New Evidence on the Role of D-Aspartate Metabolism in Regulating Brain and Endocrine System Physiology: From Preclinical Observations to Clinical Applications

 , ,
, , 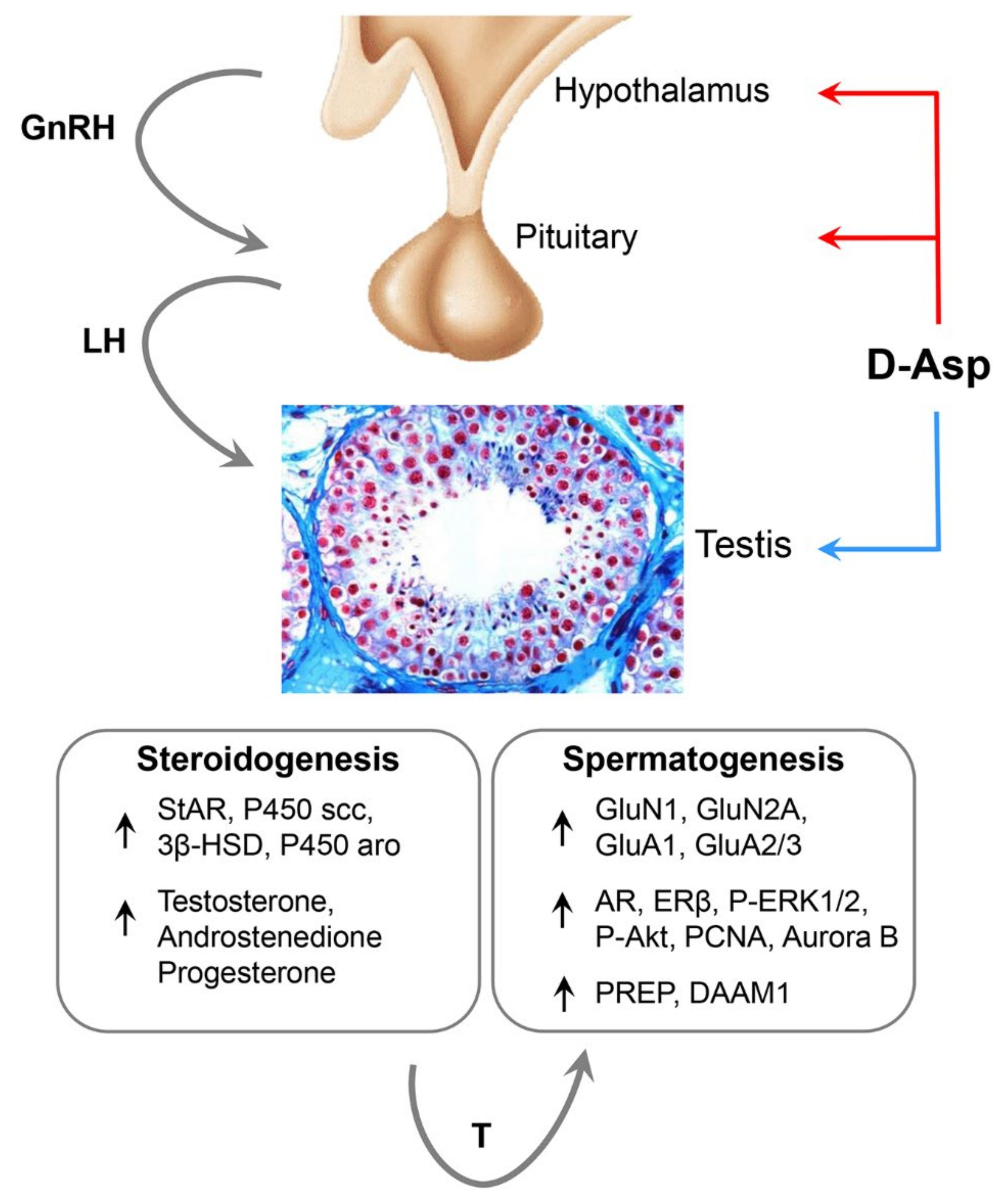{kind=link}
Abstract
:1. Introduction
2. Free D-Aspartate Distribution in the Mammalian Central Nervous System
3. Pharmacological Features of D-Aspartate
4. Animal Models with Altered D-Aspartate Metabolism
5. D-Aspartate Metabolism Alteration in Neurological and Psychiatric Disorders
6. Free D-Aspartate Distribution in Neuroendocrine and Endocrine Systems
7. D-Asp Administration on the Hypothalamus-Pituitary-Testis Axis
8. D-Asp Supplementation in Humans
9. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Genchi, G. An overview on d-amino acids. Amino Acids 2017, 49, 1521–1533. [Google Scholar] [CrossRef] [PubMed]
- D’Aniello, G.; Grieco, N.; Di Filippo, M.; Cappiello, F.; Topo, E.; D’Aniello, E.; Ronsini, S. Reproductive implication of D-aspartic acid in human pre-ovulatory follicular fluid. Hum. Reprod. 2007, 22, 3178–3183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleck, M.W.; Barrionuevo, G.; Palmer, A.M. Synaptosomal and vesicular accumulation of l-glutamate, l-aspartate and d-aspartate. Neurochem. Int. 2001, 39, 217–225. [Google Scholar] [CrossRef]
- Bastings, J.J.; Van Eijk, H.M.; Damink, S.O.; Rensen, S.S. d-amino Acids in Health and Disease: A Focus on Cancer. Nutrients 2019, 11, 2205. [Google Scholar] [CrossRef] [Green Version]
- D’Aniello, A.; Vetere, A.; Petrucelli, L. Further study on the specificity of d-amino acid oxidase and of d-aspartate oxidase and time course for complete oxidation of d-amino acids. Comp. Biochem. Physiol. Part B: Comp. Biochem. 1993, 105, 731–734. [Google Scholar] [CrossRef]
- Di Fiore, M.M.; Santillo, A.; Chieffi Baccari, G. Current knowledge of d-aspartate in glandular tissues. Amino Acids 2014, 46, 1805–1818. [Google Scholar] [CrossRef]
- Katane, M.; Homma, H. D-Aspartate Oxidase: The Sole Catabolic Enzyme Acting on Free D-Aspartate in Mammals. Chem. Biodivers. 2010, 7, 1435–1449. [Google Scholar] [CrossRef]
- Katane, M.; Kawata, T.; Nakayama, K.; Saitoh, Y.; Kaneko, Y.; Matsuda, S.; Saitoh, Y.; Miyamoto, T.; Sekine, M.; Homma, H. Characterization of the Enzymatic and Structural Properties of Human D-Aspartate Oxidase and Comparison with Those of the Rat and Mouse Enzymes. Biol. Pharm. Bull. 2015, 38, 298–305. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, S. d-Aspartate oxidase: Distribution, functions, properties, and biotechnological applications. Appl. Microbiol. Biotechnol. 2020, 104, 2883–2895. [Google Scholar] [CrossRef]
- Hashimoto, A.; Kumashiro, S.; Nishikawa, T.; Oka, T.; Takahashi, K.; Mito, T.; Takashima, S.; Doi, N.; Mizutani, Y.; Yamazaki, T.; et al. Embryonic Development and Postnatal Changes in Free d-Aspartate and d-Serine in the Human Prefrontal Cortex. J. Neurochem. 1993, 61, 348–351. [Google Scholar] [CrossRef]
- Sakai, K.; Homma, H.; Lee, J.-A.; Fukushima, T.; Santa, T.; Tashiro, K.; Iwatsubo, T.; Imai, K. Emergence of d-aspartic acid in the differentiating neurons of the rat central nervous system. Brain Res. 1998, 808, 65–71. [Google Scholar] [CrossRef]
- Hashimoto, A.; Oka, T. Free d-aspartate and d-serine in the mammalian brain and periphery. Prog. Neurobiol. 1997, 52, 325–353. [Google Scholar] [CrossRef]
- Nuzzo, T.; Sacchi, S.; Errico, F.; Keller, S.; Palumbo, O.; Florio, E.; Punzo, D.; Napolitano, F.; Copetti, M.; Carella, M.; et al. Decreased free d-aspartate levels are linked to enhanced d-aspartate oxidase activity in the dorsolateral prefrontal cortex of schizophrenia patients. npj Schizophr. 2017, 3, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Nuzzo, T.; Sekine, M.; Punzo, D.; Miroballo, M.; Katane, M.; Saitoh, Y.; Galbusera, A.; Pasqualetti, M.; Errico, F.; Gozzi, A.; et al. Dysfunctional d-aspartate metabolism in BTBR mouse model of idiopathic autism. Biochim. Biophys. Acta BBA Proteins Proteom. 2020, 1868, 140531. [Google Scholar] [CrossRef] [PubMed]
- Errico, F.; Napolitano, F.; Squillace, M.; Vitucci, D.; Blasi, G.; De Bartolomeis, A.; Bertolino, A.; D’Aniello, A.; Usiello, A. Decreased levels of d-aspartate and NMDA in the prefrontal cortex and striatum of patients with schizophrenia. J. Psychiatr. Res. 2013, 47, 1432–1437. [Google Scholar] [CrossRef] [PubMed]
- Di Fiore, M.M.; Santillo, A.; Falvo, S.; Longobardi, S.; Chieffi Baccari, G. Molecular Mechanisms Elicited by d-Aspartate in Leydig Cells and Spermatogonia. Int. J. Mol. Sci. 2016, 17, 1127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Fiore, M.M.; Boni, R.; Santillo, A.; Falvo, S.; Gallo, A.; Esposito, S.; Chieffi Baccari, G. D-Aspartic Acid in Vertebrate Reproduction: Animal Models and Experimental Designs. Biomolecules 2019, 9, 445. [Google Scholar] [CrossRef] [Green Version]
- Raspa, M.; Mahabir, E.; Paoletti, R.; Protti, M.; Mercolini, L.; Schiller, P.; Scavizzi, F. Effects of oral d-aspartate on sperm quality in B6N mice. Theriogenology 2018, 121, 53–61. [Google Scholar] [CrossRef]
- Raspa, M.; Paoletti, R.; Mahabir, E.; Scavizzi, F. d-aspartate treatment in vitro improves mouse sperm fertility in young B6N mice. Theriogenology 2020, 148, 60–67. [Google Scholar] [CrossRef]
- Dunlop, D.M.; Neidle, A.; McHale, D.M.; Lajtha, A. The presence of free D-aspartic acid in rodents and man. Biochem. Biophys. Res. Commun. 1986, 141, 27–32. [Google Scholar] [CrossRef]
- Hashimoto, A.; Oka, T.; Nishikawa, T. Anatomical Distribution and Postnatal Changes in Endogenous Free D-Aspartate and D-Serine in Rat Brain and Periphery. Eur. J. Neurosci. 1995, 7, 1657–1663. [Google Scholar] [CrossRef] [PubMed]
- De Rosa, A.; Mastrostefano, F.; Di Maio, A.; Nuzzo, T.; Saitoh, Y.; Katane, M.; Isidori, A.M.; Caputo, V.; Marotta, P.; Falco, G.; et al. Prenatal expression of d-aspartate oxidase causes early cerebral d-aspartate depletion and influences brain morphology and cognitive functions at adulthood. Amino Acids 2020, 52, 597–617. [Google Scholar] [CrossRef] [PubMed]
- Errico, F.; Nuzzo, T.; Carella, M.; Bertolino, A.; Usiello, A. The Emerging Role of Altered d-Aspartate Metabolism in Schizophrenia: New Insights from Preclinical Models and Human Studies. Front. Psychiatry 2018, 9, 559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Punzo, D.; Errico, F.; Cristino, L.; Sacchi, S.; Keller, S.; Belardo, C.; Luongo, L.; Nuzzo, T.; Imperatore, R.; Florio, E.; et al. Age-Related Changes in D-Aspartate Oxidase Promoter Methylation Control Extracellular D-Aspartate Levels and Prevent Precocious Cell Death during Brain Aging. J. Neurosci. 2016, 36, 3064–3078. [Google Scholar] [CrossRef]
- Errico, F.; Cuomo, M.; Canu, N.; Caputo, V.; Usiello, A. New insights on the influence of free d-aspartate metabolism in the mammalian brain during prenatal and postnatal life. Biochim. Biophys. Acta BBA Proteins Proteom. 2020, 1868, 140471. [Google Scholar] [CrossRef]
- Errico, F.; Napolitano, F.; Nisticò, R.; Usiello, A. New insights on the role of free d-aspartate in the mammalian brain. Amino Acids 2012, 43, 1861–1871. [Google Scholar] [CrossRef]
- Ota, N.; Shi, T.; Sweedler, J.V. d-Aspartate acts as a signaling molecule in nervous and neuroendocrine systems. Amino Acids 2012, 43, 1873–1886. [Google Scholar] [CrossRef] [Green Version]
- Wolosker, H.; D’Aniello, A.; Snyder, S.H. d-Aspartate disposition in neuronal and endocrine tissues: Ontogeny, biosynthesis and release. Neuroscience 2000, 100, 183–189. [Google Scholar] [CrossRef]
- Schell, M.J.; Cooper, O.; Snyder, S.H. D-aspartate localizations imply neuronal and neuroendocrine roles. Proc. Natl. Acad. Sci. USA 1997, 94, 2013–2018. [Google Scholar] [CrossRef] [Green Version]
- Cuomo, M.; Keller, S.; Punzo, D.; Nuzzo, T.; Affinito, O.; Coretti, L.; Carella, M.; De Rosa, V.; Florio, E.; Boscia, F.; et al. Selective demethylation of two CpG sites causes postnatal activation of the Dao gene and consequent removal of d-serine within the mouse cerebellum. Clin. Epigenetics 2019, 11, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Long, Z.; Homma, H.; Lee, J.-A.; Fukushima, T.; Santa, T.; Iwatsubo, T.; Yamada, R.-H.; Imai, K. Biosynthesis of D-aspartate in mammalian cells. FEBS Lett. 1998, 434, 231–235. [Google Scholar] [CrossRef] [Green Version]
- Long, Z.; Lee, J.A.; Okamoto, T.; Nimura, N.; Imai, K.; Homma, H. d-Aspartate in a prolactin-secreting clonal strain of rat pituitary tumor cells (GH(3)). Biochem. Biophys. Res. Commun. 2000, 276, 1143–1147. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, S.; Katane, M.; Maeda, K.; Kaneko, Y.; Saitoh, Y.; Miyamoto, T.; Sekine, M.; Homma, H. Biosynthesis of d-aspartate in mammals: The rat and human homologs of mouse aspartate racemase are not responsible for the biosynthesis of d-aspartate. Amino Acids 2015, 47, 975–985. [Google Scholar] [CrossRef] [PubMed]
- Kim, P.M.; Duan, X.; Huang, A.S.; Liu, C.Y.; Ming, G.-L.; Song, H.; Snyder, S.H. Aspartate racemase, generating neuronal D-aspartate, regulates adult neurogenesis. Proc. Natl. Acad. Sci. USA 2010, 107, 3175–3179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka-Hayashi, A.; Hayashi, S.; Inoue, R.; Ito, T.; Konno, K.; Yoshida, T.; Watanabe, M.; Yoshimura, T.; Mori, H. Is d-aspartate produced by glutamic-oxaloacetic transaminase-1 like 1 (Got1l1): A putative aspartate racemase? Amino Acids 2015, 47, 79–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horio, M.; Ishima, T.; Fujita, Y.; Inoue, R.; Mori, H.; Hashimoto, K. Decreased levels of free d-aspartic acid in the forebrain of serine racemase (Srr) knock-out mice. Neurochem. Int. 2013, 62, 843–847. [Google Scholar] [CrossRef] [PubMed]
- Ito, T.; Hayashida, M.; Kobayashi, S.; Muto, N.; Hayashi, A.; Yoshimura, T.; Mori, H. Serine racemase is involved in d-aspartate biosynthesis. J. Biochem. 2016, 160, 345–353. [Google Scholar] [CrossRef]
- Katane, M.; Kuwabara, H.; Nakayama, K.; Saitoh, Y.; Miyamoto, T.; Sekine, M.; Homma, H. Biochemical characterization of d-aspartate oxidase from Caenorhabditis elegans: Its potential use in the determination of free d-glutamate in biological samples. Biochim. Biophys. Acta BBA Proteins Proteom. 2020, 1868, 140442. [Google Scholar] [CrossRef]
- Molla, G.; Chaves-Sanjuan, A.; Savinelli, A.; Nardini, M.; Pollegioni, L.; Chaves-Sanjuan, A. Structure and kinetic properties of human d -aspartate oxidase, the enzyme-controlling d -aspartate levels in brain. FASEB J. 2019, 34, 1182–1197. [Google Scholar] [CrossRef] [Green Version]
- Puggioni, V.; Savinelli, A.; Miceli, M.; Molla, G.; Pollegioni, L.; Sacchi, S. Biochemical characterization of mouse d-aspartate oxidase. Biochim. Biophys. Acta BBA Proteins Proteom. 2020, 1868, 140472. [Google Scholar] [CrossRef]
- Katane, M.; Saitoh, Y.; Hanai, T.; Sekine, M.; Furuchi, T.; Koyama, N.; Nakagome, I.; Tomoda, H.; Hirono, S.; Homma, H. Thiolactomycin inhibits d-aspartate oxidase: A novel approach to probing the active site environment. Biochimie 2010, 92, 1371–1378. [Google Scholar] [CrossRef] [PubMed]
- Van Veldhoven, P.P.; Brees, C.; Mannaerts, G.P. d-Aspartate oxidase, a peroxisomal enzyme in liver of rat and man. Biochim. Biophys. Acta BBA Gen. Subj. 1991, 1073, 203–208. [Google Scholar] [CrossRef]
- Zaar, K.; Köst, H.-P.; Schad, A.; Völkl, A.; Baumgart, E.; Fahimi, H.D.; Baumgart-Vogt, E. Cellular and subcellular distribution of D-aspartate oxidase in human and rat brain. J. Comp. Neurol. 2002, 450, 272–282. [Google Scholar] [CrossRef] [PubMed]
- Errico, F.; Pirro, M.T.; Affuso, A.; Spinelli, P.; De Felice, M.; D’Aniello, A.; Di Lauro, R. A physiological mechanism to regulate d-aspartic acid and NMDA levels in mammals revealed by D-aspartate oxidase deficient mice. Gene 2006, 374, 50–57. [Google Scholar] [CrossRef]
- Sacchi, S.; Bernasconi, M.; Martineau, M.; Mothet, J.-P.; Ruzzene, M.; Pilone, M.S.; Pollegioni, L.; Molla, G. pLG72 Modulates Intracellular D-Serine Levels through Its Interaction with D-Amino Acid Oxidase: Effect on schizophrenia susceptibility. J. Biol. Chem. 2008, 283, 22244–22256. [Google Scholar] [CrossRef] [Green Version]
- Keller, S.; Punzo, D.; Cuomo, M.; Affinito, O.; Coretti, L.; Sacchi, S.; Florio, E.; Lembo, F.; Carella, M.; Copetti, M.; et al. DNA methylation landscape of the genes regulating D-serine and D-aspartate metabolism in post-mortem brain from controls and subjects with schizophrenia. Sci. Rep. 2018, 8, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Florio, E.; Keller, S.; Coretti, L.; Affinito, O.; Scala, G.; Errico, F.; Fico, A.; Boscia, F.; Sisalli, M.J.; Reccia, M.G.; et al. Tracking the evolution of epialleles during neural differentiation and brain development: D-Aspartate oxidase as a model gene. Epigenetics 2017, 12, 41–54. [Google Scholar] [CrossRef]
- Jagannath, V.; Marinova, Z.; Monoranu, C.-M.; Walitza, S.; Grünblatt, E. Expression of D-Amino Acid Oxidase (DAO/DAAO) and D-Amino Acid Oxidase Activator (DAOA/G72) during Development and Aging in the Human Post-mortem Brain. Front. Neuroanat. 2017, 11, 31. [Google Scholar] [CrossRef] [Green Version]
- Fagg, G.E.; Matus, A. Selective association of N-methyl aspartate and quisqualate types of L-glutamate receptor with brain postsynaptic densities. Proc. Natl. Acad. Sci. USA 1984, 81, 6876–6880. [Google Scholar] [CrossRef] [Green Version]
- Ransom, R.W.; Stec, N.L. Cooperative Modulation of [3H]MK-801 Binding to the N-Methyl-d-Aspartate Receptor-Ion Channel Complex by l-Glutamate, Glycine, and Polyamines. J. Neurochem. 1988, 51, 830–836. [Google Scholar] [CrossRef]
- Errico, F.; Nisticò, R.; Palma, G.; Federici, M.; Affuso, A.; Brilli, E.; Topo, E.; Centonze, D.; Bernardi, G.; Bozzi, Y.; et al. Increased levels of d-aspartate in the hippocampus enhance LTP but do not facilitate cognitive flexibility. Mol. Cell. Neurosci. 2008, 37, 236–246. [Google Scholar] [CrossRef] [PubMed]
- Krashia, P.; LeDonne, A.; Nobili, A.; Cordella, A.; Errico, F.; Usiello, A.; D’Amelio, M.; Mercuri, N.B.; Guatteo, E.; Carunchio, I. Persistent elevation of D-Aspartate enhances NMDA receptor-mediated responses in mouse substantia nigra pars compacta dopamine neurons. Neuropharmacology 2016, 103, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Errico, F.; Nisticò, R.; Napolitano, F.; Oliva, A.B.; Romano, R.; Barbieri, F.; Florio, T.; Russo, C.; Mercuri, N.B.; Usiello, A. Persistent increase of d-aspartate in d-aspartate oxidase mutant mice induces a precocious hippocampal age-dependent synaptic plasticity and spatial memory decay. Neurobiol. Aging 2011, 32, 2061–2074. [Google Scholar] [CrossRef] [PubMed]
- Gong, X.-Q.; Frandsen, A.; Lu, W.-Y.; Wan, Y.; Zabek, R.L.; Pickering, D.S.; Bai, D. D -Aspartate and NMDA, but not L -aspartate, block AMPA receptors in rat hippocampal neurons. Br. J. Pharmacol. 2005, 145, 449–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molinaro, G.; Pietracupa, S.; Di Menna, L.; Pescatori, L.; Usiello, A.; Battaglia, G.; Nicoletti, F.; Bruno, V.M.G. d-Aspartate activates mGlu receptors coupled to polyphosphoinositide hydrolysis in neonate rat brain slices. Neurosci. Lett. 2010, 478, 128–130. [Google Scholar] [CrossRef] [PubMed]
- Sacchi, S.; De Novellis, V.; Paolone, G.; Nuzzo, T.; Iannotta, M.; Belardo, C.; Squillace, M.; Bolognesi, P.; Rosini, E.; Motta, Z.; et al. Olanzapine, but not clozapine, increases glutamate release in the prefrontal cortex of freely moving mice by inhibiting D-aspartate oxidase activity. Sci. Rep. 2017, 7, srep46288. [Google Scholar] [CrossRef] [Green Version]
- Boccella, S.; Vacca, V.; Errico, F.; Marinelli, S.; Squillace, M.; Guida, F.; Di Maio, A.; Vitucci, D.; Palazzo, E.; De Novellis, V.; et al. D-Aspartate Modulates Nociceptive-Specific Neuron Activity and Pain Threshold in Inflammatory and Neuropathic Pain Condition in Mice. BioMed Res. Int. 2015, 2015, 905906. [Google Scholar] [CrossRef]
- D’Aniello, S.; Somorjai, I.M.L.; Garcia-Fernàndez, J.; Topo, E.; D’Aniello, A. D-Aspartic acid is a novel endogenous neurotransmitter. FASEB J. 2010, 25, 1014–1027. [Google Scholar] [CrossRef] [Green Version]
- Anderson, C.M.; Bridges, R.J.; Chamberlin, A.R.; Shimamoto, K.; Yasuda-Kamatani, Y.; Swanson, R.A. Differing effects of substrate and non-substrate transport inhibitors on glutamate uptake reversal. J. Neurochem. 2001, 79, 1207–1216. [Google Scholar] [CrossRef]
- Bak, L.K.; Schousboe, A.; Waagepetersen, H.S. Characterization of depolarization-coupled release of glutamate from cultured mouse cerebellar granule cells using dl-threo-β-benzyloxyaspartate (DL-TBOA) to distinguish between the vesicular and cytoplasmic pools. Neurochem. Int. 2003, 43, 417–424. [Google Scholar] [CrossRef]
- Guida, F.; Luongo, L.; Marmo, F.; Romano, R.; Iannotta, M.; Napolitano, F.; Belardo, C.; Marabese, I.; Aniello, A.D.; De Gregorio, D.; et al. Palmitoylethanolamide reduces pain-related behaviors and restores glutamatergic synapses homeostasis in the medial prefrontal cortex of neuropathic mice. Mol. Brain 2015, 8, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Palacín, M.; Estévez, R.; Bertran, J.; Zorzano, A. Molecular Biology of Mammalian Plasma Membrane Amino Acid Transporters. Physiol. Rev. 1998, 78, 969–1054. [Google Scholar] [CrossRef] [PubMed]
- Arkhipova, V.; Trinco, G.; Ettema, T.W.; Jensen, S.; Slotboom, D.J.; Guskov, A. Binding and transport of D-aspartate by the glutamate transporter homolog GltTk. eLife 2019, 8, 8. [Google Scholar] [CrossRef] [PubMed]
- Huang, A.S.; Beigneux, A.; Weil, Z.M.; Kim, P.M.; Molliver, M.E.; Blackshaw, S.; Nelson, R.J.; Young, S.G.; Snyder, S.H. D-Aspartate Regulates Melanocortin Formation and Function: Behavioral Alterations in D-Aspartate Oxidase-Deficient Mice. J. Neurosci. 2006, 26, 2814–2819. [Google Scholar] [CrossRef] [Green Version]
- Weil, Z.M.; Huang, A.S.; Beigneux, A.; Kim, P.M.; Molliver, M.E.; Blackshaw, S.; Young, S.G.; Nelson, R.J.; Snyder, S.H. Behavioural alterations in male mice lacking the gene for d-aspartate oxidase. Behav. Brain Res. 2006, 171, 295–302. [Google Scholar] [CrossRef]
- Jami, S.A.; Cameron, S.; Wong, J.M.; Daly, E.R.; McAllister, A.K.; Gray, J.A. Increased excitation-inhibition balance due to a loss of GABAergic synapses in the serine racemase knockout model of NMDA receptor hypofunction. bioRxiv 2020. [Google Scholar] [CrossRef]
- Errico, F.; Bonito-Oliva, A.; Bagetta, V.; Vitucci, D.; Romano, R.; Zianni, E.; Napolitano, F.; Marinucci, S.; Di Luca, M.; Calabresi, P.; et al. Higher free D-aspartate and N-methyl-d-aspartate levels prevent striatal depotentiation and anticipate l-DOPA-induced dyskinesia. Exp. Neurol. 2011, 232, 240–250. [Google Scholar] [CrossRef]
- Errico, F.; Rossi, S.; Napolitano, F.; Catuogno, V.; Topo, E.; Fisone, G.; D’Aniello, A.; Centonze, D.; Usiello, A. D-Aspartate Prevents Corticostriatal Long-Term Depression and Attenuates Schizophrenia-Like Symptoms Induced by Amphetamine and MK-801. J. Neurosci. 2008, 28, 10404–10414. [Google Scholar] [CrossRef] [Green Version]
- Kitamura, A.; Hojo, Y.; Ikeda, M.; Karakawa, S.; Kuwahara, T.; Kim, J.; Soma, M.; Kawato, S.; Tsurugizawa, T. Ingested d-Aspartate Facilitates the Functional Connectivity and Modifies Dendritic Spine Morphology in Rat Hippocampus. Cereb. Cortex 2018, 29, 2499–2508. [Google Scholar] [CrossRef]
- Bauer, D.; Hamacher, K.; Bröer, S.; Pauleit, D.; Palm, C.; Zilles, K.; Coenen, H.H.; Langen, K.-J. Preferred stereoselective brain uptake of d-serine — a modulator of glutamatergic neurotransmission. Nucl. Med. Biol. 2005, 32, 793–797. [Google Scholar] [CrossRef]
- Langen, K.-J.; Hamacher, K.; Bauer, D.; Bröer, S.; Pauleit, D.; Herzog, H.; Floeth, F.; Zilles, K.; Coenen, H.H. Preferred Stereoselective Transport of the D-isomer of cis-4-[18F]fluoro-proline at the Blood–Brain Barrier. Br. J. Pharmacol. 2005, 25, 607–616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Errico, F.; Nistico, R.; Di Giorgio, A.; Squillace, M.; Vitucci, D.; Galbusera, A.; Piccinin, S.; Mango, D.; Fazio, L.; Middei, S.; et al. Free D-aspartate regulates neuronal dendritic morphology, synaptic plasticity, gray matter volume and brain activity in mammals. Transl. Psychiatry 2014, 4, e417. [Google Scholar] [CrossRef] [PubMed]
- Topo, E.; Soricelli, A.; Di Maio, A.; D’Aniello, E.; Di Fiore, M.M.; D’Aniello, A. Evidence for the involvement of d-aspartic acid in learning and memory of rat. Amino Acids 2009, 38, 1561–1569. [Google Scholar] [CrossRef] [PubMed]
- Luo, T.; Wu, W.-H.; Chen, B.-S. NMDA receptor signaling: Death or survival? Front. Biol. 2011, 6, 468–476. [Google Scholar] [CrossRef] [Green Version]
- Hardingham, G.E.; Bading, H. The Yin and Yang of NMDA receptor signalling. Trends Neurosci. 2003, 26, 81–89. [Google Scholar] [CrossRef]
- Nuzzo, T.; Feligioni, M.; Cristino, L.; Pagano, I.; Marcelli, S.; Iannuzzi, F.; Imperatore, R.; D’Angelo, L.; Petrella, C.; Carella, M.; et al. Free D-aspartate triggers NMDA receptor-dependent cell death in primary cortical neurons and perturbs JNK activation, Tau phosphorylation, and protein SUMOylation in the cerebral cortex of mice lacking d-aspartate oxidase activity. Exp. Neurol. 2019, 317, 51–65. [Google Scholar] [CrossRef] [Green Version]
- Di Giorgi-Gerevini, V.; Melchiorri, D.; Battaglia, G.; Ricci-Vitiani, L.; Ciceroni, C.; Busceti, C.L.; Biagioni, F.; Iacovelli, L.; Canudas, A.M.; Parati, E.; et al. Endogenous activation of metabotropic glutamate receptors supports the proliferation and survival of neural progenitor cells. Cell Death Differ. 2005, 12, 1124–1133. [Google Scholar] [CrossRef] [Green Version]
- Ikonomidou, C. Triggers of apoptosis in the immature brain. Brain Dev. 2009, 31, 488–492. [Google Scholar] [CrossRef]
- Jansson, L.C.; Åkerman, K.E. The role of glutamate and its receptors in the proliferation, migration, differentiation and survival of neural progenitor cells. J. Neural Transm. 2014, 121, 819–836. [Google Scholar] [CrossRef]
- Komuro, H.; Rakic, P. Modulation of neuronal migration by NMDA receptors. Science 1993, 260, 95–97. [Google Scholar] [CrossRef]
- Coyle, J.T. NMDA Receptor and Schizophrenia: A Brief History. Schizophr. Bull. 2012, 38, 920–926. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, J.; Jurado-Coronel, J.C.; Ávila, M.F.; Sabogal, A.; Capani, F.; Barreto, G.E. NMDARs in neurological diseases: A potential therapeutic target. Int. J. Neurosci. 2014, 125, 315–327. [Google Scholar] [CrossRef] [PubMed]
- Krivoy, A.; Fischel, T.; Weizman, A. The possible involvement of metabotropic glutamate receptors in schizophrenia. Eur. Neuropsychopharmacol. 2008, 18, 395–405. [Google Scholar] [CrossRef]
- Matosin, N.; Fernandez, F.M.; Lum, J.S.; Newell, K.A. Shifting towards a model of mGluR5 dysregulation in schizophrenia: Consequences for future schizophrenia treatment. Neuropharmacology 2017, 115, 73–91. [Google Scholar] [CrossRef] [PubMed]
- Moghaddam, B.; Javitt, D.C. From Revolution to Evolution: The Glutamate Hypothesis of Schizophrenia and its Implication for Treatment. Neuropsychopharmacology 2012, 37, 4–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribeiro, F.M.; Paquet, M.; Cregan, S.P.; Ferguson, S.S.G. Group I Metabotropic Glutamate Receptor Signalling and its Implication in Neurological Disease. CNS Neurol. Disord. Drug Targets 2010, 9, 574–595. [Google Scholar] [CrossRef] [PubMed]
- Gardoni, F.; Bellone, C. Modulation of the glutamatergic transmission by Dopamine: A focus on Parkinson, Huntington and Addiction diseases. Front. Cell. Neurosci. 2015, 9, 25. [Google Scholar] [CrossRef] [Green Version]
- Mellone, M.; Gardoni, F. Glutamatergic mechanisms in l-DOPA-induced dyskinesia and therapeutic implications. J. Neural Transm. 2018, 125, 1225–1236. [Google Scholar] [CrossRef]
- Nuzzo, T.; Punzo, D.; Devoto, P.; Rosini, E.; Paciotti, S.; Sacchi, S.; Li, Q.; Thiolat, M.-L.; Véga, C.; Carella, M.; et al. The levels of the NMDA receptor co-agonist D-serine are reduced in the substantia nigra of MPTP-lesioned macaques and in the cerebrospinal fluid of Parkinson’s disease patients. Sci. Rep. 2019, 9, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Nuzzo, T.; Miroballo, M.; Casamassa, A.; Mancini, A.; Gaetani, L.; Nistico, R.; Eusebi, P.; Katane, M.; Homma, H.; Calabresi, P.; et al. Cerebrospinal fluid and serum d-serine concentrations are unaltered across the whole clinical spectrum of Alzheimer’s disease. Biochim. Biophys. Acta Proteins. Proteom. 2020, 1868, 140537. [Google Scholar] [CrossRef]
- Li, Y.; Han, H.; Yin, J.; Li, T.; Yin, Y. Role of D-aspartate on biosynthesis, racemization, and potential functions: A mini-review. Anim. Nutr. 2018, 4, 311–315. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, A.; Tanaka, H.; Ishida, T.; Horiike, K. d-Aspartate Oxidase Localisation in Pituitary and Pineal Glands of the Female Pig. J. Neuroendocr. 2010, 22, 1165–1172. [Google Scholar] [CrossRef] [PubMed]
- Topo, E.; Soricelli, A.; D’Aniello, A.; Ronsini, S.; D’Aniello, A. The role and molecular mechanism of D-aspartic acid in the release and synthesis of LH and testosterone in humans and rats. Reprod. Biol. Endocrinol. 2009, 7, 120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burrone, L.; Di Giovanni, M.; Di Fiore, M.M.; Chieffi Baccari, G.; Santillo, A. Effects of D-Aspartate Treatment on D-Aspartate Oxidase, Superoxide Dismutase, and Caspase 3 Activities in Frog (Rana esculenta) Tissues. Chem. Biodivers. 2010, 7, 1459–1466. [Google Scholar] [CrossRef]
- Di Giovanni, M.; Burrone, L.; Chieffi Baccari, G.; Topo, E.; Santillo, A. Distribution of free D-aspartic acid and D-aspartate oxidase in frogRana esculentatissues. J. Exp. Zool. Part A Ecol. Genet. Physiol. 2010, 313, 137–143. [Google Scholar] [CrossRef]
- Han, H.; Miyoshi, Y.; Koga, R.; Mita, M.; Konno, R.; Hamase, K. Changes in d-aspartic acid and d-glutamic acid levels in the tissues and physiological fluids of mice with various d-aspartate oxidase activities. J. Pharm. Biomed. Anal. 2015, 116, 47–52. [Google Scholar] [CrossRef]
- Hamase, K.; Konno, R.; Morikawa, A.; Zaitsu, K. Sensitive Determination of D-Amino Acids in Mammals and the Effect of D-Amino-Acid Oxidase Activity on Their Amounts. Biol. Pharm. Bull. 2005, 28, 1578–1584. [Google Scholar] [CrossRef] [Green Version]
- Han, H.; Miyoshi, Y.; Ueno, K.; Okamura, C.; Tojo, Y.; Mita, M.; Lindner, W.; Zaitsu, K.; Hamase, K. Simultaneous determination of d-aspartic acid and d-glutamic acid in rat tissues and physiological fluids using a multi-loop two-dimensional HPLC procedure. J. Chromatogr. B 2011, 879, 3196–3202. [Google Scholar] [CrossRef]
- Morikawa, A.; Hamase, K.; Inoue, T.; Konno, R.; Niwa, A.; Zaitsu, K. Determination of free D-aspartic acid, D-serine and D-alanine in the brain of mutant mice lacking D-amino acid oxidase activity. J. Chromatogr. B, Biomed. Sci. Appl. 2001, 757, 119–125. [Google Scholar] [CrossRef]
- Miyoshi, Y.; Koga, R.; Oyama, T.; Han, H.; Ueno, K.; Masuyama, K.; Itoh, Y.; Hamase, K. HPLC analysis of naturally occurring free d-amino acids in mammals. J. Pharm. Biomed. Anal. 2012, 69, 42–49. [Google Scholar] [CrossRef]
- Imai, K.; Fukushima, T.; Hagiwara, K.; Santa, T. Occurrence ofD-aspartic acid in rat brain pineal gland. Biomed. Chromatogr. 1995, 9, 106–109. [Google Scholar] [CrossRef] [PubMed]
- Sakai, K.; Homma, H.; Lee, J.-A.; Fukushima, T.; Santa, T.; Tashiro, K.; Iwatsubo, T.; Imai, K. D-Aspartic Acid Localization during Postnatal Development of Rat Adrenal Gland. Biochem. Biophys. Res. Commun. 1997, 235, 433–436. [Google Scholar] [CrossRef] [PubMed]
- D·Aniello, A.; Di Fiore, M.M.; Fisher, G.H.; Milone, A.; Seleni, A.; D’Aniello, S.; Perna, A.F.; Ingrosso, D. Occurrence of D-aspartic acid and N-methyl-D-aspartic acid in rat neuroendocrine tissues and their role in the modulation of luteinizing hormone and growth hormone release. FASEB J. 2000, 14, 699–714. [Google Scholar] [CrossRef]
- Wang, H.; Wolosker, H.; Pevsner, J.; Snyder, S.H.; Selkoe, D.J. Regulation of rat magnocellular neurosecretory system by D-aspartate: Evidence for biological role(s) of a naturally occurring free D-amino acid in mammals. J. Endocrinol. 2000, 167, 247–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pampillo, M.; Scimonelli, T.; Bottino, M.C.; Duvilanski, B.H.; McCann, S.M.; Seilicovich, A.; Lasaga, M. The effect of D-aspartate on luteinizing hormone-releasing hormone, α-melanocyte-stimulating hormone, GABA and dopamine release. NeuroReport 2002, 13, 2341–2344. [Google Scholar] [CrossRef] [PubMed]
- D’Aniello, A. d-Aspartic acid: An endogenous amino acid with an important neuroendocrine role. Brain Res. Rev. 2007, 53, 215–234. [Google Scholar] [CrossRef]
- Hamase, K.; Homma, H.; Takigawa, Y.; Fukushima, T.; Santa, T.; Imai, K. Regional distribution and postnatal changes of d-amino acids in rat brain. Biochim. Biophys. Acta BBA Gen. Subj. 1997, 1334, 214–222. [Google Scholar] [CrossRef]
- Han, H.; Miyoshi, Y.; Oyama, T.; Konishi, R.; Mita, M.; Hamase, K. Enantioselective micro-2D-HPLC determination of aspartic acid in the pineal glands of rodents with various melatonin contents. J. Sep. Sci. 2011, 34, 2847–2853. [Google Scholar] [CrossRef]
- Ishio, S.; Yamada, H.; Hayashi, M.; Yatsushiro, S.; Noumi, T.; Yamaguchi, A.; Moriyama, Y. d-Aspartate modulates melatonin synthesis in rat pinealocytes. Neurosci. Lett. 1998, 249, 143–146. [Google Scholar] [CrossRef]
- Karakawa, S.; Shimbo, K.; Yamada, N.; Mizukoshi, T.; Miyano, H.; Mita, M.; Lindner, W.; Hamase, K. Simultaneous analysis of d-alanine, d-aspartic acid, and d-serine using chiral high-performance liquid chromatography-tandem mass spectrometry and its application to the rat plasma and tissues. J. Pharm. Biomed. Anal. 2015, 115, 123–129. [Google Scholar] [CrossRef]
- Tsunoda, M.; Kato, M.; Fukushima, T.; Santa, T.; Homma, H.; Yanai, H.; Soga, T.; Imai, K. Determination of aspartic acid enantiomers in bio-samples by capillary electrophoresis. Biomed. Chromatogr. 1999, 13, 335–339. [Google Scholar] [CrossRef]
- Yatsushiro, S.; Yamada, H.; Kozaki, S.; Kumon, H.; Michibata, H.; Yamamoto, A.; Moriyama, Y. L-aspartate but not the D form is secreted through microvesicle-mediated exocytosis and is sequestered through Na+-dependent transporter in rat pinealocytes. J. Neurochem. 1997, 69, 340–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takigawa, Y.; Homma, H.; Lee, J.-A.; Fukushima, T.; Santa, T.; Iwatsubo, T.; Imai, K. d-Aspartate Uptake into Cultured Rat Pinealocytes and the Concomitant Effect onl-Aspartate Levels and Melatonin Secretion. Biochem. Biophys. Res. Commun. 1998, 248, 641–647. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-A.; Long, Z.; Nimura, N.; Iwatsubo, T.; Imai, K.; Homma, H. Localization, Transport, and Uptake of -Aspartate in the Rat Adrenal and Pituitary Glands. Arch. Biochem. Biophys. 2001, 385, 242–249. [Google Scholar] [CrossRef]
- Long, Z.; Sekine, M.; Adachi, M.; Furuchi, T.; Imai, K.; Nimura, N.; Homma, H. Cell density inversely regulates d- and l-aspartate levels in rat pheochromocytoma MPT1 cells. Arch. Biochem. Biophys. 2002, 404, 92–97. [Google Scholar] [CrossRef]
- Nakatsuka, S.; Hayashi, M.; Muroyama, A.; Otsuka, M.; Kozaki, S.; Yamada, H.; Moriyama, Y. d-Aspartate Is Stored in Secretory Granules and Released through a Ca2+-dependent Pathway in a Subset of Rat Pheochromocytoma PC12 Cells. J. Biol. Chem. 2001, 276, 26589–26596. [Google Scholar] [CrossRef] [Green Version]
- Santillo, A.; Falvo, S.; Chieffi, P.; Burrone, L.; Chieffi Baccari, G.; Longobardi, S.; Di Fiore, M.M. d-aspartate affects NMDA receptor-extracellular signal–regulated kinase pathway and upregulates androgen receptor expression in the rat testis. Theriogenology 2014, 81, 744–751. [Google Scholar] [CrossRef]
- Yamada, H.; Yatsushiro, S.; Yamamoto, A.; Hayashi, M.; Nishi, T.; Futai, M.; Yamaguchi, A.; Moriyama, Y. Functional expression of a GLT-1 type Na+-dependent glutamate transporter in rat pinealocytes. J. Neurochem. 1997, 69, 1491–1498. [Google Scholar] [CrossRef]
- Hamase, K.; Morikawa, A.; Zaitsu, K. d-Amino acids in mammals and their diagnostic value. J. Chromatogr. B 2002, 781, 73–91. [Google Scholar] [CrossRef]
- Topo, E.; Fisher, G.; Sorricelli, A.; Errico, F.; Usiello, A.; D’Aniello, A. Thyroid Hormones and D-Aspartic Acid, D-Aspartate Oxidase, D-Aspartate Racemase, H2O2, and ROS in Rats and Mice. Chem. Biodivers. 2010, 7, 1467–1478. [Google Scholar] [CrossRef]
- Lee, J.-A.; Homma, H.; Tashiro, K.; Iwatsubo, T.; Imai, K. d-Aspartate localization in the rat pituitary gland and retina. Brain Res. 1999, 838, 193–199. [Google Scholar] [CrossRef]
- Boni, R.; Santillo, R.; Macchia, G.; Spinelli, P.; Ferrandino, G.; D’Aniello, A. d-Aspartate and reproductive activity in sheep. Theriogenology 2006, 65, 1265–1278. [Google Scholar] [CrossRef] [PubMed]
- D’Aniello, G.; Tolino, A.; D’Aniello, A.; Errico, F.; Fisher, G.H.; Di Fiore, M.M. The role of D-aspartic acid and N-methyl-D-aspartic acid in the regulation of prolactin release. Endocrinology 2000, 141, 3862–3870. [Google Scholar] [CrossRef]
- Raucci, F.; D’Aniello, A.; Di Fiore, M.M. Stimulation of androgen production by D-aspartate through the enhancement of StAR, P450scc and 3beta-HSD mRNA levels in vivo rat testis and in culture of immature rat Leydig cells. Steroids 2014, 84, 103–110. [Google Scholar] [CrossRef]
- Burrone, L.; Santillo, A.; Pinelli, C.; Chieffi Baccari, G.; Di Fiore, M.M. Induced synthesis of P450 aromatase and 17beta-estradiol by D-aspartate in frog brain. J. Exp. Biol. 2012, 215, 3559–3565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Fiore, M.M.; Santillo, A.; Falvo, S.; Chieffi Baccari, G.; Venditti, M.; Russo, F.D.G.; Lispi, M.; D’Aniello, A. Sex hormone levels in the brain of d -aspartate-treated rats. Comptes Rendus Biol. 2018, 341, 9–15. [Google Scholar] [CrossRef]
- Santillo, A.; Pinelli, C.; Burrone, L.; Chieffi Baccari, G.; Di Fiore, M.M. d-Aspartic acid implication in the modulation of frog brain sex steroid levels. Gen. Comp. Endocrinol. 2013, 181, 72–76. [Google Scholar] [CrossRef]
- Falvo, S.; Di Fiore, M.M.; Burrone, L.; Chieffi Baccari, G.; Longobardi, S.; Santillo, A. Androgen and oestrogen modulation by D-aspartate in rat epididymis. Reprod. Fertil. Dev. 2016, 28, 1865. [Google Scholar] [CrossRef]
- Nagata, Y.; Homma, H.; Matsumoto, M.; Imai, K. Stimulation of steroidogenic acute regulatory protein (STAR) gene expression by D-aspartate in rat Leydig cells. FEBS Lett. 1999, 454, 317–320. [Google Scholar] [CrossRef] [Green Version]
- Di Nisio, A.; De Toni, L.; Ferigo, M.; Rocca, M.S.; Speltra, E.; Ferlin, A.; Foresta, C. d-Aspartic acid stimulates steroidogenesis through the delay of LH receptor internalization in a mammalian Leydig cell line. J. Endocrinol. Investig. 2015, 39, 207–213. [Google Scholar] [CrossRef]
- Bhat, G.K.; Mahesh, V.B.; Chu, Z.W.; Chorich, L.P.; Zamorano, P.L.; Brann, D.W. Localization of the N-Methyl-D-Aspartate R1 Receptor Subunit in Specific Anterior Pituitary Hormone Cell Types of the Female Rat. Neuroendocrinology 1995, 62, 178–186. [Google Scholar] [CrossRef] [PubMed]
- Storto, M.; Sallese, M.; Salvatore, L.; Poulet, R.; Condorelli, D.F.; Dell’Albani, P.; Marcello, M.F.; Romeo, R.; Piomboni, P.; Barone, N.; et al. Expression of metabotropic glutamate receptors in the rat and human testis. J. Endocrinol. 2001, 170, 71–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Giovanni, M.; Topo, E.; Santillo, A.; D’Aniello, A.; Chieffi Baccari, G. d-Aspartate binding sites in rat Harderian gland. Amino Acids 2009, 38, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Santillo, A.; Falvo, S.; Chieffi, P.; Di Fiore, M.M.; Senese, R.; Chieffi Baccari, G. D-Aspartate Induces Proliferative Pathways in Spermatogonial GC-1 Cells. J. Cell. Physiol. 2015, 231, 490–495. [Google Scholar] [CrossRef]
- Santillo, A.; Falvo, S.; Di Fiore, M.M.; Russo, F.D.G.; Chieffi, P.; Usiello, A.; Pinelli, C.; Chieffi Baccari, G. AMPA receptor expression in mouse testis and spermatogonial GC-1 cells: A study on its regulation by excitatory amino acids. J. Cell. Biochem. 2019, 120, 11044–11055. [Google Scholar] [CrossRef]
- Tomita, K.; Tanaka, H.; Kageyama, S.; Nagasawa, M.; Wada, A.; Murai, R.; Kobayashi, K.; Hanada, E.; Agata, Y.; Kawauchi, A. The Effect of d-Aspartate on Spermatogenesis in Mouse Testis1. Biol. Reprod. 2016, 94, 30. [Google Scholar] [CrossRef] [Green Version]
- Santillo, A.; Venditti, M.; Minucci, S.; Chieffi Baccari, G.; Falvo, S.; Rosati, L.; Di Fiore, M.M. D-Asp upregulates PREP and GluA2/3 expressions and induces p-ERK1/2 and p-Akt in rat testis. Reproduction 2019, 158, 357–367. [Google Scholar] [CrossRef]
- Venditti, M.; Santillo, A.; Falvo, S.; Di Fiore, M.M.; Chieffi Baccari, G.; Minucci, S. D-Aspartate Upregulates DAAM1 Protein Levels in the Rat Testis and Induces Its Localization in Spermatogonia Nucleus. Biomolecules 2020, 10, 677. [Google Scholar] [CrossRef]
- Boni, R.; Gallo, A.; Cecchini, S. Kinetic activity, membrane mitochondrial potential, lipid peroxidation, intracellular pH and calcium of frozen/thawed bovine spermatozoa treated with metabolic enhancers. Andrology 2016, 5, 133–145. [Google Scholar] [CrossRef]
- D’Aniello, G.; Ronsini, S.; Guida, F.; Spinelli, P.; D’Aniello, A. Occurrence of D-aspartic acid in human seminal plasma and spermatozoa: Possible role in reproduction. Fertil. Steril. 2005, 84, 1444–1449. [Google Scholar] [CrossRef]
- D’Aniello, A.; Ronsini, S.; Notari, T.; Grieco, N.; Infante, V.; D’Angel, N.; Mascia, F.; Di Fiore, M.M.; Fisher, G.; D’Aniello, A. D-Aspartate, a Key Element for the Improvement of Sperm Quality. Adv. Sex. Med. 2012, 2, 45–53. [Google Scholar] [CrossRef] [Green Version]
- Giacone, F.; Condorelli, R.A.; Mongioì, L.M.; Bullara, V.; La Vignera, S.; Calogero, A.E. In vitro effects of zinc, D-aspartic acid, and coenzyme-Q10 on sperm function. Endocrine 2016, 56, 408–415. [Google Scholar] [CrossRef] [PubMed]
- Fink, J.; Schoenfeld, B.J.; Nakazato, K. The role of hormones in muscle hypertrophy. Physician Sportsmed. 2018, 46, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Willoughby, D.S.; Leutholtz, B. d-Aspartic acid supplementation combined with 28 days of heavy resistance training has no effect on body composition, muscle strength, and serum hormones associated with the hypothalamo-pituitary-gonadal axis in resistance-trained men. Nutr. Res. 2013, 33, 803–810. [Google Scholar] [CrossRef] [PubMed]
- Melville, G.W.; Siegler, J.C.; Marshall, P.W. Three and six grams supplementation of d-aspartic acid in resistance trained men. J. Int. Soc. Sports Nutr. 2015, 12, 15. [Google Scholar] [CrossRef] [Green Version]
- Melville, G.; Siegler, J.C.; Marshall, P.W.M. The effects of d-aspartic acid supplementation in resistance-trained men over a three month training period: A randomised controlled trial. PLoS ONE 2017, 12, e0182630. [Google Scholar] [CrossRef] [Green Version]
- De Rosa, V.; Secondo, A.; Pannaccione, A.; Ciccone, R.; Formisano, L.; Guida, N.; Crispino, R.; Fico, A.; Polishchuk, R.; D’Aniello, A.; et al. D-Aspartate treatment attenuates myelin damage and stimulates myelin repair. EMBO Mol. Med. 2018, 11, e9278. [Google Scholar] [CrossRef]
- Nicoletti, C.G.; Monteleone, F.; Marfia, G.A.; Usiello, A.; Buttari, F.; Centonze, D.; Mori, F. Oral D-Aspartate enhances synaptic plasticity reserve in progressive multiple sclerosis. Mult. Scler. J. 2019, 26, 304–311. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Usiello, A.; Di Fiore, M.M.; De Rosa, A.; Falvo, S.; Errico, F.; Santillo, A.; Nuzzo, T.; Chieffi Baccari, G. New Evidence on the Role of D-Aspartate Metabolism in Regulating Brain and Endocrine System Physiology: From Preclinical Observations to Clinical Applications. Int. J. Mol. Sci. 2020, 21, 8718. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21228718
Usiello A, Di Fiore MM, De Rosa A, Falvo S, Errico F, Santillo A, Nuzzo T, Chieffi Baccari G. New Evidence on the Role of D-Aspartate Metabolism in Regulating Brain and Endocrine System Physiology: From Preclinical Observations to Clinical Applications. International Journal of Molecular Sciences. 2020; 21(22):8718. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21228718
Chicago/Turabian StyleUsiello, Alessandro, Maria Maddalena Di Fiore, Arianna De Rosa, Sara Falvo, Francesco Errico, Alessandra Santillo, Tommaso Nuzzo, and Gabriella Chieffi Baccari. 2020. "New Evidence on the Role of D-Aspartate Metabolism in Regulating Brain and Endocrine System Physiology: From Preclinical Observations to Clinical Applications" International Journal of Molecular Sciences 21, no. 22: 8718. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21228718