Identification of Viruses and Viroids Infecting Tomato and Pepper Plants in Vietnam by Metatranscriptomics

 ,
, 
Abstract
:1. Introduction
2. Results
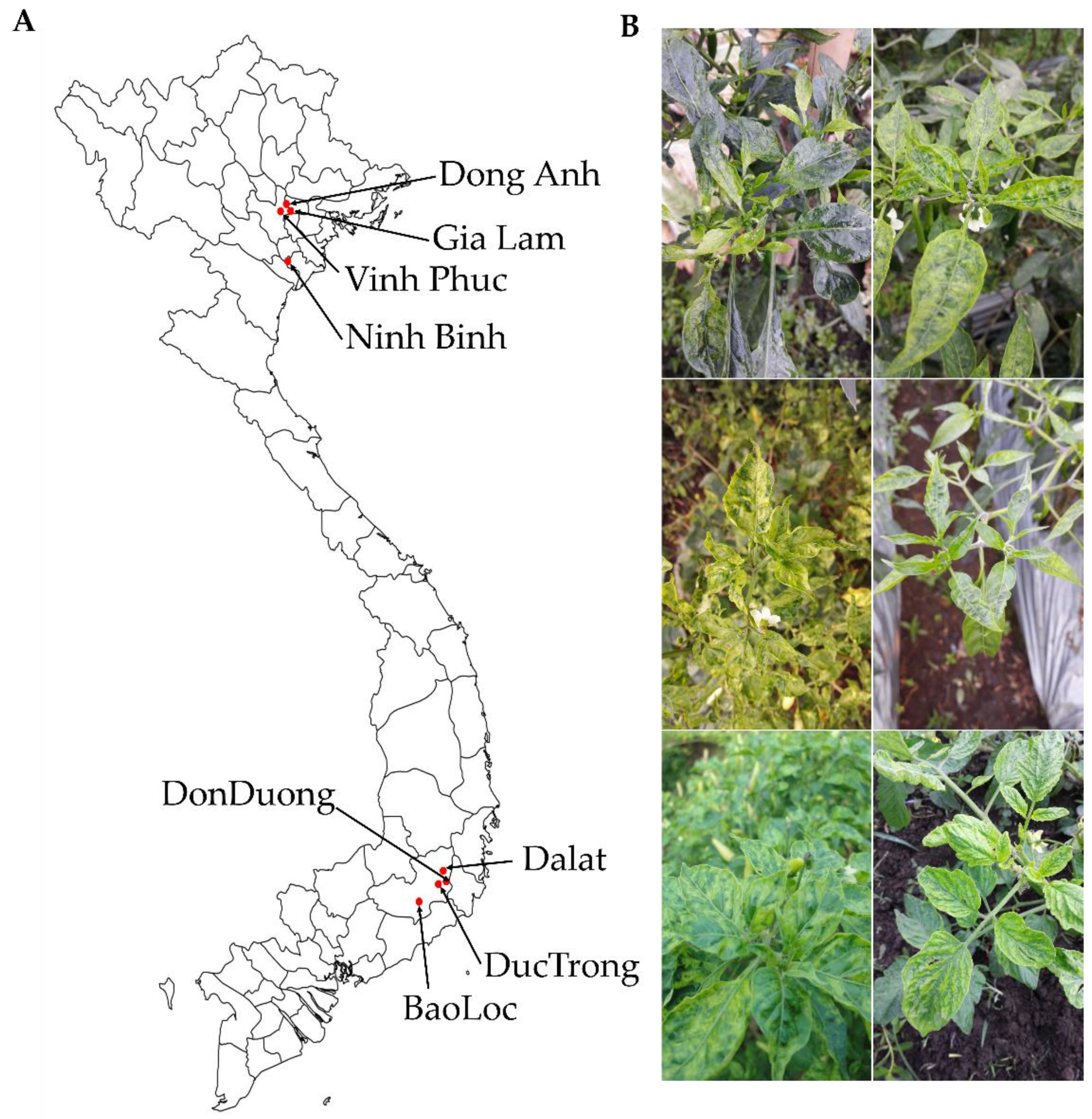
2.1. Collection of Leaf Samples and Generation of Libraries for Identification of Viruses Infecting Tomato and Pepper Plants
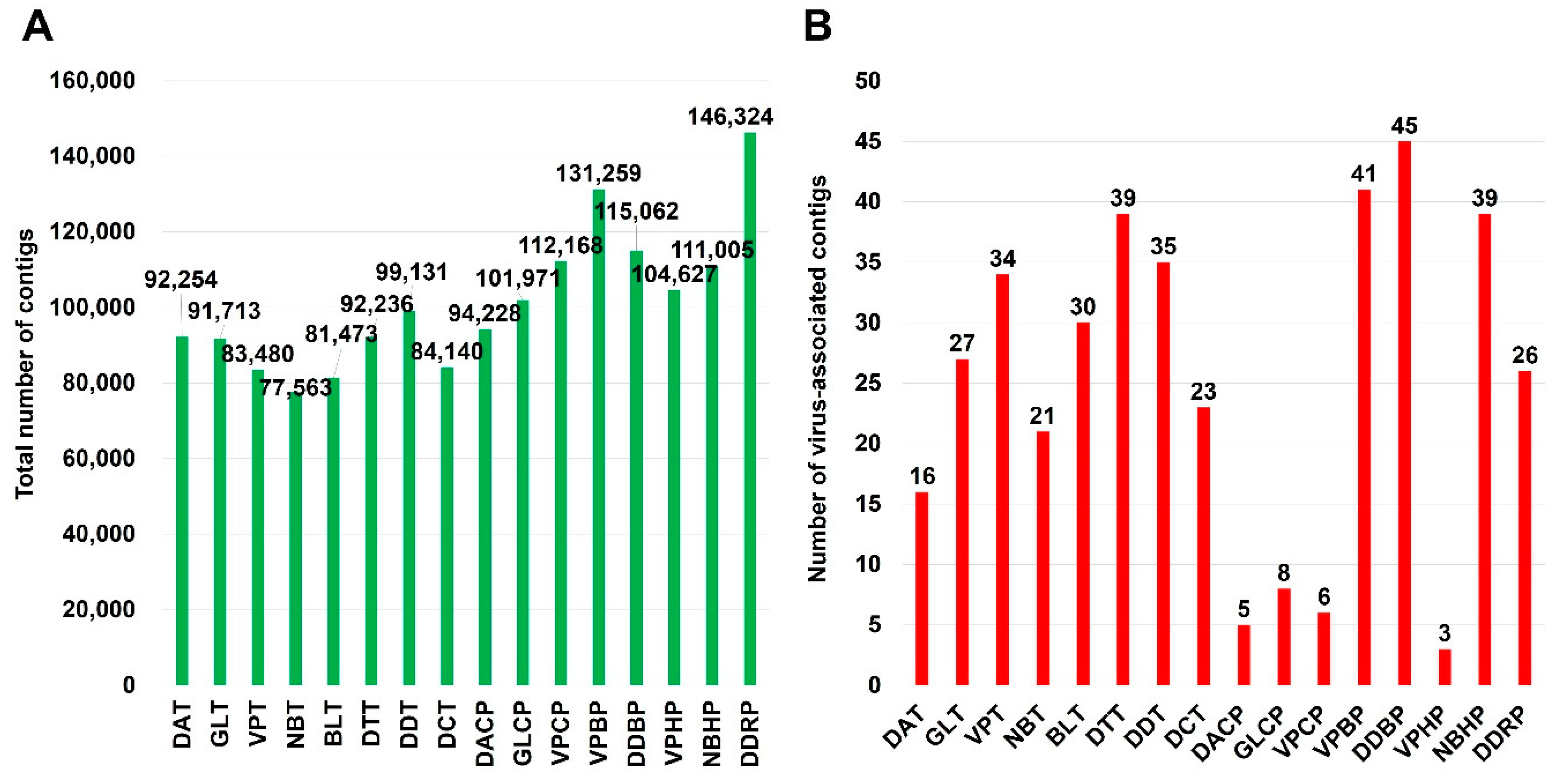
2.2. Identification of Virus-Associated Contigs
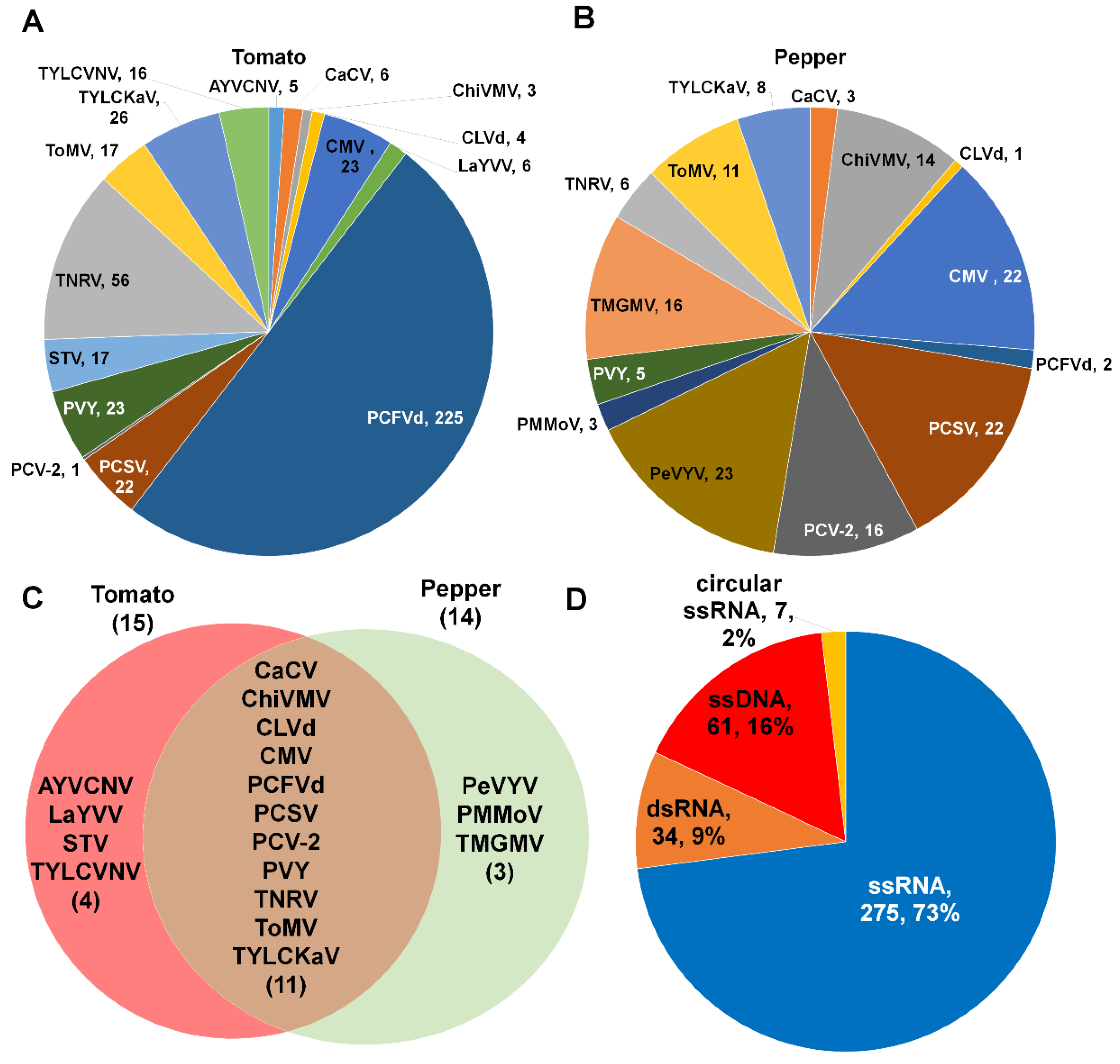
2.3. Classification of Identified Virus-Associated Contigs According to Virus Taxonomy
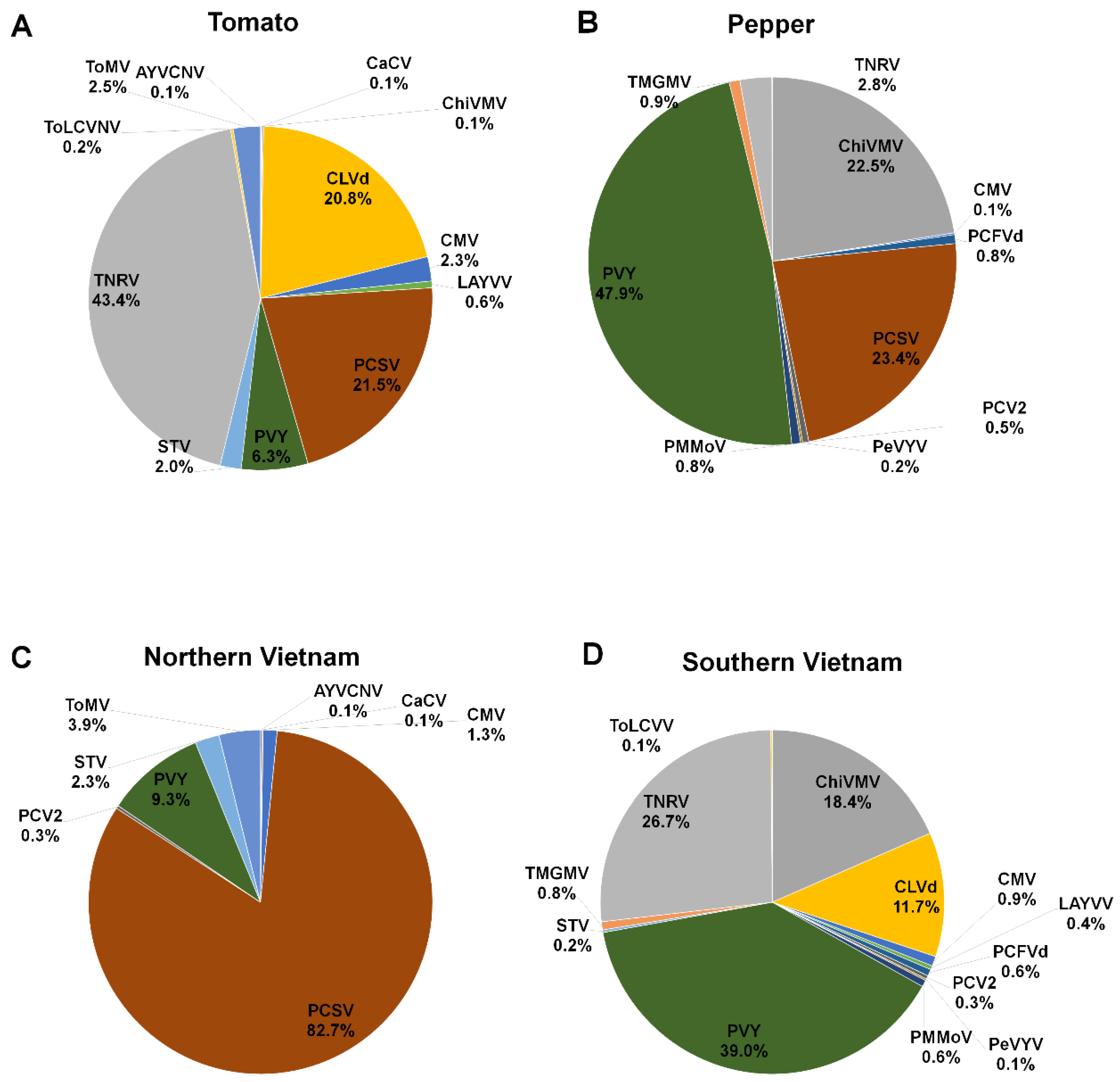
2.4. Proportion of Identified Viruses and Viroids According to Virus-Associated Contigs
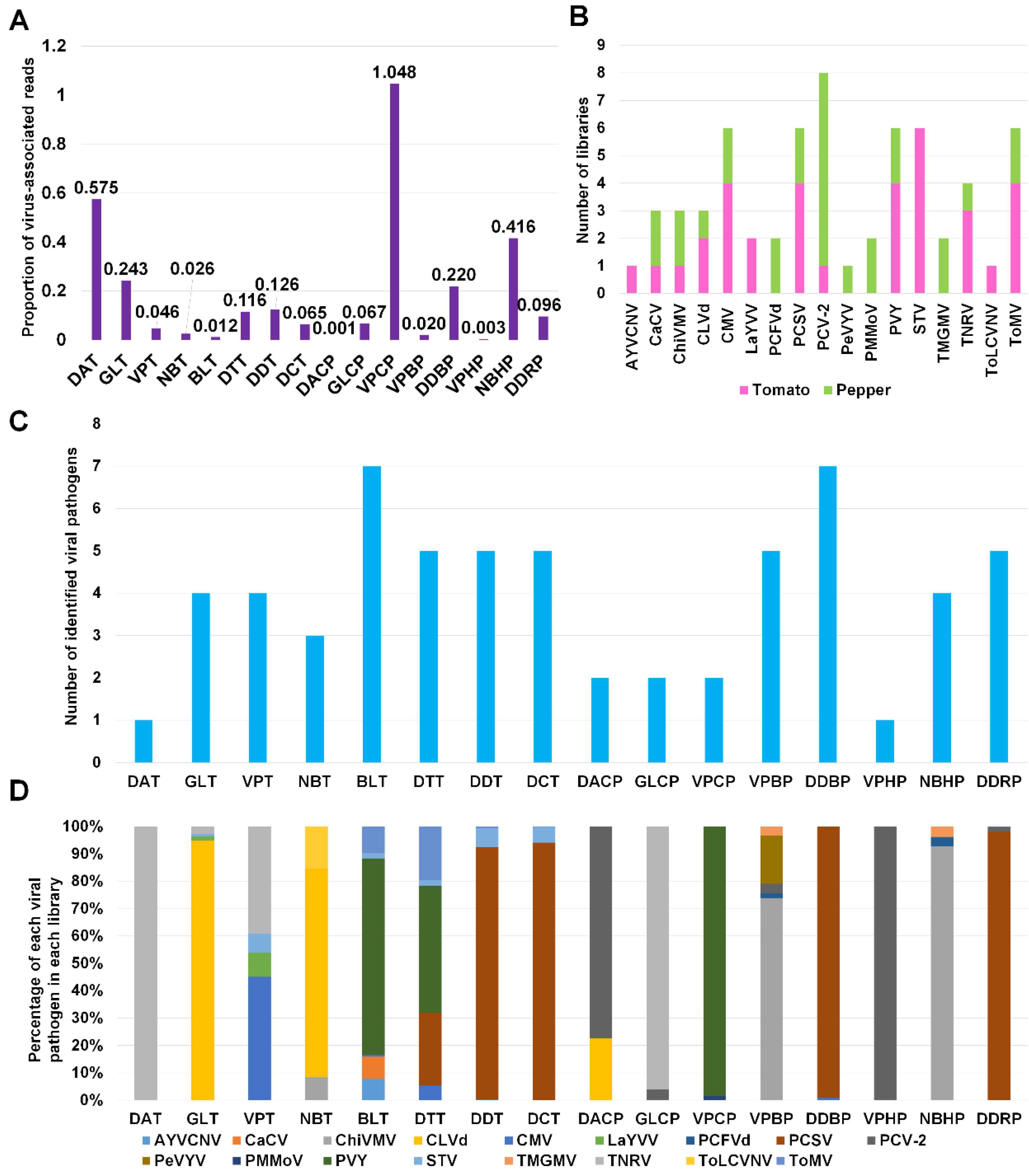
2.5. Proportion of Identified Viruses and Viroids According to Virus-Associated Reads
2.6. Proportion of Identified Viruses and Viroids According to Plant Host and Geographical Region
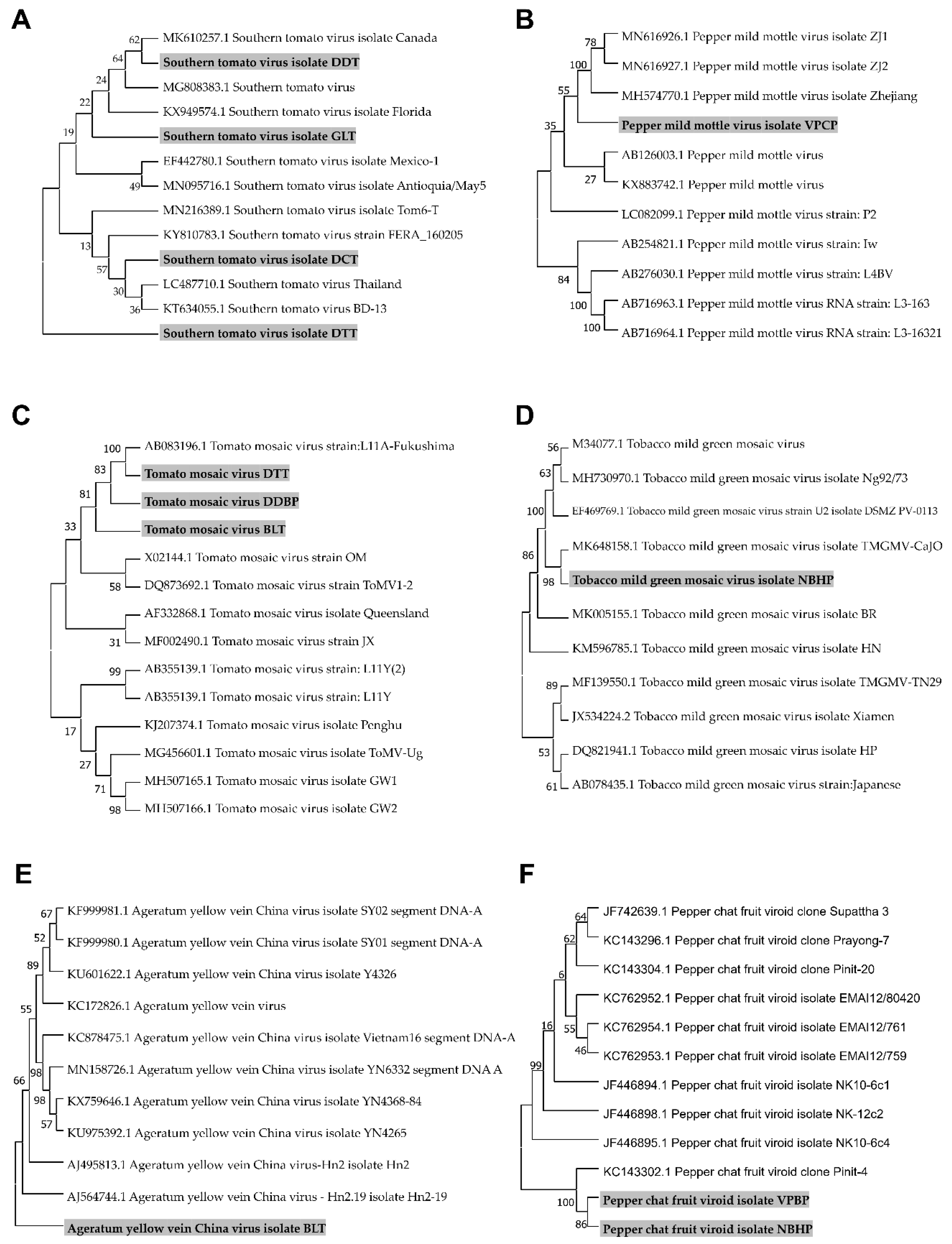
2.7. Phylogenetic Analyses for Identified Viruses and a Viroid
2.8. Validation of Results for RNA-Seq by RT-PCR
3. Discussion
4. Materials and Methods
4.1. Sample Collection and RNA Sequencing
4.2. Bioinformatic Analyses
4.3. Construction of Phylogenetic Trees
4.4. Reverse Transcription–Polymerase Chain Reaction (RT-PCR) to Validate Infection of Identified Viruses and Viroids
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Blancard, D. Tomato Diseases: Identification, Biology and Control: A Colour Handbook; CRC Press: Boca Raton, FL, USA, 2012. [Google Scholar]
- Ganesan, M.; Rajesh, M.; Solairaj, P.; Senthilkumar, T. Tomato as a pioneer in health management. Int. J. Pharm. Chem. Biol. Sci. 2012, 2, 210–217. [Google Scholar]
- Faustino, J.; Barroca, M.; Guiné, R. Study of the Drying Kinetics of Green Bell Pepper and Chemical Characterization. Food Bioprod. Process. 2007, 85, 163–170. [Google Scholar] [CrossRef]
- Blawid, R.; Van, D.; Maiss, E. Transreplication of a Tomato yellow leaf curl Thailand virus DNA-B and replication of a DNAß component by Tomato leaf curl Vietnam virus and Tomato yellow leaf curl Vietnam virus. Virus Res. 2008, 136, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Pernezny, K.; Roberts, P.D.; Murphy, J.F.; Goldberg, N.P. Compendium of Pepper Diseases; APS Press: St. Paul, MN, USA, 2003. [Google Scholar]
- Pospieszny, H.; Borodynko-Filas, N.; Hasiów-Jaroszewska, B.; Czerwonka, B.; Elena, S.F. An assessment of the transmission rate of Tomato black ring virus through tomato seeds. Plant. Prot. Sci. 2019, 56, 9–12. [Google Scholar] [CrossRef] [Green Version]
- Thakur, H.; Jindal, S.K.; Sharma, A.; Dhaliwal, M.S. Chilli leaf curl virus disease: A serious threat for chilli cultivation. J. Plant. Dis. Prot. 2018, 125, 239–249. [Google Scholar] [CrossRef]
- Yanagisawa, H.; Matsushita, Y. Host ranges European Journal of Plant Pathology and seed transmission of Tomato planta macho viroid and Pepper chat fruit viroid. Eur. J. Plant. Pathol. 2017, 149, 211–217. [Google Scholar] [CrossRef]
- Constable, F.E.; Chambers, G.A.; Penrose, L.; Daly, A.; Mackie, J.; Davis, K.; Rodoni, B.; Gibbs, M. Viroid-infected Tomato and Capsicum Seed Shipments to Australia. Viruses 2019, 11, 98. [Google Scholar] [CrossRef] [Green Version]
- Batuman, O.; Turini, T.A.; Oliveira, P.V.; Rojas, M.R.; Macedo, M.; Mellinger, H.C.; Adkins, S.; Gilbertson, R.L. First Report of a Resistance-Breaking Strain of Tomato spotted wilt virus Infecting Tomatoes With the Sw-5 Tospovirus-Resistance Gene in California. Plant. Dis. 2017, 101, 637. [Google Scholar] [CrossRef]
- Ghanim, M.; Czosnek, H. Tomato Yellow Leaf Curl Geminivirus (TYLCV-Is) Is Transmitted among Whiteflies (Bemisia tabaci) in a Sex-Related Manner. J. Virol. 2000, 74, 4738–4745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massart, S.; Olmos, A.; Jijakli, H.; Candresse, T. Current impact and future directions of high throughput sequencing in plant virus diagnostics. Virus Res. 2014, 188, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Roossinck, M.J.; Martin, D.P.; Roumagnac, P. Plant Virus Metagenomics: Advances in Virus Discovery. Phytopathology 2015, 105, 716–727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Q.; Ding, S.-W.; Zhang, Y.; Zhu, S. Identification of Viruses and Viroids by Next-Generation Sequencing and Homology-Dependent and Homology-Independent Algorithms. Annu. Rev. Phytopathol. 2015, 53, 425–444. [Google Scholar] [CrossRef]
- Candresse, T.; Marais, A.; Sorrentino, R.; Faure, C.; Theil, S.; Cadot, V.; Rolland, M.; Villemot, J.; Rabenstein, F. Complete genomic sequence of barley (Hordeum vulgare) endornavirus (HvEV) determined by next-generation sequencing. Arch. Virol. 2015, 161, 741–743. [Google Scholar] [CrossRef] [PubMed]
- Yan, T.; Zhu, J.-R.; Di, D.; Gao, Q.; Zhang, Y.; Zhang, A.; Yan, C.; Miao, H.; Wang, X.-B. Characterization of the complete genome of Barley yellow striate mosaic virus reveals a nested gene encoding a small hydrophobic protein. Virology 2015, 478, 112–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bae, M.; Jo, Y.; Choi, H.; Tran, P.-T.; Kim, K.-H. First report of tomato mosaic virus isolated from tomato and pepper in Vietnam. J. Plant. Pathol. 2018, 101, 181. [Google Scholar] [CrossRef] [Green Version]
- Choi, H.; Jo, Y.; Tran, P.-T.; Kim, K.-H. First report of ageratum yellow vein virus infecting tomato in Vietnam. J. Plant. Pathol. 2019, 101, 1267. [Google Scholar] [CrossRef] [Green Version]
- Cuong, H.V.; Hai, L.; Tiep, T.; Hao, N. Molecular characterization of Tomato leaf curl Hainan virus and Tomato leaf curl Hanoi virus in Vietnam. Int. Soc. Southeast. Asian Agric. Sci. J. 2011, 2, 70–82. [Google Scholar]
- Ha, C.; Revill, P.; Harding, R.M.; Vu, M.; Dale, J.L. Identification and sequence analysis of potyviruses infecting crops in Vietnam. Arch. Virol. 2007, 153, 45–60. [Google Scholar] [CrossRef]
- Maree, H.J.; Fox, A.; Al Rwahnih, M.; Boonham, N.; Candresse, T. Application of HTS for Routine Plant Virus Diagnostics: State of the Art and Challenges. Front. Plant. Sci. 2018, 9, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Revill, P.; Ha, C.V.; Porchun, S.C.; Vu, M.T.; Dale, J.L. The complete nucleotide sequence of two distinct geminiviruses infecting cucurbits in Vietnam. Arch. Virol. 2003, 148, 1523–1541. [Google Scholar] [CrossRef] [PubMed]
- Jo, Y.; Choi, H.; Kim, S.-M.; Kim, S.-L.; Lee, B.C.; Cho, W.K. The pepper virome: Natural co-infection of diverse viruses and their quasispecies. BMC Genom. 2017, 18, 453. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Gao, S.; Hernandez, A.G.; Wechter, W.P.; Fei, Z.; Ling, K.-S. Deep Sequencing of Small RNAs in Tomato for Virus and Viroid Identification and Strain Differentiation. PLoS ONE 2012, 7, e37127. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Sun, X.; Taylor, A.; Jiao, C.; Xu, Y.; Cai, X.; Wang, X.; Ge, C.; Pan, G.; Wang, Q.; et al. Diversity, Distribution, and Evolution of Tomato Viruses in China Uncovered by Small RNA Sequencing. J. Virol. 2017, 91, e00173-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fadhila, C.; Lal, A.; Vo, T.T.; Ho, P.T.; Hidayat, S.H.; Lee, J.; Kil, E.-J.; Lee, S. The threat of seed-transmissible pepper yellow leaf curl Indonesia virus in chili pepper. Microb. Pathog. 2020, 143, 104132. [Google Scholar] [CrossRef] [PubMed]
- Kil, E.-J.; Park, J.; Choi, E.-Y.; Byun, H.-S.; Lee, K.-Y.; An, C.G.; Kim, C.-S. Seed transmission of Tomato yellow leaf curl virus in sweet pepper (Capsicum annuum). Eur. J. Plant. Pathol. 2017, 150, 759–764. [Google Scholar] [CrossRef]
- Kil, E.-J.; Kim, S.; Lee, Y.-J.; Byun, H.-S.; Park, J.; Seo, H.; Kim, C.-S.; Shim, J.-K.; Lee, J.-H.; Kim, J.-K.; et al. Tomato yellow leaf curl virus (TYLCV-IL): A seed-transmissible geminivirus in tomatoes. Sci. Rep. 2016, 6, 19013. [Google Scholar] [CrossRef] [Green Version]
- Pospieszny, H.; Borodynko, N.; Hasiów-Jaroszewska, B.; Rymelska, N.; Elena, S.F. Transmission rate of two Polish Tomato torrado virus isolates through tomato seeds. J. Gen. Plant. Pathol. 2018, 85, 109–115. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Padilla, V.; Fortes, I.M.; Romero-Rodríguez, B.; Arroyo-Mateos, M.; Castillo, A.G.; Moyano, C.; De León, L.; Moriones, E. Revisiting Seed Transmission of the Type Strain of Tomato yellow leaf curl virus in Tomato Plants. Phytopathology 2020, 110, 121–129. [Google Scholar] [CrossRef]
- Jo, Y.; Bae, J.-Y.; Kim, S.-M.; Choi, H.; Lee, B.C.; Cho, W.K. Barley RNA viromes in six different geographical regions in Korea. Sci. Rep. 2018, 8, 13237. [Google Scholar] [CrossRef] [PubMed]
- Jo, Y.; Kim, S.-M.; Choi, H.; Yang, J.W.; Lee, B.C.; Cho, W.K. Sweet potato viromes in eight different geographical regions in Korea and two different cultivars. Sci. Rep. 2020, 10, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Rojas, M.R.; Macedo, M.A.; Maliano, M.R.; Soto-Aguilar, M.; Souza, J.O.; Briddon, R.W.; Kenyon, L.; Bustamante, R.F.R.; Zerbini, F.M.; Adkins, S.; et al. World Management of Geminiviruses. Annu. Rev. Phytopathol. 2018, 56, 637–677. [Google Scholar] [CrossRef] [PubMed]
- Sabanadzovic, S.; Valverde, R.A. Properties and detection of two cryptoviruses from pepper (Capsicum annuum). Virus Genes 2011, 43, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Sabanadzovic, S.; Valverde, R.A.; Brown, J.K.; Martin, R.R.; Tzanetakis, I. Southern tomato virus: The link between the families Totiviridae and Partitiviridae. Virus Res. 2009, 140, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Choi, G.-S.; Kim, J.-H.; Lee, D.-H.; Kim, J.-S.; Ryu, K.H. Occurrence and Distribution of Viruses Infecting Pepper in Korea. Plant. Pathol. J. 2005, 21, 258–261. [Google Scholar] [CrossRef] [Green Version]
- Olawale, A.; Samuel, B.O.; Solomon, A.S.O.; Kumar, P.L. Surveys of virus diseases on pepper (Capsicum spp.) in South-west Nigeria. Afr. J. Biotechnol. 2015, 14, 3198–3205. [Google Scholar]
- Sepúlveda, P.R.; Larraín, P.S.; Quiroz, C.E.; Rebufel, P.A.; Graña, F.S. Identification and incidence of pepper viruses in north central Chile and its association with vectors. Agric. Tec. 2005, 65, 235–245. [Google Scholar]
- Jo, Y.; Lian, S.; Chu, H.; Cho, J.K.; Yoo, S.-H.; Choi, H.; Yoon, J.-Y.; Choi, S.-K.; Lee, B.C.; Cho, W.K. Peach RNA viromes in six different peach cultivars. Sci. Rep. 2018, 8, 1844. [Google Scholar] [CrossRef] [Green Version]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [Green Version]
- Larkin, M.; Blackshields, G.; Brown, N.; Chenna, R.; Mcgettigan, P.; McWilliam, H.; Valentin, F.; Wallace, I.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, P.O.; Kumar, S.; Tamura, K.; Nei, M. MEGA: Molecular Evolutionary Genetics Analysis, Version 1.02. Syst. Biol. 1995, 44, 576. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers * | Nucleotide Sequence | Expected Size |
|---|---|---|
| PVY_diag_F | ATGGCAAATGACACAATCGATGCAG | 804 bp |
| PVY_diag_R | TCACATGTTCTTAACTCCAAGTAGAGTATG | |
| CMV_diag_F | ATGGACAAATCTGAATCAACCAGTG | 754 bp |
| CMV_diag_R | GACTGGGAGCACTCCAGATG | |
| ToMV_diag_F | CGAGAGGGGCAACAAACAT | 317 bp |
| ToMV_diag_R | ACCTGTCTCCATCTCTTTGG | |
| AYVCNV_diag_F | CTGATGTGCCCAAAGGTTGT | 406 bp |
| AYVCNV_diag_R | GCCTGCTCCTTAGACGCATA | |
| TYLCKaV_diag_F | TGCCGAAGCGTTCAATAGAT | 733 bp |
| TYLCKaV_diag_R | TGTTGCATACACAGGATTAGAGG | |
| CaCV_diag_F | AAGACCTCGAAAGAGGCAAA | 703 bp |
| CaCV_diag_R | CTTCGGAGGCAAACTATTGG | |
| TNRV_diag_F | TTGCTAGCTGGAGGAGAAGC | 786 bp |
| TNRV_diag_R | TCCTCTCCTAGTTGGCTTGC | |
| PCV2_diag_F | TTCAATCGACGGTTTCACAA | 792 bp |
| PCV2_diag_R | CCTTGACTTGAGGTCGTGGT | |
| STV_diag_F | CAAAGGGAAGACTGCTGAGG | 808 bp |
| STV_diag_R | AGCCTCTCCATCGGGATTAT | |
| ChiVMV_diag_F | GCGTAAAAGGCGAAGACTCA | 782 bp |
| ChiVMV_diag_R | GTGCCGTTCAGTGTCCTCTT | |
| PMMoV_diag_F | ATGGCTTACACAGTTTCCAGTGCCAA | 474 bp |
| PMMoV_diag_R | TTAAGGAGTTGTAGCCCAGGTGAGTCC | |
| TMGMV_diag_F | ATGCCTTATACAATCAACTCTCCG | 480 bp |
| TMGMV_diag_R | CTAAGTAGCCGGAGTTGTGGTC | |
| PeVYV_diag_F | ATGAATACGGGAGGGGTTAGG | 621 bp |
| PeVYV_diag_R | CTATTTCGGGTTGTGCAATTGC | |
| LaYVV_diag_F | ATGTCGAAGCGACCTGCAGATAT | 741 bp |
| LaYVV_diag_R | GATTTTCAGAGTAGCATACACGGGA | |
| CLVd_diag_F | CGGAACTAAACTCGTGGTTCCTG | 370 bp |
| CLVd_diag_R | AGGAACCTACTGCGGTTCCA | |
| PCFVd_diag_F | CCGGATTCTTCTAAGGGTGCCT | 317 bp |
| PCFVd_diag_R | AGATCCTCTCGGGTCCCGG | |
| ToLCVV_diag_F | GCGTTAATGCGTCCCATAAT | 528 bp |
| ToLCVV_diag_R | GCATTAAAGTCGTGGGCAAT | |
| TYLCVNV_diag_F | AGAAACGCCAAGTCTGAGGA | 314 bp |
| TYLCVNV_diag_R | GTTCGGAGACGGAGAGTTGA |
| Region * | Host Plant | Library | Total Read Bases (bp) | Total Reads | GC (%) |
|---|---|---|---|---|---|
| Dong Anh | Tomato | DAT | 5,592,948,326 | 55,375,726 | 39.75 |
| Gia Lam | Tomato | GLT | 6,595,539,572 | 65,302,372 | 42.59 |
| Vinh Phuc | Tomato | VPT | 6,078,200,604 | 60,180,204 | 42.64 |
| Ninh Binh | Tomato | NBT | 4,899,434,654 | 48,509,254 | 41.2 |
| Bao Loc city | Tomato | BLT | 6,453,511,150 | 63,896,150 | 42.39 |
| Duc Trong | Tomato | DTT | 8,004,172,432 | 79,249,232 | 42.7 |
| Don Duong | Tomato | DDT | 7,918,928,432 | 78,405,232 | 43.8 |
| Dalat | Tomato | DCT | 8,257,251,768 | 81,754,968 | 43.67 |
| DongAnh | Chili pepper | DACP | 5,522,312,764 | 54,676,364 | 41.07 |
| Gia Lam | Chili pepper | GLCP | 4,912,867,452 | 48,642,252 | 41.24 |
| Vinh Phuc | Chili pepper | VPCP | 5,229,970,486 | 51,781,886 | 40.54 |
| Vinh Phuc | Bell pepper | VPBP | 5,745,072,910 | 56,881,910 | 42.45 |
| Don Duong | Bell pepper | DDBP | 8,473,848,288 | 83,899,488 | 44.83 |
| Vinh Phuc | Hot pepper | VPHP | 5,498,428,890 | 54,439,890 | 42.86 |
| Ninh Binh | Hot pepper | NBHP | 6,356,471,966 | 62,935,366 | 42.25 |
| Don Duong | Red pepper | DDRP | 8,339,452,436 | 82,568,836 | 43.24 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, H.; Jo, Y.; Cho, W.K.; Yu, J.; Tran, P.-T.; Salaipeth, L.; Kwak, H.-R.; Choi, H.-S.; Kim, K.-H. Identification of Viruses and Viroids Infecting Tomato and Pepper Plants in Vietnam by Metatranscriptomics. Int. J. Mol. Sci. 2020, 21, 7565. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21207565
Choi H, Jo Y, Cho WK, Yu J, Tran P-T, Salaipeth L, Kwak H-R, Choi H-S, Kim K-H. Identification of Viruses and Viroids Infecting Tomato and Pepper Plants in Vietnam by Metatranscriptomics. International Journal of Molecular Sciences. 2020; 21(20):7565. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21207565
Chicago/Turabian StyleChoi, Hoseong, Yeonhwa Jo, Won Kyong Cho, Jisuk Yu, Phu-Tri Tran, Lakha Salaipeth, Hae-Ryun Kwak, Hong-Soo Choi, and Kook-Hyung Kim. 2020. "Identification of Viruses and Viroids Infecting Tomato and Pepper Plants in Vietnam by Metatranscriptomics" International Journal of Molecular Sciences 21, no. 20: 7565. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21207565