Cytosolic TaGAPC2 Enhances Tolerance to Drought Stress in Transgenic Arabidopsis Plants

Life Sciences College, Northwest A&F University, Xianyang 712100, China
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2020, 21(20), 7499; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21207499
Submission received: 24 August 2020
/
Revised: 2 October 2020
/
Accepted: 4 October 2020
/
Published: 12 October 2020
(This article belongs to the Section Molecular Biology)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Drought is a major natural disaster that seriously affects agricultural production, especially for winter wheat in boreal China. As functional proteins, the functions and mechanisms of glyceraldehyde-3-phosphate dehydrogenase in cytoplasm (GAPCs) have remained little investigated in wheat subjected to adverse environmental conditions. In this study, we cloned and characterized a GAPC isoform TaGAPC2 in wheat. Over-expression of TaGApC2-6D in Arabidopsis led to enhanced root length, reduced reactive oxygen species (ROS) production, and elevated drought tolerance. In addition, the dual-luciferase assays showed that TaWRKY28/33/40/47 could positively regulate the expression of TaGApC2-6A and TaGApC2-6D. Further results of the yeast two-hybrid system and bimolecular fluorescence complementation assay (BiFC) demonstrate that TaPLDδ, an enzyme producing phosphatidic acid (PA), could interact with TaGAPC2-6D in plants. These results demonstrate that TaGAPC2 regulated by TaWRKY28/33/40/47 plays a crucial role in drought tolerance, which may influence the drought stress conditions via interaction with TaPLDδ. In conclusion, our results establish a new positive regulation mechanism of TaGAPC2 that helps wheat fine-tune its drought response.
1. Introduction
Plants are constantly subject to various environmental stresses, including drought and salinity, which all lead to severe losses in crop yield. Under harsh stress conditions, the elaborate and complex signaling crosstalk with the environment has evolved and developed to perceive and translate these stress-signaling pathways, leading to protective responses [1,2]. All these adversities could greatly reduce crop yield and increase economic costs. A deeper physiological and genetic understanding of drought resistance is crucial for increasing food production.
Glyceraldehyde-3-P dehydrogenase (GAPDH) is widely considered to be a critical enzyme involved in the glycolytic and gluconeogenesis metabolic pathways by catalyzing the NAD-dependent conversion of glyceraldehyde-3-phosphate into 1,3-diphosphoglycerate. In all living organisms, glycolysis is an important metabolic pathway in carbohydrate metabolism [3]. In higher plants, there are four distinct isoforms of GAPDHs: GAPA, GAPB, glyceraldehyde-3-phosphate dehydrogenase in cytoplasm (GAPCs), and GAPCps. Phosphorylated and NADP-dependent GAPA and GAPB are involved in photosynthetic CO2 fixation in chloroplasts, phosphorylated and NADP-dependent GAPCs exist in the cytosol, and phosphorylated and NADP-dependent GAPCps exist in the non-green plastids. Nonphosphorylated and NADP-dependent NP-GAPDH exist in the cytosol [4,5,6].
NAD-dependent GAPDH consists of four identical subunits (C4). Generally, GAPDH is used as an internal control for the relative quantitation of gene expression because it is highly conserved in plants [7,8,9,10]. The gene structure and biochemical and functional properties of GAPDH have been investigated in mammals [11]. Recently, more and more studies have been carried out on the in vivo role of plant GAPDHs, especially in stress tolerance [12,13]. Many studies have also indicated that GAPDH proteins have many functions in addition to their roles in the glycolytic pathway [14,15,16]. In Arabidopsis, the relative expression level of AtGApDH was significantly increased in Arabidopsis roots after treatment with NaCl [17,18]. In the stable genetic Arabidopsis mutant plants (gapc1-1 and gapc2-1), the stomatal apertures and photosynthetic rate were significantly increased under drought stress, compared with the wild type [19]. The accumulation of GAPC1 has been found in the nuclei of root tip cells under cadmium treatment [20]. Under a low-concentration selenium environment, NAD-dependent GAPDHs are key regulatory factors that promote the normal growth of Arabidopsis seedlings [21]. The expression level and enzyme activity of GAPDH changes significantly during the immune response [22]. In rice, the expressions of OsGApC1-3 were all induced by drought, salt, high temperature, ABA, and methyl viologen treatments. Further, the transgenic rice plants over-expressing OsGApC3 are more resistant to salt stress compared with WT [4]. In maize, GAPC3 and GAPC4 proteins are synthesized under hypoxic conditions in the roots, which belong to the classic “anaerobic polypeptide”. Hypoxia can significantly increase the transcription level of the GApC3/4 gene, while the GApC1/2 gene maintains constitutive expression or decreases the transcription level. These studies all indicate that GAPC may play a role in the stress resistance of plants in some way.
Many previous studies, including our studies, have shown that the TaGAPC1 was up-regulated under certain abiotic stresses in Chinese Spring Triticum aestivum [23]. Thus, in order to investigate the function and stress response mechanism of TaGApC2, TaGApC2-6A/6B/6D were cloned in this study. Our results show that over-expression of TaGApC2-6D reduced the accumulation of reactive oxygen species (ROS) and promoted root growth in transgenic Arabidopsis plants under drought stress, which suggests that TaGApC2-6D plays an important role in enhancing drought tolerance by adjusting ROS content and promoting root length. Further, an osmotic-stressed full-length normalized cDNA library from Chinese Spring wheat was constructed and used to screen TaGAPC2-6D’s potential partners in a yeast two-hybrid system. This provides more detailed information for further investigation into the underlying mechanisms of TaGApC2 involved in drought-related regulatory networks.
2. Results
2.1. Characterization of TaGApC2 and Bioinformatics Analysis
In our previous study, we identified six members of the GApC gene family in the wheat genome, among which TaGAPDH3 (TraesCS6A02G213700.1), TaGAPDH6 (TraesCS6B02G243700.1), and TaGAPDH8 (TraesCS6D02G196300.2) are three copies of a gene, namely, TaGAPC2-6A, TaGAPC2-6B, and TaGAPC2-6D [24]. In the present study, three wheat GApC2 genes—TaGApC2-6A, TaGApC2-6B, and TaGApC2-6D—were cloned from Chinese spring wheat and characterized. These three homologous genes share similar gene structures and are highly similar in terms of their amino acid sequences. Additionally, the numbers of exons and introns were also almost conserved within TaGApC2 (Figure S1).
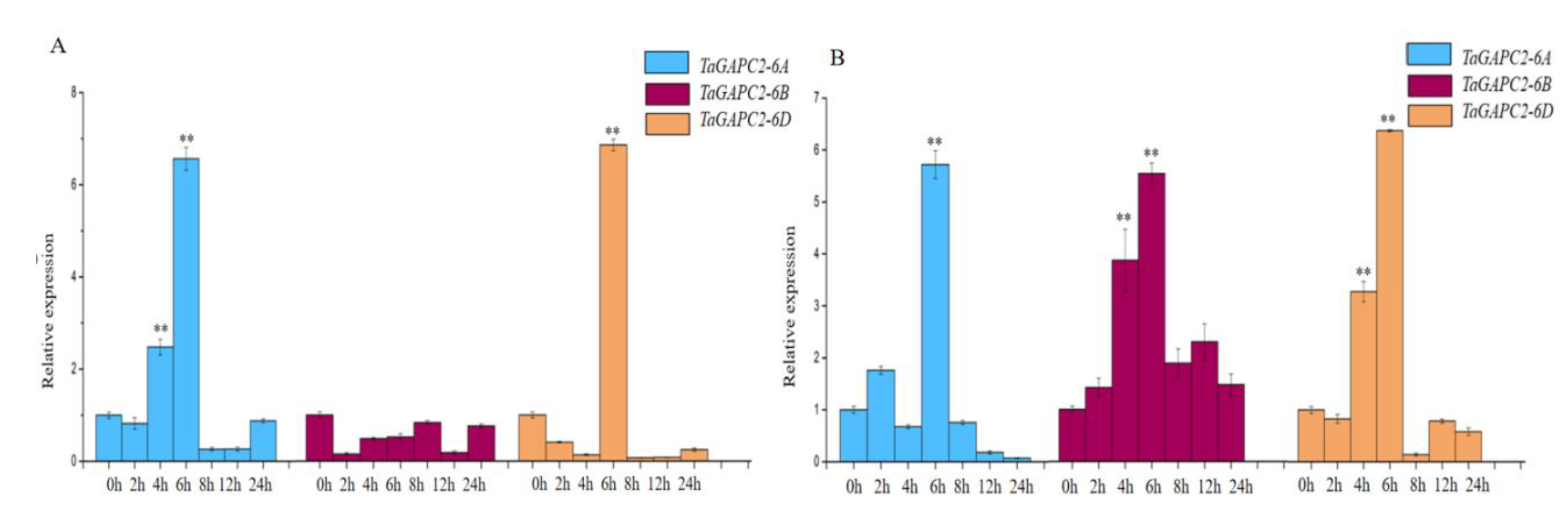
To obtain more insights into the regulation mechanism of TaGApC2 in Chinese spring wheat, the transcriptional expression patterns of TaGApC2 genes, including TaGApC2-6A, TaGApC2-6B, and TaGApC2-6D, were examined at different time points (0, 2, 4, 6, 8, 12, and 24 h) after PEG and H2O2 treatment by quantitative real-time PCR (qRT-PCR). As shown in Figure 1, real-time PCR results show that the relative expression level of TaGApC2-6A and TaGApC2-6D increased over 2–8 h and reached the highest point. In addition, these genes were highly expressed in the roots, stems, and leaves of wheat plants (Figure S2).
2.2. Interaction between TaWRKY and TaGApC2-6D and TaGApC2-6A Promoters
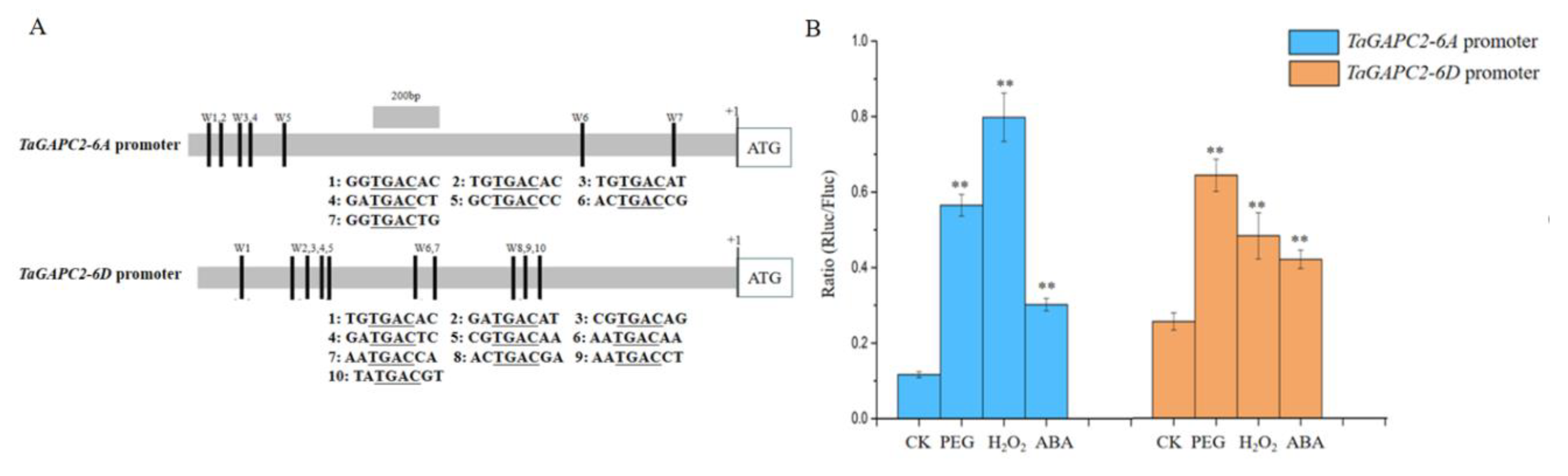
As shown in Figure 1 and Figure S2, TaGApC2-6A and TaGApC2-6D exhibited higher expression levels in different organizations and drought stress, showing that TaGApC2-6A and TaGApC2-6D might be the main effective genes for TaGApC2 in wheat. Therefore, TaGApC2-6A and TaGApC2-6D were selected for further study. We firstly amplified and sequenced approximately 2.0 kb of the promoter regions upstream of the TaGApC2-6A and TaGApC2-6D ATG start codons from Chinese spring wheat, named TaGApC2-6AP and TaGApC2-6DP. Sequence analysis showed that TaGApC2 promoter not only differs significantly in different wheat cultivars but also in different chromosomes of the same wheat cultivar (Figure S3). Furthermore, we also observed many cis-acting elements related to drought resistance in TaGApC2-6A and TaGApC2-6D promoters by bioinformatics analysis, including W-box elements; ABA-responsive elements (ABREs), MYB, and MYC binding sequences; and dehydration-responsive elements (DREs) (Table S1). TaGApC2-6AP and TaGApC2-6DP were fused to the Rluc reporter vector to detect promoter activity in tobacco leaves. The inducible activities of TaGApC2-6AP and TaGApC2-6DP under PEG, ABA, and H2O2 treatments were measured in transgenic tobacco by the Rluc/Fluc enzyme activity. The results show that TaGApC2-6AP and TaGApC2-6DP were stress-inducible promoters (Figure 2B).
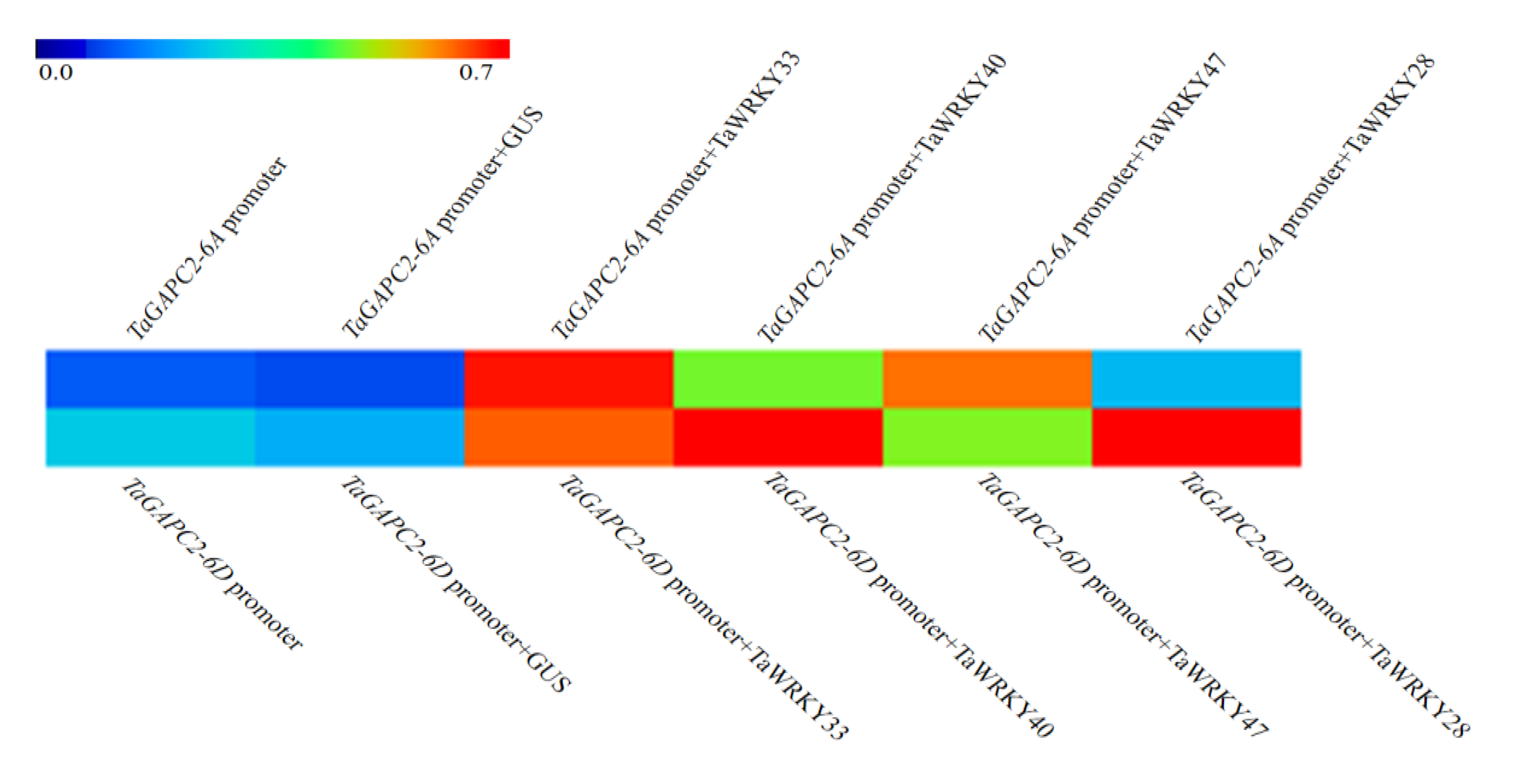
We previously observed that the transcription factor TaWRKY28/33/40/47 could bind to the special W-box in CW-TaGApC5 (TaGApC2-6D of wheat cv.Changwu) and ZY-TaGApC5 (TaGApC2-6D of wheat cv.Zhengyin) promoter. The transcriptional expression patterns of several TaWRKY genes, including TaWRKY28, 33, 40, and 47, were examined at different time points after PEG treatment by qRT-PCR [25]. TaWRKY28, 33, 40, and 47 all expressed a trend of increasing first and then decreasing under PEG stress, similar to that of TaGApC2-6A and TaGApC2-6D but before them. These results indicate that TaWRKY28, 33, 40, and 47 may also positively regulate the expression of TaGApC2-6A and TaGApC2-6D in Chinese spring wheat (Figure 1A). To further investigate whether TaWRKY28, 33, 40, and 47 could bind to TaGApC2-6A and TaGApC2-6D promoters in Chinese spring wheat, we firstly analyzed the type of W-box in TaGApC2-6A and TaGApC2-6D promoters in Chinese spring wheat. As shown in Figure 2A, these two promoters contain different types of W-box, including G/ATGACG/C/A bound by TaWRKY28, C/G/ATGACG bound by TaWRKY33, C/ATGACC bound by TaWRKY40, and C/ATGACC/G bound by TaWRKY47 [25]. To investigate the transactivation role of TaWRKY28/33/40/47 on the target TaGApC2-6A and TaGApC2-6D promoter in Chinese spring wheat, a dual-luciferase assay was conducted according to what was previously reported [26]. The promoter of TaGApC2-6A and TaGApC2-6D was cloned into the dual-luciferase report vector pC0390-RUC and transformed into Agrobacterium GV3101 and then injected together with either pC0390-TaWRKY:GUS (effector) or pC0390-GUS empty vector (control) into tobacco leaves. The Dual-Luciferase® Reporter Assay System (Promega, Madison, WI, USA) was used to assess the luciferase activity (RLuc/Fluc). The Rluc/Fluc ratio was significantly increased by simultaneous TaWRKY:GUS over-expression compared to GUS expression (Figure 3). These results indicate that TaWRKY28/33/40/47 are indeed upstream positive regulators of TaGApC2-6A and TaGApC2-6D genes in wheat cv. Chinese spring.
2.3. Morphological Changes and Drought Tolerance in Transgenic Arabidopsis Plants
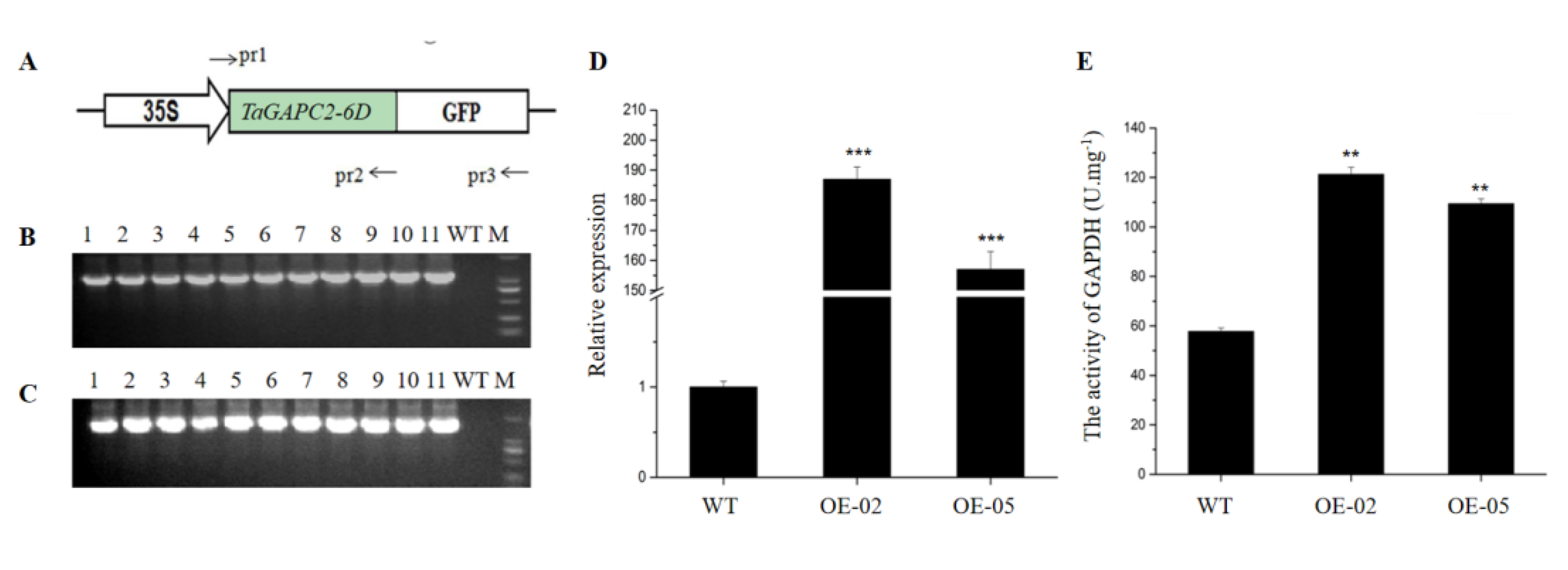
To further understand the function of TaGApC2, we constructed transgenic Arabidopsis plants harboring p35S::TaGApC2-6D, expressing TaGApC2-6D under the control of the constitutive CaMV35S promoter. We selected putative transgenic lines harboring this construct on media containing hygromycin and confirmed them using PCR with gene-specific primers and qRT-PCR. The enzyme activity of GAPDH was also measured in WT and transgenic Arabidopsis plants (Figure 4). Two independent p35S::TaGApC2-6D transgenic lines (OE-02 and 05) were selected for further analysis.
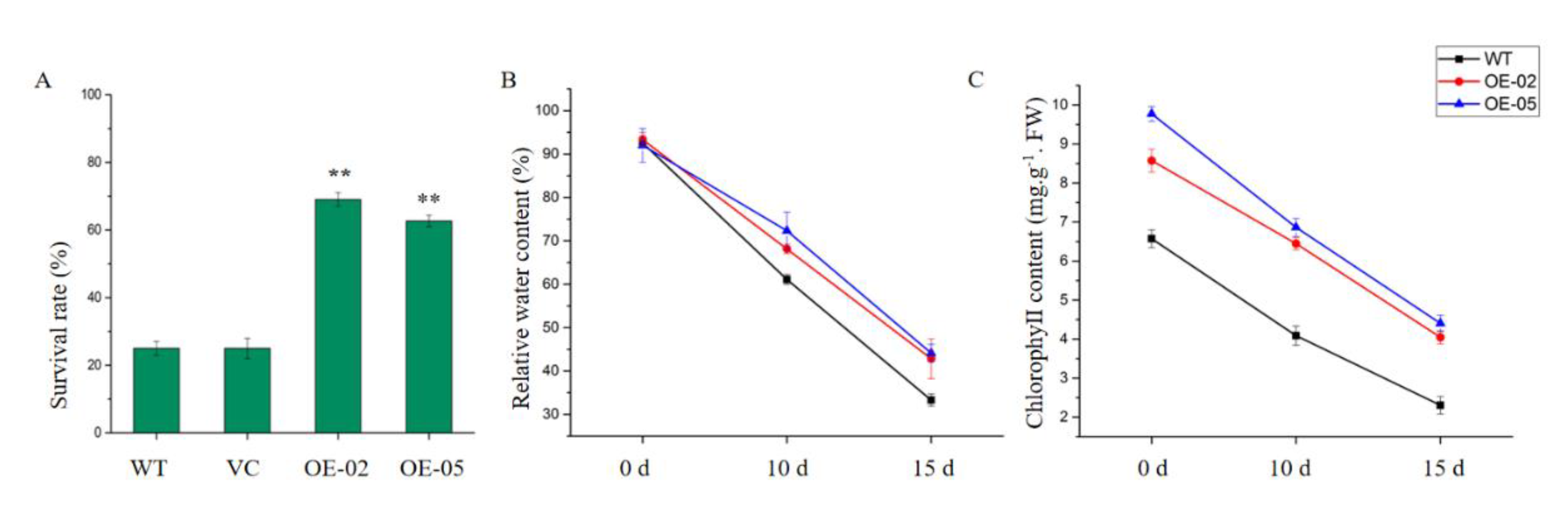
To investigate whether TaGApC2-6D over-expression was associated with drought stress tolerance, we subjected WT, vector control (VC), and transgenic three-week-old seedlings to 25 days of drought stress. At the early stage of treatment (7 days), all plants showed normal growth. At day 12 of treatment, the leaves of WT and the control line rapidly withered, whereas the growth of OE-02 and OE-05 plants was almost unaffected by drought stress. After 16 days of drought stress, both WT and the control line plants exhibited severe wilting. By contrast, the growth of OE-02 and OE-05 transgenic plants was better than that of WT. However, on the last day of drought treatment (day 25), all plants, including OE-02 and OE-05 transgenic plants, were severely wilted (Figure 5). Seven days after re-watering, 24% of the WT and 25% of the VC plants had survived, while the survival rates of the OE Arabidopsis were 68% and 62%, respectively (Figure 6A). Relative water content (RWC) and chlorophyll contents were relevant indicators for the measurement of drought tolerance. RWC and chlorophyll content were calculated in transgenic and WT Arabidopsis plants after 10 and 15 days of drought stress. As can be seen in Figure 6, the transgenic Arabidopsis lines had significantly higher RWC and chlorophyll contents than WT.
Transgenic and WT Arabidopsis seeds were grown on 1/2 MS medium for five days and then transferred to 1/2 MS medium containing 6% PEG8000. The OE-02 transgenic lines had a similar phenotype to WT plants under normal conditions. However, the total root length of the transgenic line was longer than that of WT plants under PEG8000 treatment after seven days. TaGApC2-6D more significantly promoted root growth in transgenic Arabidopsis plants than WT under PEG8000 treatment (Figure 7). The result suggests that the enhanced drought tolerance in TaGApC2-6D transgenic lines may be related to the longer root length.
2.4. Ectopic Expression of TaGApC2-6D Reduces ROS Accumulation under Drought Stress
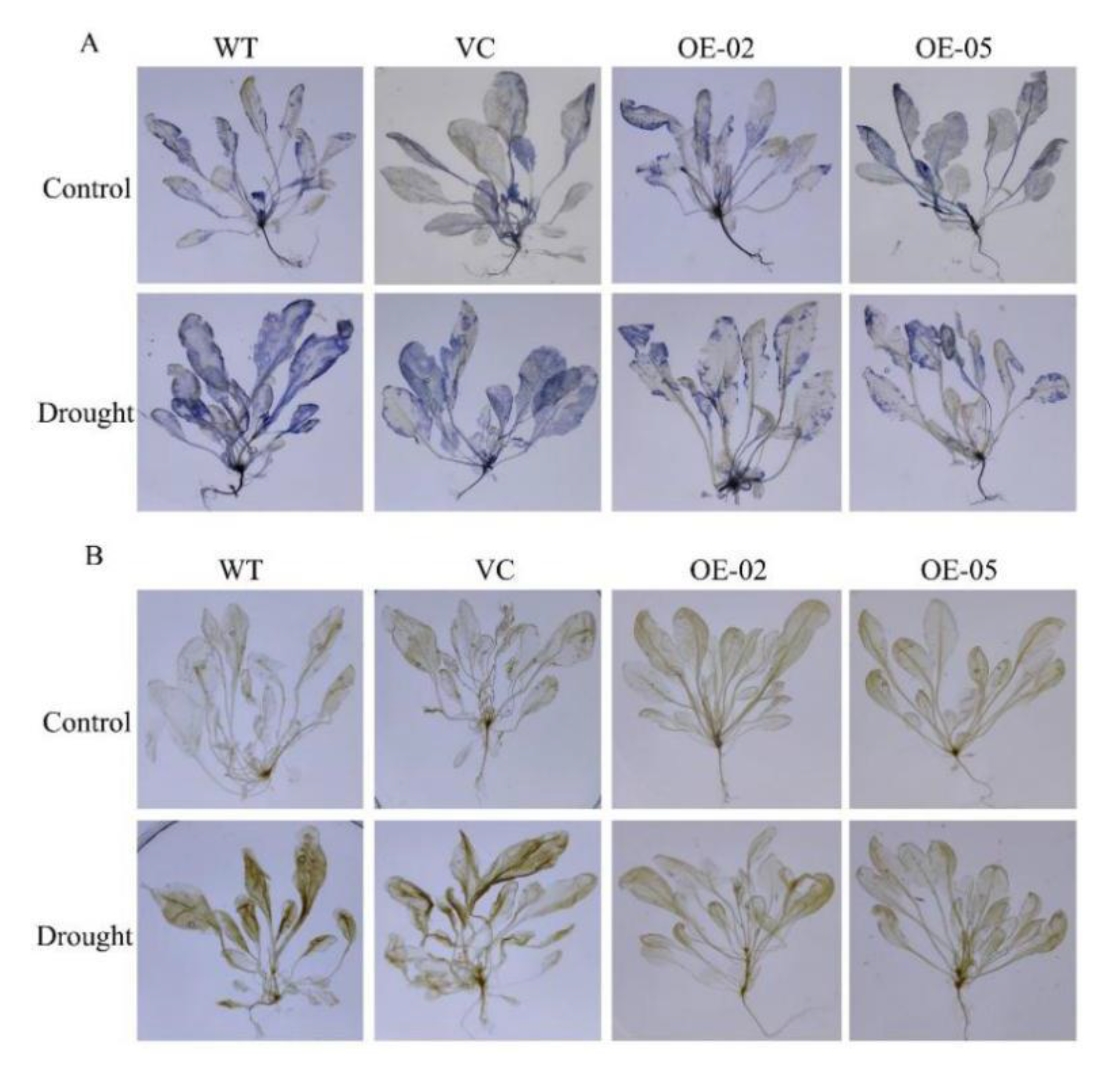
Since drought stress increases ROS production, we then investigated whether TaGApC2-6D over-expression in Arabidopsis would affect ROS accumulation under drought stress by examining O2− and H2O2 accumulation using nitroblue tetrazolium (NBT) and diaminobenzidine (DAB) staining, respectively. The results obtained from histochemical staining indicate that transgenic Arabidopsis plants and WT generated different amounts of ROS (mainly O2− and H2O2). As shown in Figure 8A, under normal conditions, little NBT staining was detected in the WT and VC, whereas after drought stress, clear NBT staining was detected in both lines. By contrast, lower NBT staining and content of O2− were detected in the OE transgenic lines, even under drought stress (Figure 8A and Figure S4). In addition, lower signals of DAB staining and content of H2O2 were detected in the OE transgenic lines than in the WT and VC lines (Figure 8B and Figure S4).
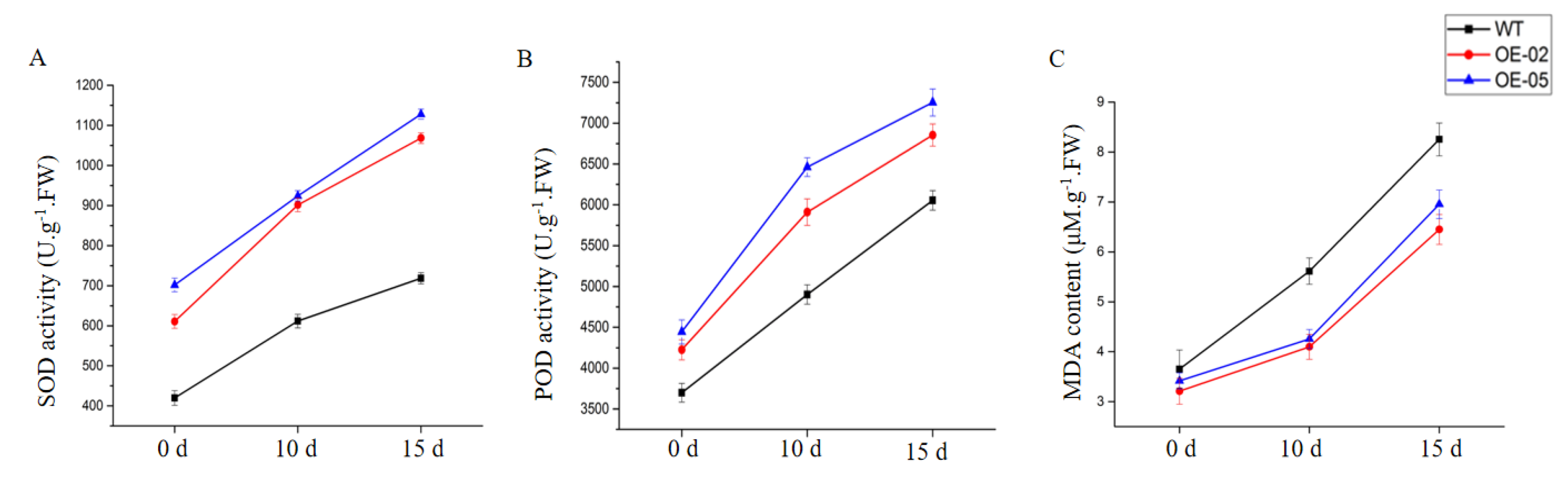
Since malondialdehyde (MDA) content is an indicator of lipid peroxidation, MDA content was measured in the leaves of WT and transgenic Arabidopsis lines. After 15 days of drought treatment, the MDA content was significantly lower in the transgenic Arabidopsis lines, relative to WT (Figure 9C). Enzymatic antioxidants play a very significant role in scavenging harmful ROS that accumulates under stress. We further analyzed the superoxide dismutase (SOD) and peroxidase (POD) of these significant antioxidant enzymes in the leaves of the plants. These activities exhibited a similar trend in all lines after 10 and 15 days of drought stress (Figure 9A,B). After 10 and 15 days of drought stress, the activities of all antioxidant enzymes were higher in the TaGApC2-6D over-expression Arabidopsis lines than in WT. These results suggest that the enhanced drought tolerance in TaGApC2-6D transgenic lines is related to the reduced MDA content and the increased antioxidant enzyme activity.
2.5. Screening of Proteins Interacting with TaGApC2-6D by Yeast Two-Hybrid
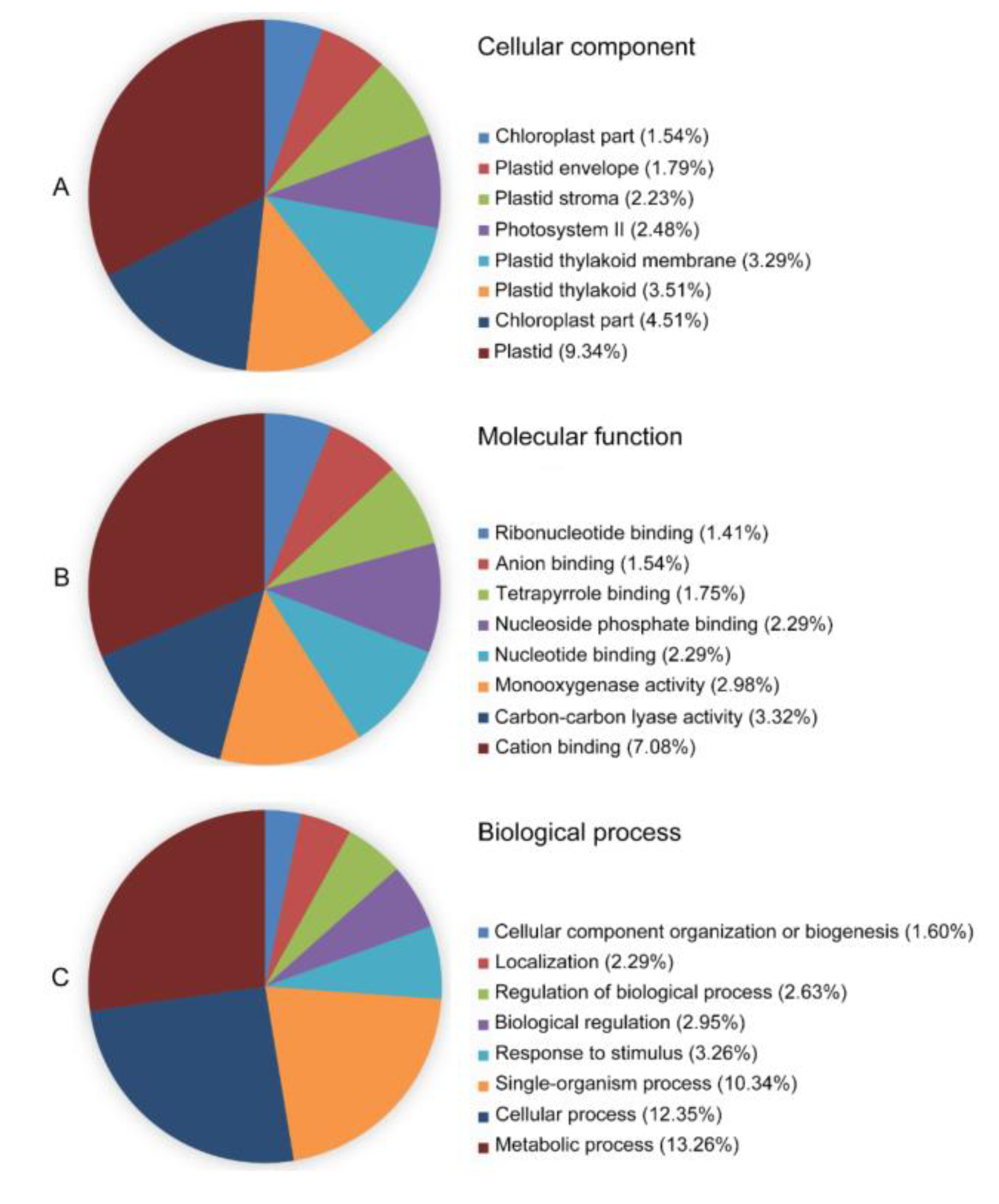
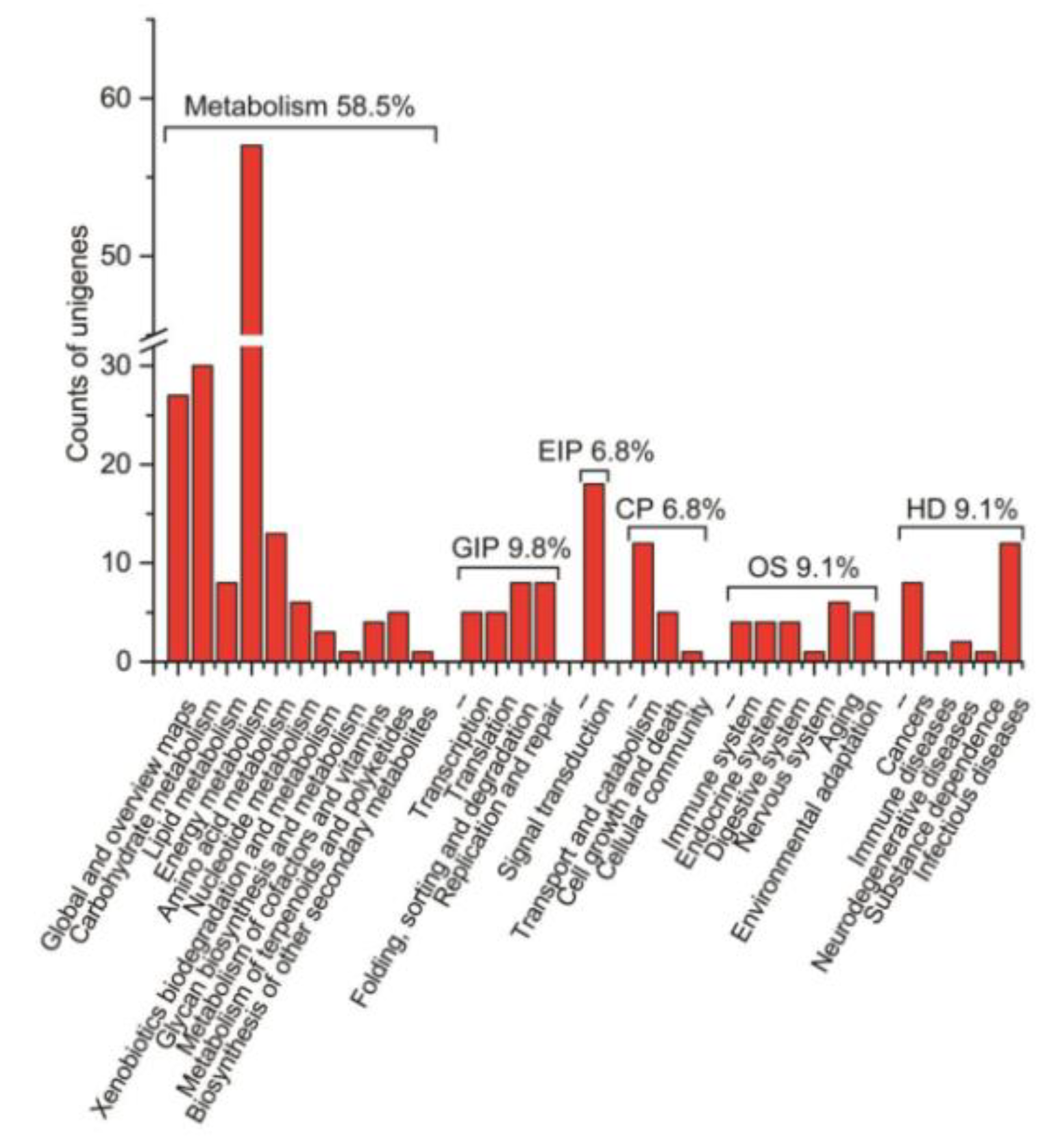
We firstly conducted an osmotic-stressed full-length normalized wheat cDNA library. The resultant library had a quantity of 5.30 × 105 cfu/mL independent clones, and its titer was 3.62 × 107 cfu/mL (Figure S5). To assess the quality of the library, a total of 1000 clones selected randomly from the cDNA library were sequenced, from which 804 high-quality expressed sequence tags (ESTs) from 963 raw sequences with an average length of 1048 bp were obtained. The 804 ESTs were clustered into 591 unigenes containing 64 contigs and 527 singletons. Based on GO annotation, the 579 of 591 unigenes were found in their homologous sequence and 330 different GO terms of cellular component (98), molecular function (71), and biological process (161) were obtained (Figure 10). Meanwhile, some transcription factors were hit, including transcription factor bHLH35, MADS-box transcription factor 5, transcription factor GLK2, and transcription factor MYB1R1-like, which were reported to play a key role in response to drought stress. The unigenes were subjected to analysis of biochemical pathways as described in the Kyoto Encyclopedia of Genes and Genomes (KEGG) database. The results show that 265 unigenes had their counterparts among 591 unigenes in the KEGG database, which relate to the six biological functions: metabolism, genetic information processing (GIP), environmental information processing (EIP), cellular processes (CP), organismal systems (OS), and human diseases. The metabolic pathways are the most abundant, making up 58.5% of the total (Figure 11). KEGG pathway analysis displayed that the cDNA library contained several plant stress response pathways, including the Ras signal pathway, MAPK signal pathway, TGF-beta signal pathway, HIF-1 signal pathway, and FoxO signaling pathway.
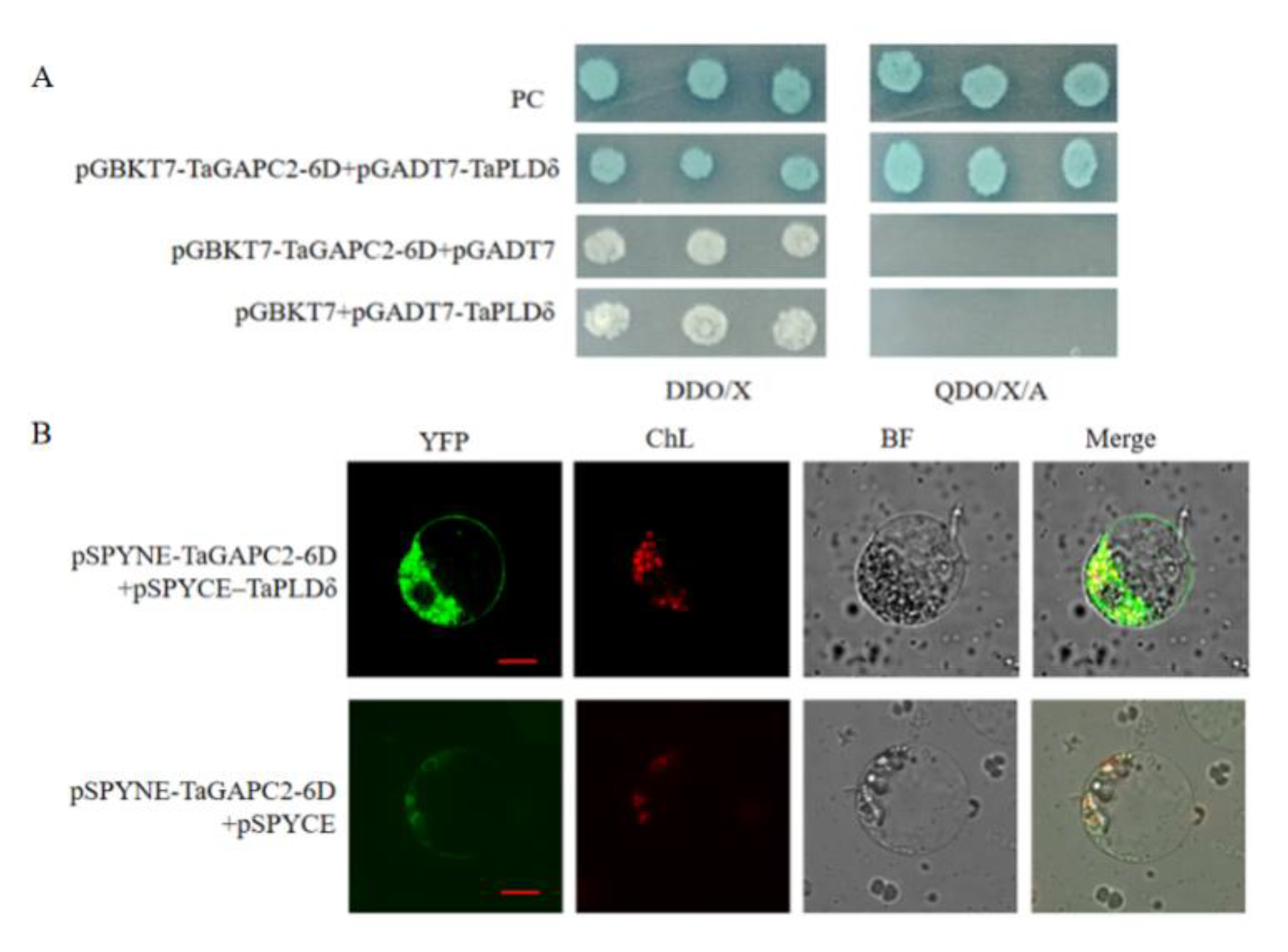
To identify proteins that interact with TaGAPC2-6D, we used full-length TaGAPC2-6D as bait in the yeast two-hybrid (Y2H) system and screened a wheat osmotic-stressed cDNA library constructed from the prey vector pGADT7-Rec. To determine the specificity of interactions between TaGAPC2-6D and interacting proteins, the bait and positive library plasmid were co-transformed into Y2HGold strain pairwise. After exclusion of the false positives, 10 clones were identified as potential interaction partners of TaGAPC2-6D (Table S2). Among the putative binding partners of the TaGAPC2-6D protein, one was TaPLDδ, which could also interact with GAPCs in Arabidopsis. The bait and prey were again co-transformed into Y2HGold strain to confirm the interaction (Figure 12A).
To further verify the interaction between TaGAPC2-6D and TaPLDδ in plant cells, the bimolecular fluorescence complementation assay (BiFC) assay was performed in tobacco leaves. YFP fluorescence was reconstituted when pSPYCE–TaPLDδ and pSPYNE–TaGAPC2-6D were co-expressed. Recombinant vectors with corresponding non-fused pSPYCE empty vectors generated no fluorescence (Figure 12B).
3. Discussion
3.1. TaWRKY Positively Regulate TaGApC2 Gene Expression
In plants, there are many drought-responsive transcription factors that regulate the transcriptional expression of downstream genes by binding specific cis-acting elements of the promoter. WRKY transcription factors are one of the largest families of transcriptional regulators in plants, and are involved in the drought-related response [27]. In plants such as wheat [28], rice [29], brachypodium distachyon [30], maize [31], soybean [32], and cotton [33], a large number of WRKYs have been identified in recent years. In our previous study, we identified that TaWRKY28, TaWRKY33, TaWRKY40, and TaWRKY47 preferentially bind to specific W-box in cw-TaGApC5 (cw-TaGApC2-6D) and zy-TaGApC5 (zy-TaGApC2-6D) by Y1H and EMSA (Electrophoretic Mobility Shift Assay) analyses [25]. In Chinese spring wheat, the real-time PCR results indicate that the expression patterns of TaGApC2-6A and TaGApC2-6D were similar to TaWRKY28/33/40/47 but lagged behind TaWRKY28/33/40/47 under drought stress (Figure 1A). Thus, we speculated that TaWRKY28/33/40/47 may also be potential positive regulators of TaGApC2-6A and TaGApC2-6D expression in Chinese spring wheat. A tobacco co-transformation experiment demonstrated that TaWRKY28/33/40/47 indeed improves transcription levels of TaGApC2-6A and TaGApC2-6D (Figure 3). These results indicate that TaWRKY28/33/40/47 positively regulates TaGApC2-6A and TaGApC2-6D gene expression.
3.2. TaGApC2 Plays a Key Role in Drought Stress
Arabidopsis thaliana is often used as a model plant to study gene functions due to its short growth cycle, small nuclear genome, and high reproduction coefficient. Niu et al. constructed over-expression TaDREB3 transgenic Arabidopsis plants to explore the function of the TaDREB3 gene under drought, salt, and heat stresses [34]. Over-expression of TaWRKY46 enhanced osmotic stress tolerance in transgenic Arabidopsis thaliana plants, which was mainly demonstrated by transgenic Arabidopsis plants forming a higher germination rate and longer root length [35]. In this study, a transgenic Arabidopsis over-expressing TaGApC2-6D was constructed through hygromycin screening and PCR identification, which provided experimental materials for studying TaGApC2-6D function. RWC, MDA, and chlorophyll content have been widely used to reflect drought tolerance in plants. The leaves of TaGApC2-6D overexpressing Arabidopsis plants exhibited higher survival rate, RWC, and chlorophyll content after drought treatment than WT plants (Figure 6). Further phenotypic analysis showed that TaGApC2-6D over-expressing Arabidopsis plants possessed longer roots at 6% concentrations of PEG (Figure 7).
In plants, ROS are the key cellular signals in response to multiple stresses, such as drought, wounding, and low and high temperatures. H2O2 and O2− are the major and most stable type of ROS. Abiotic stress, such as drought stress, could cause excessive accumulation of ROS raising lipid peroxidation to interfere with normal physiological processes and ultimately leading to programmed cell death [36,37,38]. In order to explore the effect of TaGApC2-6D on antioxidant enzyme activity, the antioxidant enzyme activities (POD, SOD) were measured in wild-type and transgenic Arabidopsis under drought stress. The results show that the H2O2 and O2− accumulation in transgenic Arabidopsis over-expressing TaGApC2-6D were less and the enzyme activities of POD and SOD were enhanced, compared with WT Arabidopsis (Figure 8 and Figure 9). The increased ROS clearance ability reduced cell membrane damage and enhanced drought resistance. All the results suggest that over-expression of TaGApC2-6D enhanced drought tolerance in Arabidopsis plants. A similar finding was also verified in other TaGApDH [39].
3.3. Interaction between TapLDδ and TaGApC2 May Contribute to Plant Drought Tolerance
To date, little is known about genes in wheat contributing to the strong adaptation in drought areas. The molecular mechanism for wheat’s adaptation to drought environments needs to be explained. It has been shown that full-length cDNA clones can be used to study the function of gene products [40]. In order to explore the biology of a plant response mechanism—genes such as those responsible for survival, dehydration avoidance, and development—a full-length cDNA library was constructed using wheat leaves from a drought condition. The high-quality-expression cDNA library can offer molecular resources for the analysis of genes involved in biology and for studying its protein function [41]. Our library storage was 5.30 × 105 cfu/mL, and the titer was 3.62 × 107 cfu/mL. The insert fragment size was greater than 0.75 kb, which proved that the cDNA library is of high quality and has a high amount of information. The normalized cDNA library can also increase the possibility of selecting the mRNA with low expression [42].
In this study, we have screened 10 proteins that potentially interact with TaGAPC2-6D from the osmotic-stressed cDNA library. A large number of studies have shown that GAPC could interact with some stress-related proteins to protect plants from damage under abiotic stress. In Arabidopsis plants, GAPC could interact with E3 ubiquitin ligase (SINAL7) to make it monoubiquitinated. Arabidopsis over-expressing SINAL7 showed increased concentrations of hexose, sucrose, and plant biomass, accompanied by elevated drought resistance and delayed leaf senescence, which provided important clues revealing that GAPC is involved in the resistance mechanism [43]. The receptor protein kinase FERONIA (FERONIA, FER) interacts with GAPC to catalyze a key reaction in glycolysis and promote energy production [44]. In this work, a wheat PLDδ protein was selected as a target protein to verify potential interactions with TaGAPC2 in yeast, and this interaction was further confirmed through bimolecular fluorescence complementation (Figure 12B). In the plant, PLD has been identified to respond to multiple stresses [45]. For example, pLDα3 positively responds to hyperosmotic stress [46], pLDα1 could respond to water stress by promoting ABA-regulated stomatal closure to reduce water loss, and pLDα1 is also involved in the process by which lipid messenger PA produces peroxide products [47,48]. We propose that TaGAPC2 may improve the drought tolerance of plants by interacting with PLDδ to promote ABA-regulated stomata closure. Our findings offer evidence of TaGAPC2’s roles in drought resistance on a molecular level. Further studies on whether TaGAPC2 interacts with other proteins will be required to thoroughly understand TaGAPC2’s molecular function.
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
Common wheat (Triticum aestivum L. cv. Chinese Spring) was used in this study. The seeds were surface-sterilized with 75% ethanol for 2 min and rinsed three times with sterile water. Seeds were placed on moistened filter paper in petri dishes for germination in darkness at 22 °C for 24 h. The sprout seedlings were grown in a chamber at 22 °C in a 16/8 h (light/dark) cycle with a growth medium using Murashige and Skoog (MS) liquid medium.
To construct an osmotic-stressed cDNA library, 10-day-old seedlings were immersed in 20% PEG8000 (polyethylene glycol) solution. The treated leaf samples were collected at 0, 1, 2, 6, 12, and 24 h after the treatment, immediately frozen in liquid nitrogen, and kept at −80 °C for RNA isolation.
Arabidopsis thaliana seeds were surface-sterilized with 70% (v/v) ethanol for 2 min followed by 4% (v/v) hypochlorite, cold-treated for three days at 4 °C for vernalization, and germinated on 1/2 MS medium agar plates. Plants were grown with a 16/8 h (light/dark) cycle photoperiod at 22 °C. Seedlings were transplanted to soil 10 days after germination.
4.2. Total RNA Extraction and Real-Time pCR Analysis
Total RNA was extracted using an RNAisoTM Plus kit (TaKaRa, Dalian, China) and treated with RNase-free DNase I (TaKaRa, Dalian, China). The quality and purity of RNA were evaluated by Bioanalyzer 2100 and 1% agarose gel electrophoresis. First-strand cDNA was synthesized using a PrimeScript First-Strand cDNA Synthesis Kit (TaKaRa, Dalian, China). qRT-PCR was conducted using a Bio-Rad CFX96 system (BioRad, Hercules, CA, USA). The β-actin (AB181991.1) was used as the internal transcript level control for qRT-PCR using SYBR® Premix Ex Taq™ (TaKaRa, Dalian, China). Detailed information on the primer sequences is provided in the supporting information (Table S3). All real-time PCR reactions were performed in triplicate to ensure reproducibility of the results.
4.3. Examination of Plant Stress Tolerance
The coding sequence of TaGApC2-6D was cloned into pCAMBIA1302 under the control of the CaMV 35S promoter, resulting in 35S::TaGAPC2-6D construct. This construct was used in transformation mediated by Agrobacterium to obtain transgenic Arabidopsis lines. Hygromycin-resistant Arabidopsis transformants carrying TaGApC2-6D were generated using the inflorescence infiltration method [49]. Transformed plants were cultured on 1/2 MS medium containing 30 µg/mL hygromycin in a day/night regime of 16/8 h under white light at 22 °C for 10 days and then transferred to soil.
The Enzyme-Linked Immunosorbent Assay (ELISA) Kits (Meimian Biotech, Lianyungang, China) was applied to detect the enzyme activities of GAPDH from the aboveground part of three-week-old Arabidopsis plants, which were homogenized with cold PBS. Then, the supernatants were collected after centrifugation and added to test wells for further reactions following the operating manual. The absorbance at 450 nm was measured using a microplate reader. All results were determined by the standard curve equation and normalized to the protein concentration of corresponding samples.
Homozygous T3 seeds of transgenic Arabidopsis lines were used for phenotypic analysis. Arabidopsis seeds were grown on 1/2 MS agar plates that were routinely kept for three days in darkness at 4 °C to break dormancy and then transferred to a tissue culture room at 22 °C for five days. For drought treatment, 5-day-old seedlings were then transferred to 1/2 MS agar plates containing 6% PEG8000 for seven days. The total root lengths of the Arabidopsis plants were measured [50]. To analyze the drought tolerance in transgenic plants, the seedlings cultivated with soil in pots for three weeks at uniform developmental stages were subjected to drought stress treatment by withholding water. After being treated for 25 days, the plants were rewatered for seven days and then the survival rates of the different plant lines were recorded. Three biological experiments were performed, and at least 50 plants were used for the calculation.
H2O2 content detection kit and O2− content detection kit (Solarbio, Beijing, China) were used to detect H2O2 content and O2− content. O2− and H2O2 accumulation was measured as previously described [26]. Determination of the RWC and the chlorophyll content of Arabidopsis leaves after drought stress were performed as previously described [26]. The MDA content and SOD and POD activity were determined as described [26].
4.4. Construction of the cDNA Library
The cDNA library was constructed using the Make Your Own “Mate&Plate™” Library System User Manual (Clontech, Mountain View, CA, USA) according to the manufacturer’s instruction. The first single-stranded cDNA (ss cDNA) were synthesized by 1 μg of total RNA using CDS III/3′ PCR primer and Moloney Murine Leukemia Virus (MMLV) Reverse Transcriptase. The double-stranded cDNA (dscDNA) was acquired by 2 μL of sscDNA product using Advantage 2 Polymerase Mix (Clontech) via long-distance PCR (LD-PCR). Then, the dscDNA products were digested with proteinase K, treated with DSN solution, and purified with a CHROMA SPINTM TE-400 column (Clontech). The purified dscDNA was ligated with pGADT7-Rec vector (SmaI-lineared) via homologous recombination using Clone ExpressTM II One Step Cloning Kit (Clontech). The products were transformed into Escherichia coli DH5α strains, which were stored in 50% glycerol at −80 °C.
The transformed strains were diluted using LB medium (Amp) with the ratio 1:105 and then spread on 15 cm agar LB plates (Amp/IPTG/X-gal) and incubated at 37 °C for 12 h. The library titer and clone capacity were calculated by counting the clones from the plates. The library titer (cfu/mL) = colonies on LB/IPTG/X-gal × dilution factor ÷ volume (mL) plated. Colonies were randomly selected to identify the size of the inserted sequences and amplified with T7 SP and 3′AD primers. The library plasmid was isolated by High Pure Maxi Plasmid Kit (TianGen Biotech, Beijing, China).
4.5. Sequencing and Analysis of Expressed Sequence Tags (ESTs)
The transformed bacteria were randomly selected and inserted, and cDNA was sequenced from the 5′ end. The Codon Code Aligner program was used to remove vector sequences and assemble EST sequences into contigs. ESTs greater than 200 bp in length were regarded as high-quality ESTs after removal of empty ESTs and vector ESTs. The PHRED program was used for base calling and quality assessment [51]. The EST sequences were clustered and assembled into successive consensus sequences (contigs) using the Codon Code Aligner program, which had high stringency parameters of minimum percent identity of 99%, minimum overlap length of 50, and default parameters for the rest. A cluster containing only one sequence was classified as a singleton. All similarity searches were batch-executed locally using BlastN, BlastX, and TBlastX tools [52,53]. The Blast2go software package was used for GO annotations. By comparing the results with the Kyoto Encyclopedia of Genes and Genomes (KEGG, ftp://ftp.genome.jp/pub/kegg/) database, the functions of most unigenes were obtained. Finally, the sequences were classified as functional categories using the GO database [54].
4.6. Yeast Two-Hybrid Screen
To identify proteins that interact with TaGAPC2-6D, we screened a wheat drought-treated full-length normalized cDNA library in the yeast two-hybrid system. A fragment of TaGApC2-6D with open reading frame (ORF) was amplified from Triticum aestivum L. cv. Chinese Spring cDNAs using primers and subsequently cloned into the pGBKT7 vector (Clontech) as baits. The bait construct was transformed into the yeast strain Y2HGold and tested for auto-activation of MEL1 and AUR1-C reporter genes. The library plasmid was used as the prey. The prey vectors were transformed into the yeast strains Y2HGold containing bait vector and without auto-activation according to the manufacturer protocol. The transform products were plated onto synthetic dropout (SD) medium without leucine (Leu), tryptophan (Trp), and histidine (His) (TDO). Then, all the colonies that grew on TDO were screened and patched out onto higher stringency SD medium without Leu, Trp, His, and adenine (Ade) (QDO), QDO containing Aureobasidin A (AbA) (QDO/A), and QDO containing AbA and X-α-gal (X) (QDO/A/X), in turn. All plasmids of the colonies that grew on QDO/A/X were isolated using the yeast plasmid mini preparation kit (Beyotime, Shanghai, China), and the plasmids inserts were PCR-amplified using the universal primer on the pGBKT7 vector. To confirm positive interactions, the baits and positive library plasmids were co-transformed into Y2HGold stain according to the manufacturer protocol.
4.7. Bimolecular Fluorescence Complementation (BiFC) Assay
BiFC assay was performed in Nicotiana benthamiana leaves. The ORFs of TapLDδ and TaGApC2-6D without termination codons were recombined with C-terminal part of YFP in the pSPYCE-35S/pUC-SPYCE vector and N-terminal part of YFP in the pSPYNE-35S/pUC-SPYNE vector, respectively. The pSPYCE-TaPLDδ and pSPYNE-TaGAPC2-6D were co-transformed into Nicotiana benthamiana leaves with corresponding empty vectors as negative controls. The Nicotiana benthamiana leaves’ protoplasts were observed for fluorescence at 48 h after transformation using a confocal laser scanning microscope (Andor, Belfast, UK).
5. Conclusions
Taken together, the results presented in this study provide evidence that the transcription factor TaWRKY positively regulates the expression level of the TaGApC2 gene by specially binding to its promoter in Chinese spring wheat. Over-expression of TaGApC2 in Arabidopsis led to lower ROS production, longer root length, and higher drought tolerance under drought stress. TaGAPC2 can interact with TaPLDδ, and it plays a vital role in plant drought tolerance. Our research provides a better understanding of the molecular mechanism of TaGAPC2 in response to drought stress and offers a promising strategy to increase crop stress tolerance using bioengineering technology.
Supplementary Materials
Supplementary materials can be found at https://0-www-mdpi-com.brum.beds.ac.uk/1422-0067/21/20/7499/s1.
Author Contributions
L.Z. designed the experiments, performed the experiments, and analyzed the corresponding results. H.Z. performed the experiments. S.Y. supervised this whole process and reviewed this paper. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by The National Nature Science Foundation of China (No. 31671609; NO. 31271625).
Acknowledgments
We would like to thank the National Natural Science Foundation of China for their financial support.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase |
| GAPC | Glyceraldehyde-3-phosphate dehydrogenase in cytoplasm |
| qRT-PCR | Quantitative real-time PCR |
| Y2H | Yeast two-hybrid |
| BiFC | Bimolecular fluorescence complementation assay |
| SDO | SD/-Trp |
| SDO/X | SD/-Trp/X-α-Gal |
| SDO/X/A | SD/-Trp/X-α-Gal/AbA |
| DDO/X/A | SD/-Leu/-Trp/X-α-Gal/AbA |
| QDO/X/A | SD/-Ade/-His/-Leu/-Trp/X-α-Gal/AbA |
| ROS | Reactive oxygen species |
References
- Singh, T.R.S.; Sushma, T.; Vinod; Naik, B.K.; Suresh, C.; Rupesh, D.; Niharika, M.; Sanjay, S.; Kumar, S.N.; Tomar, S.M.S. Molecular and Morpho-Agronomical Characterization of Root Architecture at Seedling and Reproductive Stages for Drought Tolerance in Wheat. PLoS ONE 2016, 11, e0156528. [Google Scholar]
- Ge, P.; Ma, C.; Wang, S.; Gao, L.; Li, X.; Guo, G.; Ma, W.; Yan, Y. Comparative proteomic analysis of grain development in two spring wheat varieties under drought stress. Anal. Bioanal. Chem. 2012, 402, 1297–1313. [Google Scholar] [CrossRef] [PubMed]
- Danshina, P.V.; Schmalhausen, E.V.; Avetisyan, A.V.; Muronetz, V.I.; Danshina, P.V. Mildly Oxidized Glyceraldehyde-3-Phosphate Dehydrogenase as a Possible Regulator of Glycolysis. IUBMB Life 2001, 51, 309–314. [Google Scholar] [PubMed]
- Zhang, X.-H.; Rao, X.-L.; Shi, H.-T.; Li, R.-J.; Lu, Y.-T. Overexpression of a cytosolic glyceraldehyde-3-phosphate dehydrogenase gene OsGAPC3 confers salt tolerance in rice. Plant Cell Tissue Organ Cult. 2011, 107, 1–11. [Google Scholar] [CrossRef]
- Muoz-Bertomeu, J.; Bermúdez, M.A.; Segura, J.; Ros, R. Arabidopsis plants deficient in plastidial glyceraldehyde-3-phosphate dehydrogenase show alterations in abscisic acid (ABA) signal transduction: Interaction between ABA and primary metabolism. J. Exp. Bot. 2011, 62, 1229–1239. [Google Scholar] [CrossRef] [Green Version]
- Petersen, J.; Brinkmann, H.; Cerff, R. Origin, Evolution, and Metabolic Role of a Novel Glycolytic GAPDH Enzyme Recruited by Land Plant Plastids. J. Mol. Evol. 2003, 57, 16. [Google Scholar] [CrossRef]
- Morgante, C.V.; Guimarães, P.M.; Martins, A.C.; Araújo, A.C.; Brasileiro, A.C. Reference genes for quantitative reverse transcription-polymerase chain reaction expression studies in wild and cultivated peanut. BMC Res. Notes 2011, 4, 339. [Google Scholar] [CrossRef] [Green Version]
- Zhong, H.Y.; Chen, J.W.; Li, C.Q.; Chen, L.; Wu, J.Y.; Chen, J.Y.; Lu, W.J.; Li, J.G. Selection of reliable reference genes for expression studies by reverse transcription quantitative real-time PCR in litchi under different experimental conditions. Plant Cell Rep. 2011, 30, 641. [Google Scholar] [CrossRef]
- Wu, Y.; Wu, M.; He, G.; Zhang, X.; Zhang, C. Glyceraldehyde-3-phosphate dehydrogenase: A universal internal control for Western blots in prokaryotic and eukaryotic cells. Anal. Biochem. 2012, 423, 15–22. [Google Scholar] [CrossRef]
- Zhu, X.; Yuan, M.; Shakeel, M.; Zhang, Y.; Wang, S.; Wang, X.; Zhan, S.; Kang, T.; Li, J. Selection and Evaluation of Reference Genes for Expression Analysis Using qRT-PCR in the Beet Armyworm Spodoptera exigua (Hübner) (Lepidoptera: Noctuidae). PLoS ONE 2014, 9, e84730. [Google Scholar] [CrossRef]
- Colell, A.; Ricci, J.E.; Tait, S.; Milasta, S.; Maurer, U.; Bouchier-Hayes, L.; Fitzgerald, P.; Guio-Carrion, A.; Waterhouse, N.J.; Li, C.W. GAPDH and autophagy preserve survival after apoptotic cytochrome c release in the absence of caspase activation. Cell 2007, 129, 983–997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Astier, J.; Wawer, I.; Bessonbard, A.; Olivier, L.; Jeandroz, S.; Terenzi, H.; Dobrowoslka, G.; Wendehenne, D. GAPDH, NtOSAK and CDC48, a conserved chaperone-like AAA-ATPase, as nitric oxide targets in response to (a)biotic stresses. Nitric Oxide 2012, 27, S9. [Google Scholar] [CrossRef]
- Sánchez-Bel, P.; Egea, I.; Sánchez-Ballesta, M.T.; Martinez-Madrid, C.; Fernandez-Garcia, N.; Romojaro, F.; Olmos, E.; Estrella, E.; Bolarín, M.C.; Flores, F.B. Understanding the mechanisms of chilling injury in bell pepper fruits using the proteomic approach. J. Proteom. 2012, 75, 5463–5478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liiv, I.; Haljasorg, U.; Kai, K.; Maslovskaja, J.; Laan, M.; Peterson, P.R. AIRE-induced apoptosis is associated with nuclear translocation of stress sensor protein GAPDH. Biochem. Biophys. Res. Commun. 2012, 423, 32. [Google Scholar] [CrossRef] [PubMed]
- Elaine, F.; Rosa, G.; Laura, A.; Karla, G.; Juan, A.; Josefa, B.; Laura, B. Protein interaction studies point to new functions for Escherichia coli glyceraldehyde-3-phosphate dehydrogenase. Res. Microbiol. 2013, 164, 145–154. [Google Scholar]
- Maloney, C.A.; Hay, S.M.; Reid, M.D.; Duncan, G.; Nicol, F.; Sinclair, K.D.; Rees, W.D. A methyl-deficient diet fed to rats during the pre—And peri-conception periods of development modifies the hepatic proteome in the adult offspring. Genes Nutr. 2013, 8, 181–190. [Google Scholar] [CrossRef] [Green Version]
- Udo, R.; Edda, R.L.; Stephan, C. Proteome changes in Arabidopsis thaliana roots upon exposure to Cd2+. J. Exp. Bot. 2006, 57, 4003–4013. [Google Scholar]
- Jiang, Y.; Yang, B.; Harris, N.S.; Deyholos, M.K. Comparative proteomic analysis of NaCl stress-responsive proteins in Arabidopsis roots. J. Exp. Bot. 2007, 58, 3591–3607. [Google Scholar] [CrossRef] [Green Version]
- Guo, L.; Devaiah, S.P.; Narasimhan, R.; Pan, X.; Zhang, Y.; Zhang, W.; Wang, X. Cytosolic glyceraldehyde-3-phosphate dehydrogenases interact with phospholipase Dδ to transduce hydrogen peroxide signals in the Arabidopsis response to stress. Plant Cell 2012, 24, 2200–2212. [Google Scholar] [CrossRef] [Green Version]
- Vescovi, M.; Zaffagnini, M.; Festa, M.; Trost, P.; Lo Schiavo, F.; Costa, A. Nuclear accumulation of cytosolic glyceraldehyde-3-phosphate dehydrogenase in cadmium-stressed Arabidopsis roots. Plant Physiol. 2013, 162, 333–346. [Google Scholar] [CrossRef] [Green Version]
- Takeda, T.; Fukui, Y. Possible role of NAD-dependent glyceraldehyde-3-phosphate dehydrogenase in growth promotion of Arabidopsis seedlings by low levels of selenium. J. Agric. Chem. Soc. Jpn. 2015, 79, 1579–1586. [Google Scholar]
- Henry, E.; Fung, N.; Liu, J.; Drakakaki, G.; Coaker, G. Beyond Glycolysis: GAPDHs Are Multi-functional Enzymes Involved in Regulation of ROS, Autophagy, and Plant Immune Responses. PLoS Genet. 2015, 11, e1005199. [Google Scholar] [CrossRef] [PubMed]
- Fei, Y.; Xue, Y.; Du, P.; Yang, S.; Deng, X. Expression analysis and promoter methylation under osmotic and salinity stress of TaGAPC1 in wheat (Triticum aestivum L). Protoplasma 2016, 254, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Deng, R.; Guo, Z.; Yang, S.; Deng, X. Genome-wide identification and characterization of Glyceraldehyde-3-phosphate dehydrogenase genes family in wheat (Triticum Aestivum). BMC Genom. 2016, 17, 240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.; Zhang, L.; Ji, H.; Xu, Z.; Zhou, Y.; Yang, S. The specific W-boxes of GAPC5 promoter bound by TaWRKY are involved in drought stress response in wheat. Plant Sci. 2020, 296, 110460. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Xu, Z.; Ji, H.; Zhou, Y.; Yang, S. TaWRKY40 transcription factor positively regulate the expression of TaGAPC1 to enhance drought tolerance. BMC Genom. 2019, 20, 795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rushton, P.J.; Somssich, I.E.; Ringler, P.; Shen, Q.J. WRKY transcription factors. Trends Plant Sci. 2010, 15, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Liu, S.; Meng, C.; Qin, L.; Kong, L.; Xia, G. WRKY Transcription Factors in Wheat and Their Induction by Biotic and Abiotic Stress. Plant Mol. Biol. Rep. 2013, 31, 1053–1067. [Google Scholar] [CrossRef]
- Xu, H.; Watanabe, K.A.; Zhang, L.; Shen, Q.J. WRKY transcription factor genes in wild rice Oryza nivara. DNA Res. 2016, 23, 311–323. [Google Scholar] [CrossRef] [Green Version]
- Wen, F.; Zhu, H.; Li, P.; Jiang, M.; Mao, W.; Chermaine, O.; Chu, Z. Genome-Wide Evolutionary Characterization and Expression Analyses of WRKY Family Genes in Brachypodium distachyon. DNA Res. 2014, 21, 327–339. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Ru, J.; Liu, Y.; Yang, J.; Li, M.; Xu, Z.; Fu, J. The Maize WRKY Transcription Factor ZmWRKY40 Confers Drought Resistance in Transgenic Arabidopsis. Int. J. Mol. Sci. 2018, 19, 2580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Chen, J.; Wang, L.; Wang, S. Genome-Wide Investigation of WRKY Transcription Factors Involved in Terminal Drought Stress Response in Common Bean. Front. Plant Sci. 2017, 8, 171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dou, L.; Zhang, X.; Pang, C.; Song, M.; Wei, H. Genome-wide analysis of the WRKY gene family in cotton. Mol. Genet. Genom. 2014, 289, 1103–1121. [Google Scholar] [CrossRef] [PubMed]
- Niu, X.; Luo, T.; Zhao, H.; Su, Y.; Li, H. Identification of wheat DREB genes and functional characterization of TaDREB3 in response to abiotic stresses. Gene 2020, 740, 144514. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Tang, Y.; Zhou, C.; Zhang, L.; Lv, J. A wheat WRKY transcription factor TaWRKY46 enhances tolerance to osmotic stress in transgenic Arabidopsis plants. Int. J. Mol. Sci. 2020, 21, 1321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, H.; Li, S.; Wang, Z.; Cheng, X.; Li, F.; Mei, F.; Chen, N.; Kang, Z. Regulatory changes in TaSNAC8-6A are associated with drought tolerance in wheat seedlings. Plant Biotechnol. J. 2020, 18, 1078–1092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Touzy, G.; Rincent, R.; Bogard, M.; Lafarge, S.; Dubreuil, P.; Mini, A.; Deswarte, J.; Beauchene, K.; Gouis, J.L.; Praud, S. Using environmental clustering to identify specific drought tolerance QTLs in bread wheat (T. aestivum L.). Theor. Appl. Genet. 2019, 132, 2859–2880. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.-H.; Zeng, Q.-D.; Tai, L.; Li, B.-B.; Zhang, P.-P.; Nie, X.-M.; Wang, P.-Q.; Liu, W.-T.; Li, W.-Q.; Kang, Z.-S.; et al. Interaction between TaNOX7 and TaCDPK13 Contributes to Plant Fertility and Drought Tolerance by Regulating ROS Production. J. Agric. Food Chem. 2020, 68, 7333–7347. [Google Scholar] [CrossRef]
- Li, X.; Wei, W.; Li, F.; Zhang, L.; Deng, X.; Liu, Y.; Yang, S. The Plastidial Glyceraldehyde-3-Phosphate Dehydrogenase Is Critical for Abiotic Stress Response in Wheat. Int. J. Mol. Sci. 2019, 20, 1104. [Google Scholar] [CrossRef] [Green Version]
- Jonathan, K.; Jason, I.; Michael, B.; Usdin, T.B. Evaluation of vector-primed cDNA library production from microgram quantities of total RNA. Nucleic Acids Res. 2004, 32, e183. [Google Scholar]
- Gao, J.X.; Jing, J.; Yu, C.J.; Chen, J. Construction of a High-Quality Yeast Two-Hybrid Library and Its Application in Identification of Interacting Proteins with Brn1 in Curvularia lunata. Plant Pathol. J. 2015, 31, 108–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhulidov, P.A.; Bogdanova, E.A.; Shcheglov, A.S.; Shagina, I.A.; Wagner, L.L.; Khazpekov, G.L.; Kozhemyako, V.V.; Lukyanov, S.A.; Shagin, D.A. A method for the preparation of normalized cDNA libraries enriched with full-length sequences. Russ. J. Bioorg. Chem. 2005, 31, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Peralta, D.A.; Alejandro, A.; Gomez-Casati, D.F.; Busi, M.V. Over-expression of SINAL7 increases biomass and drought tolerance, and also delays senescence in Arabidopsis. J. Biotechnol. 2018, 283, 11–21. [Google Scholar] [CrossRef]
- Yang, T.; Wang, L.; Li, C.; Liu, Y.; Zhu, S.; Qi, Y.; Liu, X.; Lin, Q.; Luan, S.; Yu, F. Receptor protein kinase FERONIA controls leaf starch accumulation by interacting with glyceraldehyde-3-phosphate dehydrogenase. Biochem. Biophys. Res. Commun. 2015, 465, 77–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Devaiah, S.P.; Zhang, W.; Welti, R. Signaling functions of phosphatidic acid. Prog. Lipid Res. 2006, 45, 250–278. [Google Scholar] [CrossRef]
- Hong, Y.; Pan, X.; Welti, R.; Wang, X. Phospholipase Dα3 Is Involved in the Hyperosmotic Response in Arabidopsis. Plant Cell 2008, 20, 803–816. [Google Scholar] [CrossRef] [Green Version]
- Sang, Y.; Zheng, S.; Li, W.; Huang, B.; Wang, X. Regulation of plant water loss by manipulating the expression of phospholipase Dα. Plant J. 2010, 28, 135–144. [Google Scholar] [CrossRef]
- Welti, R. Profiling Membrane Lipids in Plant Stress Responses role of phospholipase Dα in freezing-induced lipid changes in arabidopsis. J. Biol. Chem. 2002, 277, 31994–32002. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.; Sun, W.; Lv, H.; Luo, M.; Zeng, S.; Sitakanta, P.; Yuan, L.; Wang, Y.; John, S. A R2R3-MYB Transcription Factor from Epimedium sagittatum Regulates the Flavonoid Biosynthetic Pathway. PLoS ONE 2013, 8, e70778. [Google Scholar] [CrossRef]
- Xu, Z.S.; Ni, Z.Y.; Liu, L.; Nie, L.N.; Li, L.C.; Chen, M.; Ma, Y.Z. Characterization of the TaAIDFa gene encoding a CRT/DRE-binding factor responsive to drought, high-salt, and cold stress in wheat. Mol. Genet. Genom. 2008, 280, 497–508. [Google Scholar] [CrossRef]
- Ewing, B.; Hillier, L.D.; Wendl, M.C.; Green, P. Base-Calling of Automated Sequencer Traces UsingPhred. I. Accuracy Assessment. Genome Res. 1998, 8, 175–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Natarajan, P.; Kanagasabapathy, D.; Gunadayalan, G.; Panchalingam, J.; Shree, N.; Sugantham, P.A.; Singh, K.K.; Madasamy, P. Gene discovery from Jatropha curcas by sequencing of ESTs from normalized and full-length enriched cDNA library from developing seeds. BMC Genom. 2010, 11, 606. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.H.; Kim, N.S.; Lim, D.; Lee, K.T.; Oh, J.H.; Park, H.S.; Jang, G.W.; Kim, H.Y.; Jeon, M.; Choi, B.H. Generation and analysis of large-scale expressed sequence tags (ESTs) from a full-length enriched cDNA library of porcine backfat tissue. BMC Genom. 2006, 7, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T. Gene Ontology: Tool for the unification of biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Transcription profiles of TaGApC2 in Chinese spring wheat leaves under 20% PEG8000 (A) and 10 mM H2O2 (B). The β-actin gene was used as an internal reference. The vertical ordinate is the fold change; the horizontal ordinate is the treatment time. The data represent three independent experiments. The standard deviation (SD) is indicated at each point. Significant differences were assessed by one-sided paired t-tests (** p < 0.01).
Figure 1.
Transcription profiles of TaGApC2 in Chinese spring wheat leaves under 20% PEG8000 (A) and 10 mM H2O2 (B). The β-actin gene was used as an internal reference. The vertical ordinate is the fold change; the horizontal ordinate is the treatment time. The data represent three independent experiments. The standard deviation (SD) is indicated at each point. Significant differences were assessed by one-sided paired t-tests (** p < 0.01).

Figure 2.
The TaGApC2 promoter in Chinese spring wheat responds to abiotic stress in tobacco. (A) Schematic diagram of the distribution of the W-boxes in the TaGApC2 promoter. (B) The Rluc/Fluc enzyme activity suggested that TaGApC2 promoters are stress-inducible. The data represent three independent experiments. The standard deviation (SD) is indicated at each point. Significant differences were assessed by one-sided paired t-tests (** p < 0.01).
Figure 2.
The TaGApC2 promoter in Chinese spring wheat responds to abiotic stress in tobacco. (A) Schematic diagram of the distribution of the W-boxes in the TaGApC2 promoter. (B) The Rluc/Fluc enzyme activity suggested that TaGApC2 promoters are stress-inducible. The data represent three independent experiments. The standard deviation (SD) is indicated at each point. Significant differences were assessed by one-sided paired t-tests (** p < 0.01).

Figure 3.
Rluc/Fluc value analysis of the interaction between TaWRKYs and TaGApC2 promoters of Chinese spring wheat. TaGApC2-6A promoter and TaGApC2-6D promoter. The GUS effector was used as internal control. The data represent three independent experiments. The standard deviation (SD) is indicated at each point. Significant differences were assessed by one-sided paired t-tests.
Figure 3.
Rluc/Fluc value analysis of the interaction between TaWRKYs and TaGApC2 promoters of Chinese spring wheat. TaGApC2-6A promoter and TaGApC2-6D promoter. The GUS effector was used as internal control. The data represent three independent experiments. The standard deviation (SD) is indicated at each point. Significant differences were assessed by one-sided paired t-tests.

Figure 4.
Detection of transgenic plants. (A) Diagram of the 35S:TaGAPC2-6D constructs. TaGApC2-6D CDS was fused with the GFP-coding region driven by a 35S promoter. The primers pr1, pr2, and pr3, used to analyze TaGApC2-6D and TaGApCs-GFp in transgenic Arabidopsis plants, are shown. (B) PCR analysis of TaGApC2-6D over-expressing transgenic Arabidopsis using primer pairs of pr1 and pr2. (C) PCR analysis of TaGApC2-6D over-expressing transgenic Arabidopsis using primer pairs of pr1 and pr3. (D) Quantitative real-time PCR (qRT-PCR) validation of TaGApC2-6D in the aboveground part of the three-week-old Arabidopsis plant. The AtTubulin gene was used as an internal reference (*** p < 0.001). (E) The enzyme activities of glyceraldehyde-3-P dehydrogenase (GAPDH) in the aboveground part of the three-week-old Arabidopsis plant (** p < 0.01).
Figure 4.
Detection of transgenic plants. (A) Diagram of the 35S:TaGAPC2-6D constructs. TaGApC2-6D CDS was fused with the GFP-coding region driven by a 35S promoter. The primers pr1, pr2, and pr3, used to analyze TaGApC2-6D and TaGApCs-GFp in transgenic Arabidopsis plants, are shown. (B) PCR analysis of TaGApC2-6D over-expressing transgenic Arabidopsis using primer pairs of pr1 and pr2. (C) PCR analysis of TaGApC2-6D over-expressing transgenic Arabidopsis using primer pairs of pr1 and pr3. (D) Quantitative real-time PCR (qRT-PCR) validation of TaGApC2-6D in the aboveground part of the three-week-old Arabidopsis plant. The AtTubulin gene was used as an internal reference (*** p < 0.001). (E) The enzyme activities of glyceraldehyde-3-P dehydrogenase (GAPDH) in the aboveground part of the three-week-old Arabidopsis plant (** p < 0.01).

Figure 5.
The phenotype of TaGApC2-6D over-expressing Arabidopsis plants after withholding water for 25 days. D, drought; R, re-watered; WT, wild type.
Figure 5.
The phenotype of TaGApC2-6D over-expressing Arabidopsis plants after withholding water for 25 days. D, drought; R, re-watered; WT, wild type.

Figure 6.
Physiological changes and tolerance assay in wild-type and transgenic plants under drought stress. (A) The survival rate of wild-type and transgenic Arabidopsis plants after drought treatment. At least 50 plants were counted and averaged for each line (** p < 0.01). (B) The relative water content (RWC) of wild-type and transgenic Arabidopsis plants after drought treatment. (C) The chlorophyll content of wild-type and transgenic Arabidopsis plants after drought treatment. The data represent three independent experiments. Asterisks indicate a significant difference between WT and transgenic Arabidopsis lines.
Figure 6.
Physiological changes and tolerance assay in wild-type and transgenic plants under drought stress. (A) The survival rate of wild-type and transgenic Arabidopsis plants after drought treatment. At least 50 plants were counted and averaged for each line (** p < 0.01). (B) The relative water content (RWC) of wild-type and transgenic Arabidopsis plants after drought treatment. (C) The chlorophyll content of wild-type and transgenic Arabidopsis plants after drought treatment. The data represent three independent experiments. Asterisks indicate a significant difference between WT and transgenic Arabidopsis lines.

Figure 7.
Total root lengths of transgenic Arabidopsis lines under mock drought stress. Phenotypes of WT and TaGApC2-6D transgenic Arabidopsis seedlings under 1/2 MS medium with or without 6% PEG8000. (A) Root lengths phenotype of WT and TaGApC2-6D transgenic Arabidopsis seedlings grown on 1/2 MS medium with or without 6% PEG8000 for seven days. (B) Root lengths of WT and TaGApC2-6D transgenic Arabidopsis seedlings grown on 1/2 MS medium with or without 6% PEG8000 for seven days. Data are means ± SD of three independent experiments, and asterisks indicate a significant difference between WT and transgenic Arabidopsis lines (** p < 0.01).
Figure 7.
Total root lengths of transgenic Arabidopsis lines under mock drought stress. Phenotypes of WT and TaGApC2-6D transgenic Arabidopsis seedlings under 1/2 MS medium with or without 6% PEG8000. (A) Root lengths phenotype of WT and TaGApC2-6D transgenic Arabidopsis seedlings grown on 1/2 MS medium with or without 6% PEG8000 for seven days. (B) Root lengths of WT and TaGApC2-6D transgenic Arabidopsis seedlings grown on 1/2 MS medium with or without 6% PEG8000 for seven days. Data are means ± SD of three independent experiments, and asterisks indicate a significant difference between WT and transgenic Arabidopsis lines (** p < 0.01).

Figure 8.
Changes in O2− and H2O2 levels in wild-type (WT) and transgenic plants subjected to drought stress. Drought-stressed seedlings were incubated in nitroblue tetrazolium (NBT) or diaminobenzidine (DAB) solution. Blue staining indicates O2− accumulation (A). Brown staining indicates H2O2 accumulation (B). Control, plants growing under normal conditions; drought, plants growing after drought stress treatment (12 days).
Figure 8.
Changes in O2− and H2O2 levels in wild-type (WT) and transgenic plants subjected to drought stress. Drought-stressed seedlings were incubated in nitroblue tetrazolium (NBT) or diaminobenzidine (DAB) solution. Blue staining indicates O2− accumulation (A). Brown staining indicates H2O2 accumulation (B). Control, plants growing under normal conditions; drought, plants growing after drought stress treatment (12 days).

Figure 9.
Effects of drought stress on superoxide dismutase (SOD) (A), peroxidase (POD) (B), and malondialdehyde (MDA) levels (C) in WT and transgenic plants after drought stress. The data represent three independent experiments.
Figure 9.
Effects of drought stress on superoxide dismutase (SOD) (A), peroxidase (POD) (B), and malondialdehyde (MDA) levels (C) in WT and transgenic plants after drought stress. The data represent three independent experiments.

Figure 10.
Functional annotation and classification of 804 Triticum aestivum annotated UniESTs using the BLAST2GO software. The three GO categories, cellular component (A), molecular function (B), and biological process (C), are presented.
Figure 10.
Functional annotation and classification of 804 Triticum aestivum annotated UniESTs using the BLAST2GO software. The three GO categories, cellular component (A), molecular function (B), and biological process (C), are presented.

Figure 11.
Functional annotation and classification of 804 Triticum aestivum annotated UniESTs using the KEGG pathway.
Figure 11.
Functional annotation and classification of 804 Triticum aestivum annotated UniESTs using the KEGG pathway.

Figure 12.
Identification of TaPLDδ interacting with TaGAPC2-6D. (A) Confirmation of true positive clones by small-scale yeast two-hybrid (Y2H) assay. SD/-Leu/-Trp/X-α-Gal (DDO/X), SD/-Leu/-Trp/X-α-Gal; SD/-Ade/-His/-Leu/-Trp/X-α-Gal/AbA (QDO/X/A), SD/-Ade/-His/-Leu/-Trp/X-α-Gal/AbA. PC indicates a positive control. (B) Bimolecular fluorescence complementation assay (BiFC) assay of the interaction between TaPLDδ and TaGAPC2-6D protein in tobacco leaf protoplasts. The pSPYNE-TaGAPC2-6D and pSPYCE-TaPLDδ constructs were co-infiltrated into tobacco by Agrobacterium. YFP fluorescence was detected by confocal laser scanning microscopy. Co-transformants of pSPYNE-TaGAPC2-6D and pSPYCE were used as negative controls. Scalebar = 100 μm.
Figure 12.
Identification of TaPLDδ interacting with TaGAPC2-6D. (A) Confirmation of true positive clones by small-scale yeast two-hybrid (Y2H) assay. SD/-Leu/-Trp/X-α-Gal (DDO/X), SD/-Leu/-Trp/X-α-Gal; SD/-Ade/-His/-Leu/-Trp/X-α-Gal/AbA (QDO/X/A), SD/-Ade/-His/-Leu/-Trp/X-α-Gal/AbA. PC indicates a positive control. (B) Bimolecular fluorescence complementation assay (BiFC) assay of the interaction between TaPLDδ and TaGAPC2-6D protein in tobacco leaf protoplasts. The pSPYNE-TaGAPC2-6D and pSPYCE-TaPLDδ constructs were co-infiltrated into tobacco by Agrobacterium. YFP fluorescence was detected by confocal laser scanning microscopy. Co-transformants of pSPYNE-TaGAPC2-6D and pSPYCE were used as negative controls. Scalebar = 100 μm.

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhang, L.; Zhang, H.; Yang, S. Cytosolic TaGAPC2 Enhances Tolerance to Drought Stress in Transgenic Arabidopsis Plants. Int. J. Mol. Sci. 2020, 21, 7499. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21207499
AMA Style
Zhang L, Zhang H, Yang S. Cytosolic TaGAPC2 Enhances Tolerance to Drought Stress in Transgenic Arabidopsis Plants. International Journal of Molecular Sciences. 2020; 21(20):7499. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21207499
Chicago/Turabian StyleZhang, Lin, Hanwen Zhang, and Shushen Yang. 2020. "Cytosolic TaGAPC2 Enhances Tolerance to Drought Stress in Transgenic Arabidopsis Plants" International Journal of Molecular Sciences 21, no. 20: 7499. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21207499
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.