Adipokines and Inflammation: Focus on Cardiovascular Diseases

 , , , , ,
, , , , , {kind=link}
Abstract
:1. Introduction
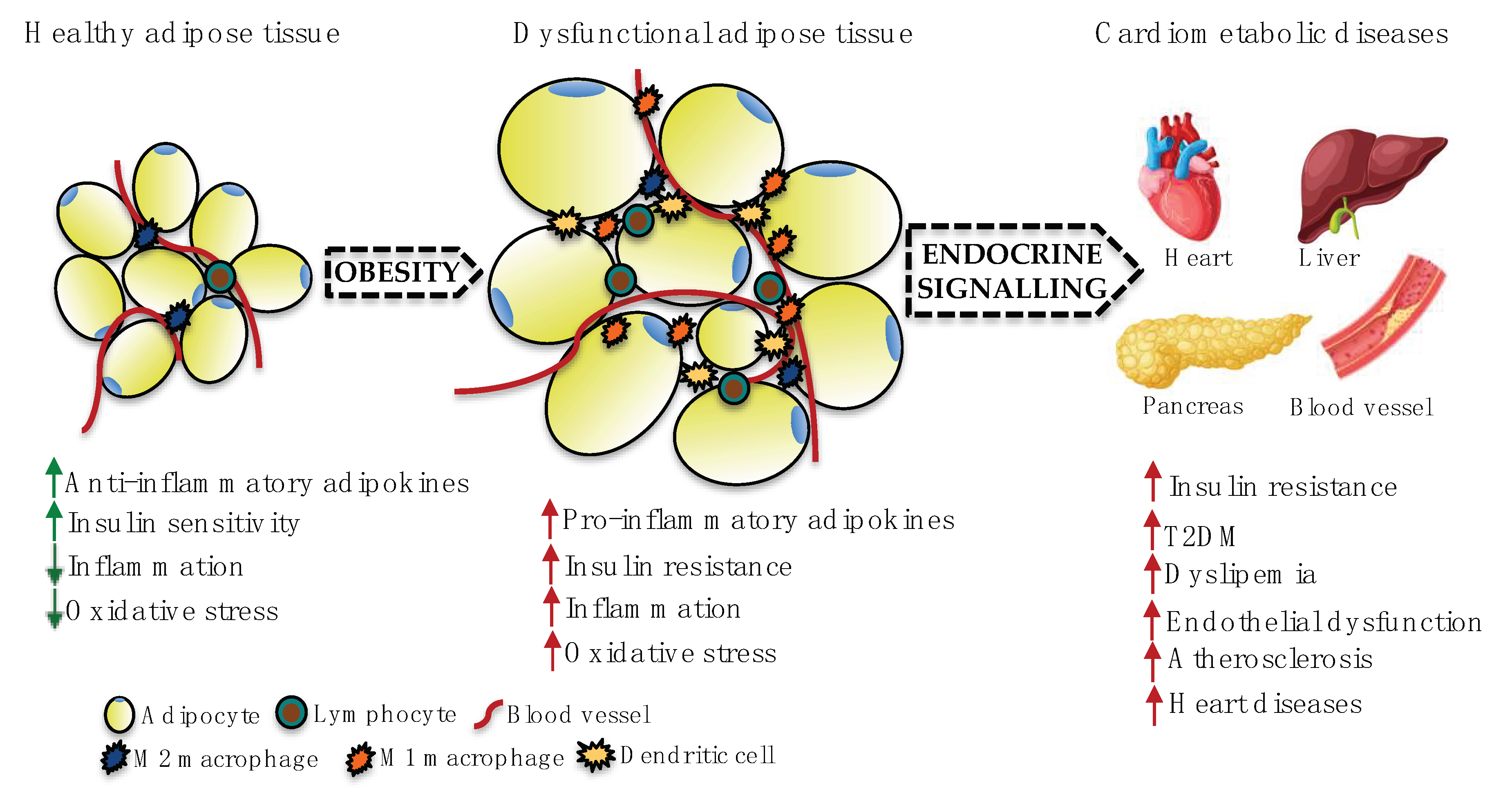
2. Adipose Tissue Dysfunction and CVDs
3. Role of Some Adipokines in Inflammatory Processes Associated with CVDs
3.1. Leptin
3.2. Chemerin
3.3. Resistin
3.4. Oncostatin M
3.5. Adiponectin
3.6. Nesfatin-1
3.7. Relaxin
3.8. Omentin
3.9. Meteorin-Like Hormone
3.10. Fibroblast Growth Factor 21
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AKT | AKT serine/threonine kinase 1 |
| AMI | Acute myocardial infarction |
| ApoE−/− | Apolipoprotein E double knockout |
| ARE | Antioxidant response elements |
| BMI | Body mass index |
| CAD | Coronary artery disease |
| CAP1 | Adenylyl cyclase-associated protein 1 |
| CaV1.2 | L-type Ca2+ channel |
| CCRL2 | C-C chemokine receptor-like 2 |
| cGMP | Cyclic guanosine monophosphate |
| CMDs | Cardiometabolic diseases |
| CMKLR1 | Chemokine-like receptor 1 |
| COX2 | Cyclooxygenase-2 |
| CRP | C-reactive protein |
| CVDs | Cardiovascular diseases |
| DCN | Decorin |
| EAT | Epicardial adipose tissue |
| ERK | Extracellular signal-regulated kinase |
| FGF-21 | Fibroblast growth factor-21 |
| eNOS | Endothelial nitric oxide synthase |
| FAS | Factor-associated suicide |
| GPR1 | G protein-coupled receptor 1 |
| GR | Glucocorticoid receptor |
| HbA1c | Glycated haemoglobin |
| HDL | High-density lipoprotein |
| HUVECs | Human umbilical vein endothelial cells |
| I/R | Ischemia/reperfusion |
| ICAM-1 | Intracellular molecular adhesion 1 |
| IFNγ | Interferon γ |
| IGF-1R | Insulin growth factor-1 receptor |
| IL | Interleukin |
| JAK | Janus kinase |
| JNK | C-Jun N-terminal kinases |
| KD | Kawasaki disease |
| LepR | Leptin receptor |
| LIF | Leukaemia inhibitory factor |
| LIFR | Leukaemia inhibitory factor receptor |
| LPS | Lipopolysaccharide |
| MAPK | Mitogen-activated protein kinase |
| MCP-1 | Monocyte chemoattractant protein-1 |
| Metrnl | Meteorin-like |
| MI | Myocardial infarction |
| MyD88 | Myeloid differentiation factor 88 |
| NFAT | Nuclear factor of activated T-cells |
| NF-κβ | Nuclear factor |
| NLRP3 | NOD-, LRR- and pyrin domain-containing protein 3 |
| NO | Nitric oxide |
| Nox | Nicotinamide adenine dinucleotide phosphate oxidase |
| Nrf2 | Nuclear factor erythroid 2-related factor 2 |
| NUCB2 | Nucleobindin 2 |
| OSM | Oncostatin M |
| OSMR | Oncostatin M receptor |
| oxLDL | Oxidised low-density lipoprotein |
| PBMCs | Peripheral blood mononuclear cells |
| PECAM | Platelet endothelial cell adhesion molecule |
| PI3K | Phosphoinositide 3-kinase |
| PGE2 | Prostaglandin E2 |
| PPARδ | Peroxisome proliferator-activated receptor δ |
| PVAT | Perivascular adipose tissue |
| RARRES2 | Retinoic acid receptor responder 2 |
| ROR-1 | Tyrosine kinase-like orphan receptor 1 |
| ROS | Reactive oxygen species |
| RXFP1 | Relaxin family peptide receptor 1 |
| SAT | Subcutaneous adipose tissue |
| STAT | Signal transducer and activator of transcription protein |
| T2DM | Type 2 diabetes mellitus |
| TGF-β | Transforming growth factor β |
| TIG2 | Tazarotene-induced gene 2 |
| TLR-4 | Toll-like receptor 4 |
| TNFα | Tumour necrosis factor α |
| TXNIP | Thioredoxin-interacting protein |
| VAT | Visceral adipose tissue |
| VCAM-1 | Vascular cell molecular adhesion 1 |
| VEGF | Vascular endothelial growth factor |
| VSMCs | Vascular smooth muscle cells |
| WAT | White adipose tissue |
References
- World Health Organization. Cardiovascular Diseases. Available online: https://www.who.int/health-topics/cardiovascular-diseases#tab=tab_1 (accessed on 22 July 2020).
- Vegiopoulos, A.; Rohm, M.; Herzig, S. Adipose tissue: Between the extremes. EMBO J. 2017, 36, 1999–2017. [Google Scholar] [CrossRef] [PubMed]
- Dodson, M.V.; Du, M.; Wang, S.; Bergen, W.G.; Fernyhough-Culver, M.; Basu, U.; Poulos, S.P.; Hausman, G.J. Adipose depots differ in cellularity, adipokines produced, gene expression, and cell systems. Adipocyte 2014, 3, 236–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koliaki, C.; Liatis, S.; Kokkinos, A. Obesity and cardiovascular disease: Revisiting an old relationship. Metabolism 2019, 92, 98–107. [Google Scholar] [CrossRef]
- Chait, A.; den Hartigh, L.J. Adipose Tissue Distribution, Inflammation and Its Metabolic Consequences, Including Diabetes and Cardiovascular Disease. Front. Cardiovasc. Med. 2020, 7, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Longo, M.; Zatterale, F.; Naderi, J.; Parrillo, L.; Formisano, P.; Raciti, G.A.; Beguinot, F.; Miele, C. Adipose Tissue Dysfunction as Determinant of Obesity-Associated Metabolic Complications. Int. J. Mol. Sci. 2019, 20, 2358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, C.H.; Lee, W.J.; Song, K.-H. Metabolically healthy obesity: A friend or foe? Kor. J. Intern. Med. 2017, 32, 611–621. [Google Scholar] [CrossRef] [Green Version]
- Guzik, T.J.; Skiba, D.S.; Touyz, R.M.; Harrison, D.G. The role of infiltrating immune cells in dysfunctional adipose tissue. Cardiovasc. Res. 2017, 113, 1009–1023. [Google Scholar] [CrossRef] [Green Version]
- Fuster, J.J.; Ouchi, N.; Gokce, N.; Walsh, K. Obesity-induced Changes in Adipose Tissue Microenvironment and Their Impact on Cardiovascular Disease. Circ. Res. 2016. [Google Scholar] [CrossRef] [Green Version]
- Unamuno, X.; Gómez-Ambrosi, J.; Rodríguez, A.; Becerril, S.; Frühbeck, G.; Catalán, V. Adipokine dysregulation and adipose tissue inflammation in human obesity. Eur. J. Clin. Invest. 2018, 48, e12997. [Google Scholar] [CrossRef] [Green Version]
- Bremer, A.A.; Jialal, I. Adipose tissue dysfunction in nascent metabolic syndrome. J. Obes. 2013, 2013, 393192. [Google Scholar] [CrossRef] [Green Version]
- Ferrara, D.; Montecucco, F.; Dallegri, F.; Carbone, F. Impact of different ectopic fat depots on cardiovascular and metabolic diseases. J. Cell. Physiol. 2019, 234, 21630–21641. [Google Scholar] [CrossRef]
- Rodríguez, A.; Becerril, S.; Hernández-Pardos, A.W.; Frühbeck, G. Adipose tissue depot differences in adipokines and effects on skeletal and cardiac muscle. Curr. Opin. Pharmacol. 2020, 52, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Landecho, M.F.; Tuero, C.; Valentí, V.; Bilbao, I.; de la Higuera, M.; Frühbeck, G. Relevance of Leptin and Other Adipokines in Obesity-Associated Cardiovascular Risk. Nutrients 2019, 11, 2664. [Google Scholar] [CrossRef] [Green Version]
- Neeland, I.J.; Poirier, P.; Després, J.-P. Cardiovascular and Metabolic Heterogeneity of Obesity. Circulation 2018, 137, 1391–1406. [Google Scholar] [CrossRef]
- McLaughlin, T.; Lamendola, C.; Liu, A.; Abbasi, F. Preferential Fat Deposition in Subcutaneous Versus Visceral Depots Is Associated with Insulin Sensitivity. J. Clin. Endocrinol. Metab. 2011, 96, E1756–E1760. [Google Scholar] [CrossRef] [Green Version]
- Fain, J.N.; Madan, A.K.; Hiler, M.L.; Cheema, P.; Bahouth, S.W. Comparison of the release of adipokines by adipose tissue, adipose tissue matrix, and adipocytes from visceral and subcutaneous abdominal adipose tissues of obese humans. Endocrinology 2004, 145, 2273–2282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González, N.; Moreno-Villegas, Z.; González-Bris, A.; Egido, J.; Lorenzo, Ó. Regulation of visceral and epicardial adipose tissue for preventing cardiovascular injuries associated to obesity and diabetes. Cardiovasc. Diabetol. 2017, 16, 44. [Google Scholar] [CrossRef]
- Packer, M. Epicardial Adipose Tissue May Mediate Deleterious Effects of Obesity and Inflammation on the Myocardium. J. Am. Coll. Cardiol. 2018, 71, 2360–2372. [Google Scholar] [CrossRef]
- Zhou, M.; Wang, H.; Chen, J.; Zhao, L. Epicardial adipose tissue and atrial fibrillation: Possible mechanisms, potential therapies, and future directions. Pacing Clin. Electrophysiol. 2020, 43, 133–145. [Google Scholar] [CrossRef] [PubMed]
- Le Jemtel, T.H.; Samson, R.; Ayinapudi, K.; Singh, T.; Oparil, S. Epicardial Adipose Tissue and Cardiovascular Disease. Curr. Hypertens. Rep. 2019, 21. [Google Scholar] [CrossRef] [PubMed]
- Patel, V.B.; Shah, S.; Verma, S.; Oudit, G.Y. Epicardial adipose tissue as a metabolic transducer: Role in heart failure and coronary artery disease. Heart Fail. Rev. 2017, 22, 889–902. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhao, L.; Liang, H.; Zhang, C.; Guan, L.; Li, M. A new measurement site for echocardiographic epicardial adipose tissue thickness and its value in predicting metabolic syndrome. Adv. Clin. Exp. Med. 2019, 28, 1403–1408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keresztesi, A.A.; Asofie, G.; Simion, M.A.; Jung, H. Correlation between epicardial adipose tissue thickness and the degree of coronary artery atherosclerosis. Turk. J. Med. Sci. 2018, 48, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Villasante Fricke, A.C.; Iacobellis, G. Epicardial Adipose Tissue: Clinical Biomarker of Cardio-Metabolic Risk. Int. J. Mol. Sci. 2019, 20, 5989. [Google Scholar] [CrossRef] [Green Version]
- Erkan, A.F.; Tanindi, A.; Kocaman, S.A.; Ugurlu, M.; Tore, H.F. Epicardial Adipose Tissue Thickness Is an Independent Predictor of Critical and Complex Coronary Artery Disease by Gensini and Syntax Scores. Texas Hear. Inst. J. 2016, 43, 29–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lima-Martínez, M.M.; Colmenares, L.; Campanelli, Y.; Paoli, M.; Rodney, M.; Santos, R.D.; Iacobellis, G. Epicardial adipose tissue thickness and type 2 diabetes risk according to the FINDRISC modified for Latin America. Clín. Investig. Arterioscler. 2019, 31, 15–22. [Google Scholar] [CrossRef]
- Hu, H.; Garcia-Barrio, M.; Jiang, Z.; Chen, Y.E.; Chang, L. Roles of Perivascular Adipose Tissue in Hypertension and Atherosclerosis. Antioxid. Redox Signal. 2020, ars.2020.8103. [Google Scholar] [CrossRef] [PubMed]
- Quesada, I.; Cejas, J.; García, R.; Cannizzo, B.; Redondo, A.; Castro, C. Vascular dysfunction elicited by a cross talk between periaortic adipose tissue and the vascular wall is reversed by pioglitazone. Cardiovasc. Ther. 2018, 36, e12322. [Google Scholar] [CrossRef]
- Qi, X.-Y.; Qu, S.-L.; Xiong, W.-H.; Rom, O.; Chang, L.; Jiang, Z.-S. Perivascular adipose tissue (PVAT) in atherosclerosis: A double-edged sword. Cardiovasc. Diabetol. 2018, 17, 134. [Google Scholar] [CrossRef] [Green Version]
- Spiroglou, S.G.; Kostopoulos, C.G.; Varakis, J.N.; Papadaki, H.H. Adipokines in periaortic and epicardial adipose tissue: Differential expression and relation to atherosclerosis. J. Atheroscler. Thromb. 2010, 17, 115–130. [Google Scholar] [CrossRef] [Green Version]
- Nosalski, R.; Guzik, T.J. Perivascular adipose tissue inflammation in vascular disease. Br. J. Pharmacol. 2017, 174, 3496–3513. [Google Scholar] [CrossRef] [Green Version]
- Anthony, S.R.; Guarnieri, A.R.; Gozdiff, A.; Helsley, R.N.; Phillip Owens, A.; Tranter, M. Mechanisms linking adipose tissue inflammation to cardiac hypertrophy and fibrosis. Clin. Sci. 2019, 133, 2329–2344. [Google Scholar] [CrossRef] [PubMed]
- Ahmadieh, S.; Kim, H.W.; Weintraub, N.L. Potential role of perivascular adipose tissue in modulating atherosclerosis. Clin. Sci. 2020, 134, 3–13. [Google Scholar] [CrossRef] [Green Version]
- Greenstein, A.S.; Khavandi, K.; Withers, S.B.; Sonoyama, K.; Clancy, O.; Jeziorska, M.; Laing, I.; Yates, A.P.; Pemberton, P.W.; Malik, R.A.; et al. Local Inflammation and Hypoxia Abolish the Protective Anticontractile Properties of Perivascular Fat in Obese Patients. Circulation 2009, 119, 1661–1670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeVallance, E.; Branyan, K.W.; Lemaster, K.C.; Anderson, R.; Marshall, K.L.; Olfert, I.M.; Smith, D.M.; Kelley, E.E.; Bryner, R.W.; Frisbee, J.C.; et al. Exercise training prevents the perivascular adipose tissue-induced aortic dysfunction with metabolic syndrome. Redox Biol. 2019, 26, 101285. [Google Scholar] [CrossRef]
- Han, F.; Hou, N.; Liu, Y.; Huang, N.; Pan, R.; Zhang, X.; Mao, E.; Sun, X. Liraglutide improves vascular dysfunction by regulating a cAMP-independent PKA-AMPK pathway in perivascular adipose tissue in obese mice. Biomed. Pharmacother. 2019, 120, 109537. [Google Scholar] [CrossRef]
- Dong, M.; Ren, J. What fans the fire: Insights into mechanisms of leptin in metabolic syndrome-associated heart diseases. Curr. Pharm. Des. 2014, 20, 652–658. [Google Scholar] [CrossRef] [PubMed]
- Considine, R.V.; Sinha, M.K.; Heiman, M.L.; Kriauciunas, A.; Stephens, T.W.; Nyce, M.R.; Ohannesian, J.P.; Marco, C.C.; McKee, L.J.; Bauer, T.L. Serum immunoreactive-leptin concentrations in normal-weight and obese humans. N. Engl. J. Med. 1996, 334, 292–295. [Google Scholar] [CrossRef]
- Martin, S.S.; Qasim, A.; Reilly, M.P. Leptin Resistance. J. Am. Coll. Cardiol. 2008, 52, 1201–1210. [Google Scholar] [CrossRef] [Green Version]
- Katsiki, N.; Mikhailidis, D.P.; Banach, M. Leptin, cardiovascular diseases and type 2 diabetes mellitus. Acta Pharmacol. Sin. 2018, 39, 1176–1188. [Google Scholar] [CrossRef] [Green Version]
- Gruzdeva, O.; Borodkina, D.; Uchasova, E.; Dyleva, Y.; Barbarash, O. Leptin resistance: Underlying mechanisms and diagnosis. Diabetes Metab. Syndr. Obes. Targets Ther. 2019, 12, 191–198. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.-W.; Lee, M.; Oh, K.-J. Adipose Tissue-Derived Signatures for Obesity and Type 2 Diabetes: Adipokines, Batokines and MicroRNAs. J. Clin. Med. 2019, 8, 854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gainsford, T.; Willson, T.A.; Metcalf, D.; Handman, E.; McFarlane, C.; Ng, A.; Nicola, N.A.; Alexander, W.S.; Hilton, D.J. Leptin can induce proliferation, differentiation, and functional activation of hemopoietic cells. Proc. Natl. Acad. Sci. USA 1996, 93, 14564–14568. [Google Scholar] [CrossRef] [Green Version]
- Feijóo-Bandín, S.; Portolés, M.; Roselló-Lletí, E.; Rivera, M.; González-Juanatey, J.R.R.; Lago, F. 20 years of leptin: Role of leptin in cardiomyocyte physiology and physiopathology. Life Sci. 2015, 140, 10–18. [Google Scholar] [CrossRef] [PubMed]
- An, H.S.; Lee, J.Y.; Choi, E.B.; Jeong, E.A.; Shin, H.J.; Kim, K.E.; Park, K.-A.; Jin, Z.; Lee, J.E.; Koh, J.S.; et al. Caloric restriction reverses left ventricular hypertrophy through the regulation of cardiac iron homeostasis in impaired leptin signaling mice. Sci. Rep. 2020, 10, 7176. [Google Scholar] [CrossRef]
- Ren, J. Leptin and hyperleptinemia—From friend to foe for cardiovascular function. J. Endocrinol. 2004, 181, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soderberg, S.; Ahren, B.; Jansson, J.-H.; Johnson, O.; Hallmans, G.; Asplund, K.; Olsson, T. Leptin is associated with increased risk of myocardial infarction. J. Intern. Med. 1999, 246, 409–418. [Google Scholar] [CrossRef]
- Kain, D.; Simon, A.J.; Greenberg, A.; Ben Zvi, D.; Gilburd, B.; Schneiderman, J. Cardiac leptin overexpression in the context of acute MI and reperfusion potentiates myocardial remodeling and left ventricular dysfunction. PLoS ONE 2018, 13, e0203902. [Google Scholar] [CrossRef]
- Demarchi, A.; Mazzucchelli, I.; Somaschini, A.; Cornara, S.; Dusi, V.; Mirizzi, A.M.; Ruffinazzi, M.; Crimi, G.; Ferlini, M.; Gnecchi, M.; et al. Leptin affects the inflammatory response after STEMI. Nutr. Metab. Cardiovasc. Dis. 2020, 30, 922–924. [Google Scholar] [CrossRef] [PubMed]
- Ekmen, N.; Helvaci, A.; Gunaldi, M.; Sasani, H.; Yildirmak, S.T. Leptin as an important link between obesity and cardiovascular risk factors in men with acute myocardial infarction. Ind. Heart J. 2016, 68, 132–137. [Google Scholar] [CrossRef] [Green Version]
- Khafaji, H.A.R.H.; Bener, A.B.; Rizk, N.M.; Al Suwaidi, J. Elevated serum leptin levels in patients with acute myocardial infarction; correlation with coronary angiographic and echocardiographic findings. BMC Res. Notes 2012, 5, 262. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Yang, P.; Li, T.; Gao, J.; Zhang, Y. Leptin Expression in Human Epicardial Adipose Tissue Is Associated with Local Coronary Atherosclerosis. Med. Sci. Monit. 2019, 25, 9913–9922. [Google Scholar] [CrossRef]
- Gruzdeva, O.; Uchasova, E.; Dyleva, Y.; Borodkina, D.; Akbasheva, O.; Antonova, L.; Matveeva, V.; Belik, E.; Ivanov, S.; Sotnikov, A.; et al. Adipocytes Directly Affect Coronary Artery Disease Pathogenesis via Induction of Adipokine and Cytokine Imbalances. Front. Immunol. 2019, 10, 2163. [Google Scholar] [CrossRef]
- Drosos, I.; Chalikias, G.; Pavlaki, M.; Kareli, D.; Epitropou, G.; Bougioukas, G.; Mikroulis, D.; Konstantinou, F.; Giatromanolaki, A.; Ritis, K.; et al. Differences between perivascular adipose tissue surrounding the heart and the internal mammary artery: Possible role for the leptin-inflammation-fibrosis-hypoxia axis. Clin. Res. Cardiol. 2016, 105, 887–900. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Hang, T.; Cheng, X.-M.; Li, D.-M.; Zhang, Q.-G.; Wang, L.-J.; Peng, Y.-P.; Gong, J.-B. Associations of C1q/TNF-Related Protein-9 Levels in Serum and Epicardial Adipose Tissue with Coronary Atherosclerosis in Humans. Biomed Res. Int. 2015, 2015, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puurunen, V.-P.; Kiviniemi, A.; Lepojärvi, S.; Piira, O.-P.; Hedberg, P.; Junttila, J.; Ukkola, O.; Huikuri, H. Leptin predicts short-term major adverse cardiac events in patients with coronary artery disease. Ann. Med. 2017, 49, 448–454. [Google Scholar] [CrossRef] [PubMed]
- Puurunen, V.P.; Lepojärvi, E.S.; Piira, O.P.; Hedberg, P.; Junttila, M.J.; Ukkola, O.; Huikuri, H.V. High plasma leptin levels are associated with impaired diastolic function in patients with coronary artery disease. Peptides 2016, 84, 17–21. [Google Scholar] [CrossRef]
- Farcaş, A.D.; Rusu, A.; Stoia, M.A.; Vida-Simiti, L.A. Plasma leptin, but not resistin, TNF-α and adiponectin, is associated with echocardiographic parameters of cardiac remodeling in patients with coronary artery disease. Cytokine 2018, 103, 46–49. [Google Scholar] [CrossRef]
- Chen, M.-C.; Wang, J.-H.; Lee, C.-J.; Hsu, B.-G. Association between hyperleptinemia and cardiovascular outcomes in patients with coronary artery disease. Ther. Clin. Risk Manag. 2018, 14, 1855–1862. [Google Scholar] [CrossRef] [Green Version]
- Du, Y.; Yang, S.-H.; Li, S.; Zhao, X.; Zhang, Y.; Sun, D.; Zhu, C.-G.; Wu, N.-Q.; Guo, Y.-L.; Xu, R.-X.; et al. Increased Serum Leptin Levels in New-Onset, Untreated Female Patients with Coronary Artery Disease and Positively Associated with Inflammatory Markers. Ann. Nutr. Metab. 2018, 72, 142–148. [Google Scholar] [CrossRef]
- Bickel, C.; Schnabel, R.B.; Zeller, T.; Lackner, K.J.; Rupprecht, H.J.; Blankenberg, S.; Sinning, C.; Westermann, D. Predictors of leptin concentration and association with cardiovascular risk in patients with coronary artery disease: Results from the Athero Gene study. Biomarkers 2017, 22, 210–218. [Google Scholar] [CrossRef]
- Knudson, J.D.; Dincer, Ü.D.; Zhang, C.; Swafford, A.N.; Koshida, R.; Picchi, A.; Focardi, M.; Dick, G.M.; Tune, J.D. Leptin receptors are expressed in coronary arteries, and hyperleptinemia causes significant coronary endothelial dysfunction. Am. J. Physiol. Circ. Physiol. 2005, 289, H48–H56. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Qiao, J.; Hu, J.; Fan, M.; Zhao, Y.; Su, H.; Wang, Z.; Yu, Q.; Ma, Q.; Li, Y.; et al. Leptin promotes endothelial dysfunction in chronic kidney disease by modulating the MTA1-mediated WNT/β-catenin pathway. Mol. Cell. Biochem. 2020. [Google Scholar] [CrossRef] [PubMed]
- Korda, M.; Kubant, R.; Patton, S.; Malinski, T. Leptin-induced endothelial dysfunction in obesity. Am. J. Physiol. Circ. Physiol. 2008, 295, H1514–H1521. [Google Scholar] [CrossRef] [Green Version]
- De Rosa, S.; Cirillo, P.; Pacileo, M.; Di Palma, V.; Paglia, A.; Chiariello, M. Leptin stimulated C-reactive protein production by human coronary artery endothelial cells. J. Vasc. Res. 2009, 46, 609–617. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Hoffmann, M.; Wolk, R.; Shamsuzzaman, A.S.M.; Somers, V.K. Leptin induces C-reactive protein expression in vascular endothelial cells. Arterioscler. Thromb. Vasc. Biol. 2007, 27, e302–e307. [Google Scholar] [CrossRef] [Green Version]
- Bouloumie, A.; Marumo, T.; Lafontan, M.; Busse, R. Leptin induces oxidative stress in human endothelial cells. FASEB J. 1999, 13, 1231–1238. [Google Scholar] [CrossRef]
- Cirillo, P.; Angri, V.; De Rosa, S.; Calì, G.; Petrillo, G.; Maresca, F.; D’Ascoli, G.-L.; Maietta, P.; Brevetti, L.; Chiariello, M. Pro-atherothrombotic effects of leptin in human coronary endothelial cells. Thromb. Haemost. 2010, 103, 1065–1075. [Google Scholar] [CrossRef] [Green Version]
- Rourke, J.L.; Dranse, H.J.; Sinal, C.J. Towards an integrative approach to understanding the role of chemerin in human health and disease. Obes. Rev. 2013, 14, 245–262. [Google Scholar] [CrossRef]
- Ernst, M.C.; Sinal, C.J. Chemerin: At the crossroads of inflammation and obesity. Trends Endocrinol. Metab. 2010, 21, 660–667. [Google Scholar] [CrossRef]
- Mattern, A.; Zellmann, T.; Beck-Sickinger, A.G. Processing, signaling, and physiological function of chemerin. IUBMB Life 2014, 66, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Du, X.-Y.; Leung, L.L.K. Proteolytic regulatory mechanism of chemerin bioactivity. Acta Biochim. Biophys. Sin. 2009, 41, 973–979. [Google Scholar] [CrossRef] [Green Version]
- Wittamer, V.; Franssen, J.-D.; Vulcano, M.; Mirjolet, J.-F.; Le Poul, E.; Migeotte, I.; Brézillon, S.; Tyldesley, R.; Blanpain, C.C.; Detheux, M.; et al. Specific Recruitment of Antigen-presenting Cells by Chemerin, a Novel Processed Ligand from Human Inflammatory Fluids. J. Exp. Med. 2003, 198, 977–985. [Google Scholar] [CrossRef] [PubMed]
- Zabel, B.A.; Nakae, S.; Zúñiga, L.; Kim, J.-Y.; Ohyama, T.; Alt, C.; Pan, J.; Suto, H.; Soler, D.; Allen, S.J.; et al. Mast cell-expressed orphan receptor CCRL2 binds chemerin and is required for optimal induction of IgE-mediated passive cutaneous anaphylaxis. J. Exp. Med. 2008, 205, 2207–2220. [Google Scholar] [CrossRef]
- Monnier, J.; Lewén, S.; O’Hara, E.; Huang, K.; Tu, H.; Butcher, E.C.; Zabel, B.A. Expression, regulation, and function of atypical chemerin receptor CCRL2 on endothelial cells. J. Immunol. 2012, 189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monnier, J.; Zabel, B.; Butcher, E. Regulation of the Atypical Chemerin Receptor, CCRL2, on Activated Brain Endothelial Cells. Clin. Immunol. 2010, 135, S63. [Google Scholar] [CrossRef]
- De Henau, O.; Degroot, G.-N.; Imbault, V.; Robert, V.; De Poorter, C.; Mcheik, S.; Galés, C.; Parmentier, M.; Springael, J.-Y. Signaling Properties of Chemerin Receptors CMKLR1, GPR1 and CCRL2. PLoS ONE 2016, 11, e0164179. [Google Scholar] [CrossRef]
- Kennedy, A.J.; Davenport, A.P. International Union of Basic and Clinical Pharmacology CIII: Chemerin Receptors CMKLR1 (Chemerin 1) and GPR1 (Chemerin 2) Nomenclature, Pharmacology, and Function. Pharmacol. Rev. 2018, 70, 174–196. [Google Scholar] [CrossRef] [Green Version]
- Nagpal, S.; Patel, S.; Jacobe, H.; DiSepio, D.; Ghosn, C.; Malhotra, M.; Teng, M.; Duvic, M.; Chandraratna, R.A. Tazarotene-induced gene 2 (TIG2), a novel retinoid-responsive gene in skin. J. Invest. Dermatol. 1997, 109, 91–95. [Google Scholar] [CrossRef] [Green Version]
- Özcan, E.; Saygun, N.I.; Ilıkçı, R.; Karslıoğlu, Y.; Muşabak, U.; Yeşillik, S. Evaluation of chemerin and its receptors, ChemR23 and CCRL2, in gingival tissues with healthy and periodontitis. Odontology 2018, 106. [Google Scholar] [CrossRef]
- Chamberland, J.P.; Berman, R.L.; Aronis, K.N.; Mantzoros, C.S. Chemerin is expressed mainly in pancreas and liver, is regulated by energy deprivation, and lacks day/night variation in humans. Eur. J. Endocrinol. 2013, 169, 453–462. [Google Scholar] [CrossRef] [Green Version]
- Kostopoulos, C.G.; Spiroglou, S.G.; Varakis, J.N.; Apostolakis, E.; Papadaki, H.H. Chemerin and CMKLR1 expression in human arteries and periadventitial fat: A possible role for local chemerin in atherosclerosis? BMC Cardiovasc. Disord. 2014, 14, 56. [Google Scholar] [CrossRef]
- Berg, V.; Sveinbjörnsson, B.; Bendiksen, S.; Brox, J.; Meknas, K.; Figenschau, Y. Human articular chondrocytes express ChemR23 and chemerin; ChemR23 promotes inflammatory signalling upon binding the ligand chemerin(21-157). Arthritis Res. Ther. 2010, 12, R228. [Google Scholar] [CrossRef] [Green Version]
- Bongrani, A.; Mellouk, N.; Rame, C.; Cornuau, M.; Guérif, F.; Froment, P.; Dupont, J. Ovarian Expression of Adipokines in Polycystic Ovary Syndrome: A Role for Chemerin, Omentin, and Apelin in Follicular Growth Arrest and Ovulatory Dysfunction? Int. J. Mol. Sci. 2019, 20, 3778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, K.; Yoshizawa, H.; Seki, T.; Shirai, R.; Yamashita, T.; Okano, T.; Shibata, K.; Wakamatsu, M.J.; Mori, Y.; Morita, T.; et al. Chemerin-9, a potent agonist of chemerin receptor (ChemR23), prevents atherogenesis. Clin. Sci. 2019, 133, 1779–1796. [Google Scholar] [CrossRef]
- Goralski, K.B.; McCarthy, T.C.; Hanniman, E.A.; Zabel, B.A.; Butcher, E.C.; Parlee, S.D.; Muruganandan, S.; Sinal, C.J. Chemerin, a novel adipokine that regulates adipogenesis and adipocyte metabolism. J. Biol. Chem. 2007, 282, 28175–28188. [Google Scholar] [CrossRef] [Green Version]
- Alfadda, A.A.; Sallam, R.M.; Chishti, M.A.; Moustafa, A.S.; Fatma, S.; Alomaim, W.S.; Al-Naami, M.Y.; Bassas, A.F.; Chrousos, G.P.; Jo, H. Differential patterns of serum concentration and adipose tissue expression of chemerin in obesity: Adipose depot specificity and gender dimorphism. Mol. Cells 2012, 33, 591–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-García, M.Á.; Montes-Nieto, R.; Fernández-Durán, E.; Insenser, M.; Luque-Ramírez, M.; Escobar-Morreale, H.F. Evidence for Masculinization of Adipokine Gene Expression in Visceral and Subcutaneous Adipose Tissue of Obese Women With Polycystic Ovary Syndrome (PCOS). J. Clin. Endocrinol. Metab. 2013, 98, E388–E396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zylla, S.; Pietzner, M.; Kühn, J.-P.; Völzke, H.; Dörr, M.; Nauck, M.; Friedrich, N. Serum chemerin is associated with inflammatory and metabolic parameters-results of a population-based study. Obesity 2017, 25, 468–475. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Mi, S.; Zhang, F.; Gong, F.; Lai, Y.; Gao, F.; Zhang, X.; Wang, L.; Tao, H. Association of chemerin mRNA expression in human epicardial adipose tissue with coronary atherosclerosis. Cardiovasc. Diabetol. 2011, 10, 87. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.; Chen, Y.; Chen, S.; Wu, X.; Nong, W. Correlation between adiponectin, chemerin, vascular endothelial growth factor and epicardial fat volume in patients with coronary artery disease. Exp. Ther. Med. 2019, 19, 1095–1102. [Google Scholar] [CrossRef] [Green Version]
- Goralski, K.B.; Sinal, C.J. Elucidation of chemerin and chemokine-like receptor-1 function in adipocytes by adenoviral-mediated shRNA knockdown of gene expression. Methods Enzymol. 2009, 460. [Google Scholar] [CrossRef]
- Takahashi, M.; Takahashi, Y.; Takahashi, K.; Zolotaryov, F.N.; Hong, K.S.; Kitazawa, R.; Iida, K.; Okimura, Y.; Kaji, H.; Kitazawa, S.; et al. Chemerin enhances insulin signaling and potentiates insulin-stimulated glucose uptake in 3T3-L1 adipocytes. FEBS Lett. 2008, 582, 573–578. [Google Scholar] [CrossRef] [Green Version]
- Kralisch, S.; Weise, S.; Sommer, G.; Lipfert, J.; Lossner, U.; Bluher, M.; Stumvoll, M.; Fasshauer, M. Interleukin-1ß induces the novel adipokine chemerin in adipocytes in vitro. Regul. Pept. 2009, 154, 102–106. [Google Scholar] [CrossRef] [PubMed]
- Sell, H.; Laurencikiene, J.; Taube, A.; Eckardt, K.; Cramer, A.; Horrighs, A.; Arner, P.; Eckel, J. Chemerin is a Novel Adipocyte-Derived Factor Inducing Insulin Resistance in Primary Human Skeletal Muscle Cells. Diabetes 2009, 58, 2731–2740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, R.; Liu, S.; Guo, B.; Chang, L.; Li, Y. Chemerin Induces Insulin Resistance in Rat Cardiomyocytes in Part through the ERK1/2 Signaling Pathway. Pharmacology 2014, 94, 259–264. [Google Scholar] [CrossRef]
- Ernst, M.C.; Issa, M.; Goralski, K.B.; Sinal, C.J. Chemerin Exacerbates Glucose Intolerance in Mouse Models of Obesity and Diabetes. Endocrinology 2010, 151, 1998–2007. [Google Scholar] [CrossRef]
- Takahashi, M.; Okimura, Y.; Iguchi, G.; Nishizawa, H.; Yamamoto, M.; Suda, K.; Kitazawa, R.; Fujimoto, W.; Takahashi, K.; Zolotaryov, F.N.; et al. Chemerin regulates β-cell function in mice. Sci. Rep. 2011, 1, 123. [Google Scholar] [CrossRef]
- Ernst, M.C.; Haidl, I.D.; Zúñiga, L.A.; Dranse, H.J.; Rourke, J.L.; Zabel, B.A.; Butcher, E.C.; Sinal, C.J. Disruption of the Chemokine-Like Receptor-1 (CMKLR1) Gene Is Associated with Reduced Adiposity and Glucose Intolerance. Endocrinology 2012, 153, 672–682. [Google Scholar] [CrossRef] [Green Version]
- El-Deeb, T.S.; Bakkar, S.M.; Eltoony, L.; Zakhary, M.M.; Kamel, A.A.; Nafee, A.M.; Hetta, H.F. The adipokine Chemerin and Fetuin-A Serum Levels in Type 2 Diabetes Mellitus: Relation to Obesity and Inflammatory Markers. Egypt. J. Immunol. 2018, 25, 191–202. [Google Scholar]
- Ba, H.-J.; Xu, L.-L.; Qin, Y.-Z.; Chen, H.-S. Serum Chemerin Levels Correlate with Determinants of Metabolic Syndrome in Obese Children and Adolescents. Clin. Med. Insights. Pediatr. 2019, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, M.; Zhou, X.; Xu, J.; Yang, B.; Yu, J.; Gong, Q.; Zhang, X.; Sun, X.; Zhang, Q.; Xia, J.; et al. Association of serum chemerin and inflammatory factors with type 2 diabetes macroangiopathy and waist-to-stature ratio. Bosn. J. Basic Med. Sci. 2019, 19, 328–335. [Google Scholar] [CrossRef] [PubMed]
- Skuratovskaia, D.; Zatolokin, P.; Vulf, M.; Mazunin, I.; Litvinova, L. Interrelation of chemerin and TNF-α with mtDNA copy number in adipose tissues and blood cells in obese patients with and without type 2 diabetes. BMC Med. Genomics 2019, 12, 40. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Lin, X.; Wang, X. Decrease in serum chemerin through aerobic exercise plus dieting and its association with mitigation of cardio-metabolic risk in obese female adolescents. J. Pediatr. Endocrinol. Metab. 2018, 31, 127–135. [Google Scholar] [CrossRef]
- Chakaroun, R.; Raschpichler, M.; Klöting, N.; Oberbach, A.; Flehmig, G.; Kern, M.; Schön, M.R.; Shang, E.; Lohmann, T.; Dreßler, M.; et al. Effects of weight loss and exercise on chemerin serum concentrations and adipose tissue expression in human obesity. Metabolism 2012, 61, 706–714. [Google Scholar] [CrossRef] [Green Version]
- Sell, H.; Divoux, A.; Poitou, C.; Basdevant, A.; Bouillot, J.-L.; Bedossa, P.; Tordjman, J.; Eckel, J.; Clément, K. Chemerin Correlates with Markers for Fatty Liver in Morbidly Obese Patients and Strongly Decreases after Weight Loss Induced by Bariatric Surgery. J. Clin. Endocrinol. Metab. 2010, 95, 2892–2896. [Google Scholar] [CrossRef] [Green Version]
- Kolahdouzi, S.; Baghadam, M.; Kani-Golzar, F.A.; Saeidi, A.; Jabbour, G.; Ayadi, A.; De Sousa, M.; Zouita, A.; Abderrahmane, A.B.; Zouhal, H. Progressive circuit resistance training improves inflammatory biomarkers and insulin resistance in obese men. Physiol. Behav. 2019, 205, 15–21. [Google Scholar] [CrossRef]
- Catalán, V.; Gómez-Ambrosi, J.; Rodríguez, A.; Ramírez, B.; Rotellar, F.; Valentí, V.; Silva, C.; Gil, M.J.; Salvador, J.; Frühbeck, G. Increased levels of chemerin and its receptor, chemokine-like receptor-1, in obesity are related to inflammation: Tumor necrosis factor-α stimulates mRNA levels of chemerin in visceral adipocytes from obese patients. Surg. Obes. Relat. Dis. 2013, 9, 306–314. [Google Scholar] [CrossRef] [PubMed]
- Roman, A.A.; Parlee, S.D.; Sinal, C.J. Chemerin: A potential endocrine link between obesity and type 2 diabetes. Endocrine 2012, 42, 243–251. [Google Scholar] [CrossRef]
- Ghosh, A.R.; Bhattacharya, R.; Bhattacharya, S.; Nargis, T.; Rahaman, O.; Duttagupta, P.; Raychaudhuri, D.; Liu, C.S.C.; Roy, S.; Ghosh, P.; et al. Adipose Recruitment and Activation of Plasmacytoid Dendritic Cells Fuel Metaflammation. Diabetes 2016, 65, 3440–3452. [Google Scholar] [CrossRef] [Green Version]
- Cash, J.L.; Hart, R.; Russ, A.; Dixon, J.P.C.; Colledge, W.H.; Doran, J.; Hendrick, A.G.; Carlton, M.B.L.; Greaves, D.R. Synthetic chemerin-derived peptides suppress inflammation through ChemR23. J. Exp. Med. 2008, 205, 767–775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamawaki, H.; Kameshima, S.; Usui, T.; Okada, M.; Hara, Y. A novel adipocytokine, chemerin exerts anti-inflammatory roles in human vascular endothelial cells. Biochem. Biophys. Res. Commun. 2012, 423, 152–157. [Google Scholar] [CrossRef] [PubMed]
- Laranjeira, S.; Regan-Komito, D.; Iqbal, A.J.; Greaves, D.R.; Payne, S.J.; Orlowski, P. A model for the optimization of anti-inflammatory treatment with chemerin. Interface Focus 2018, 8, 20170007. [Google Scholar] [CrossRef] [Green Version]
- Valcamonica, E.; Chighizola, C.B.; Comi, D.; De Lucia, O.; Pisoni, L.; Murgo, A.; Salvi, V.; Sozzani, S.; Meroni, P.L. Levels of chemerin and interleukin 8 in the synovial fluid of patients with inflammatory arthritides and osteoarthritis. Clin. Exp. Rheumatol. 2014, 32, 243–250. [Google Scholar] [PubMed]
- Patnaik, K.; Pradeep, A.R.; Nagpal, K.; Karvekar, S.; Singh, P.; Raju, A. Human chemerin correlation in gingival crevicular fluid and tear fluid as markers of inflammation in chronic periodontitis and type-2 diabetes mellitus. J. Investig. Clin. Dent. 2017, 8, e12181. [Google Scholar] [CrossRef]
- Calvet, J.; Orellana, C.; Gratacós, J.; Berenguer-Llergo, A.; Caixàs, A.; Chillarón, J.J.; Pedro-Botet, J.; García-Manrique, M.; Navarro, N.; Larrosa, M. Synovial fluid adipokines are associated with clinical severity in knee osteoarthritis: A cross-sectional study in female patients with joint effusion. Arthritis Res. Ther. 2016, 18, 207. [Google Scholar] [CrossRef] [Green Version]
- Huang, K.; Du, G.; Li, L.; Liang, H.; Zhang, B. Association of chemerin levels in synovial fluid with the severity of knee osteoarthritis. Biomarkers 2012, 17, 16–20. [Google Scholar] [CrossRef] [PubMed]
- Zabel, B.A.; Ohyama, T.; Zuniga, L.; Kim, J.-Y.; Johnston, B.; Allen, S.J.; Guido, D.G.; Handel, T.M.; Butcher, E.C. Chemokine-like receptor 1 expression by macrophages in vivo: Regulation by TGF-β and TLR ligands. Exp. Hematol. 2006, 34, 1106–1114. [Google Scholar] [CrossRef]
- Parolini, S.; Santoro, A.; Marcenaro, E.; Luini, W.; Massardi, L.; Facchetti, F.; Communi, D.; Parmentier, M.; Majorana, A.; Sironi, M.; et al. The role of chemerin in the colocalization of NK and dendritic cell subsets into inflamed tissues. Blood 2007, 109, 3625–3632. [Google Scholar] [CrossRef]
- Zabel, B.A.; Silverio, A.M.; Butcher, E.C. Chemokine-Like Receptor 1 Expression and Chemerin-Directed Chemotaxis Distinguish Plasmacytoid from Myeloid Dendritic Cells in Human Blood. J. Immunol. 2005, 174, 244–251. [Google Scholar] [CrossRef] [Green Version]
- Nishida, K.; Otsu, K. Inflammation and metabolic cardiomyopathy. Cardiovasc. Res. 2017, 113, 389–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hart, R.; Greaves, D.R. Chemerin Contributes to Inflammation by Promoting Macrophage Adhesion to VCAM-1 and Fibronectin through Clustering of VLA-4 and VLA-5. J. Immunol. 2010, 185, 3728–3739. [Google Scholar] [CrossRef] [Green Version]
- Lehrke, M.; Becker, A.; Greif, M.; Stark, R.; Laubender, R.P.; von Ziegler, F.; Lebherz, C.; Tittus, J.; Reiser, M.; Becker, C.; et al. Chemerin is associated with markers of inflammation and components of the metabolic syndrome but does not predict coronary atherosclerosis. Eur. J. Endocrinol. 2009, 161, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Gu, P.; Jiang, W.; Lu, B.; Shi, Z. Chemerin is associated with inflammatory markers and metabolic syndrome phenotypes in hypertension patients. Clin. Exp. Hypertens. 2014, 36, 326–332. [Google Scholar] [CrossRef]
- Sawicka, K.; Michalska-Jakubus, M.; Potembska, E.; Kowal, M.; Pietrzak, A.; Krasowska, D. Visfatin and chemerin levels correspond with inflammation and might reflect the bridge between metabolism, inflammation and fibrosis in patients with systemic sclerosis. Adv. Dermatol. Allergol. 2019, 36, 551–565. [Google Scholar] [CrossRef] [Green Version]
- Kammerer, A.; Staab, H.; Herberg, M.; Kerner, C.; Klöting, N.; Aust, G. Increased circulating chemerin in patients with advanced carotid stenosis. BMC Cardiovasc. Disord. 2018, 18, 65. [Google Scholar] [CrossRef] [Green Version]
- Weigert, J.; Obermeier, F.; Neumeier, M.; Wanninger, J.; Filarsky, M.; Bauer, S.; Aslanidis, C.; Rogler, G.; Ott, C.; Schäffler, A.; et al. Circulating levels of chemerin and adiponectin are higher in ulcerative colitis and chemerin is elevated in Crohn’s disease. Inflamm. Bowel Dis. 2010, 16, 630–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiaotao, L.; Xiaoxia, Z.; Yue, X.; Liye, W. Serum chemerin levels are associated with the presence and extent of coronary artery disease. Coron. Artery Dis. 2012, 23, 412–416. [Google Scholar] [CrossRef] [PubMed]
- Dong, B.; Ji, W.; Zhang, Y. Elevated Serum Chemerin Levels are Associated with the Presence of Coronary Artery Disease in Patients with Metabolic Syndrome. Intern. Med. 2011, 50, 1093–1097. [Google Scholar] [CrossRef] [Green Version]
- Aksan, G.; İnci, S.; Nar, G.; Soylu, K.; Gedikli, Ö.; Yüksel, S.; Özdemir, M.; Nar, R.; Meriç, M.; Şahin, M. Association of serum chemerin levels with the severity of coronary artery disease in patients with metabolic syndrome. Int. J. Clin. Exp. Med. 2014, 7, 5461–5468. [Google Scholar] [PubMed]
- Motawi, T.M.K.; Mahdy, S.G.; El-Sawalhi, M.M.; Ali, E.N.; El-Telbany, R.F.A. Serum levels of chemerin, apelin, vaspin, and omentin-1 in obese type 2 diabetic Egyptian patients with coronary artery stenosis. Can. J. Physiol. Pharmacol. 2018, 96, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Mussbacher, M.; Salzmann, M.; Brostjan, C.; Hoesel, B.; Schoergenhofer, C.; Datler, H.; Hohensinner, P.; Basílio, J.; Petzelbauer, P.; Assinger, A.; et al. Cell Type-Specific Roles of NF-κB Linking Inflammation and Thrombosis. Front. Immunol. 2019, 10, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kojok, K.; El-Kadiry, A.E.-H.; Merhi, Y. Role of NF-κB in Platelet Function. Int. J. Mol. Sci. 2019, 20, 4185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dimitriadis, G.K.; Kaur, J.; Adya, R.; Miras, A.D.; Mattu, H.S.; Hattersley, J.G.; Kaltsas, G.; Tan, B.K.; Randeva, H.S. Chemerin induces endothelial cell inflammation: Activation of nuclear factor-kappa beta and monocyte-endothelial adhesion. Oncotarget 2018, 9, 16678–16690. [Google Scholar] [CrossRef] [Green Version]
- Neves, K.B.; Nguyen Dinh Cat, A.; Lopes, R.A.M.; Rios, F.J.; Anagnostopoulou, A.; Lobato, N.S.; de Oliveira, A.M.; Tostes, R.C.; Montezano, A.C.; Touyz, R.M. Chemerin Regulates Crosstalk Between Adipocytes and Vascular Cells Through Nox. Hypertension 2015, 66, 657–666. [Google Scholar] [CrossRef] [PubMed]
- Shang, J.; Wang, L.; Zhang, Y.; Zhang, S.; Ning, L.; Zhao, J.; Cheng, G.; Liu, D.; Xiao, J.; Zhao, Z. Chemerin/ChemR23 axis promotes inflammation of glomerular endothelial cells in diabetic nephropathy. J. Cell. Mol. Med. 2019, 23, 3417–3428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Xiong, W.; Luo, Y.; Chen, H.; He, Y.; Cao, Y.; Dong, S. Adipokine Chemerin Stimulates Progression of Atherosclerosis in ApoE −/− Mice. Biomed Res. Int. 2019, 2019, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Er, L.; Hsu, L.-A.; Juang, J.-M.; Chiang, F.-T.; Teng, M.-S.; Tzeng, I.-S.; Wu, S.; Lin, J.-F.; Ko, Y.-L. Circulating Chemerin Levels, but not the RARRES2 Polymorphisms, Predict the Long-Term Outcome of Angiographically Confirmed Coronary Artery Disease. Int. J. Mol. Sci. 2019, 20, 1174. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Xiao, M.; Zhang, L.; Zhao, Y.; Yang, Q. Association of serum chemerin concentrations with the presence of atrial fibrillation. Ann. Clin. Biochem. 2017, 54, 342–347. [Google Scholar] [CrossRef] [Green Version]
- Zhao, D.; Bi, G.; Feng, J.; Huang, R.; Chen, X. Association of Serum Chemerin Levels with Acute Ischemic Stroke and Carotid Artery Atherosclerosis in a Chinese Population. Med. Sci. Monit. 2015, 21, 3121–3128. [Google Scholar] [CrossRef] [Green Version]
- Gu, P.; Cheng, M.; Hui, X.; Lu, B.; Jiang, W.; Shi, Z. Elevating circulation chemerin level is associated with endothelial dysfunction and early atherosclerotic changes in essential hypertensive patients. J. Hypertens. 2015, 33, 1624–1632. [Google Scholar] [CrossRef] [PubMed]
- Aydin, K.; Canpolat, U.; Akin, S.; Dural, M.; Karakaya, J.; Aytemir, K.; Ozer, N.; Gurlek, A. Chemerin is not associated with subclinical atherosclerosis markers in prediabetes and diabetes. Anatol. J. Cardiol. 2015, 16, 749–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Penas, D.; Feijóo-Bandín, S.; García-Rúa, V.; Mosquera-Leal, A.; Durán, D.; Varela, A.; Portolés, M.; Roselló-Lletí, E.; Rivera, M.; Diéguez, C.; et al. The adipokine chemerin induces apoptosis in cardiomyocytes. Cell. Physiol. Biochem. 2015, 37, 176–192. [Google Scholar] [CrossRef] [PubMed]
- Kutlay, Ö.; Kaygısız, Z.; Kaygısız, B. The Effect of Chemerin on Cardiac Parameters and Gene Expressions in Isolated Perfused Rat Heart. Balkan Med. J. 2018. [Google Scholar] [CrossRef] [PubMed]
- Hanthazi, A.; Jespers, P.; Vegh, G.; Degroot, G.-N.; Springael, J.-Y.; Lybaert, P.; Dewachter, L.; Mc Entee, K. Chemerin influences endothelin- and serotonin-induced pulmonary artery vasoconstriction in rats. Life Sci. 2019, 231, 116580. [Google Scholar] [CrossRef] [PubMed]
- Ferland, D.J.; Darios, E.S.; Neubig, R.R.; Sjögren, B.; Truong, N.; Torres, R.; Dexheimer, T.S.; Thompson, J.M.; Watts, S.W. Chemerin-induced arterial contraction is Gi- and calcium-dependent. Vascul. Pharmacol. 2017, 88, 30–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darios, E.S.; Winner, B.M.; Charvat, T.; Krasinksi, A.; Punna, S.; Watts, S.W. The adipokine chemerin amplifies electrical field-stimulated contraction in the isolated rat superior mesenteric artery. Am. J. Physiol. Circ. Physiol. 2016, 311, H498–H507. [Google Scholar] [CrossRef]
- Kennedy, A.J.; Yang, P.; Read, C.; Kuc, R.E.; Yang, L.; Taylor, E.J.A.; Taylor, C.W.; Maguire, J.J.; Davenport, A.P. Chemerin Elicits Potent Constrictor Actions via Chemokine-Like Receptor 1 (CMKLR1), not G-Protein-Coupled Receptor 1 (GPR1), in Human and Rat Vasculature. J. Am. Heart Assoc. 2016, 5. [Google Scholar] [CrossRef] [Green Version]
- Lobato, N.S.; Neves, K.B.; Filgueira, F.P.; Fortes, Z.B.; Carvalho, M.H.C.; Webb, R.C.; Oliveira, A.M.; Tostes, R.C. The adipokine chemerin augments vascular reactivity to contractile stimuli via activation of the MEK-ERK1/2 pathway. Life Sci. 2012, 91, 600–606. [Google Scholar] [CrossRef]
- Kunimoto, H.; Kazama, K.; Takai, M.; Oda, M.; Okada, M.; Yamawaki, H. Chemerin promotes the proliferation and migration of vascular smooth muscle and increases mouse blood pressure. Am. J. Physiol. Circ. Physiol. 2015, 309, H1017–H1028. [Google Scholar] [CrossRef] [Green Version]
- Weng, C.; Shen, Z.; Li, X.; Jiang, W.; Peng, L.; Yuan, H.; Yang, K.; Wang, J. Effects of chemerin/CMKLR1 in obesity-induced hypertension and potential mechanism. Am. J. Transl. Res. 2017, 9, 3096–3104. [Google Scholar]
- Ferland, D.J.; Flood, E.D.; Garver, H.; Yeh, S.T.; Riney, S.; Mullick, A.E.; Fink, G.D.; Watts, S.W. Different blood pressure responses in hypertensive rats following chemerin mRNA inhibition in dietary high fat compared to dietary high-salt conditions. Physiol. Genomics 2019, 51, 553–561. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Yang, G.; Dong, J.; Liu, Y.; Zong, H.; Liu, H.; Boden, G.; Li, L. Elevated plasma levels of chemerin in newly diagnosed type 2 diabetes mellitus with hypertension. J. Investig. Med. 2010, 58, 883–886. [Google Scholar] [CrossRef]
- Meric, M.; Soylu, K.; Avci, B.; Yuksel, S.; Gulel, O.; Yenercag, M.; Coksevim, M.; Uzun, A. Evaluation of plasma chemerin levels in patients with non-dipper blood pressure patterns. Med. Sci. Monit. 2014, 20, 698–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watts, S.W.; Dorrance, A.M.; Penfold, M.E.; Rourke, J.L.; Sinal, C.J.; Seitz, B.; Sullivan, T.J.; Charvat, T.T.; Thompson, J.M.; Burnett, R.; et al. Chemerin connects fat to arterial contraction. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 1320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steppan, C.M.; Brown, E.J.; Wright, C.M.; Bhat, S.; Banerjee, R.R.; Dai, C.Y.; Enders, G.H.; Silberg, D.G.; Wen, X.; Wu, G.D.; et al. A family of tissue-specific resistin-like molecules. Proc. Natl. Acad. Sci. USA 2001, 98, 502–506. [Google Scholar] [CrossRef] [Green Version]
- Steppan, C.M.; Bailey, S.T.; Bhat, S.; Brown, E.J.; Banerjee, R.R.; Wright, C.M.; Patel, H.R.; Ahima, R.S.; Lazar, M.A. The hormone resistin links obesity to diabetes. Nature 2001, 409, 307–312. [Google Scholar] [CrossRef]
- Moon, B.; Kwan, J.J.-M.; Duddy, N.; Sweeney, G.; Begum, N. Resistin inhibits glucose uptake in L6 cells independently of changes in insulin signaling and GLUT4 translocation. Am. J. Physiol. Metab. 2003, 285, E106–E115. [Google Scholar] [CrossRef]
- Rajala, M.W.; Obici, S.; Scherer, P.E.; Rossetti, L. Adipose-derived resistin and gut-derived resistin-like molecule–β selectively impair insulin action on glucose production. J. Clin. Invest. 2003, 111, 225–230. [Google Scholar] [CrossRef] [Green Version]
- Acquarone, E.; Monacelli, F.; Borghi, R.; Nencioni, A.; Odetti, P. Resistin: A reappraisal. Mech. Ageing Dev. 2019, 178, 46–63. [Google Scholar] [CrossRef] [PubMed]
- Bokarewa, M.; Nagaev, I.; Dahlberg, L.; Smith, U.; Tarkowski, A. Resistin, an Adipokine with Potent Proinflammatory Properties. J. Immunol. 2005. [Google Scholar] [CrossRef] [PubMed]
- Lehrke, M.; Reilly, M.P.; Millington, S.C.; Iqbal, N.; Rader, D.J.; Lazar, M.A. An Inflammatory Cascade Leading to Hyperresistinemia in Humans. PLoS Med. 2004, 1, e45. [Google Scholar] [CrossRef] [PubMed]
- Plutzky, J. Inflammatory pathways in atherosclerosis and acute coronary syndromes. Am. J. Cardiol. 2001, 88, 10–15. [Google Scholar] [CrossRef]
- Chen, C.; Jiang, J.; Lü, J.-M.; Chai, H.; Wang, X.; Lin, P.H.; Yao, Q. Resistin decreases expression of endothelial nitric oxide synthase through oxidative stress in human coronary artery endothelial cells. Am. J. Physiol. Circ. Physiol. 2010, 299, H193–H201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, H.S.; Park, K.H.; Cho, Y.M.; Chung, S.S.; Cho, H.J.; Cho, S.Y.; Kim, S.J.; Kim, S.Y.; Lee, H.K.; Park, K.S. Resistin is secreted from macrophages in atheromas and promotes atherosclerosis. Cardiovasc. Res. 2006, 69, 76–85. [Google Scholar] [CrossRef]
- Langheim, S.; Dreas, L.; Veschini, L.; Maisano, F.; Foglieni, C.; Ferrarello, S.; Sinagra, G.; Zingone, B.; Alfieri, O.; Ferrero, E.; et al. Increased expression and secretion of resistin in epicardial adipose tissue of patients with acute coronary syndrome. Am. J. Physiol. Circ. Physiol. 2010, 298, H746–H753. [Google Scholar] [CrossRef] [Green Version]
- Kawanami, D.; Maemura, K.; Takeda, N.; Harada, T.; Nojiri, T.; Imai, Y.; Manabe, I.; Utsunomiya, K.; Nagai, R. Direct reciprocal effects of resistin and adiponectin on vascular endothelial cells: A new insight into adipocytokine–endothelial cell interactions. Biochem. Biophys. Res. Commun. 2004, 314, 415–419. [Google Scholar] [CrossRef]
- Verma, S.; Li, S.-H.; Wang, C.-H.; Fedak, P.W.M.; Li, R.-K.; Weisel, R.D.; Mickle, D.A.G. Resistin Promotes Endothelial Cell Activation. Circulation 2003, 108, 736–740. [Google Scholar] [CrossRef]
- Luo, J.; Huang, L.; Wang, A.; Liu, Y.; Cai, R.; Li, W.; Zhou, M.-S. Resistin-Induced Endoplasmic Reticulum Stress Contributes to the Impairment of Insulin Signaling in Endothelium. Front. Pharmacol. 2018, 9, 1226. [Google Scholar] [CrossRef]
- Noval Rivas, M.; Arditi, M. Kawasaki disease: Pathophysiology and insights from mouse models. Nat. Rev. Rheumatol. 2020, 16, 391–405. [Google Scholar] [CrossRef]
- Liu, R.; He, B.; Gao, F.; Liu, Q.; Yi, Q. Relationship between adipokines and coronary artery aneurysm in children with Kawasaki disease. Transl. Res. 2012, 160, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Si, F.; Feng, S.; Yi, Q.; Liu, R. Resistin Enhances Inflammatory Cytokine Production in Coronary Artery Tissues by Activating the NF- κ B Signaling. Biomed Res. Int. 2016, 2016, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, T.-Y.; Hsiao, Y.-W.; Guo, S.-M.; Chang, S.-L.; Lin, Y.-J.; Lo, L.-W.; Hu, Y.-F.; Chung, F.-P.; Chao, T.-F.; Liao, J.-N.; et al. Resistin as a Biomarker for the Prediction of Left Atrial Substrate and Recurrence in Patients with Drug-Refractory Atrial Fibrillation Undergoing Catheter Ablation. Int. Heart J. 2020, 61, 517–523. [Google Scholar] [CrossRef] [PubMed]
- Zarling, J.M.; Shoyab, M.; Marquardt, H.; Hanson, M.B.; Lioubin, M.N.; Todaro, G.J. Oncostatin M: A growth regulator produced by differentiated histiocytic lymphoma cells. Proc. Natl. Acad. Sci. USA 1986, 83, 9739–9743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akarsu, M.; Hurşitoğlu, M.; Toprak, Z.; Yoldemir, Ş.A.; Altun, Ö.; Toprak, I.D.; Özcan, M.; Yürüyen, G.; Uğurlukişi, B.; Erdem, M.G.; et al. Relationships among oncostatin M, insulin resistance, and chronic inflammation: A pilot study. Arch. Endocrinol. Metab. 2019, 64, 38–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demyanets, S.; Kaun, C.; Rychli, K.; Pfaffenberger, S.; Kastl, S.P.; Hohensinner, P.J.; Rega, G.; Katsaros, K.M.; Afonyushkin, T.; Bochkov, V.N.; et al. Oncostatin M-enhanced vascular endothelial growth factor expression in human vascular smooth muscle cells involves PI3K-, p38 MAPK-, Erk1/2- and STAT1/STAT3-dependent pathways and is attenuated by interferon-γ. Basic Res. Cardiol. 2011, 106, 217–231. [Google Scholar] [CrossRef]
- Nguyen, M.T.; Prima, M.J.; Song, J.-A.; Kim, J.; Do, B.H.; Yoo, J.; Park, S.; Jang, J.; Lee, S.; Lee, E.; et al. Prokaryotic soluble overexpression and purification of oncostatin M using a fusion approach and genetically engineered E. coli strains. Sci. Rep. 2019, 9, 13706. [Google Scholar] [CrossRef]
- White, U.A.; Stewart, W.C.; Stephens, J.M. Gp130 cytokines exert differential patterns of crosstalk in adipocytes both in vitro and in vivo. Obesity 2011. [Google Scholar] [CrossRef] [Green Version]
- Lörchner, H.; Pöling, J.; Gajawada, P.; Hou, Y.; Polyakova, V.; Kostin, S.; Adrian-Segarra, J.M.; Boettger, T.; Wietelmann, A.; Warnecke, H.; et al. Myocardial healing requires Reg3β-dependent accumulation of macrophages in the ischemic heart. Nat. Med. 2015, 21, 353–362. [Google Scholar] [CrossRef]
- Luo, P.; Wang, P.-X.; Li, Z.-Z.; Zhang, X.-J.; Jiang, X.; Gong, J.; Qin, J.-J.; Guo, J.; Zhu, X.; Yang, S.; et al. Hepatic Oncostatin M Receptor β Regulates Obesity-Induced Steatosis and Insulin Resistance. Am. J. Pathol. 2016, 186, 1278–1292. [Google Scholar] [CrossRef] [Green Version]
- Richards, C.D. The Enigmatic Cytokine Oncostatin M and Roles in Disease. ISRN Inflamm. 2013, 2013, 1–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Li, J.; Qin, J.-J.; Cheng, W.-L.; Zhu, X.; Gong, F.-H.; She, Z.; Huang, Z.; Xia, H.; Li, H. Oncostatin M receptor β deficiency attenuates atherogenesis by inhibiting JAK2/STAT3 signaling in macrophages. J. Lipid Res. 2017, 58, 895–906. [Google Scholar] [CrossRef] [Green Version]
- Komori, T.; Morikawa, Y. Oncostatin M in the development of metabolic syndrome and its potential as a novel therapeutic target. Anat. Sci. Int. 2018, 93, 169–176. [Google Scholar] [CrossRef]
- Schnittker, D.; Kwofie, K.; Ashkar, A.; Trigatti, B.; Richards, C.D. Oncostatin M and TLR-4 Ligand Synergize to Induce MCP-1, IL-6, and VEGF in Human Aortic Adventitial Fibroblasts and Smooth Muscle Cells. Mediat. Inflamm. 2013, 2013, 1–14. [Google Scholar] [CrossRef]
- Rychli, K.; Kaun, C.; Hohensinner, P.J.; Rega, G.; Pfaffenberger, S.; Vyskocil, E.; Breuss, J.M.; Furnkranz, A.; Uhrin, P.; Zaujec, J.; et al. The inflammatory mediator oncostatin M induces angiopoietin 2 expression in endothelial cells in vitro and in vivo. J. Thromb. Haemost. 2010, 8, 596–604. [Google Scholar] [CrossRef] [PubMed]
- Martini, E.; Kunderfranco, P.; Peano, C.; Carullo, P.; Cremonesi, M.; Schorn, T.; Carriero, R.; Termanini, A.; Colombo, F.S.; Jachetti, E.; et al. Single-Cell Sequencing of Mouse Heart Immune Infiltrate in Pressure Overload-Driven Heart Failure Reveals Extent of Immune Activation. Circulation 2019, 140, 2089–2107. [Google Scholar] [CrossRef] [PubMed]
- Pöling, J.; Gajawada, P.; Richter, M.; Lörchner, H.; Polyakova, V.; Kostin, S.; Shin, J.; Boettger, T.; Walther, T.; Rees, W.; et al. Therapeutic targeting of the oncostatin M receptor-β prevents inflammatory heart failure. Basic Res. Cardiol. 2014, 109, 396. [Google Scholar] [CrossRef]
- Frangogiannis, N.G. The reparative function of cardiomyocytes in the infarcted myocardium. Cell Metab. 2015, 21, 797–798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Setiadi, H.; Yago, T.; Liu, Z.; McEver, R.P. Endothelial signaling by neutrophil-released oncostatin M enhances P-selectin–dependent inflammation and thrombosis. Blood Adv. 2019, 3, 168–183. [Google Scholar] [CrossRef] [Green Version]
- Kirchmer, M.N.; Franco, A.; Albasanz-Puig, A.; Murray, J.; Yagi, M.; Gao, L.; Dong, Z.M.; Wijelath, E.S. Modulation of vascular smooth muscle cell phenotype by STAT-1 and STAT-3. Atherosclerosis 2014, 234, 169–175. [Google Scholar] [CrossRef]
- Kakutani, Y.; Shioi, A.; Shoji, T.; Okazaki, H.; Koyama, H.; Emoto, M.; Inaba, M. Oncostatin M Promotes Osteoblastic Differentiation of Human Vascular Smooth Muscle Cells Through JAK3-STAT3 Pathway. J. Cell. Biochem. 2015, 116, 1325–1333. [Google Scholar] [CrossRef]
- Shioi, A.; Ikari, Y. Plaque calcification during atherosclerosis progression and regression. J. Atheroscler. Thromb. 2018, 25, 294–303. [Google Scholar] [CrossRef] [Green Version]
- Ghadge, A.A.; Khaire, A.A.; Kuvalekar, A.A. Adiponectin: A potential therapeutic target for metabolic syndrome. Cytokine Growth Factor Rev. 2018, 39, 151–158. [Google Scholar] [CrossRef]
- Oh, D.K.; Ciaraldi, T.; Henry, R.R. Adiponectin in health and disease. Diabetes Obes. Metab. 2007, 9, 282–289. [Google Scholar] [CrossRef] [PubMed]
- Harwood, H.J.; Heal, D.J.; Smith, S.L.; Jones, R.B. The adipocyte as an endocrine organ in the regulation of metabolic homeostasis. Neuropharmacology 2012, 63, 57–75. [Google Scholar] [CrossRef]
- Jackson, M.B.; Ahima, R.S. Neuroendocrine and metabolic effects of adipocyte-derived hormones. Clin. Sci. 2006, 110, 143–152. [Google Scholar] [CrossRef] [Green Version]
- Chou, I.-P.; Chiu, Y.-P.; Ding, S.-T.; Liu, B.-H.; Lin, Y.Y.; Chen, C.-Y. Adiponectin receptor 1 overexpression reduces lipid accumulation and hypertrophy in the heart of diet-induced obese mice—Possible involvement of oxidative stress and autophagy. Endocr. Res. 2014, 39, 173–179. [Google Scholar] [CrossRef]
- Koentges, C.; König, A.; Pfeil, K.; Hölscher, M.E.; Schnick, T.; Wende, A.R.; Schrepper, A.; Cimolai, M.C.; Kersting, S.; Hoffmann, M.M.; et al. Myocardial mitochondrial dysfunction in mice lacking adiponectin receptor 1. Basic Res. Cardiol. 2015, 110, 37. [Google Scholar] [CrossRef]
- Braun, M.; Hettinger, N.; Koentges, C.; Pfeil, K.; Cimolai, M.C.; Hoffmann, M.M.; Osterholt, M.; Doenst, T.; Bode, C.; Bugger, H. Myocardial mitochondrial and contractile function are preserved in mice lacking adiponectin. PLoS ONE 2015, 10, e0119416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berg, A.H.; Combs, T.P.; Scherer, P.E. ACRP30/adiponectin: An adipokine regulating glucose and lipid metabolism. Trends Endocrinol. Metab. 2002, 13, 84–89. [Google Scholar] [CrossRef]
- Hui, X.; Lam, K.S.L.; Vanhoutte, P.M.; Xu, A. Adiponectin and cardiovascular health: An update. Br. J. Pharmacol. 2012, 165, 574–590. [Google Scholar] [CrossRef] [Green Version]
- Ouchi, N.; Ohishi, M.; Kihara, S.; Funahashi, T.; Nakamura, T.; Nagaretani, H.; Kumada, M.; Ohashi, K.; Okamoto, Y.; Nishizawa, H.; et al. Association of hypoadiponectinemia with impaired vasoreactivity. Hypertension 2003, 42, 231–234. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Xia, L.; Zhang, F.; Zhu, D.; Xin, C.; Wang, H.; Zhang, F.; Guo, X.; Lee, Y.; Zhang, L.; et al. A novel mechanism of diabetic vascular endothelial dysfunction: Hypoadiponectinemia-induced NLRP3 inflammasome activation. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 1556–1567. [Google Scholar] [CrossRef] [PubMed]
- Iwashima, Y.; Katsuya, T.; Ishikawa, K.; Ouchi, N.; Ohishi, M.; Sugimoto, K.; Fu, Y.; Motone, M.; Yamamoto, K.; Matsuo, A.; et al. Hypoadiponectinemia Is an Independent Risk Factor for Hypertension. Hypertension 2004, 43, 1318–1323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chow, W.-S.; Cheung, B.M.Y.; Tso, A.W.K.; Xu, A.; Wat, N.M.S.; Fong, C.H.Y.; Ong, L.H.Y.; Tam, S.; Tan, K.C.B.; Janus, E.D.; et al. Hypoadiponectinemia as a Predictor for the Development of Hypertension. Hypertension 2007, 49, 1455–1461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashimoto, N.; Kanda, J.; Nakamura, T.; Horie, A.; Kurosawa, H.; Hashimoto, T.; Sato, K.; Kushida, S.; Suzuki, M.; Yano, S.; et al. Association of hypoadiponectinemia in men with early onset of coronary heart disease and multiple coronary artery stenoses. Metabolism 2006, 55, 1653–1657. [Google Scholar] [CrossRef]
- Kumada, M.; Kihara, S.; Sumitsuji, S.; Kawamoto, T.; Matsumoto, S.; Ouchi, N.; Arita, Y.; Okamoto, Y.; Shimomura, I.; Hiraoka, H.; et al. Association of Hypoadiponectinemia With Coronary Artery Disease in Men. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 85–89. [Google Scholar] [CrossRef] [Green Version]
- Mohty, D.; Pibarot, P.; Côté, N.; Cartier, A.; Audet, A.; Després, J.P.; Mathieu, P. Hypoadiponectinemia Is Associated with Valvular Inflammation and Faster Disease Progression in Patients with Aortic Stenosis. Cardiology 2011, 118, 140–146. [Google Scholar] [CrossRef]
- Yanai, H.; Yoshida, H. Beneficial Effects of Adiponectin on Glucose and Lipid Metabolism and Atherosclerotic Progression: Mechanisms and Perspectives. Int. J. Mol. Sci. 2019, 20, 1190. [Google Scholar] [CrossRef] [Green Version]
- Devaraj, S.; Torok, N.; Dasu, M.R.; Samols, D.; Jialal, I. Adiponectin Decreases C-Reactive Protein Synthesis and Secretion from Endothelial Cells. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 1368–1374. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Wei, Y.; Wang, L.; Wang, X.; Du, X.; Sun, Z.; Dong, N.; Chen, X. Decreased adiponectin and increased inflammation expression in epicardial adipose tissue in coronary artery disease. Cardiovasc. Diabetol. 2011, 10, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hattori, Y.; Nakano, Y.; Hattori, S.; Tomizawa, A.; Inukai, K.; Kasai, K. High molecular weight adiponectin activates AMPK and suppresses cytokine-induced NF-κB activation in vascular endothelial cells. FEBS Lett. 2008, 582, 1719–1724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Mahadev, K.; Fuchsel, L.; Ouedraogo, R.; Xu, S.-Q.; Goldstein, B.J. Adiponectin suppresses IκB kinase activation induced by tumor necrosis factor-α or high glucose in endothelial cells: Role of cAMP and AMP kinase signaling. Am. J. Physiol. Metab. 2007, 293, E1836–E1844. [Google Scholar] [CrossRef] [PubMed]
- Ouedraogo, R.; Gong, Y.; Berzins, B.; Wu, X.; Mahadev, K.; Hough, K.; Chan, L.; Goldstein, B.J.; Scalia, R. Adiponectin deficiency increases leukocyte-endothelium interactions via upregulation of endothelial cell adhesion molecules in vivo. J. Clin. Invest. 2007, 117, 1718–1726. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Park, Y.; Zhang, C. Coronary and Aortic Endothelial Function Affected by Feedback Between Adiponectin and Tumor Necrosis Factor α in Type 2 Diabetic Mice. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 2156–2163. [Google Scholar] [CrossRef] [Green Version]
- Shimabukuro, M.; Higa, N.; Asahi, T.; Oshiro, Y.; Takasu, N.; Tagawa, T.; Ueda, S.; Shimomura, I.; Funahashi, T.; Matsuzawa, Y. Hypoadiponectinemia Is Closely Linked to Endothelial Dysfunction in Man. J. Clin. Endocrinol. Metab. 2003, 88, 3236–3240. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Tao, L.; Yuan, Y.; Jiao, X.; Lau, W.B.; Wang, Y.; Christopher, T.; Lopez, B.; Chan, L.; Goldstein, B.; et al. Endothelial dysfunction in adiponectin deficiency and its mechanisms involved. J. Mol. Cell. Cardiol. 2009, 46, 413–419. [Google Scholar] [CrossRef] [Green Version]
- Deng, G.; Long, Y.; Yu, Y.-R.; Li, M.-R. Adiponectin directly improves endothelial dysfunction in obese rats through the AMPK-eNOS Pathway. Int. J. Obes. 2010, 34, 165–171. [Google Scholar] [CrossRef] [Green Version]
- Kelley, N.; Jeltema, D.; Duan, Y.; He, Y. The NLRP3 Inflammasome: An Overview of Mechanisms of Activation and Regulation. Int. J. Mol. Sci. 2019, 20, 3328. [Google Scholar] [CrossRef] [Green Version]
- Van de Voorde, J.; Pauwels, B.; Boydens, C.; Decaluwé, K. Adipocytokines in relation to cardiovascular disease. Metabolism 2013, 62, 1513–1521. [Google Scholar] [CrossRef]
- Piñeiro, R.; Iglesias, M.J.; Gallego, R.; Raghay, K.; Eiras, S.; Rubio, J.; Diéguez, C.; Gualillo, O.; González-Juanatey, J.R.; Lago, F. Adiponectin is synthesized and secreted by human and murine cardiomyocytes. FEBS Lett. 2005, 579, 5163–5169. [Google Scholar] [CrossRef] [Green Version]
- Boddu, N.J.; Theus, S.; Luo, S.; Wei, J.Y.; Ranganathan, G. Is the lack of adiponectin associated with increased ER/SR stress and inflammation in the heart? Adipocyte 2014, 3, 10–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, Y.; Wang, X.F.; Lang, Z.Q.; Jin, Y.C.; Fu, J.R.; Xv, X.M.; Sun, S.T.; Xin, X.; Zhang, L.S. Adiponectin is protective against endoplasmic reticulum stress-induced apoptosis of endothelial cells in sepsis. Braz. J. Med. Biol. Res. 2018, 51, e7747. [Google Scholar] [CrossRef] [PubMed]
- Ding, W.; Zhang, X.; Huang, H.; Ding, N.; Zhang, S.; Hutchinson, S.Z.; Zhang, X. Adiponectin protects rat myocardium against chronic intermittent hypoxia-induced injury via inhibition of endoplasmic reticulum stress. PLoS ONE 2014, 9, e94545. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Bian, Y.; Bai, R.; Li, H.; Fu, M.; Xiao, C. Globular adiponectin attenuates myocardial ischemia/reperfusion injury by upregulating endoplasmic reticulum Ca2+-ATPase activity and inhibiting endoplasmic reticulum stress. J. Cardiovasc. Pharmacol. 2013, 62, 143–153. [Google Scholar] [CrossRef]
- Bian, Y.-F.; Hao, X.-Y.; Gao, F.; Yang, H.-Y.; Zang, N.; Xiao, C.-S. Adiponectin attenuates hypoxia/reoxygenation-induced cardiomyocyte injury through inhibition of endoplasmic reticulum stress. J. Investig. Med. 2011, 59, 921–925. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Liu, Y.; Yu, Q.; Yang, Q.; Li, B.; Sun, L.; Yan, W.; Cai, X.; Gao, E.; Xiong, L.; et al. TNF-α antagonism ameliorates myocardial ischemia-reperfusion injury in mice by upregulating adiponectin. Am. J. Physiol. Heart Circ. Physiol. 2015, 308, H1583–H1591. [Google Scholar] [CrossRef] [Green Version]
- Oh-I, S.; Shimizu, H.; Satoh, T.; Okada, S.; Adachi, S.; Inoue, K.; Eguchi, H.; Yamamoto, M.; Imaki, T.; Hashimoto, K.; et al. Identification of nesfatin-1 as a satiety molecule in the hypothalamus. Nature 2006, 443, 709–712. [Google Scholar] [CrossRef]
- Feijóo-Bandín, S.; Rodríguez-Penas, D.; García-Rúa, V.; Mosquera-Leal, A.; González-Juanatey, J.R.; Lago, F. Nesfatin-1: A new energy-regulating peptide with pleiotropic functions. Implications at cardiovascular level. Endocrine 2016, 52, 11–29. [Google Scholar] [CrossRef]
- Ramanjaneya, M.; Chen, J.; Brown, J.E.; Tripathi, G.; Hallschmid, M.; Patel, S.; Kern, W.; Hillhouse, E.W.; Lehnert, H.; Tan, B.K.; et al. Identification of nesfatin-1 in human and murine adipose tissue: A novel depot-specific adipokine with increased levels in obesity. Endocrinology 2010, 151, 3169–3180. [Google Scholar] [CrossRef] [Green Version]
- St-Pierre, D.H.; Martin, J.; Shimizu, H.; Tagaya, Y.; Tsuchiya, T.; Marceau, S.; Biertho, L.; Bastien, M.; Caron-Cantin, S.-M.; Simard, S.; et al. Association between nesfatin-1 levels and metabolic improvements in severely obese patients who underwent biliopancreatic derivation with duodenal switch. Peptides 2016, 86, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, R.; Reingold, B.K.; Gao, X.; Gaidhu, M.P.; Tsushima, R.G.; Unniappan, S. Nesfatin-1 exerts a direct, glucose-dependent insulinotropic action on mouse islet β- and MIN6 cells. J. Endocrinol. 2011, 208, R9–R16. [Google Scholar] [CrossRef] [Green Version]
- Bonnet, M.S.; Pecchi, E.; Trouslard, J.; Jean, A.; Dallaporta, M.; Troadec, J.-D. Central nesfatin-1-expressing neurons are sensitive to peripheral inflammatory stimulus. J. Neuroinflamm. 2009, 6, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, L.; Xu, K.; Li, J.; Zhou, X.; Xu, L.; Wu, Z.; Ma, C.; Ran, J.; Hu, P.; Bao, J.; et al. Nesfatin-1 suppresses interleukin-1β-induced inflammation, apoptosis, and cartilage matrix destruction in chondrocytes and ameliorates osteoarthritis in rats. Aging 2020, 12, 1760–1777. [Google Scholar] [CrossRef]
- Wang, Z.-Z.; Chen, S.-C.; Zou, X.-B.; Tian, L.-L.; Sui, S.-H.; Liu, N.-Z. Nesfatin-1 alleviates acute lung injury through reducing inflammation and oxidative stress via the regulation of HMGB1. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 5071–5081. [Google Scholar] [CrossRef]
- Tang, C.-H.; Fu, X.-J.; Xu, X.-L.; Wei, X.-J.; Pan, H.-S. The anti-inflammatory and anti-apoptotic effects of nesfatin-1 in the traumatic rat brain. Peptides 2012, 36, 39–45. [Google Scholar] [CrossRef]
- Özsavcí, D.; Erşahin, M.; Şener, A.; Özakpinar, Ö.B.; Toklu, H.Z.; Akakín, D.; Şener, G.; Yeğen, B.Ç. The novel function of nesfatin-1 as an anti-inflammatory and antiapoptotic peptide in subarachnoid hemorrhage-induced oxidative brain damage in rats. Neurosurgery 2011, 68, 1699–1708. [Google Scholar] [CrossRef]
- Acik, V.; Matyar, S.; Arslan, A.; İstemen, İ.; Olguner, S.K.; Arslan, B.; Gezercan, Y.; Ökten, A.İ. Relationshıp of spontaneous subarachnoid haemorrhage and cerebral aneurysm to serum Visfatin and Nesfatin-1 levels. Clin. Neurol. Neurosurg. 2020, 194, 105837. [Google Scholar] [CrossRef] [PubMed]
- Feijóo-Bandín, S.; Rodríguez-Penas, D.; García-Rúa, V.; Mosquera-Leal, A.; Otero, M.F.; Pereira, E.; Rubio, J.; Martínez, I.; Seoane, L.M.; Gualillo, O.; et al. Nesfatin-1 in human and murine cardiomyocytes: Synthesis, secretion, and mobilization of GLUT-4. Endocrinology 2013, 154, 4757–4767. [Google Scholar] [CrossRef] [Green Version]
- Ibe, S.; Kishimoto, Y.; Niki, H.; Saita, E.; Umei, T.; Miura, K.; Ikegami, Y.; Ohmori, R.; Kondo, K.; Momiyama, Y. Associations between plasma nesfatin-1 levels and the presence and severity of coronary artery disease. Heart Vessels 2019, 34, 965–970. [Google Scholar] [CrossRef]
- Serdar Kuyumcu, M.; Kuyumcu, A.; Yayla, Ç.; Bilal Özbay, M.; Ünal, S.; Açar, B.; Nural, C.; Şenat, A.; Samur, G. The Relationship between Nesfatin-1 Levels and SYNTAX Score in Patients with Non-ST Segment Elevation Myocardial Infarction. Acta Cardiol. Sin. 2018, 34, 386–393. [Google Scholar] [CrossRef] [PubMed]
- Kuyumcu, A.; Kuyumcu, M.S.; Ozbay, M.B.; Ertem, A.G.; Samur, G. Nesfatin-1: A novel regulatory peptide associated with acute myocardial infarction and Mediterranean diet. Peptides 2019, 114, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Dai, H.; Li, X.; He, T.; Wang, Y.; Wang, Z.; Wang, S.; Xing, M.; Sun, W.; Ding, H. Decreased plasma nesfatin-1 levels in patients with acute myocardial infarction. Peptides 2013, 46, 167–171. [Google Scholar] [CrossRef] [PubMed]
- Sivri, S.; Sökmen, E.; Çelik, M.; Güçlü, K. Nesfatin-1 Levels Predict Angiographic No-Reflow in Patients with ST-Segment Elevation Myocardial Infarction. Acta Cardiol. Sin. 2020, 36, 318–325. [Google Scholar] [CrossRef] [PubMed]
- Kirisci, M.; Yardimci, M.M.; Kocarslan, A.; Sokmen, A.; Doganer, A.; Gunes, H. Nesfatin 1: A promising biomarker predicting successful reperfusion after coronary artery bypass surgery. Bratisl. Med. J. 2020, 121, 282–286. [Google Scholar] [CrossRef] [PubMed]
- Tasatargil, A.; Kuscu, N.; Dalaklioglu, S.; Adiguzel, D.; Celik-Ozenci, C.; Ozdem, S.; Barutcigil, A.; Ozdem, S. Cardioprotective effect of nesfatin-1 against isoproterenol-induced myocardial infarction in rats: Role of the Akt/GSK-3β pathway. Peptides 2017, 95, 1–9. [Google Scholar] [CrossRef]
- Naseroleslami, M.; Sharifi, M.; Rakhshan, K.; Mokhtari, B.; Aboutaleb, N. Nesfatin-1 attenuates injury in a rat model of myocardial infarction by targeting autophagy, inflammation, and apoptosis. Arch. Physiol. Biochem. 2020, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Feijóo-Bandín, S.; Aragón-Herrera, A.; Rodríguez-Penas, D.; Portolés, M.; Roselló-Lletí, E.; Rivera, M.; González-Juanatey, J.R.; Lago, F. Relaxin-2 in Cardiometabolic Diseases: Mechanisms of Action and Future Perspectives. Front. Physiol. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Olefsky, J.M.; Saekow, M.; Kroc, R.L. Potentiation of insulin binding and insulin action by purified porcine relaxin. Ann. N. Y. Acad. Sci. 1982, 380, 200–216. [Google Scholar] [CrossRef]
- Jarrett, J.C.; Ballejo, G.; Saleem, T.H.; Tsibris, J.C.M.; Spellacy, W.N. The effect of prolactin and relaxin on insulin binding by adipocytes from pregnant women. Am. J. Obstet. Gynecol. 1984, 149, 250–255. [Google Scholar] [CrossRef]
- Bani, G.; Bianchi, S.; Formigli, L.; Bigazzi, M. Responsiveness of Mouse Parametrial Fat to Relaxin. Cells Tissues Organs 1989, 134, 128–132. [Google Scholar] [CrossRef]
- Martin, B.; Romero, G.; Salama, G. Cardioprotective actions of relaxin. Mol. Cell. Endocrinol. 2019, 487, 45–53. [Google Scholar] [CrossRef]
- Masini, E.; Bani, D.; Bigazzi, M.; Mannaioni, P.F.; Bani-Sacchi, T. Effects of relaxin on mast cells. In vitro and in vivo studies in rats and guinea pigs. J. Clin. Invest. 1994, 94, 1974–1980. [Google Scholar] [CrossRef] [PubMed]
- Bani, D.; Bigazzi, M.; Masini, E.; Bani, G.; Sacchi, T.B. Relaxin depresses platelet aggregation: In vitro studies on isolated human and rabbit platelets. Lab. Invest. 1995, 73, 709–716. [Google Scholar]
- Bani, D.; Masini, E.; Bello, M.G.; Bigazzi, M.; Bani Sacchi, T.; Sacchi, T.B. Relaxin protects against myocardial injury caused by ischemia and reperfusion in rat heart. Am. J. Pathol. 1998, 152, 1367–1376. [Google Scholar]
- Masini, E.; Bani, D.; Bello, M.G.; Bigazzi, M.; Mannaioni, P.F.; Sacchi, T.B. Relaxin counteracts myocardial damage induced by ischemia-reperfusion in isolated guinea pig hearts: Evidence for an involvement of nitric oxide. Endocrinology 1997, 138, 4713–4720. [Google Scholar] [CrossRef]
- Nistri, S.; Cinci, L.; Perna, A.M.; Masini, E.; Mastroianni, R.; Bani, D. Relaxin induces mast cell inhibition and reduces ventricular arrhythmias in a swine model of acute myocardial infarction. Pharmacol. Res. 2008, 57, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Nistri, S.; Chiappini, L.; Sassoli, C.; Bani, D. Relaxin inhibits lipopolysaccharide-induced adhesion of neutrophils to coronary endothelial cells by a nitric oxide-mediated mechanism. FASEB J. 2003, 17, 2109–2111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masini, E.; Nistri, S.; Vannacci, A.; Bani Sacchi, T.; Novelli, A.; Bani, D. Relaxin Inhibits the Activation of Human Neutrophils: Involvement of the Nitric Oxide Pathway. Endocrinology 2004, 145, 1106–1112. [Google Scholar] [CrossRef] [Green Version]
- Valle Raleigh, J.; Mauro, A.G.; Devarakonda, T.; Marchetti, C.; He, J.; Kim, E.; Filippone, S.; Das, A.; Toldo, S.; Abbate, A.; et al. Reperfusion therapy with recombinant human relaxin-2 (Serelaxin) attenuates myocardial infarct size and NLRP3 inflammasome following ischemia/reperfusion injury via eNOS-dependent mechanism. Cardiovasc. Res. 2017, 2, 609–619. [Google Scholar] [CrossRef] [Green Version]
- Toldo, S.; Mauro, A.G.; Cutter, Z.; Abbate, A. Inflammasome, pyroptosis, and cytokines in myocardial ischemia-reperfusion injury. Am. J. Physiol. Circ. Physiol. 2018, 315, H1553–H1568. [Google Scholar] [CrossRef] [PubMed]
- Beiert, T.; Tiyerili, V.; Knappe, V.; Effelsberg, V.; Linhart, M.; Stöckigt, F.; Klein, S.; Schierwagen, R.; Trebicka, J.; Nickenig, G.; et al. Relaxin reduces susceptibility to post-infarct atrial fibrillation in mice due to anti-fibrotic and anti-inflammatory properties. Biochem. Biophys. Res. Commun. 2017, 490, 643–649. [Google Scholar] [CrossRef]
- Beiert, T.; Knappe, V.; Tiyerili, V.; Stöckigt, F.; Effelsberg, V.; Linhart, M.; Steinmetz, M.; Klein, S.; Schierwagen, R.; Trebicka, J.; et al. Chronic lower-dose relaxin administration protects from arrhythmia in experimental myocardial infarction due to anti-inflammatory and anti-fibrotic properties. Int. J. Cardiol. 2018, 250, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Mas, J.; Lax, A.; Asensio-Lopez, M.C.; Lencina, M.; Fernandez-del Palacio, M.J.; Soriano-Filiu, A.; de Boer, R.A.; Pascual-Figal, D.A. Early Anti-inflammatory and Pro-angiogenic Myocardial Effects of Intravenous Serelaxin Infusion for 72 H in an Experimental Rat Model of Acute Myocardial Infarction. J. Cardiovasc. Transl. Res. 2017, 10, 460–469. [Google Scholar] [CrossRef]
- Gao, X.-M.; Su, Y.; Moore, S.; Han, L.-P.; Kiriazis, H.; Lu, Q.; Zhao, W.-B.; Ruze, A.; Fang, B.-B.; Duan, M.-J.; et al. Relaxin mitigates microvascular damage and inflammation following cardiac ischemia—Reperfusion. Basic Res. Cardiol. 2019, 114, 30. [Google Scholar] [CrossRef]
- Van den Berg, N.W.E.; de Groot, J.R. Myocardial infarction, atrial fibrillation and mortality: Timing is everything. Neth. Heart. J. 2015, 23, 428–429. [Google Scholar] [CrossRef] [PubMed]
- Samuel, C.S.; Royce, S.G.; Hewitson, T.D.; Denton, K.M.; Cooney, T.E.; Bennett, R.G. Anti-fibrotic actions of relaxin. Br. J. Pharmacol. 2017, 174, 962–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.; Zhu, H.; Yang, Q.; Sun, Y. Effects of relaxin on cardiac fibrosis, apoptosis, and tachyarrhythmia in rats with myocardial infarction. Biomed. Pharmacother. 2016, 84, 348–355. [Google Scholar] [CrossRef]
- Martin, B.; Gabris-Weber, B.A.; Reddy, R.; Romero, G.; Chattopadhyay, A.; Salama, G. Relaxin reverses inflammatory and immune signals in aged hearts. PLoS ONE 2018, 13, e0190935. [Google Scholar] [CrossRef] [Green Version]
- Brecht, A.; Bartsch, C.; Baumann, G.; Stangl, K.; Dschietzig, T. Relaxin inhibits early steps in vascular inflammation. Regul. Pept. 2011, 166, 76–82. [Google Scholar] [CrossRef]
- Dschietzig, T.; Bartsch, C.; Baumann, G.; Stangl, K. RXFP1-inactive relaxin activates human glucocorticoid receptor: Further investigations into the relaxin-GR pathway. Regul. Pept. 2009, 154, 77–84. [Google Scholar] [CrossRef]
- Bathgate, R.A.D.; Halls, M.L.; van der Westhuizen, E.T.; Callander, G.E.; Kocan, M.; Summers, R.J. Relaxin family peptides and their receptors. Physiol. Rev. 2013, 93, 405–480. [Google Scholar] [CrossRef] [PubMed]
- Collino, M.; Rogazzo, M.; Pini, A.; Benetti, E.; Rosa, A.C.; Chiazza, F.; Fantozzi, R.; Bani, D.; Masini, E. Acute treatment with relaxin protects the kidney against ischaemia/reperfusion injury. J. Cell. Mol. Med. 2013, 17, 1494–1505. [Google Scholar] [CrossRef] [PubMed]
- Boehnert, M.U.; Armbruster, F.P.; Hilbig, H. Relaxin as a protective substance in the preserving solution for liver transplantation: Spectrophotometric in vivo imaging of local oxygen supply in an isolated perfused rat liver model. Ann. N. Y. Acad. Sci. 2009, 1160, 320–321. [Google Scholar] [CrossRef] [PubMed]
- Komiya, T.; Tanigawa, Y.; Hirohashi, S. Cloning of the Novel Gene Intelectin, Which Is Expressed in Intestinal Paneth Cells in Mice. Biochem. Biophys. Res. Commun. 1998, 251, 759–762. [Google Scholar] [CrossRef]
- Suzuki, Y.A.; Shin, K.; Lönnerdal, B. Molecular Cloning and Functional Expression of a Human Intestinal Lactoferrin Receptor ‡. Biochemistry 2001, 40, 15771–15779. [Google Scholar] [CrossRef]
- Lee, J.-K.; Schnee, J.; Pang, M.; Wolfert, M.; Baum, L.G.; Moremen, K.W.; Pierce, M. Human homologs of the Xenopus oocyte cortical granule lectin XL35. Glycobiology 2001, 11, 65–73. [Google Scholar] [CrossRef] [Green Version]
- Schäffler, A.; Neumeier, M.; Herfarth, H.; Fürst, A.; Schölmerich, J.; Büchler, C. Genomic structure of human omentin, a new adipocytokine expressed in omental adipose tissue. Biochim. Biophys. Acta Gene Struct. Expr. 2005, 1732, 96–102. [Google Scholar] [CrossRef]
- Yang, R.-Z.; Lee, M.-J.; Hu, H.; Pray, J.; Wu, H.-B.; Hansen, B.C.; Shuldiner, A.R.; Fried, S.K.; McLenithan, J.C.; Gong, D.-W. Identification of omentin as a novel depot-specific adipokine in human adipose tissue: Possible role in modulating insulin action. Am. J. Physiol. Metab. 2006, 290, E1253–E1261. [Google Scholar] [CrossRef]
- Fain, J.N.; Sacks, H.S.; Buehrer, B.; Bahouth, S.W.; Garrett, E.; Wolf, R.Y.; Carter, R.A.; Tichansky, D.S.; Madan, A.K. Identification of omentin mRNA in human epicardial adipose tissue: Comparison to omentin in subcutaneous, internal mammary artery periadventitial and visceral abdominal depots. Int. J. Obes. 2008, 32, 810–815. [Google Scholar] [CrossRef] [Green Version]
- Svensson, H.; Odén, B.; Edén, S.; Lönn, M. Adiponectin, chemerin, cytokines, and dipeptidyl peptidase 4 are released from human adipose tissue in a depot-dependent manner: An in vitro system including human serum albumin. BMC Endocr. Disord. 2014, 14, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, Y.; Ji, Q.; Cai, L.; Huang, F.; Lai, Y.; Liu, Y.; Yu, J.; Han, B.; Zhu, E.; Zhang, J.; et al. Association between omentin-1 expression in human epicardial adipose tissue and coronary atherosclerosis. Cardiovasc. Diabetol. 2016, 15, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matloch, Z.; Kratochvílová, H.; Cinkajzlová, A.; Lipš, M.; Kopecký, P.; Pořízka, M.; Haluzíková, D.; Lindner, J.; Mráz, M.; Kloučková, J.; et al. Changes in Omentin Levels and Its mRNA Expression in Epicardial Adipose Tissue in Patients Undergoing Elective Cardiac Surgery: The Influence of Type 2 Diabetes and Coronary Heart Disease. Physiol. Res. 2018, 67, 881–890. [Google Scholar] [CrossRef] [PubMed]
- Harada, K.; Shibata, R.; Ouchi, N.; Tokuda, Y.; Funakubo, H.; Suzuki, M.; Kataoka, T.; Nagao, T.; Okumura, S.; Shinoda, N.; et al. Increased expression of the adipocytokine omentin in the epicardial adipose tissue of coronary artery disease patients. Atherosclerosis 2016, 251, 299–304. [Google Scholar] [CrossRef] [PubMed]
- Berti, L.; Hartwig, S.; Irmler, M.; Rädle, B.; Siegel-Axel, D.; Beckers, J.; Lehr, S.; Al-Hasani, H.; Häring, H.-U.; Hrabě de Angelis, M.; et al. Impact of fibroblast growth factor 21 on the secretome of human perivascular preadipocytes and adipocytes: A targeted proteomics approach. Arch. Physiol. Biochem. 2016, 122, 281–288. [Google Scholar] [CrossRef]
- De Souza Batista, C.M.; Yang, R.-Z.; Lee, M.-J.; Glynn, N.M.; Yu, D.-Z.; Pray, J.; Ndubuizu, K.; Patil, S.; Schwartz, A.; Kligman, M.; et al. Omentin Plasma Levels and Gene Expression Are Decreased in Obesity. Diabetes 2007, 56, 1655–1661. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Trasancos, Á.; Agra, R.M.; García-Acuña, J.M.; Fernández, Á.L.; González-Juanatey, J.R.; Eiras, S. Omentin treatment of epicardial fat improves its anti-inflammatory activity and paracrine benefit on smooth muscle cells. Obesity 2017, 25, 1042–1049. [Google Scholar] [CrossRef]
- Zhou, H.; Zhang, Z.; Qian, G.; Zhou, J. Omentin-1 attenuates adipose tissue inflammation via restoration of TXNIP/NLRP3 signaling in high-fat diet-induced obese mice. Fundam. Clin. Pharmacol. 2020. [Google Scholar] [CrossRef]
- Wang, J.; Gao, Y.; Lin, F.; Han, K.; Wang, X. Omentin-1 attenuates lipopolysaccharide (LPS)-induced U937 macrophages activation by inhibiting the TLR4/MyD88/NF-κB signaling. Arch. Biochem. Biophys. 2020, 679, 108187. [Google Scholar] [CrossRef]
- Yıldız, S.S.; Sahin, I.; Cetinkal, G.; Aksan, G.; Kucuk, S.H.; Keskin, K.; Cetin, S.; Sigirci, S.; Avcı, İ.İ.; Kilci, H.; et al. Usefulness of Serum Omentin-1 Levels for the Prediction of Adverse Cardiac Events in Patients with Hypertrophic Cardiomyopathy. Med. Princ. Pract. 2018, 27, 107–114. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.-H.; Dou, L.-Z.; Gu, C.; Wang, X.-Q. Plasma levels of omentin-1 and visfatin in senile patients with coronary heart disease and heart failure. Asian Pac. J. Trop. Med. 2014, 7, 55–62. [Google Scholar] [CrossRef] [Green Version]
- Narumi, T.; Watanabe, T.; Kadowaki, S.; Kinoshita, D.; Yokoyama, M.; Honda, Y.; Otaki, Y.; Nishiyama, S.; Takahashi, H.; Arimoto, T.; et al. Impact of serum omentin-1 levels on cardiac prognosis in patients with heart failure. Cardiovasc. Diabetol. 2014, 13, 84. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Lin, Y.; Zhang, S.; Wang, Z.; Zhang, J.; Chang, C.; Liu, L.; Ji, Q.; Liu, X. Circulating Omentin-1 Levels Are Decreased in Dilated Cardiomyopathy Patients with Overt Heart Failure. Dis. Markers 2016, 2016, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Biscetti, F.; Nardella, E.; Bonadia, N.; Angelini, F.; Pitocco, D.; Santoliquido, A.; Filipponi, M.; Landolfi, R.; Flex, A. Association between plasma omentin-1 levels in type 2 diabetic patients and peripheral artery disease. Cardiovasc. Diabetol. 2019, 18, 74. [Google Scholar] [CrossRef] [PubMed]
- Onur, I.; Oz, F.; Yildiz, S.; Kuplay, H.; Yucel, C.; Sigirci, S.; Elitok, A.; Pilten, S.; Kasali, K.; Yasar Cizgici, A.; et al. A decreased serum omentin-1 level may be an independent risk factor for peripheral arterial disease. Int. Angiol. 2014, 33, 455–460. [Google Scholar]
- Baig, M.; Alghalayini, K.W.; Gazzaz, Z.J.; Atta, H. Association of Serum Omentin-1, Chemerin, and Leptin with Acute Myocardial Infarction and its Risk Factors. Pakistan J. Med. Sci. 2020, 36, 1183–1188. [Google Scholar] [CrossRef] [PubMed]
- Kadoglou, N.P.E.; Tahmatzidis, D.K.; Giannakoulas, C.; Kapelouzou, A.; Gkontopoulos, A.; Parissis, J.; Lampropoulos, S.; Kottas, G. Serum levels of novel adipokines, omentin-1 and chemerin, in patients with acute myocardial infarction: KOZANI STUDY. J. Cardiovasc. Med. 2015, 16, 341–346. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Liu, F.; Han, F.; Lv, L.; Tang, C.-E.; Luo, F. Omentin-1 Ameliorated Free Fatty Acid-Induced Impairment in Proliferation, Migration, and Inflammatory States of HUVECs. Cardiol. Res. Pract. 2020, 20, 3054379. [Google Scholar] [CrossRef] [Green Version]
- Çimen, A.R.; Cerit, E.T.; Iyidir, O.T.; Karakus, R.; Uyar, B.B.; Toruner, F.B.; Cakir, N.; Arslan, M. Serum omentin-1 levels and endothelial dysfunction in obesity. Acta Endocrinol. 2017, 13, 138–143. [Google Scholar] [CrossRef]
- Hayashi, M.; Morioka, T.; Hatamori, M.; Kakutani, Y.; Yamazaki, Y.; Kurajoh, M.; Motoyama, K.; Mori, K.; Fukumoto, S.; Shioi, A.; et al. Plasma omentin levels are associated with vascular endothelial function in patients with type 2 diabetes at elevated cardiovascular risk. Diabetes Res. Clin. Pract. 2019, 148, 160–168. [Google Scholar] [CrossRef]
- Moreno-Navarrete, J.M.; Ortega, F.; Castro, A.; Sabater, M.; Ricart, W.; Fernández-Real, J.M. Circulating Omentin as a Novel Biomarker of Endothelial Dysfunction. Obesity 2011, 19, 1552–1559. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Fang, S.; Liu, X.; Li, J.; Wang, X.; Cui, J.; Chen, T.; Li, Z.; Yang, F.; Tian, J.; et al. Omentin-1 protects against high glucose-induced endothelial dysfunction via the AMPK/PPARδ signaling pathway. Biochem. Pharmacol. 2020, 174, 113830. [Google Scholar] [CrossRef] [PubMed]
- Yamawaki, H.; Kuramoto, J.; Kameshima, S.; Usui, T.; Okada, M.; Hara, Y. Omentin, a novel adipocytokine inhibits TNF-induced vascular inflammation in human endothelial cells. Biochem. Biophys. Res. Commun. 2011, 408, 339–343. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X.; Li, X.; Liu, F.; Tan, H.; Shang, D. Omentin inhibits TNF-α-induced expression of adhesion molecules in endothelial cells via ERK/NF-κB pathway. Biochem. Biophys. Res. Commun. 2012, 425, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Binti Kamaruddin, N.A.; Fong, L.Y.; Tan, J.J.; Abdullah, M.N.H.; Singh Cheema, M.; Bin Yakop, F.; Yong, Y.K. Cytoprotective Role of Omentin Against Oxidative Stress-Induced Vascular Endothelial Cells Injury. Molecules 2020, 25, 2534. [Google Scholar] [CrossRef]
- Luo, L.; Liu, M. Adipose tissue in control of metabolism. J. Endocrinol. 2016, 231, R77–R99. [Google Scholar] [CrossRef] [Green Version]
- Ushach, I.; Arrevillaga-Boni, G.; Heller, G.N.; Pone, E.; Hernandez-Ruiz, M.; Catalan-Dibene, J.; Hevezi, P.; Zlotnik, A. Meteorin-like/Meteorin-β Is a Novel Immunoregulatory Cytokine Associated with Inflammation. J. Immunol. 2018, 201, 3669–3676. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.O.; Byun, W.S.; Kang, M.J.; Han, J.A.; Moon, J.; Shin, M.-J.; Lee, H.J.; Chung, J.H.; Lee, J.-S.; Son, C.-G.; et al. The myokine meteorin-like (metrnl) improves glucose tolerance in both skeletal muscle cells and mice by targeting AMPKα2. FEBS J. 2020, 287, 2087–2104. [Google Scholar] [CrossRef] [Green Version]
- Rao, R.R.; Long, J.Z.; White, J.P.; Svensson, K.J.; Lou, J.; Lokurkar, I.; Jedrychowski, M.P.; Ruas, J.L.; Wrann, C.D.; Lo, J.C.; et al. Meteorin-like is a hormone that regulates immune-adipose interactions to increase beige fat thermogenesis. Cell 2014, 157, 1279–1291. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Kang, Y.E.; Kim, J.M.; Choung, S.; Joung, K.H.; Kim, H.J.; Ku, B.J. Serum Meteorin-like protein levels decreased in patients newly diagnosed with type 2 diabetes. Diabetes Res. Clin. Pract. 2018, 135, 7–10. [Google Scholar] [CrossRef]
- Dadmanesh, M.; Aghajani, H.; Fadaei, R.; Ghorban, K. Lower serum levels of Meteorin-like/Subfatin in patients with coronary artery disease and type 2 diabetes mellitus are negatively associated with insulin resistance and inflammatory cytokines. PLoS ONE 2018, 13, e0204180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.-X.; Ji, H.-H.; Yao, M.-P.; Wang, L.; Wang, Y.; Zhou, P.; Liu, Y.; Zheng, X.-F.; He, H.-W.; Wang, L.-S.; et al. Serum Metrnl is associated with the presence and severity of coronary artery disease. J. Cell. Mol. Med. 2019, 23, 271–280. [Google Scholar] [CrossRef] [PubMed]
- El-Ashmawy, H.M.; Selim, F.O.; Hosny, T.A.M.; Almassry, H.N. Association of low serum Meteorin like (Metrnl) concentrations with worsening of glucose tolerance, impaired endothelial function and atherosclerosis. Diabetes Res. Clin. Pract. 2019, 150, 57–63. [Google Scholar] [CrossRef]
- Xu, L.; Cai, Y.; Wang, Y.; Xu, C. Meteorin-Like (METRNL) Attenuates Myocardial Ischemia/Reperfusion Injury-Induced Cardiomyocytes Apoptosis by Alleviating Endoplasmic Reticulum Stress via Activation of AMPK-PAK2 Signaling in H9C2 Cells. Med. Sci. Monit. 2020, 26, e924564. [Google Scholar] [CrossRef]
- Fisher, F.M.; Maratos-Flier, E. Understanding the Physiology of FGF21. Annu. Rev. Physiol. 2016, 78, 223–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haberka, M.; Machnik, G.; Kowalówka, A.; Biedroń, M.; Skudrzyk, E.; Regulska-Ilow, B.; Gajos, G.; Manka, R.; Deja, M.; Okopień, B.; et al. Epicardial, paracardial and perivascular fat quantity, genes expression and serum cytokines in coronary artery disease and diabetes. Pol. Arch. Intern. Med. 2019, 129, 738–746. [Google Scholar] [CrossRef] [Green Version]
- Akyildiz, Z.I.; Polat, S.; Yurekli, B.S.; Kocabas, G.U.; Tuluce, K.; Tuluce, S.Y.; Kocabas, U.; Bozkaya, G.; Yuksel, A.; Nazli, C. Epicardial fat, body mass index, and triglyceride are independent contributors of serum fibroblast growth factor 21 level in obese premenopausal women. J. Endocrinol. Invest. 2015, 38, 361–366. [Google Scholar] [CrossRef] [PubMed]
- Kotulák, T.; Drápalová, J.; Kopecký, P.; Lacinová, Z.; Kramář, P.; Říha, H.; Netuka, I.; Malý, J.; Housa, D.; Bláha, J.; et al. Increased circulating and epicardial adipose tissue mRNA expression of fibroblast growth factor-21 after cardiac surgery: Possible role in postoperative inflammatory response and insulin resistance. Physiol. Res. 2011, 60, 757–767. [Google Scholar] [CrossRef]
- Wang, W.-F.; Li, S.-M.; Ren, G.-P.; Zheng, W.; Lu, Y.-J.; Yu, Y.-H.; Xu, W.-J.; Li, T.-H.; Zhou, L.-H.; Liu, Y.; et al. Recombinant murine fibroblast growth factor 21 ameliorates obesity-related inflammation in monosodium glutamate-induced obesity rats. Endocrine 2015, 49, 119–129. [Google Scholar] [CrossRef]
- Wang, N.; Zhao, T.-T.; Li, S.; Sun, X.; Li, Z.; Li, Y.; Li, D.-S.; Wang, W.-F. Fibroblast Growth Factor 21 Exerts its Anti-inflammatory Effects on Multiple Cell Types of Adipose Tissue in Obesity. Obesity 2019, 27, 399–408. [Google Scholar] [CrossRef]
- Keinicke, H.; Sun, G.; Mentzel, C.M.J.; Fredholm, M.; John, L.M.; Andersen, B.; Raun, K.; Kjaergaard, M. FGF21 regulates hepatic metabolic pathways to improve steatosis and inflammation. Endocr. Connect. 2020, 9, 755–768. [Google Scholar] [CrossRef]
- Singhal, G.; Fisher, F.M.; Chee, M.J.; Tan, T.G.; El Ouaamari, A.; Adams, A.C.; Najarian, R.; Kulkarni, R.N.; Benoist, C.; Flier, J.S.; et al. Fibroblast Growth Factor 21 (FGF21) Protects against High Fat Diet Induced Inflammation and Islet Hyperplasia in Pancreas. PLoS ONE 2016, 11, e0148252. [Google Scholar] [CrossRef]
- Gao, J.; Liu, Q.; Li, J.; Hu, C.; Zhao, W.; Ma, W.; Yao, M.; Xing, L. Fibroblast Growth Factor 21 dependent TLR4/MYD88/NF-κB signaling activation is involved in lipopolysaccharide-induced acute lung injury. Int. Immunopharmacol. 2020, 80, 106219. [Google Scholar] [CrossRef]
- Zhang, J.; Cheng, Y.; Gu, J.; Wang, S.; Zhou, S.; Wang, Y.; Tan, Y.; Feng, W.; Fu, Y.; Mellen, N.; et al. Fenofibrate increases cardiac autophagy via FGF21/SIRT1 and prevents fibrosis and inflammation in the hearts of Type 1 diabetic mice. Clin. Sci. 2016, 130, 625–641. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.G.; Yi, S.-A.; Choi, S.-E.; Kang, Y.; Kim, T.H.; Jeon, J.Y.; Bae, M.A.; Ahn, J.H.; Jeong, H.; Hwang, E.S.; et al. TM-25659-Induced Activation of FGF21 Level Decreases Insulin Resistance and Inflammation in Skeletal Muscle via GCN2 Pathways. Mol. Cells 2015, 38, 1037–1043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, N.; Li, J.-Y.; Li, S.; Guo, X.-C.; Wu, T.; Wang, W.-F.; Li, D.-S. Fibroblast growth factor 21 regulates foam cells formation and inflammatory response in Ox-LDL-induced THP-1 macrophages. Biomed. Pharmacother. 2018, 108, 1825–1834. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; He, J.; Li, S.; Song, L.; Guo, X.; Yao, W.; Zou, D.; Gao, X.; Liu, Y.; Bai, F.; et al. Fibroblast growth factor 21 (FGF21) inhibits macrophage-mediated inflammation by activating Nrf2 and suppressing the NF-κB signaling pathway. Int. Immunopharmacol. 2016, 38, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Zheng, Q.; Chen, J.; Tan, X.; Li, Q.; Ding, L.; Zhang, R.; Lin, X. FGF21 mitigates atherosclerosis via inhibition of NLRP3 inflammasome-mediated vascular endothelial cells pyroptosis. Exp. Cell Res. 2020, 393, 112108. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Gou, Z.; Li, Y.; Wang, Y.; Zhu, J.; Xu, G.; Zhang, Q. Fibroblast growth factor 21 inhibits atherosclerosis in apoE-/- mice by ameliorating Fas-mediated apoptosis. Lipids Health Dis. 2018, 17, 203. [Google Scholar] [CrossRef] [Green Version]
- Xiaolong, L.; Dongmin, G.; Liu, M.; Zuo, W.; Huijun, H.; Qiufen, T.; XueMei, H.; Wensheng, L.; Yuping, P.; Jun, L.; et al. FGF21 induces autophagy-mediated cholesterol efflux to inhibit atherogenesis via RACK1 up-regulation. J. Cell. Mol. Med. 2020, 24, 4992–5006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Qi, Y.-F.; Chang, J.-R.; Lu, W.-W.; Zhang, J.-S.; Wang, S.-P.; Cheng, S.-J.; Zhang, M.; Fan, Q.; Lv, Y.; et al. Possible role of fibroblast growth factor 21 on atherosclerosis via amelioration of endoplasmic reticulum stress-mediated apoptosis in apoE(-/-) mice. Heart Vessels 2015, 30, 657–668. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Pan, X.; Wu, F.; Ye, D.; Zhang, Y.; Wang, Y.; Jin, L.; Lian, Q.; Huang, Y.; Ding, H.; et al. Fibroblast Growth Factor 21 Prevents Atherosclerosis by Suppression of Hepatic Sterol Regulatory Element-Binding Protein-2 and Induction of Adiponectin in Mice. Circulation 2015, 131, 1861–1871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, H.; Cheng, J.; Zhou, Q.; Peng, J.; Pan, Y.; Han, H. Fibroblast growth factor 21 attenuates inflammation and oxidative stress in atherosclerotic rat via enhancing the Nrf1-ARE signaling pathway. Int. J. Clin. Exp. Pathol. 2018, 11, 1308–1317. [Google Scholar]
- Zhang, Y.; Liu, Z.; Zhou, M.; Liu, C. Therapeutic effects of fibroblast growth factor-21 against atherosclerosis via the NF-κB pathway. Mol. Med. Rep. 2018, 17, 1453–1460. [Google Scholar] [CrossRef] [PubMed]
- Ying, L.; Li, N.; He, Z.; Zeng, X.; Nan, Y.; Chen, J.; Miao, P.; Ying, Y.; Lin, W.; Zhao, X.; et al. Fibroblast growth factor 21 Ameliorates diabetes-induced endothelial dysfunction in mouse aorta via activation of the CaMKK2/AMPKα signaling pathway. Cell Death Dis. 2019, 10, 665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basurto, L.; Gregory, M.A.; Hernández, S.B.; Sánchez-Huerta, L.; Martínez, A.D.; Manuel-Apolinar, L.; Avelar, F.J.; Alonso, L.A.M.; Sánchez-Arenas, R. Monocyte chemoattractant protein-1 (MCP-1) and fibroblast growth factor-21 (FGF-21) as biomarkers of subclinical atherosclerosis in women. Exp. Gerontol. 2019, 124, 110624. [Google Scholar] [CrossRef] [PubMed]
- Yafei, S.; Elsewy, F.; Youssef, E.; Ayman, M.; El-Shafei, M. Fibroblast growth factor 21 association with subclinical atherosclerosis and arterial stiffness in type 2 diabetes. Diabetes Metab. Syndr. 2019, 13, 882–888. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Qian, L.; Zhang, L.; Zhang, J.; Zhou, J.; Li, Y.; Hou, X.; Fang, Q.; Li, H.; Jia, W. Fibroblast Growth Factor 21 is Related to Atherosclerosis Independent of Nonalcoholic Fatty Liver Disease and Predicts Atherosclerotic Cardiovascular Events. J. Am. Heart Assoc. 2020, 9, e015226. [Google Scholar] [CrossRef]
- Wang, X.; Huang, X.; Hou, J. Relationship between Serum fibroblast growth factor 21 levels and morphological atherosclerotic plaque characteristics in patients with coronary heart disease. Eur. Heart J. Suppl. 2016, 18, F37. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Y.; Liu, L.; Xu, A.; Zhou, P.; Long, Z.; Tu, Y.; Chen, X.; Tang, W.; Huang, G.; Zhou, Z. Serum fibroblast growth factor 21 levels are related to subclinical atherosclerosis in patients with type 2 diabetes. Cardiovasc. Diabetol. 2015, 14, 72. [Google Scholar] [CrossRef] [Green Version]
- Miyazaki, Y.; Saita, E.; Kishimoto, Y.; Ibe, S.; Seki, T.; Miura, K.; Suzuki-Sugihara, N.; Ikegami, Y.; Ohmori, R.; Kondo, K.; et al. Low Plasma Levels of Fibroblast Growth Factor-21 in Patients with Peripheral Artery Disease. J. Atheroscler. Thromb. 2018, 25, 821–828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.; Feng, A.; Lin, S.; Yu, L.; Lin, X.; Yan, X.; Lu, X.; Zhang, C. Fibroblast growth factor-21 prevents diabetic cardiomyopathy via AMPK-mediated antioxidation and lipid-lowering effects in the heart. Cell Death Dis. 2018, 9, 227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanajak, P.; Sa-Nguanmoo, P.; Wang, X.; Liang, G.; Li, X.; Jiang, C.; Chattipakorn, S.C.; Chattipakorn, N. Fibroblast growth factor 21 (FGF21) therapy attenuates left ventricular dysfunction and metabolic disturbance by improving FGF21 sensitivity, cardiac mitochondrial redox homoeostasis and structural changes in pre-diabetic rats. Acta Physiol. 2016, 217, 287–299. [Google Scholar] [CrossRef]
- Li, S.; Zhu, Z.; Xue, M.; Yi, X.; Liang, J.; Niu, C.; Chen, G.; Shen, Y.; Zhang, H.; Zheng, J.; et al. Fibroblast growth factor 21 protects the heart from angiotensin II-induced cardiac hypertrophy and dysfunction via SIRT1. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 1241–1252. [Google Scholar] [CrossRef] [PubMed]
- Planavila, A.; Redondo-Angulo, I.; Ribas, F.; Garrabou, G.; Casademont, J.; Giralt, M.; Villarroya, F. Fibroblast growth factor 21 protects the heart from oxidative stress. Cardiovasc. Res. 2015, 106, 19–31. [Google Scholar] [CrossRef] [Green Version]
- Planavila, A.; Redondo, I.; Hondares, E.; Vinciguerra, M.; Munts, C.; Iglesias, R.; Gabrielli, L.A.; Sitges, M.; Giralt, M.; van Bilsen, M.; et al. Fibroblast growth factor 21 protects against cardiac hypertrophy in mice. Nat. Commun. 2013, 4, 2019. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Xu, C.; Liu, Y.; Li, Y.; Du, S.; Zhang, R.; Sun, Y.; Zhang, R.; Wang, Y.; Xue, H.; et al. Fibroblast growth factor 21 inhibited ischemic arrhythmias via targeting miR-143/EGR1 axis. Basic Res. Cardiol. 2020, 115, 9. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feijóo-Bandín, S.; Aragón-Herrera, A.; Moraña-Fernández, S.; Anido-Varela, L.; Tarazón, E.; Roselló-Lletí, E.; Portolés, M.; Moscoso, I.; Gualillo, O.; González-Juanatey, J.R.; et al. Adipokines and Inflammation: Focus on Cardiovascular Diseases. Int. J. Mol. Sci. 2020, 21, 7711. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21207711
Feijóo-Bandín S, Aragón-Herrera A, Moraña-Fernández S, Anido-Varela L, Tarazón E, Roselló-Lletí E, Portolés M, Moscoso I, Gualillo O, González-Juanatey JR, et al. Adipokines and Inflammation: Focus on Cardiovascular Diseases. International Journal of Molecular Sciences. 2020; 21(20):7711. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21207711
Chicago/Turabian StyleFeijóo-Bandín, Sandra, Alana Aragón-Herrera, Sandra Moraña-Fernández, Laura Anido-Varela, Estefanía Tarazón, Esther Roselló-Lletí, Manuel Portolés, Isabel Moscoso, Oreste Gualillo, José Ramón González-Juanatey, and et al. 2020. "Adipokines and Inflammation: Focus on Cardiovascular Diseases" International Journal of Molecular Sciences 21, no. 20: 7711. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21207711