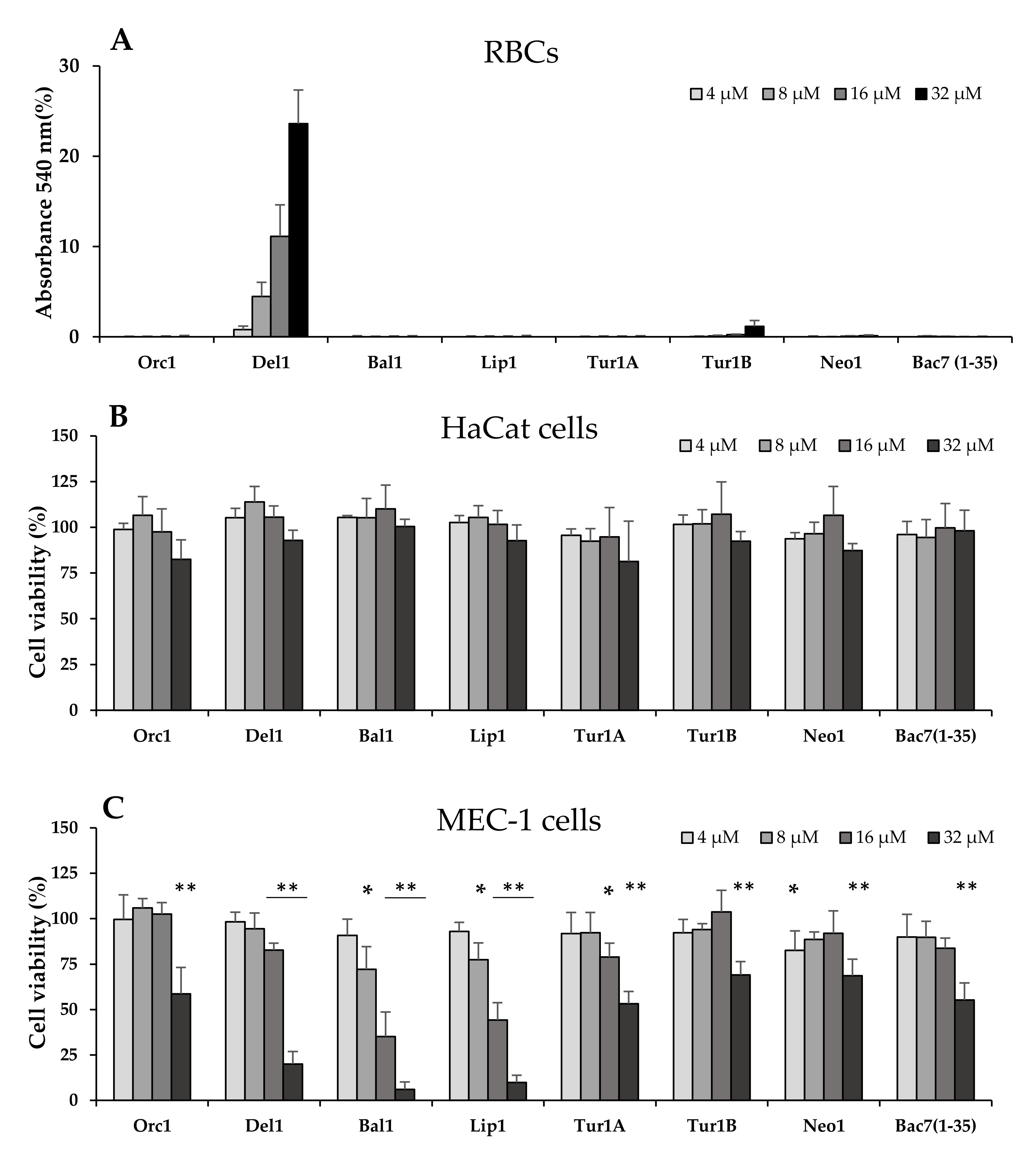
2.6. Evaluation of Potential Hemolytic and Cytotoxic Effects on Eukaryotic Cells
3. Discussion
4. Materials and Methods
4.1. Peptides
4.2. Microbial Strains
4.3. Circular Dichroism (CD) Spectra
4.4. Minimum Inhibitory Concentration (MIC) Determination
4.5. Assessment of Bacterial Membrane Permeabilization
4.6. In Vitro Transcription/Translation Assay
4.7. Determination of Cytotoxicity
4.7.1. Hemolysis Assay
4.7.2. MTT Cytotoxicity Assay
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AMPs | Antimicrobial peptides |
| ESKAPE | Enterococcus faecium, Staphylococcus aureus, Klebsiella pneumoniae, Acinetobacter baumannii, Pseudomonas aeruginosa, Enterobacter spp. |
| cePrAMPs | Cetacean proline-rich antimicrobial peptides |
| GRAVY | Grand average of hydropathy |
| ESTs | Expressed sequence tags |
| MIC | Minimum Inhibitory concentration |
| CD | Circular dichroism |
| SPB | Sodium phosphate buffer |
| SDS | Sodium dodecylsulphate |
| ONPG | O-Nitrophenyl-β-D-galactopyranoside |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium (bromide) |
References
- Ventola, C.L. The antibiotic resistance crisis—Part 1: Causes and threats. Pharm. Ther. 2015, 40, 277–283. [Google Scholar]
- Mulani, M.S.; Kamble, E.E.; Kumkar, S.N.; Tawre, M.S.; Pardesi, K.R. Emerging strategies to combat ESKAPE pathogens in the era of antimicrobial resistance: A review. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef]
- Ma, Y.-X.; Wang, C.-Y.; Li, Y.-Y.; Li, J.; Wan, Q.-Q.; Chen, J.-H.; Tay, F.R.; Niu, L.-N. Considerations and caveats in combating eskape pathogens against nosocomial infections. Adv. Sci. 2020, 7, 1901872. [Google Scholar] [CrossRef] [Green Version]
- Theuretzbacher, U.; Outterson, K.; Engel, A.; Karlén, A. The global preclinical antibacterial pipeline. Nat. Rev. Microbiol. 2020, 18, 275–285. [Google Scholar] [CrossRef] [Green Version]
- Pacios, O.; Blasco, L.; Bleriot, I.; Fernandez-Garcia, L.; González Bardanca, M.; Ambroa, A.; López, M.; Bou, G.; Tomás, M. Strategies to combat multidrug-resistant and persistent infectious diseases. Antibiotics 2020, 9, 65. [Google Scholar] [CrossRef] [Green Version]
- Boto, A.; Pérez de la Lastra, J.M.; González, C.C. The road from host-defense peptides to a new generation of antimicrobial drugs. Molecules 2018, 23, 311. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.H.; Lu, T.K. Development and challenges of antimicrobial peptides for therapeutic applications. Antibiotics 2020, 9, 24. [Google Scholar] [CrossRef] [Green Version]
- Lei, J.; Sun, L.; Huang, S.; Zhu, C.; Li, P.; He, J.; Mackey, V.; Coy, D.H.; He, Q. The antimicrobial peptides and their potential clinical applications. Am. J. Transl. Res. 2019, 11, 3919–3931. [Google Scholar]
- Naafs, M.A.B. The antimicrobial peptides: Ready for clinical trials? BJSTR 2018, 7. [Google Scholar] [CrossRef] [Green Version]
- Wimley, W.C.; Hristova, K. Antimicrobial peptides: Successes, challenges and unanswered questions. J. Membr. Biol. 2011, 239, 27–34. [Google Scholar] [CrossRef] [Green Version]
- Pletzer, D.; Hancock, R.E.W. Antibiofilm peptides: Potential as broad-spectrum agents. J. Bacteriol. 2016, 198, 2572–2578. [Google Scholar] [CrossRef] [Green Version]
- Uhlig, T.; Kyprianou, T.; Martinelli, F.G.; Oppici, C.A.; Heiligers, D.; Hills, D.; Calvo, X.R.; Verhaert, P. The emergence of peptides in the pharmaceutical business: From exploration to exploitation. EuPA Open Proteom. 2014, 4, 58–69. [Google Scholar] [CrossRef] [Green Version]
- Hancock, R.E.W.; Sahl, H.-G. Antimicrobial and host-defense peptides as new anti-infective therapeutic strategies. Nat. Biotechnol. 2006, 24, 1551–1557. [Google Scholar] [CrossRef]
- Le, C.-F.; Fang, C.-M.; Sekaran, S.D. Intracellular Targeting mechanisms by antimicrobial peptides. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef] [Green Version]
- Scocchi, M.; Mardirossian, M.; Runti, G.; Benincasa, M. Non-membrane permeabilizing modes of action of antimicrobial peptides on bacteria. Curr. Top. Med. Chem. 2015, 16, 76–88. [Google Scholar] [CrossRef]
- Mattiuzzo, M.; Bandiera, A.; Gennaro, R.; Benincasa, M.; Pacor, S.; Antcheva, N.; Scocchi, M. Role of the escherichia coli sbma in the antimicrobial activity of proline-rich peptides. Mol. Microbiol. 2007, 66, 151–163. [Google Scholar] [CrossRef]
- Krizsan, A.; Knappe, D.; Hoffmann, R. Influence of the YjiL-MdtM gene cluster on the antibacterial activity of proline-rich antimicrobial peptides overcoming escherichia coli resistance induced by the missing sbma transporter system. Antimicrob. Agents Chemother. 2015, 59, 5992–5998. [Google Scholar] [CrossRef] [Green Version]
- Krizsan, A.; Volke, D.; Weinert, S.; Sträter, N.; Knappe, D.; Hoffmann, R. Insect-derived proline-rich antimicrobial peptides kill bacteria by inhibiting bacterial protein translation at the 70 s ribosome. Angew. Chem. Int. Edit. 2014, 53, 12236–12239. [Google Scholar] [CrossRef]
- Mardirossian, M.; Grzela, R.; Giglione, C.; Meinnel, T.; Gennaro, R.; Mergaert, P.; Scocchi, M. The host antimicrobial peptide Bac71-35 binds to bacterial ribosomal proteins and inhibits protein synthesis. Chem. Biol. 2014, 21, 1639–1647. [Google Scholar] [CrossRef] [Green Version]
- Mardirossian, M.; Barrière, Q.; Timchenko, T.; Müller, C.; Pacor, S.; Mergaert, P.; Scocchi, M.; Wilson, D.N. Fragments of the nonlytic proline-rich antimicrobial peptide Bac5 kill escherichia coli cells by inhibiting protein synthesis. Antimicrob. Agents Chemother. 2018, 62. [Google Scholar] [CrossRef] [Green Version]
- Mardirossian, M.; Pérébaskine, N.; Benincasa, M.; Gambato, S.; Hofmann, S.; Huter, P.; Müller, C.; Hilpert, K.; Innis, C.A.; Tossi, A.; et al. The dolphin proline-rich antimicrobial peptide Tur1A inhibits protein synthesis by targeting the bacterial ribosome. Cell Chem. Biol. 2018, 25, 530–539. [Google Scholar] [CrossRef] [PubMed]
- Graf, M.; Mardirossian, M.; Nguyen, F.; Seefeldt, A.C.; Guichard, G.; Scocchi, M.; Innis, C.A.; Wilson, D.N. Proline-rich antimicrobial peptides targeting protein synthesis. Nat. Prod. Rep. 2017, 34, 702–711. [Google Scholar] [CrossRef] [PubMed]
- Scocchi, M.; Tossi, A.; Gennaro, R. Proline-rich antimicrobial peptides: Converging to a non-lytic mechanism of action. Cell. Mol. Life Sci. 2011, 68, 2317–2330. [Google Scholar] [CrossRef] [PubMed]
- Podda, E.; Benincasa, M.; Pacor, S.; Micali, F.; Mattiuzzo, M.; Gennaro, R.; Scocchi, M. Dual mode of action of Bac7, a proline-rich antibacterial peptide. Biochim. Biophys. Acta (BBA) Gen. Subj. 2006, 1760, 1732–1740. [Google Scholar] [CrossRef]
- Tomasinsig, L.; Zanetti, M. The cathelicidins: Structure, function and evolution. Curr. Protein Pept. Sci. 2005, 6, 23–34. [Google Scholar] [CrossRef]
- Scocchi, M.; Skerlavaj, B.; Romeo, D.; Gennaro, R. Proteolytic cleavage by neutrophil elastase converts inactive storage proforms to antibacterial bactenecins. Eur. J. Biochem. 1992, 209, 589–595. [Google Scholar] [CrossRef]
- Zanetti, M. The role of cathelicidins in the innate host defenses of mammals. Curr. Issues Mol. Biol. 2005, 7, 179–196. [Google Scholar]
- Tossi, A.; D’este, F.; Skerlavaj, B.; Gennaro, R. Structural and functional diversity of cathelicidins. In Antimicrobial Peptides: Discovery, Design and Novel Therapeutic Strategies; CABI: Omaha, NE, USA, 2010; pp. 20–48. [Google Scholar]
- O’Leary, M.A.; Gatesy, J. Impact of increased character sampling on the phylogeny of cetartiodactyla (Mammalia): Combined analysis including fossils. Cladistics 2008, 24, 397–442. [Google Scholar] [CrossRef]
- Spaulding, M.; O’Leary, M.A.; Gatesy, J. Relationships of cetacea (Artiodactyla) among mammals: Increased taxon sampling alters interpretations of key fossils and character evolution. PLoS ONE 2009, 4, e7062. [Google Scholar] [CrossRef]
- Seefeldt, A.C.; Graf, M.; Pérébaskine, N.; Nguyen, F.; Arenz, S.; Mardirossian, M.; Scocchi, M.; Wilson, D.N.; Innis, C.A. Structure of the mammalian antimicrobial peptide Bac7(1–16) bound within the exit tunnel of a bacterial ribosome. Nucleic Acids Res. 2016, 44, 2429–2438. [Google Scholar] [CrossRef] [Green Version]
- Gagnon, M.G.; Roy, R.N.; Lomakin, I.B.; Florin, T.; Mankin, A.S.; Steitz, T.A. Structures of proline-rich peptides bound to the ribosome reveal a common mechanism of protein synthesis inhibition. Nucleic Acids Res. 2016, 44, 2439–2450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seefeldt, A.C.; Nguyen, F.; Antunes, S.; Perebaskine, N.; Graf, M.; Arenz, S.; Inampudi, K.K.; Douat, C.; Guichard, G.; Wilson, D.N.; et al. The proline-rich antimicrobial peptide Onc112 inhibits translation by blocking and destabilizing the initiation complex. Nat. Struct. Mol. Biol. 2015, 22, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Roy, R.N.; Lomakin, I.B.; Gagnon, M.G.; Steitz, T.A. The mechanism of inhibition of protein synthesis by the proline-rich peptide oncocin. Nat. Struct. Mol. Biol. 2015, 22, 466–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Florin, T.; Maracci, C.; Graf, M.; Karki, P.; Klepacki, D.; Berninghausen, O.; Beckmann, R.; Vázquez-Laslop, N.; Wilson, D.N.; Rodnina, M.V.; et al. An antimicrobial peptide that inhibits translation by trapping release factors on the ribosome. Nat. Struct. Mol. Biol. 2017, 24, 752–757. [Google Scholar] [CrossRef] [PubMed]
- Benincasa, M.; Scocchi, M.; Podda, E.; Skerlavaj, B.; Dolzani, L.; Gennaro, R. Antimicrobial activity of Bac7 fragments against drug-resistant clinical isolates. Peptides 2004, 25, 2055–2061. [Google Scholar] [CrossRef]
- Tossi, A.; Sandri, L.; Giangaspero, A. Amphipathic, alpha-helical antimicrobial peptides. Biopolymers 2000, 55, 4–30. [Google Scholar] [CrossRef]
- Lehrer, R.I.; Barton, A.; Ganz, T. Concurrent assessment of inner and outer membrane permeabilization and bacteriolysis in E. coli by multiple-wavelength spectrophotometry. J. Immunol. Methods 1988, 108, 153–158. [Google Scholar] [CrossRef]
- Knappe, D.; Kabankov, N.; Herth, N.; Hoffmann, R. Insect-derived short proline-rich and murine cathelicidin-related antimicrobial peptides act synergistically on gram-negative bacteria in vitro. Fut. Med. Chem. 2016, 8, 1035–1045. [Google Scholar] [CrossRef]
- Benincasa, M.; Pelillo, C.; Zorzet, S.; Garrovo, C.; Biffi, S.; Gennaro, R.; Scocchi, M. The proline-rich peptide Bac7(1-35) reduces mortality from salmonella typhimurium in a mouse model of infection. BMC Microbiol. 2010, 10. [Google Scholar] [CrossRef] [Green Version]
- Hansen, A.; Schäfer, I.; Knappe, D.; Seibel, P.; Hoffmann, R. Intracellular toxicity of proline-rich antimicrobial peptides shuttled into mammalian cells by the cell-penetrating peptide penetratin. Antimicrob. Agents Chemother. 2012, 56, 5194–5201. [Google Scholar] [CrossRef] [Green Version]
- Otvos, L.; Bokonyi, K.; Varga, I.; Otvos, B.I.; Hoffmann, R.; Ertl, H.C.; Wade, J.D.; McManus, A.M.; Craik, D.J.; Bulet, P. Insect peptides with improved protease-resistance protect mice against bacterial infection. Protein Sci. 2000, 9, 742–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mardirossian, M.; Sola, R.; Beckert, B.; Valencic, E.; Collis, D.W.P.; Borišek, J.; Armas, F.; Di Stasi, A.; Buchmann, J.; Syroegin, E.A.; et al. Peptide inhibitors of bacterial protein synthesis with broad spectrum and SbmA-independent bactericidal activity against clinical pathogens. J. Med. Chem. 2020. [Google Scholar] [CrossRef] [PubMed]
- Knappe, D.; Zahn, M.; Sauer, U.; Schiffer, G.; Sträter, N.; Hoffmann, R. Rational design of oncocin derivatives with superior protease stabilities and antibacterial activities based on the high-resolution structure of the oncocin-DnaK complex. ChemBioChem 2011, 12, 874–876. [Google Scholar] [CrossRef] [PubMed]
- Kragol, G.; Hoffmann, R.; Chattergoon, M.A.; Lovas, S.; Cudic, M.; Bulet, P.; Condie, B.A.; Rosengren, K.J.; Montaner, L.J.; Otvos, L. Identification of crucial residues for the antibacterial activity of the proline-rich peptide, pyrrhocoricin. Eur. J. Biochem. 2002, 269, 4226–4237. [Google Scholar] [CrossRef] [PubMed]
- Tim, C.; Simon, R.H.; Matthew, B.; Julian, P.; Jacqueline, A.M. Artemis: An integrated platform for visualization and analysis of high-throughput sequence-based experimental data. Bioinformatics 2012, 28, 464–469. [Google Scholar] [CrossRef] [Green Version]
- Kuipers, B.J.H.; Gruppen, H. Prediction of molar extinction coefficients of proteins and peptides using UV absorption of the constituent amino acids at 214 Nm to enable quantitative reverse phase high-performance liquid chromatography-mass spectrometry analysis. J. Agric. Food Chem. 2007, 55, 5445–5451. [Google Scholar] [CrossRef] [PubMed]
- Baba, T.; Ara, T.; Hasegawa, M.; Takai, Y.; Okumura, Y.; Baba, M.; Datsenko, K.A.; Tomita, M.; Wanner, B.L.; Mori, H. Construction of escherichia coli K-12 in-frame, single-gene knockout Mutants: The keio collection. Mol. Syst. Biol. 2006, 2. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Peptide | Sequence a | q b | H(GI) c |
|---|---|---|---|---|
| Orcinus orca | Orc1 | RRIPFWPPNLPGPRRPPWFLPDFRIPRIPRKR | +8 | −1.1 |
| Delphinapterus leucas | Del1 | RRIPFWPIPLRWQWPPPWFPPSFPIPRISRKR | +7 | −0.8 |
| Neophocaena asiaeorientalis | Neo1 d | RRIRFPFPPFPWQWPPAGF*ptfhipriprkq * | +3 | −0.6 |
| Tursiops truncatus | Tur1A | RRIRFRPPYLPRPGRRPRFPPPFPIPRIPRIP | +10 | −1.1 |
| Tur1B | RRIPFWPPNWPGPWLPPWSPPDFRIPRILRKR | +6 | −1 | |
| Balaenoptera acutorostrata | Bal1 e | RRIRFRPPRLPRPRPRPWIPPRFPFPRIPGKR | +12 | −1.5 |
| Lipotes vexillifer | Lip1 e | RRIRIRPPRLPRPRPRPWFPPRFPIPRIPGKR | +12 | −1.4 |
| Bos taurus | Bac7(1-35) | RRIRPRPPRLPRPRPRPLPFPRPGPRPIPRPLPFP | +11 | −1.4 |
| Microorganism and Strain | MIC * (µM) | |||||||
|---|---|---|---|---|---|---|---|---|
| Orc1 | Del1 | Bal1 | Lip1 | Tur1A | Tur1B | Neo1 | Bac71-35 | |
| E. coli ATCC 25922 | 6 | 6 | 1 | 1 | 1 | 8 | 16 | 1 |
| E. faecium ATCC 19434 | 16 | 4 | 4 | 4 | 64 | 16 | >64 | 64 |
| S. aureus ATCC 25923 | 32 | 8 | 16 | 16 | >64 | 32 | 16 | >64 |
| K. pneumoniae ATCC 700603 | 32 | >64 | 1 | 1 | 2 | >64 | >64 | 2 |
| A. baumannii ATCC 19606 | 2 | 4 | 1 | 1 | 1 | 4 | 16 | 2 |
| P. aeruginosa ATCC 27853 | 32 | 16 | 2 | 2 | 16 | 32 | 64 | 16 |
| E. agglomerans—clinical isolate | 8 | 8 | 0.5 | 0.5 | 1 | 16 | 64 | 0.5 |
| S. maltophilia ATCC 13637 | 2 | 4 | 1 | 0.5 | 1 | 8 | 24 | 1 |
| C. albicans ATCC 90029 | 4 | 8 | 1.5 | 2 | 2 | 8 | 32 | 2 |
| E. coli BW25113 | 8 | 8 | 1 | 1 | 0.75 | 16 | 24 | 0.75 |
| E. coli BW25113—Δ SbmA | 2 | 8 | 2 | 1.5 | 1.5 | 8 | 16 | 8 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sola, R.; Mardirossian, M.; Beckert, B.; Sanghez De Luna, L.; Prickett, D.; Tossi, A.; Wilson, D.N.; Scocchi, M. Characterization of Cetacean Proline-Rich Antimicrobial Peptides Displaying Activity against ESKAPE Pathogens. Int. J. Mol. Sci. 2020, 21, 7367. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21197367
Sola R, Mardirossian M, Beckert B, Sanghez De Luna L, Prickett D, Tossi A, Wilson DN, Scocchi M. Characterization of Cetacean Proline-Rich Antimicrobial Peptides Displaying Activity against ESKAPE Pathogens. International Journal of Molecular Sciences. 2020; 21(19):7367. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21197367
Chicago/Turabian StyleSola, Riccardo, Mario Mardirossian, Bertrand Beckert, Laura Sanghez De Luna, Dennis Prickett, Alessandro Tossi, Daniel N. Wilson, and Marco Scocchi. 2020. "Characterization of Cetacean Proline-Rich Antimicrobial Peptides Displaying Activity against ESKAPE Pathogens" International Journal of Molecular Sciences 21, no. 19: 7367. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21197367