Erythrocyte, Platelet, Serum Ferritin, and P-Selectin Pathophysiology Implicated in Severe Hypercoagulation and Vascular Complications in COVID-19

 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Blood Proteins Results
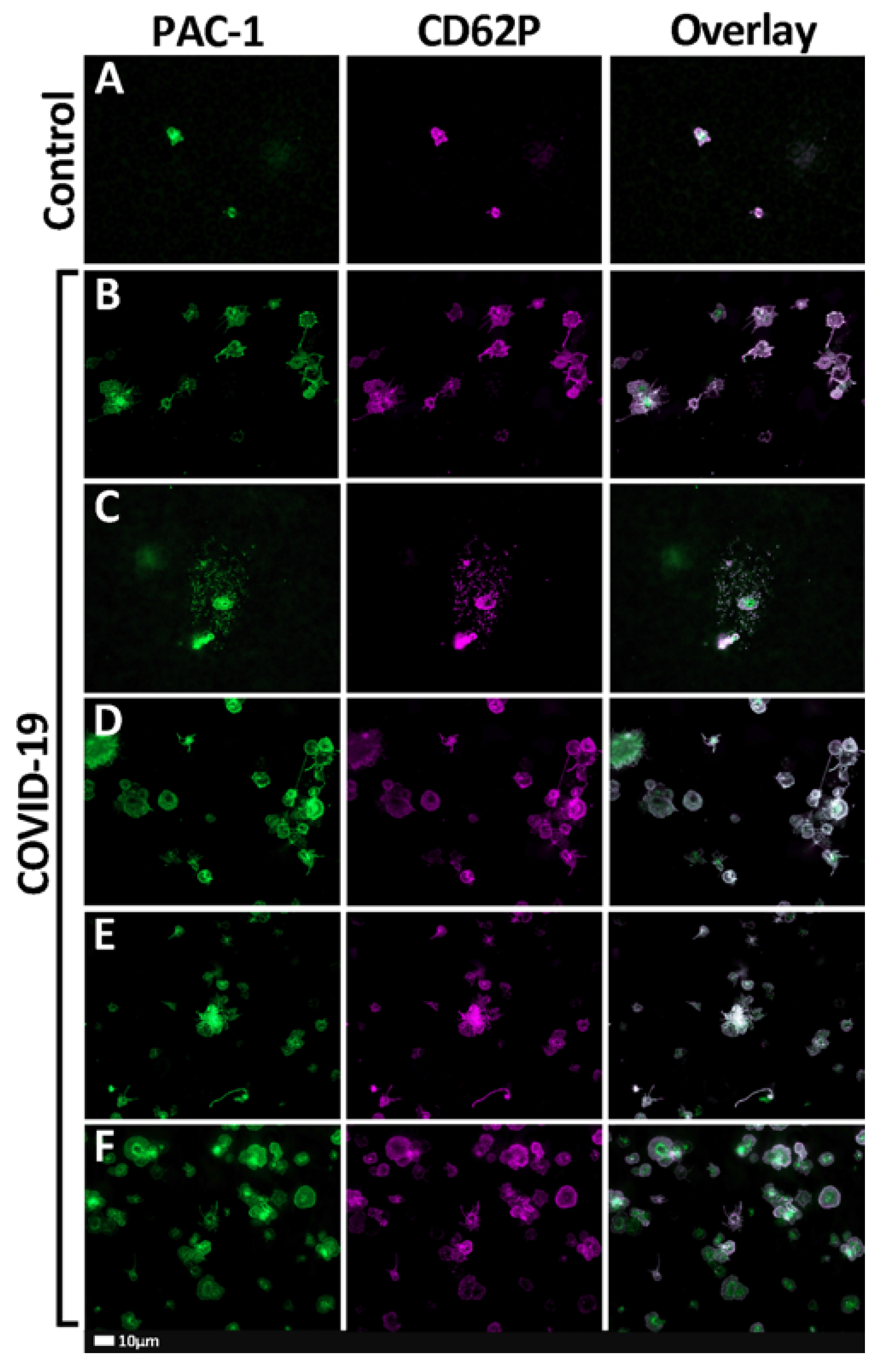
2.2. Fluorescent Microscopy Results
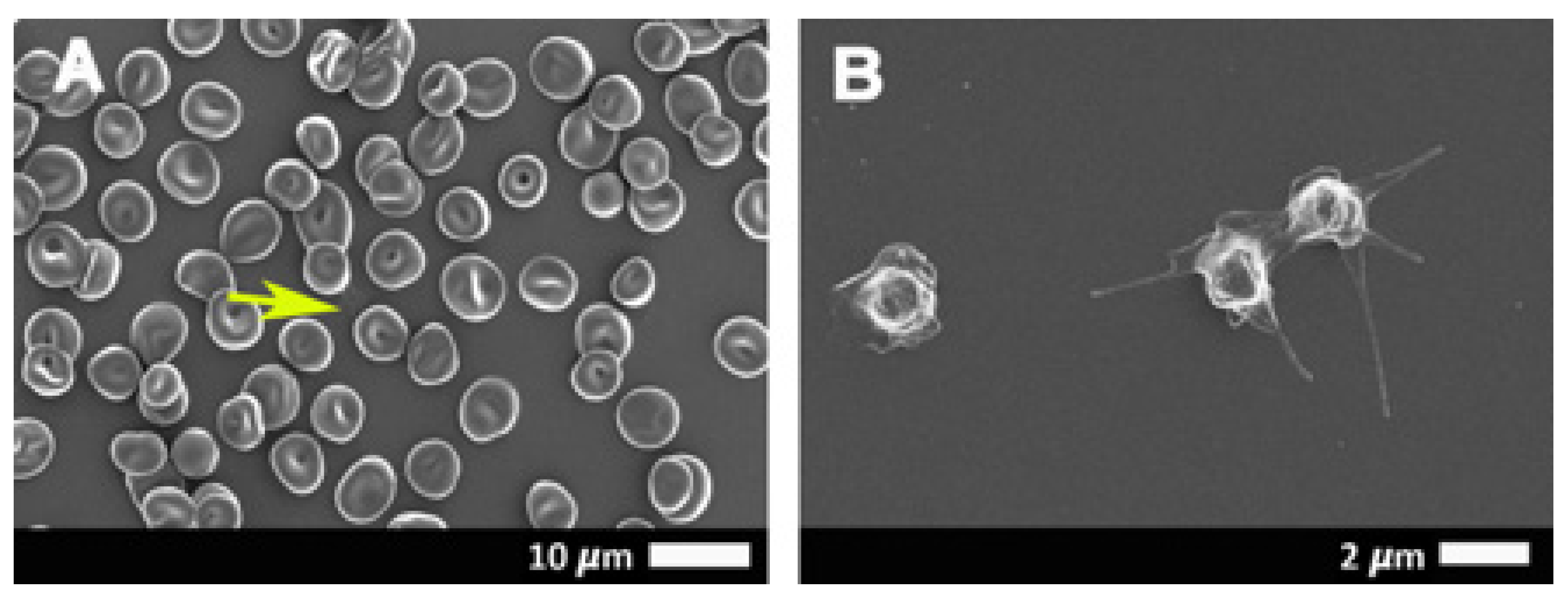
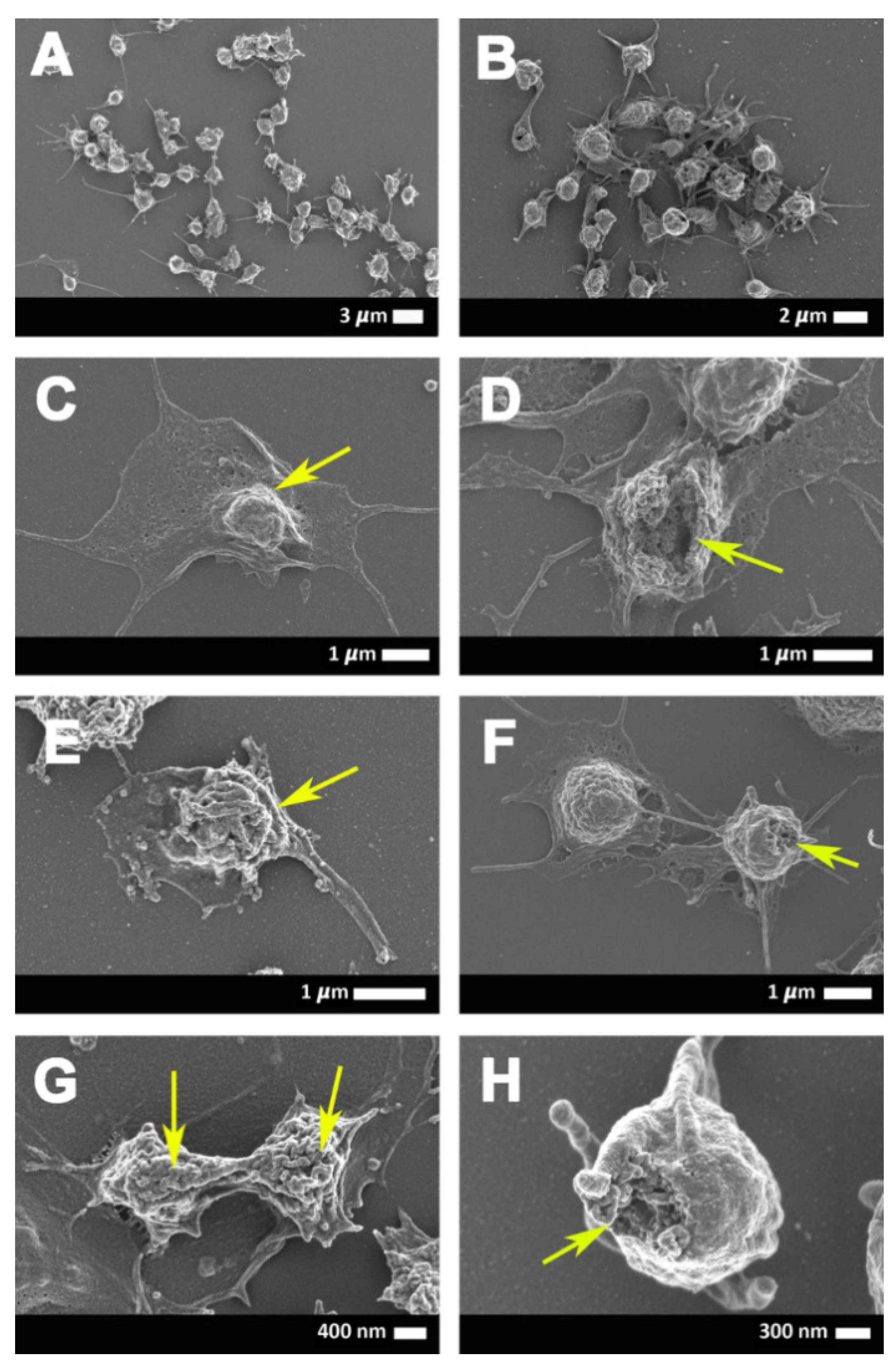
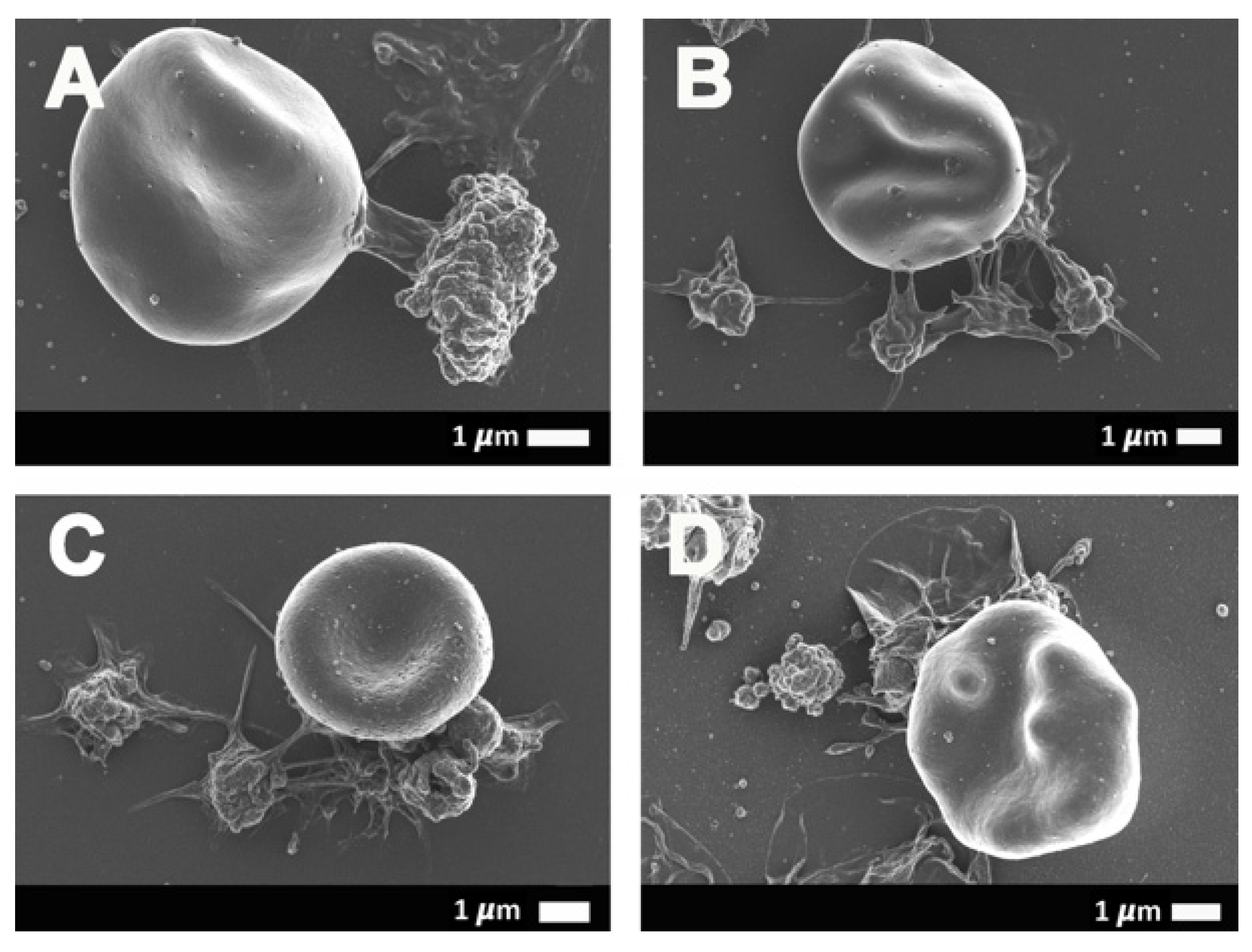
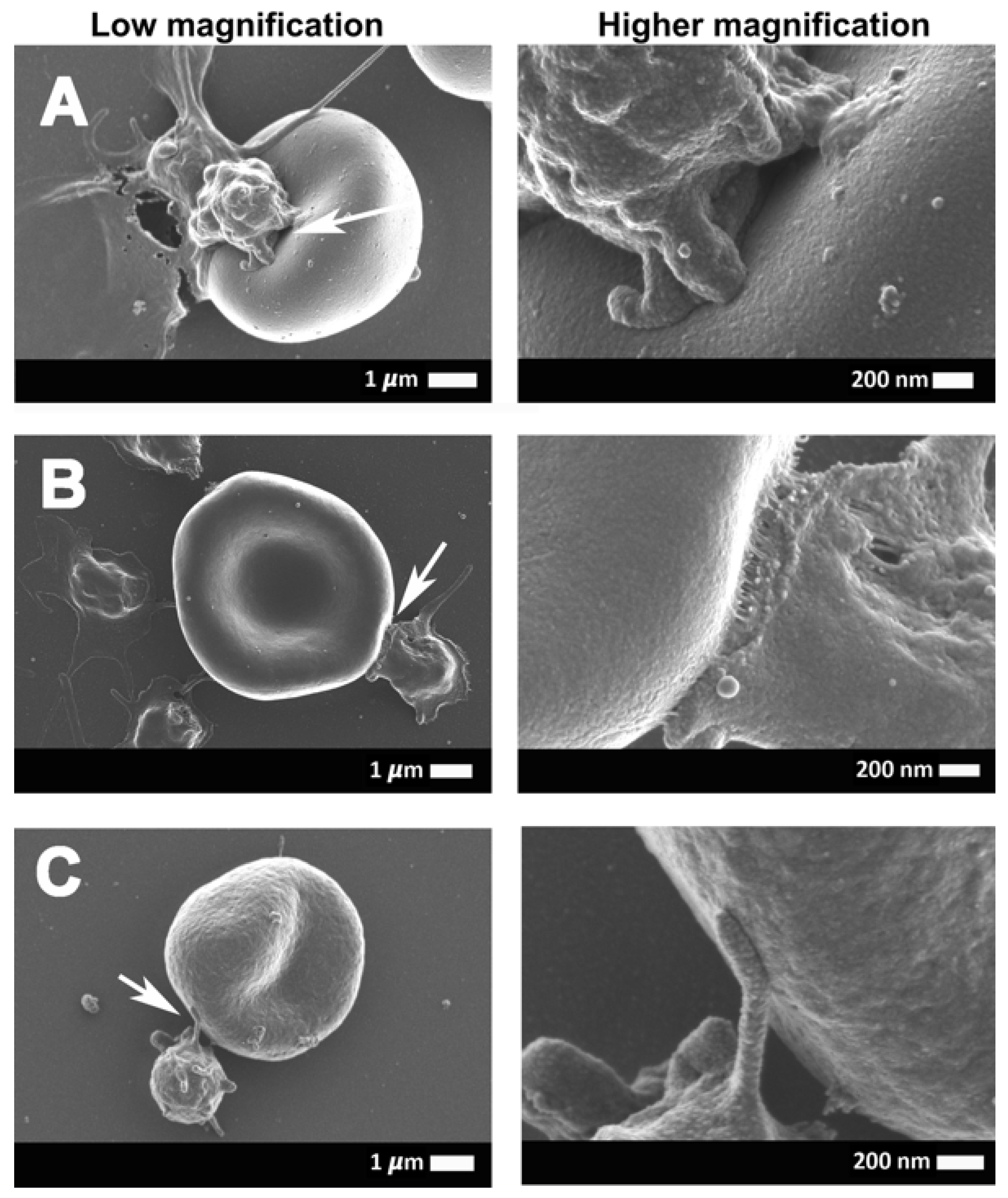
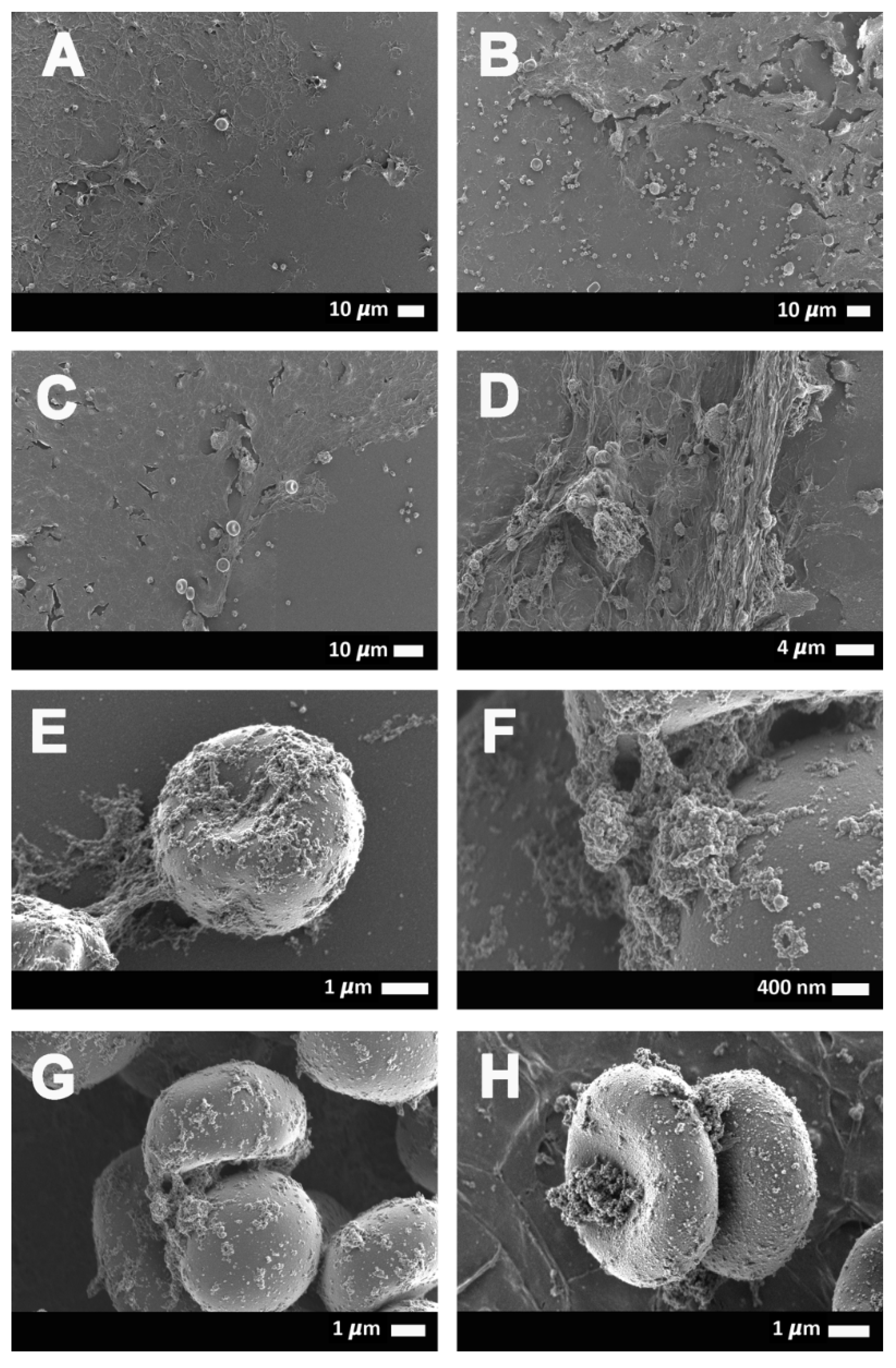
2.3. Scanning Electron Microscopy Results
2.4. Viscoelasticity and Viscometry Results
3. Discussion
4. Materials and Methods
4.1. Ethical Statement
4.2. Patient Samples
4.2.1. Covid-19 Patients
4.2.2. Healthy Samples
4.3. Fluorescence Microscopy
4.4. Scanning Electron Microscopy
4.5. P-Selectin ELISA, CRP, and Serum Ferritin
4.6. Thromboelastography (TEG®)
4.7. Viscometry
4.8. Statistics
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| COVID-19 | Coronavirus disease 2019 |
| sP-selectin | Soluble P-selectin |
| SEM | Scanning electron microscope |
| TEG® | Thromboelastography |
| R value | reaction time measured in minutes |
| Alpha-angle | (α, degrees) |
| MA | Maximal amplitude measured in mm |
| MRTG | Maximum rate of thrombus generation, measured in Dyn·cm−2·s−1 |
| TMRTG | Time to maximum rate of thrombus, measured in minutes generation |
| TTG | Total thrombus generation, measured in Dyn·cm−2 |
References
- Willyard, C. Coronavirus blood-clot mystery intensifies. Nature 2020, 581, 250. [Google Scholar] [CrossRef] [PubMed]
- Matacic, C. Blood vessel injury may spur disease’s fatal second phase. Science 2020, 368, 1039–1040. [Google Scholar] [CrossRef] [PubMed]
- Kander, T. Coagulation disorder in COVID-19. Lancet Haematol. 2020, 7, e630–e632. [Google Scholar] [CrossRef]
- Liao, D.; Zhou, F.; Luo, L.; Xu, M.; Wang, H.; Xia, J.; Gao, Y.; Cai, L.; Wang, Z.; Yin, P.; et al. Haematological characteristics and risk factors in the classification and prognosis evaluation of COVID-19: A retrospective cohort study. Lancet Haematol. 2020, 7, e671–e678. [Google Scholar] [CrossRef]
- Teuwen, L.A.; Geldhof, V.; Pasut, A.; Carmeliet, P. COVID-19: The vasculature unleashed. Nat. Rev. Immunol. 2020, 20, 389–391. [Google Scholar] [CrossRef] [PubMed]
- Yuriditsky, E.; Horowitz, J.M.; Merchan, C.; Ahuja, T.; Brosnahan, S.B.; McVoy, L.; Berger, J.S. Thromboelastography Profiles of Critically Ill Patients With Coronavirus Disease 2019. Crit. Care Med. 2020. [Google Scholar] [CrossRef]
- Thachil, J.; Tang, N.; Gando, S.; Falanga, A.; Cattaneo, M.; Levi, M.; Clark, C.; Iba, T. ISTH interim guidance on recognition and management of coagulopathy in COVID-19. J. Thromb. Haemost. 2020, 18, 1023–1026. [Google Scholar] [CrossRef]
- Levi, M.; Thachil, J.; Iba, T.; Levy, J.H. Coagulation abnormalities and thrombosis in patients with COVID-19. Lancet Haematol. 2020, 7, e438–e440. [Google Scholar] [CrossRef]
- Iba, T.; Levy, J.H.; Levi, M.; Thachil, J. Coagulopathy in COVID-19. J. Thromb. Haemost. 2020, 18, 2103–2109. [Google Scholar] [CrossRef]
- Levi, M.; Thachil, J. Coronavirus Disease 2019 Coagulopathy: Disseminated Intravascular Coagulation and Thrombotic Microangiopathy-Either, Neither, or Both. Semin. Thromb. Hemost. 2020, 46, 781–784. [Google Scholar] [CrossRef]
- Iba, T.; Levy, J.H.; Connors, J.M.; Warkentin, T.E.; Thachil, J.; Levi, M. The unique characteristics of COVID-19 coagulopathy. Crit. Care 2020, 24, 360. [Google Scholar] [CrossRef]
- Al-Samkari, H.; Leaf, R.S.K.; Dzik, W.H.; Carlson, J.C.T.; Fogerty, A.E.; Waheed, A.; Goodarzi, K.; Bendapudi, P.K.; Bornikova, L.; Gupta, S.; et al. COVID and Coagulation: Bleeding and Thrombotic Manifestations of SARS-CoV2 Infection. Blood 2020, 136, 489–500. [Google Scholar] [CrossRef] [PubMed]
- Dorgalaleh, A. Bleeding and Bleeding Risk in COVID-19. Semin. Thromb. Hemost. 2020, 46, 815–818. [Google Scholar] [CrossRef] [PubMed]
- Grobler, C.; Maphumulo, S.C.; Grobbelaar, L.M.; Bredenkamp, J.C.; Laubscher, G.J.; Lourens, P.J.; Currin, A.; Currin, A.; Pretorius, E. COVID-19: The Rollercoaster of Fibrin(ogen), D-dimer, von Willebrand Factor, P-selectin and Their Interactions with Endothelial Cells, Platelets and Erythrocytes. Int. J. Mol. Sci. 2020, 21, 5168. [Google Scholar] [CrossRef] [PubMed]
- Ackermann, M.; Verleden, S.E.; Kuehnel, M.; Haverich, A.; Welte, T.; Laenger, F.; Vanstapel, A.; Werlein, C.; Stark, H.; Tzankov, A.; et al. Pulmonary Vascular Endothelialitis, Thrombosis, and Angiogenesis in Covid-19. N. Engl. J. Med. 2020, 383, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Goshua, G.; Pine, A.B.; Meizlish, M.L.; Chang, C.H.; Zhang, H.; Bahel, P.; Baluha, A.; Bar, N.; Bona, R.D.; Burns, A.J.; et al. Endotheliopathy in COVID-19-associated coagulopathy: Evidence from a single-centre, cross-sectional study. Lancet Haematol. 2020, 7, e575–e582. [Google Scholar] [CrossRef]
- Pretorius, E.; Venter, C.; Laubscher, G.J.; Lourens, P.J.; Steenkamp, J.; Kell, D.B. Prevalence of amyloid blood clots in COVID-19 plasma. Accepted for publication in Cadiovas Diabetology.
- Kell, D.B.; Pretorius, E. Proteins behaving badly. Substoichiometric molecular control and amplification of the initiation and nature of amyloid fibril formation: Lessons from and for blood clotting. Prog. Biophys. Mol. Biol. 2017, 123, 16–41. [Google Scholar] [CrossRef]
- Kell, D.B.; Pretorius, E. To what extent are the terminal stages of sepsis, septic shock, SIRS, and multiple organ dysfunction syndrome actually driven by a toxic prion/amyloid form of fibrin? Semin. Thromb. Hemost. 2018, 44, 224–238. [Google Scholar]
- Pretorius, E.; Bester, J.; Page, M.J.; Kell, D.B. The potential of LPS-binding protein to reverse amyloid formation in plasma fibrin of individuals with Alzheimer-type dementia. Front. Aging Neurosci. 2018, 10, 257. [Google Scholar] [CrossRef]
- Pretorius, E.; Mbotwe, S.; Bester, J.; Robinson, C.J.; Kell, D.B. Acute induction of anomalous and amyloidogenic blood clotting by molecular amplification of highly substoichiometric levels of bacterial lipopolysaccharide. J. R. Soc. Interface 2016, 123, 20160539. [Google Scholar] [CrossRef] [Green Version]
- Pretorius, E.; Page, M.J.; Engelbrecht, L.; Ellis, G.C.; Kell, D.B. Substantial fibrin amyloidogenesis in type 2 diabetes assessed using amyloid-selective fluorescent stains. Cardiovasc. Diabetol. 2017, 16, 141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pretorius, E.; Page, M.J.; Hendricks, L.; Nkosi, N.B.; Benson, S.R.; Kell, D.B. Both lipopolysaccharide and lipoteichoic acids potently induce anomalous fibrin amyloid formation: Assessment with novel Amytracker™ stains. bioRxiv preprint. J. R. Soc. Interface 2018, 15, 20170941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neri, T.; Nieri, D.; Celi, A. P-selectin blockade in COVID-19-related ARDS. Am. J. Physiol. Lung Cell. Mol. Physiol. 2020, 318, L1237–L1238. [Google Scholar] [CrossRef] [PubMed]
- Blann, A.D.; Nadar, S.K.; Lip, G.Y.H. The adhesion molecule P-selectin and cardiovascular disease. Eur. Heart J. 2003, 24, 2166–2179. [Google Scholar] [CrossRef] [Green Version]
- Lorant, D.E.; Topham, M.K.; E Whatley, R.; McEver, R.P.; McIntyre, T.M.; Prescott, S.M.; A Zimmerman, G. Inflammatory roles of P-selectin. J. Clin. Investig. 1993, 92, 559–570. [Google Scholar] [CrossRef] [Green Version]
- Cambien, B.; Wagner, D.D. A new role in hemostasis for the adhesion receptor P-selectin. Trends Mol. Med. 2004, 10, 179–186. [Google Scholar] [CrossRef]
- Manne, B.K.; Denorme, F.; Middleton, E.A.; Portier, I.; Rowley, J.W.; Stubben, C.J.; Petrey, A.C.; Tolley, N.D.; Guo, L.; Cody, M.; et al. Platelet Gene Expression and Function in COVID-19 Patients. Blood 2020, 136, 1317–1329. [Google Scholar] [CrossRef]
- Kell, D.B. Iron behaving badly: Inappropriate iron chelation as a major contributor to the aetiology of vascular and other progressive inflammatory and degenerative diseases. BMC Med. Genom. 2009, 2, 2. [Google Scholar] [CrossRef]
- Pretorius, E.; Vermeulen, N.; Bester, J.; Lipinski, B.; Kell, D.B. A novel method for assessing the role of iron and its functional chelation in fibrin fibril formation: The use of scanning electron microscopy. Toxicol. Mech. Methods 2013, 23, 352–359. [Google Scholar] [CrossRef]
- Kell, D.B.; Pretorius, E. Serum ferritin is an important disease marker, and is mainly a leakage product from damaged cells. Metallomics 2014, 6, 748–773. [Google Scholar] [CrossRef] [Green Version]
- De Villiers, S.; Bester, J.; Kell, D.B.; Pretorius, E. Erythrocyte health and the possible role of amyloidogenic blood clotting in the evolving haemodynamics of female migraine-with-aura pathophysiology: Results from a pilot study. Front. Neurol. 2019, 10, 1262. [Google Scholar] [CrossRef]
- Pretorius, E. The adaptability of red blood cells. Cardiovasc. Diabetol. 2013, 12, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lipinski, B.; Pretorius, E.; Oberholzer, H.M.; van der Spuy, W.J. Interaction of fibrin with red blood cells: The role of iron. Ultrastruct. Pathol. 2012, 36, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Du Plooy, J.N.; Bester, J.; Pretorius, E. Eryptosis in Haemochromatosis: Implications for rheology. Clin. Hemorheol. Microcirc. 2018, 69, 457–469. [Google Scholar] [CrossRef]
- Lipinski, B.; Pretorius, E. Novel pathway of ironinduced blood coagulation: Implications for diabetes mellitus and its complications. Pol. Arch. Med. Wewn. 2012, 122, 115–122. [Google Scholar] [PubMed]
- Olumuyiwa-Akeredolu, O.O.; Page, M.J.; Soma, P.; Pretorius, E. Platelets: Emerging facilitators of cellular crosstalk in rheumatoid arthritis. Nat. Rev. Rheumatol. 2019, 15, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Van Rooy, M.J.; Duim, W.; Ehlers, R.; Buys, A.V.; Pretorius, E. Platelet hyperactivity and fibrin clot structure in transient ischemic attack individuals in the presence of metabolic syndrome: A microscopy and thromboelastography study. Cardiovasc. Diabetol. 2015, 14, 86. [Google Scholar] [CrossRef] [Green Version]
- Soma, P.; Swanepoel, A.C.; du Plooy, J.N.; Mqoco, T.; Pretorius, E. Flow cytometric analysis of platelets type 2 diabetes mellitus reveals ‘angry’ platelets. Cardiovasc. Diabetol. 2016, 15, 52. [Google Scholar] [CrossRef] [Green Version]
- Kell, D.B. Towards a unifying, systems biology understanding of large-scale cellular death and destruction caused by poorly liganded iron: Parkinson’s, Huntington’s, Alzheimer’s, prions, bactericides, chemical toxicology and others as examples. Arch. Toxicol. 2010, 84, 825–889. [Google Scholar] [CrossRef] [Green Version]
- Sproston, N.R.; Ashworth, J.J. Role of C-Reactive Protein at Sites of Inflammation and Infection. Front. Immunol. 2018, 9, 754. [Google Scholar] [CrossRef]
- Quantikine. [ELISA catalog]. 2020. Available online: https://resources.rndsystems.com/pdfs/datasheets/dpse00.pdf (accessed on 30 September 2020).
- Lu, Q.; Malinauskas, R.A. Comparison of two platelet activation markers using flow cytometry after in vitro shear stress exposure of whole human blood. Artif. Organs 2011, 35, 137–144. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Demographics | |||
|---|---|---|---|
| p-value (data normally distributed; unpaired t-test) | 0.6 | ||
| Mean age of healthy individuals (n = 13) | 55.6 (±10.7) | ||
| Mean age of COVID-19 (n = 37) | 53.1 (±14.7) | ||
| Serum Ferritin (µg·mL−1) (COVID-19: n = 33; controls: n = 13) | |||
| p-value (Mann-Whitney U test, unpaired nonparametric data expressed as median) (Q1–Q3) | 0.0009 (***) | ||
| Median of healthy individuals | 113.5 (55.8–149.3) | ||
| Median of COVID-19 patients | 306.6 (162.0–699.0) | ||
| CRP (µg·mL−1) (COVID-19: n = 37; controls: n = 13) | |||
| p-value (Mann-Whitney U test, unpaired nonparametric data expressed as median) (Q1–Q3) | <0.0001 (***) | ||
| Median of healthy individuals | 0.8 (0.3–2.1) | ||
| Median of COVID-19 patients | 44.1 (13.2–108.0) | ||
| Soluble P-selectin (ng·mL−1) (COVID-19: n = 30; controls: n = 10) | |||
| Our P-selectin (ng·mL−1) analysis | Goshua et al., 2020 analysis (ng·mL−1) | ||
| p-value | 0.0009 (***) | p-value | 0.001 |
| Median of healthy individuals (n = 10) | 26.7 (23.9–27.2) | Median of healthy individuals (n = 13) | 9.5 (8.5–11.3) |
| Median of COVID-19 patients (n = 30) | 17.36 (14.3–22.3) | Median of ICU COVID-19 patients (n = 48) | 15.9 (4.8) |
| Median of non-ICU COVID-19 patients (n = 20) | 11.2 (3.1) | ||
| Our ELISA kit average ranges for control data | 15–55 (Citrate and EDTA plasma) | Their ELISA kit insert average for control data | 25.8 (citrate plasma) and 18.3 to 57.4 (EDTA plasma) |
| Platelet-poor plasma viscosity (mPa·second) | |||
| p-value (data normally distributed; unpaired t-test) (control: n = 10; COVID-19: n = 7) | 0.007 (**) | ||
| Mean viscosity of healthy individuals | 2.0 (±0.2) | ||
| Mean viscosity of COVID-19 samples | 2.8 (±0.8) | ||
| Thromboelastography® of platelet-poor plasma (PPP) | |||
| Parameter | Plasma from healthy samples (n = 10) | Plasma from COVID-19 samples (n = 7) | p-value |
| R | 16.41 (±6.1) | 12.9 (±5.2) | 0.1 |
| α angle | 52.4 (34.4–61.3) | 46.0 (33.9–63.3) | 0.7 |
| MA | 32.1 (24.5–36.0) | 48.2 (35.0–53.8) | 0.04 (*) |
| MRTG | 4.7 (±3.0) | 7.5 (±3) | 0.04 (*) |
| TMRTG | 17.6 (13.1–23.6) | 13.2 (11.3–18.4) | 0.4 |
| TTG | 237 (162.6–415.6) | 577.7 (422.0–642.3) | 0.009 (**) |
| TEG® Parameters | Explanation |
|---|---|
| R value: reaction time measured in minutes | Time of latency from start of test to initial fibrin formation (amplitude of 2 mm); i.e., initiation time. |
| Alpha-angle (α, degrees) | The rate of fibrin crosslinking indicated by degrees. |
| MA: maximal amplitude measured in mm | Maximum clot size: it reflects the ultimate strength of the fibrin clot, i.e., overall stability of the clot. The larger the MA, the more hypercoagulable the clot. |
| Maximum rate of thrombus generation (MRTG) measured in Dyn·cm−2·s−1 | The maximum velocity of clot growth observed or maximum rate of thrombus generation using G, where G is the elastic modulus strength of the thrombus in dynes per cm−2. |
| Time to maximum rate of thrombus generation (TMRTG) measured in minutes | The time interval observed before the maximum speed of the clot growth. |
| Total thrombus generation (TTG) measured in Dyn·cm−2 | The clot strength: the amount of total resistance (to movement of the cup and pin) generated during clot formation. This is the total area under the velocity curve during clot growth, representing the amount of clot strength generated during clot growth. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Venter, C.; Bezuidenhout, J.A.; Laubscher, G.J.; Lourens, P.J.; Steenkamp, J.; Kell, D.B.; Pretorius, E. Erythrocyte, Platelet, Serum Ferritin, and P-Selectin Pathophysiology Implicated in Severe Hypercoagulation and Vascular Complications in COVID-19. Int. J. Mol. Sci. 2020, 21, 8234. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21218234
Venter C, Bezuidenhout JA, Laubscher GJ, Lourens PJ, Steenkamp J, Kell DB, Pretorius E. Erythrocyte, Platelet, Serum Ferritin, and P-Selectin Pathophysiology Implicated in Severe Hypercoagulation and Vascular Complications in COVID-19. International Journal of Molecular Sciences. 2020; 21(21):8234. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21218234
Chicago/Turabian StyleVenter, Chantelle, Johannes Andries Bezuidenhout, Gert Jacobus Laubscher, Petrus Johannes Lourens, Janami Steenkamp, Douglas B. Kell, and Etheresia Pretorius. 2020. "Erythrocyte, Platelet, Serum Ferritin, and P-Selectin Pathophysiology Implicated in Severe Hypercoagulation and Vascular Complications in COVID-19" International Journal of Molecular Sciences 21, no. 21: 8234. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21218234