Genetic Carriers and Genomic Distribution of cadA6—A Novel Variant of a Cadmium Resistance Determinant Identified in Listeria spp.

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Isolation and Taxonomic Position of Listeria spp. Isolates
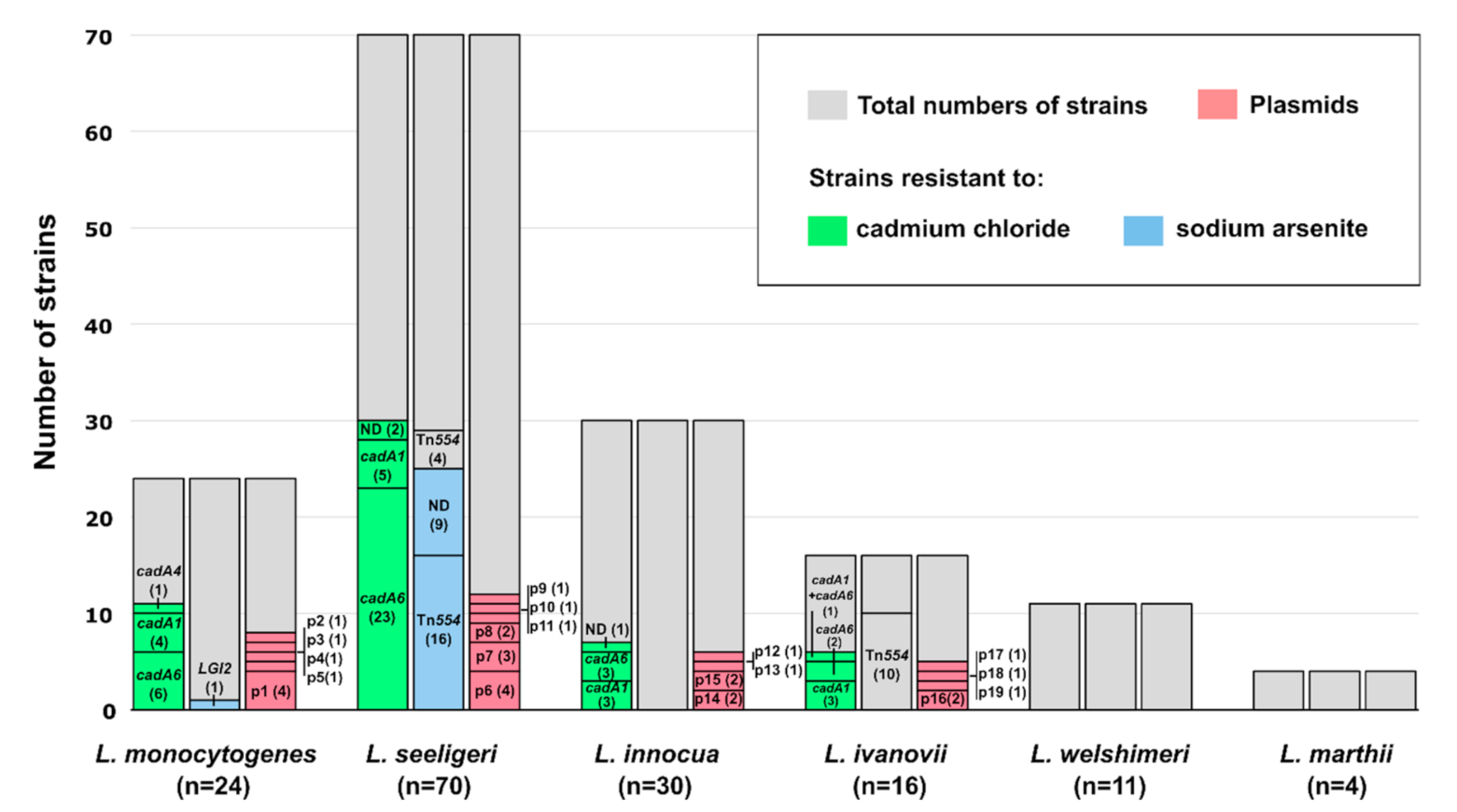
2.2. Susceptibility of Listeria spp. Strains to Heavy Metals and Metalloids
2.3. Plasmid Occurrence and Distribution in Listeria spp. Strains
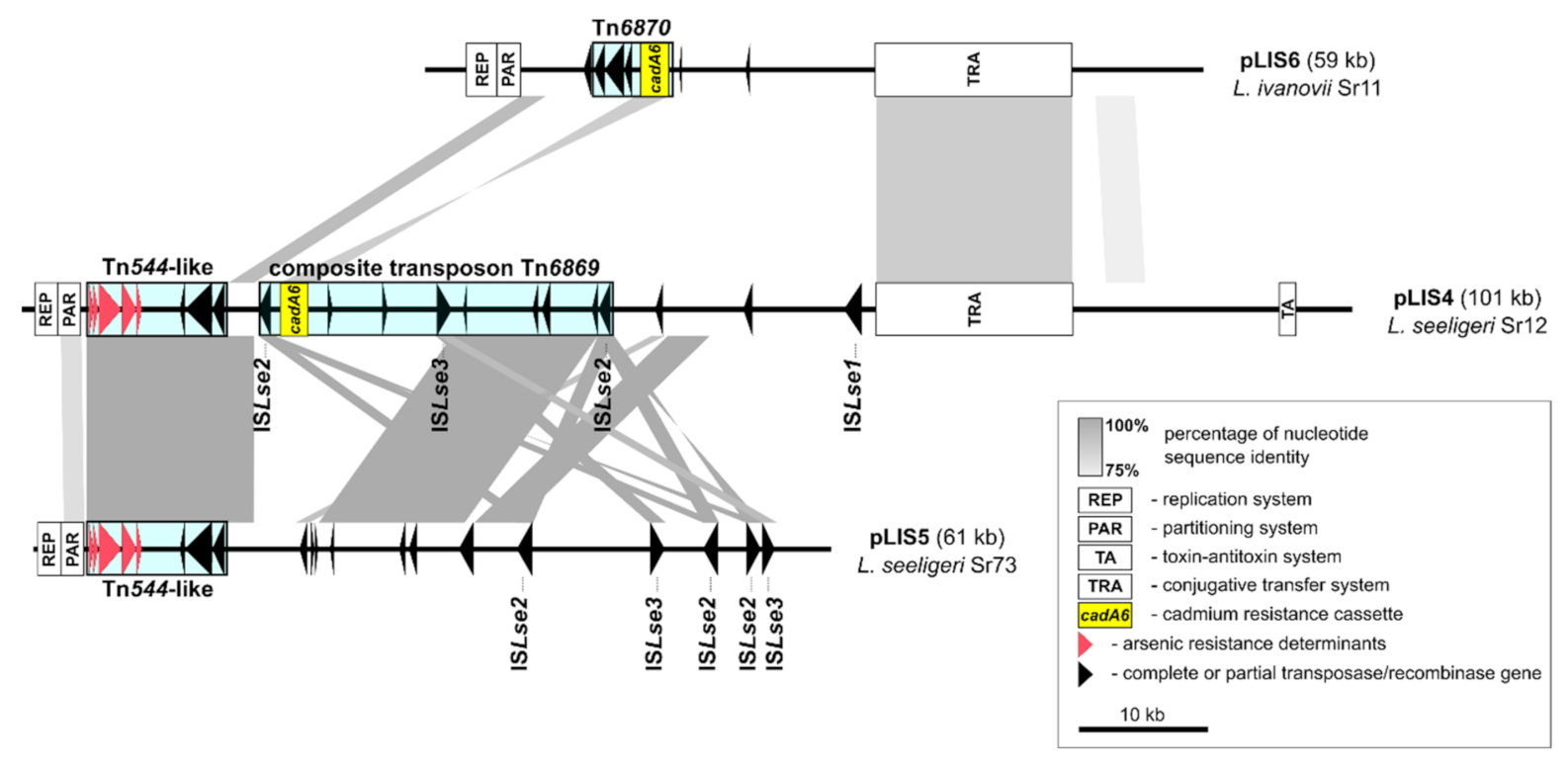
2.4. Nucleotide Sequences of L. seeligeri and L. ivanovii Replicons Containing Resistance Determinants
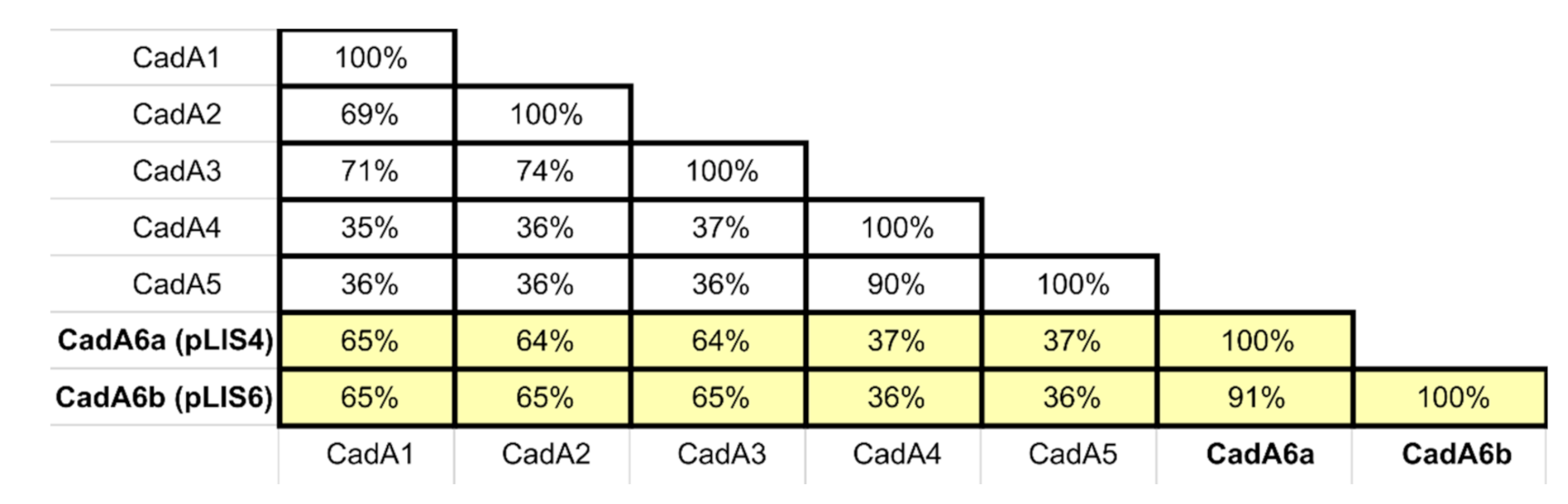
2.5. Identification of cadA6—A Novel Cadmium Resistance Cassette Variant
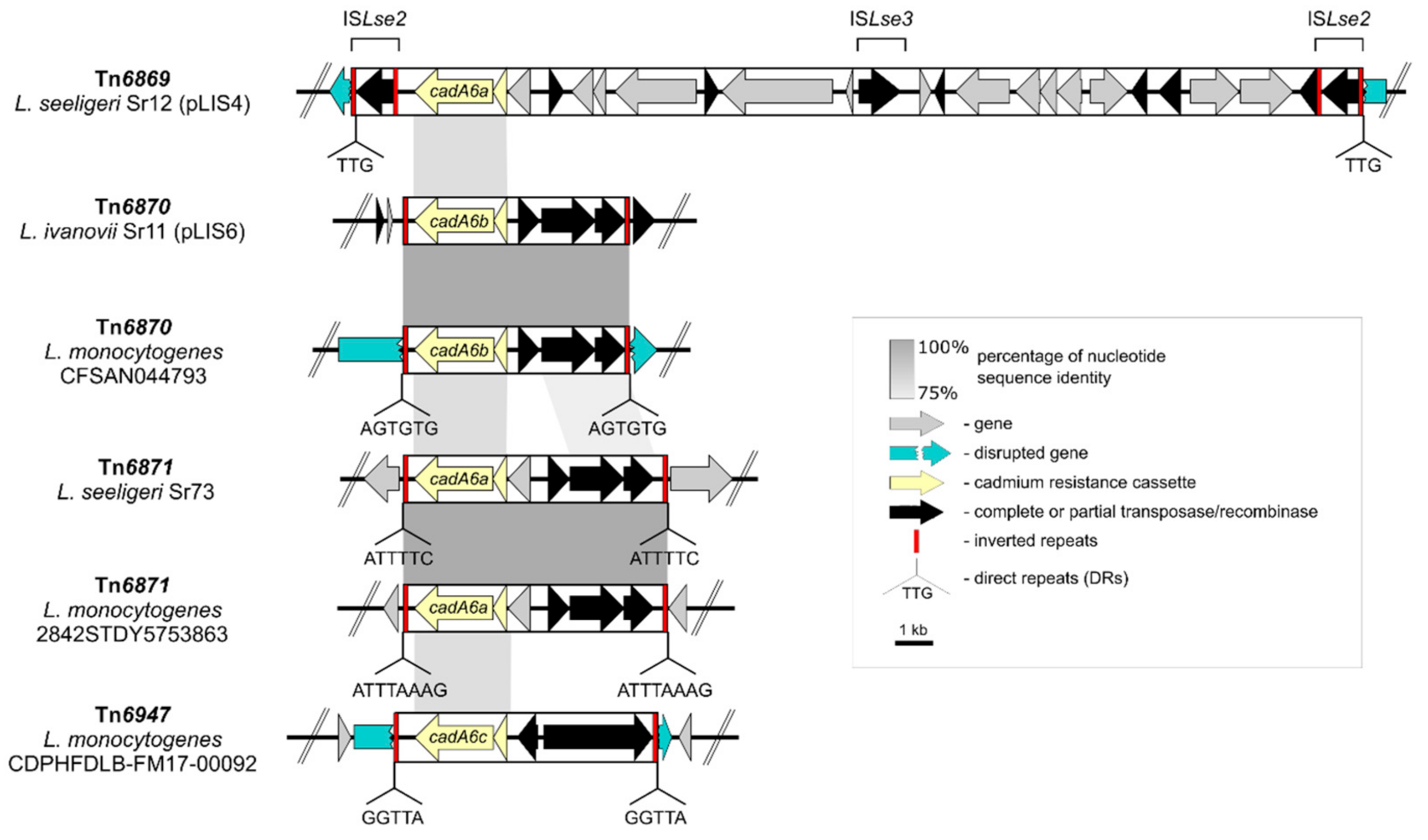
2.6. Transposons Harboring the cadA6 Efflux System
2.7. Functional Analysis of the cadA6 Resistance Cassette
3. Materials and Methods
3.1. Bacterial Strains and Growth Conditions
3.2. Soil and Water Sample Collection and Isolation of Listeria spp.
3.3. Identification of Listeria Species
3.4. Bacterial Mating between Listeria spp. Strains
3.5. Heavy-Metal and Metalloid Susceptibility
3.6. Isolation of Genomic DNA
3.7. Plasmid DNA Isolation and Restriction Fragment Length Polymorphism (RFLP) Profile Analysis
3.8. Plasmid Curing
3.9. Detection of Resistance Determinants by PCR
3.10. Functional Analysis of the cadA6 Resistance Cassette
3.10.1. Construction of a Mobilizable Shuttle Vector pDKEL
3.10.2. Cloning of the Cadmium Resistance Cassette into pDKEL
3.10.3. Introduction of Constructed Plasmids into L. monocytogenes 10403S
3.10.4. Heavy-Metal Susceptibility of L. monocytogenes 10403S Strains Harboring the Constructed Plasmids
3.11. DNA Sequencing
3.12. Bioinformatic Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Leclercq, A.; Moura, A.; Vales, G.; Tessaud-Rita, N.; Aguilhon, C.; Lecuit, M. Listeria thailandensis sp. nov. Int. J. Syst. Evol. Microbiol. 2019, 69, 74–81. [Google Scholar] [CrossRef]
- Girardin, H.; Morris, C.E.; Albagnac, C.; Dreux, N.; Glaux, C.; Nguyen-The, C. Behaviour of the pathogen surrogates Listeria innocua and Clostridium sporogenes during production of parsley in fields fertilized with contaminated amendments. FEMS Microbiol. Ecol. 2005, 54, 287–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaya, P.; Saralegui, C.; Medina, M.; Nuñez, M. Occurrence of Listeria monocytogenes and other Listeria spp. in raw caprine milk. J. Dairy Sci. 1996, 79, 1936–1941. [Google Scholar] [CrossRef]
- Hofer, E.; Ribeiro, R.; Feitosa, D.P. Species and serovars of the genus Listeria isolated from different sources in Brazil from 1971 to 1997. Mem. Inst. Oswaldo Cruz 2000, 95, 615–620. [Google Scholar] [CrossRef]
- Kalorey, D.R.; Warke, S.R.; Kurkure, N.V.; Rawool, D.B.; Barbuddhe, S.B. Listeria species in bovine raw milk: A large survey of Central India. Food Control 2008, 19, 109–112. [Google Scholar] [CrossRef]
- MacGowan, A.P.; Bowker, K.; McLauchlin, J.; Bennett, P.M.; Reeves, D.S. The occurrence and seasonal changes in the isolation of Listeria spp. in shop bought food stuffs, human faeces, sewage and soil from urban sources. Int. J. Food Microbiol. 1994, 21, 325–334. [Google Scholar] [CrossRef]
- Meloni, D.; Galluzzo, P.; Mureddu, A.; Piras, F.; Griffiths, M.; Mazzette, R. Listeria monocytogenes in RTE foods marketed in Italy: Prevalence and automated EcoRI ribotyping of the isolates. Int. J. Food Microbiol. 2009, 129, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Wilson, I.G. Occurrence of Listeria species in ready to eat foods. Epidemiol. Infect. 1995, 115, 519–526. [Google Scholar] [CrossRef] [Green Version]
- Pesavento, G.; Ducci, B.; Nieri, D.; Comodo, N.; Lo Nostro, A. Prevalence and antibiotic susceptibility of Listeria spp. isolated from raw meat and retail foods. Food Control 2010, 21, 708–713. [Google Scholar] [CrossRef]
- Sheridan, J.J.; Duffy, G.; McDowell, D.A.; Blair, I.S. The occurence and initial numbers of Listeria in Irish meat and fish products and the recovery of injured cells from frozen products. Int. J. Food Microbiol. 1994, 22, 105–113. [Google Scholar] [CrossRef]
- Bauwensm, L.; Vercammen, F.; De Meurichy, W. Occurrence of Listeria Spp. in captive antelope herds and their environment. J. Zoo Wildl. Med. 2001, 32, 514–518. [Google Scholar]
- Linke, K.; Rückerl, I.; Brugger, K.; Karpiskova, R.; Walland, J.; Muri-Klinger, S.; Tichy, A.; Wagner, M.; Stessl, B. Reservoirs of Listeria species in three environmental ecosystems. Appl. Environ. Microbiol. 2014, 80, 5583–5592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sauders, B.D.; Overdevest, J.; Fortes, E.; Windham, K.; Schukken, Y.; Lembo, A.; Wiedmann, M. Diversity of Listeria species in urban and natural environments. Appl. Environ. Microbiol. 2012, 78, 4420–4433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.; Rakic-Martinez, M.; Graves, L.M.; Ward, T.J.; Siletzky, R.M.; Kathariou, S. Genetic determinants for cadmium and arsenic resistance among Listeria monocytogenes serotype 4B isolates from sporadic human listeriosis patients. Appl. Environ. Microbiol. 2013, 79, 2471–2476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.; Ward, T.J.; Jima, D.D.; Parsons, C.; Kathariou, S. The arsenic resistance-associated Listeria genomic island LGI2 exhibits sequence and integration site diversity and a propensity for three Listeria monocytogenes clones with enhanced virulence. Appl. Environ. Microbiol. 2017, 83, e01189-17. [Google Scholar] [CrossRef] [Green Version]
- Mullapudi, S.; Siletzky, R.M.; Kathariou, S. Diverse cadmium resistance determinants in Listeria monocytogenes isolates from the turkey processing plant environment. Appl. Environ. Microbiol. 2010, 76, 627–630. [Google Scholar] [CrossRef] [Green Version]
- Ratani, S.S.; Siletzky, R.M.; Dutta, V.; Yildirim, S.; Osborne, J.A.; Lin, W.; Hitchins, A.D.; Ward, T.J.; Kathariou, S. Heavy metal and disinfectant resistance of Listeria monocytogenes from foods and food processing plants. Appl. Environ. Microbiol. 2012, 78, 6938–6945. [Google Scholar] [CrossRef] [Green Version]
- Korsak, D.; Chmielowska, C.; Szuplewska, M.; Bartosik, D. Prevalence of plasmid-borne benzalkonium chloride resistance cassette bcrABC and cadmium resistance cadA genes in nonpathogenic Listeria spp. isolated from food and food-processing environments. Int. J. Food Microbiol. 2019, 290, 247–253. [Google Scholar] [CrossRef]
- Korsak, D.; Szuplewska, M. Characterization of nonpathogenic Listeria species isolated from food and food processing environment. Int. J. Food Microbiol. 2016, 238, 274–280. [Google Scholar] [CrossRef]
- Katharios-Lanwermeyer, S.; Rakic-Martinez, M.; Elhanafi, D.; Ratani, S.; Tiedje, J.M.; Kathariou, S. Coselection of cadmium and benzalkonium chloride resistance in conjugative transfers from nonpathogenic Listeria spp. to other Listeriae. Appl. Environ. Microbiol. 2012, 78, 7549–7556. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.; Deng, Y.; Fan, R.; Shi, L.; Bai, J.; Yan, H. Coresistance to benzalkonium chloride disinfectant and heavy metal ions in Listeria monocytogenes and Listeria innocua swine isolates from China. Foodborne Pathog. Dis. 2019, 16, 696–703. [Google Scholar] [CrossRef] [PubMed]
- Mullapudi, S.; Siletzky, R.M.; Kathariou, S. Heavy-metal and benzalkonium chloride resistance of Listeria monocytogenes isolates from the environment of Turkey-processing plants. Appl. Environ. Microbiol. 2008, 74, 1464–1468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuenne, C.; Voget, S.; Pischimarov, J.; Oehm, S.; Goesmann, A.; Daniel, R.; Hain, T.; Chakraborty, T. Comparative analysis of plasmids in the genus Listeria. PLoS ONE 2010, 5, e12511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lebrun, M.; Audurier, A.; Cossart, P. Plasmid-borne cadmium resistance genes in Listeria monocytogenes are similar to cadA and cadC of Staphylococcus aureus and are induced by cadmium. J. Bacteriol. 1994, 176, 3040–3048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lebrun, M.; Loulergue, J.; Chaslus-Dancla, E.; Audurier, A. Plasmids in Listeria monocytogenes in relation to cadmium resistance. Appl. Environ. Microbiol. 1992, 58, 3183–3186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dutta, V.; Elhanafi, D.; Kathariou, S. Conservation and distribution of the benzalkonium chloride resistance cassette bcrABC in Listeria monocytogenes. Appl. Environ. Microbiol. 2013, 79, 6067–6074. [Google Scholar] [CrossRef] [Green Version]
- Elhanafi, D.; Dutta, V.; Kathariou, S. Genetic characterization of plasmid-associated benzalkonium chloride resistance determinants in a Listeria monocytogenes strain from the 1998–1999 outbreak. Appl. Environ. Microbiol. 2010, 76, 8231–8238. [Google Scholar] [CrossRef] [Green Version]
- Kuenne, C.; Billion, A.; Mraheil, M.A.; Strittmatter, A.; Daniel, R.; Goesmann, A.; Barbuddhe, S.; Hain, T.; Chakraborty, T. Reassessment of the Listeria monocytogenes pan-genome reveals dynamic integration hotspots and mobile genetic elements as major components of the accessory genome. BMC Genom. 2013, 14, 47. [Google Scholar] [CrossRef] [Green Version]
- Parsons, C.; Lee, S.; Kathariou, S. Heavy metal resistance determinants of the foodborne pathogen Listeria monocytogenes. Genes 2018, 10, 11. [Google Scholar] [CrossRef] [Green Version]
- Parsons, C.; Lee, S.; Jayeola, V.; Kathariou, S. Novel cadmium resistance determinant in Listeria monocytogenes. Appl. Environ. Microbiol. 2017, 83, e02580-16. [Google Scholar] [CrossRef] [Green Version]
- Kubier, A.; Wilkin, R.T.; Pichler, T. Cadmium in soils and groundwater: A review. Appl. Geochem. 2019, 108, 104388. [Google Scholar] [CrossRef] [PubMed]
- Tóth, G.; Hermann, T.; Da Silva, M.R.; Montanarella, L. Heavy metals in agricultural soils of the European Union with implications for food safety. Environ. Int. 2016, 88, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Cai, K.; Yu, Y.; Zhang, M.; Kim, K. Concentration, source, and total health risks of cadmium in multiple media in densely populated areas, China. Int. J. Environ. Res. Public Health 2019, 16, 2269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Wu, Q.; Hu, W.; Huang, B.; Dong, L.; Liu, G. Using multi-medium factors analysis to assess heavy metal health risks along the Yangtze River in Nanjing, Southeast China. Environ. Pollut. 2018, 243, 1047–1056. [Google Scholar] [CrossRef] [PubMed]
- Parsons, C.; Lee, S.; Kathariou, S. Dissemination and conservation of cadmium and arsenic resistance determinants in Listeria and other Gram-positive bacteria. Mol. Microbiol. 2020, 113, 560–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pombinho, R.; Camejo, A.; Vieira, A.; Reis, O.; Carvalho, F.; Almeida, M.T.; Pinheiro, J.C.; Sousa, S.; Cabanes, D. Listeria monocytogenes CadC regulates cadmium efflux and fine-tunes lipoprotein localization to escape the host immune response and promote infection. J. Infect. Dis. 2017, 215, 1468–1479. [Google Scholar] [CrossRef]
- Hingston, P.; Brenner, T.; Truelstrup Hansen, L.; Wang, S. Comparative analysis of Listeria monocytogenes plasmids and expression levels of plasmid-encoded genes during growth under salt and acid stress conditions. Toxins 2019, 11, 426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corbett, D.; Schuler, S.; Glenn, S.; Andrew, P.W.; Cavet, J.S.; Roberts, I.S. The combined actions of the copper-responsive repressor CsoR and copper-metallochaperone CopZ modulate CopA-mediated copper efflux in the intracellular pathogen Listeria monocytogenes. Mol. Microbiol. 2011, 81, 457–472. [Google Scholar] [CrossRef]
- Bell, F.Y. Copper Tolerance of Listeria Monocytogenes Strain DRDC8. Ph.D. Thesis, University of Adelaide, Adelaide, Australia, 2010. [Google Scholar]
- Francis, M.S.; Thomas, C.J. Mutants in the CtpA copper transporting P-type ATPase reduce virulence of Listeria monocytogenes. Microb. Pathog. 1997, 22, 67–78. [Google Scholar] [CrossRef]
- Parsons, C.; Costolo, B.; Brown, P.; Kathariou, S. Penicillin-binding protein encoded by pbp4 is involved in mediating copper stress in Listeria monocytogenes. FEMS Microbiol. Lett. 2017, 364. [Google Scholar] [CrossRef]
- Kremer, P.H.C.; Lees, J.A.; Koopmans, M.M.; Ferwerda, B.; Arends, A.W.M.; Feller, M.M.; Schipper, K.; Valls Seron, M.; van der Ende, A.; Brouwer, M.C.; et al. Benzalkonium tolerance genes and outcome in Listeria monocytogenes meningitis. Clin. Microbiol. Infect. 2017, 23, 265.e1–265.e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charpentier, E.; Courvalin, P. Emergence of the trimethoprim resistance gene dfrD in Listeria monocytogenes BM4293. Antimicrob. Agents Chemother. 1997, 41, 1134–1136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poyart-Salmeron, C.; Carlier, C.; Trieu-Cuot, P.; Courvalin, P.; Courtieu, A.L. Transferable plasmid-mediated antibiotic resistance in Listeria monocytogenes. Lancet 1990, 335, 1422–1426. [Google Scholar] [CrossRef]
- Bertsch, D.; Anderegg, J.; Lacroix, C.; Meile, L.; Stevens, M.J.A. PDB2011, a 7.6kb multidrug resistance plasmid from Listeria innocua replicating in Gram-positive and Gram-negative hosts. Plasmid 2013, 70, 284–287. [Google Scholar] [CrossRef] [PubMed]
- Pöntinen, A.; Aalto-Araneda, M.; Lindström, M.; Korkeala, H. Heat resistance mediated by pLM58 plasmid-borne ClpL in Listeria monocytogenes. mSphere 2017, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naditz, A.L.; Dzieciol, M.; Wagner, M.; Schmitz-Esser, S. Plasmids contribute to food processing environment–associated stress survival in three Listeria monocytogenes ST121, ST8, and ST5 strains. Int. J. Food Microbiol. 2019, 299, 39–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLauchlin, J.; Hampton, M.D.; Shah, S.; Threlfall, E.J.; Wieneke, A.A.; Curtis, G.D.W. Subtyping of Listeria monocytogenes on the basis of plasmid profiles and arsenic and cadmium susceptibility. J. Appl. Microbiol. 1997, 83, 381–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dusza-Dobek, A. Geochemical studies of soils in some city parks of Warsaw. Biul. Państw. Inst. Geol. 2012, 450, 35–46, (In Polish with an English summary). [Google Scholar]
- Krogulec, E.; Wierchowiec, J.; Bojakowska, I.; Kwecko, P.; Tomassi-Morawiec, H.; Hrybowicz, G. Objaśnienia do Mapy geośrodowiskowej Polski 1:50 000 Arkusz Kampinos (521). Additional Information to a Geo-Environmental Map of Poland 1:50 000. Sheet Kampinos (521). Available online: http://bazadata.pgi.gov.pl/data/mgsp/txt/mgsp0521.pdf (accessed on 10 October 2020).
- Rutkowski, P.; Diatta, J.; Konatowska, M.; Andrzejewska, A.; Tyburski, Ł.; Przybylski, P. Geochemical referencing of natural forest contamination in Poland. Forests 2020, 11, 157. [Google Scholar] [CrossRef] [Green Version]
- Chmielowska, C.; Korsak, D.; Szuplewska, M.; Grzelecka, M.; Maćkiw, E.; Stasiak, M.; Macion, A.; Skowron, K.; Bartosik, D. Benzalkonium chloride and heavy metal resistance profiles of Listeria monocytogenes strains isolated from fish, fish products and food-producing factories in Poland. Food Microbiol. 2020. in review. [Google Scholar]
- Fox, E.M.; Allnutt, T.; Bradbury, M.I.; Fanning, S.; Chandry, P.S. Comparative genomics of the Listeria monocytogenes ST204 subgroup. Front. Microbiol. 2016, 7, 2057. [Google Scholar] [CrossRef] [Green Version]
- Schmitz-Esser, S.; Müller, A.; Stessl, B.; Wagner, M. Genomes of sequence type 121 Listeria monocytogenes strains harbor highly conserved plasmids and prophages. Front. Microbiol. 2015, 6, 380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- den Bakker, H.C.; Bowen, B.M.; Rodriguez-Rivera, L.D.; Wiedmann, M. FSL J1-208, a virulent uncommon phylogenetic lineage IV Listeria monocytogenes strain with a small chromosome size and a putative virulence plasmid carrying internalin-like genes. Appl. Environ. Microbiol. 2012, 78, 1876–1889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Platt, R.; Drescher, C.; Park, S.-K.; Phillips, G.J. Genetic system for reversible integration of DNA constructs and lacZ gene fusions into the Escherichia coli chromosome. Plasmid 2000, 43, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Bishop, D.K.; Hinrichs, D.J. Adoptive transfer of immunity to Listeria monocytogenes. The influence of in vitro stimulation on lymphocyte subset requirements. J. Immunol. 1987, 139, 2005–2009. [Google Scholar]
- Huang, B.; Eglezos, S.; Heron, B.A.; Smith, H.; Graham, T.; Bates, J.; Savill, J. Comparison of multiplex PCR with conventional biochemical methods for the identification of Listeria spp. isolates from food and clinical samples in Queensland, Australia. J. Food Prot. 2007, 70, 1874–1880. [Google Scholar] [CrossRef]
- Lane, D. 16S/23S rRNA Sequencing. In Nucleic Acid Techniques in Bacterial Systematics; Stackebrandt, E., Goodfellow, M., Eds.; John Wiley and Sons: New York, NY, USA, 1991; pp. 115–176. [Google Scholar]
- Sambrook, J.; Green, M. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2012. [Google Scholar]
- Anderson, D.G.; McKay, L.L. Simple and rapid method for isolating large plasmid DNA from lactic streptococci. Appl. Environ. Microbiol. 1983, 46, 549–552. [Google Scholar] [CrossRef] [Green Version]
- Margolles, A.; De Los Reyes-Gavilán, C.G. Characterization of plasmids from Listeria monocytogenes and Listeria innocua strains isolated from short-ripened cheeses. Int. J. Food Microbiol. 1998, 39, 231–236. [Google Scholar] [CrossRef]
- Glaser, P.; Frangeul, L.; Buchrieser, C.; Rusniok, C.; Amend, A.; Baquero, F.; Berche, P.; Bloecker, H.; Brandt, P.; Chakraborty, T.; et al. Comparative genomics of Listeria species. Science 2001, 294, 849–852. [Google Scholar]
- Philippe, N.; Alcaraz, J.P.; Coursange, E.; Geiselmann, J.; Schneider, D. Improvement of pCVD442, a suicide plasmid for gene allele exchange in bacteria. Plasmid 2004, 51, 246–255. [Google Scholar] [CrossRef]
- Dziewit, L.; Adamczuk, M.; Szuplewska, M.; Bartosik, D. DIY series of genetic cassettes useful in construction of versatile vectors specific for Alphaproteobacteria. J. Microbiol. Methods 2011, 86, 166–174. [Google Scholar] [CrossRef] [PubMed]
- Haima, P.; Bron, S.; Venema, G. The effect of restriction on shotgun cloning and plasmid stability in Bacillus subtilis Marburg. Mol. Gen. Genet. MGG 1987, 209, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Kovach, M.E.; Elzer, P.H.; Hill, D.S.; Robertson, G.T.; Farris, M.A.; Roop, R.M.; Peterson, K.M. Four new derivatives of the broad-host-range cloning vector pBBR1MCS, carrying different antibiotic-resistance cassettes. Gene 1995, 166, 175–176. [Google Scholar] [CrossRef]
- Ditta, G.; Stanfield, S.; Corbin, D.; Helinski, D.R. Broad host range DNA cloning system for Gram-negative bacteria: Construction of a gene bank of Rhizobium meliloti. Proc. Natl. Acad. Sci. USA 1980, 77, 7347–7351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wattam, A.R.; Davis, J.J.; Assaf, R.; Boisvert, S.; Brettin, T.; Bun, C.; Conrad, N.; Dietrich, E.M.; Disz, T.; Gabbard, J.L.; et al. Improvements to PATRIC, the all-bacterial Bioinformatics Database and Analysis Resource Center. Nucleic Acids Res. 2017, 45, D535–D542. [Google Scholar] [CrossRef] [PubMed]
- Rutherford, K.; Parkhill, J.; Crook, J.; Horsnell, T.; Rice, P.; Rajandream, M.-A.; Barrell, B. Artemis: Sequence visualization and annotation. Bioinformatics 2000, 16, 944–945. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, M.J.; Petty, N.K.; Beatson, S.A. Easyfig: A genome comparison visualizer. Bioinformatics 2011, 27, 1009–1010. [Google Scholar] [CrossRef]
- Siguier, P.; Perochon, J.; Lestrade, L.; Mahillon, J.; Chandler, M. ISfinder: The reference centre for bacterial insertion sequences. Nucleic Acids Res. 2006, 34, D32–D36. [Google Scholar] [CrossRef] [Green Version]
- Tansirichaiya, S.; Rahman, M.A.; Roberts, A.P. The Transposon Registry. Mob. DNA 2019, 10, 40. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.A.; Clamp, M.; Barton, G.J. Jalview Version 2--a multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bal, N.; Wu, C.C.; Catty, P.; Guillain, F.; Mintz, E. Cd2+ and the N-terminal metal-binding domain protect the putative membranous CPC motif of the Cd2+-ATPase of Listeria monocytogenes. Biochem. J. 2003, 369, 681–685. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′→3′) | Annealing [Tm, °C ] | Amplified DNA Fragment | Reference | |
|---|---|---|---|---|---|
| Targeted Gene (Original Source) | Size [bp] | ||||
| Identification of Resistance Determinants by PCR | |||||
| cadA1-F cadA1-R | CAGAGCACTTTACTGACCATCAATCGTT CTTCTTCATTTAACGTTCCAGCAAAAA | 53 | cadA1 (Tn5422) | 594 | [16] |
| cadA2-F cadA2-R | ACAAGTTAGATCAAAAGAGTCTTTTATT ATCTTCTTCATTTAGTGTTCCTGCAAAT | 53 | cadA2 (plM80) | 590 | [16] |
| cadA3-F cadA3-R | GCGATGATTGATAATGTCGATTACAAAT TGGTAATTTCTTTAAGTCATCTCCCATT | 52 | cadA3 (ICELm1 of EGD-e strain) | 468 | [16] |
| cadA4-F cadA4-R | GCATACGTACGAACCAGAAG CAGTGTTTCTGCTTTTGCTCC | 51 | cadA4 (genomic island LGI2) | 1135 | [14] |
| cadA5-F cadA5-R | GTGATTGTTAATAGGCAGGAAG GTGGTATACCCAACATTG | 47 | cadA5 (genomic island LGI2-1) | 1178 | [52] |
| cadA6-F cadA6-R | ACTTGTACAAACTGTGCATC AGTAATGTTGCTCCCATATCG | 51 | cadA6 (pLIS4, pLIS6) | 2027 | This study |
| arsA1-F arsA1-R | CAACTTTGACCCTGTGGAG CTTTCCATTCAATCACTGCG | 50 | arsA1 (genomic island LGI2) | 1466 | [14] |
| arsA2-F arsA2-R | CAACCAGATCAGTTACCATTAAC TGCTTCTCCAGAGATTTCTTCTG | 50 | arsA2 (genomic island LGI2) | 1710 | [14] |
| arsATn554F arsATn554R | TAACCAATAAGCCAACACCG CTTCTTTCCACTTCCCGAGC | 64 | arsA (Tn554–like transposon) | 1219 | [15] |
| FB001 LM2004 | CACACTGTAATGTTAACTGG TCCAAGTGCTGAGAATATAC | 47 | ctpA (plasmid-associated) | 396 | [39] |
| Cloning of cadA6 Genes | |||||
| cadA6aF cadA6aR | GCGACTAGTGCGCCCTCTAATGGAATAAC ATGCGGCCGCTGCTTCTCGCTTCTTTGC | 49 | cadA6a and cadC6a cassette (pLIS4, strain Sr12) | 2843 | This study |
| cadA6bF cadA6bR | CGCGACTAGTTCTCCAGGTCTCAATTTGTC AAGCGGCCGCCCACAAAAGGTGCTTAGTTC | 49 | cadA6b and cadC6b cassette (pLIS6 strain Sr11) | 3072 | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chmielowska, C.; Korsak, D.; Szmulkowska, B.; Krop, A.; Lipka, K.; Krupińska, M.; Bartosik, D. Genetic Carriers and Genomic Distribution of cadA6—A Novel Variant of a Cadmium Resistance Determinant Identified in Listeria spp. Int. J. Mol. Sci. 2020, 21, 8713. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21228713
Chmielowska C, Korsak D, Szmulkowska B, Krop A, Lipka K, Krupińska M, Bartosik D. Genetic Carriers and Genomic Distribution of cadA6—A Novel Variant of a Cadmium Resistance Determinant Identified in Listeria spp. International Journal of Molecular Sciences. 2020; 21(22):8713. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21228713
Chicago/Turabian StyleChmielowska, Cora, Dorota Korsak, Barbara Szmulkowska, Alicja Krop, Kinga Lipka, Martyna Krupińska, and Dariusz Bartosik. 2020. "Genetic Carriers and Genomic Distribution of cadA6—A Novel Variant of a Cadmium Resistance Determinant Identified in Listeria spp." International Journal of Molecular Sciences 21, no. 22: 8713. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21228713