Rapamycin Eyedrops Increased CD4+Foxp3+ Cells and Prevented Goblet Cell Loss in the Aged Ocular Surface

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
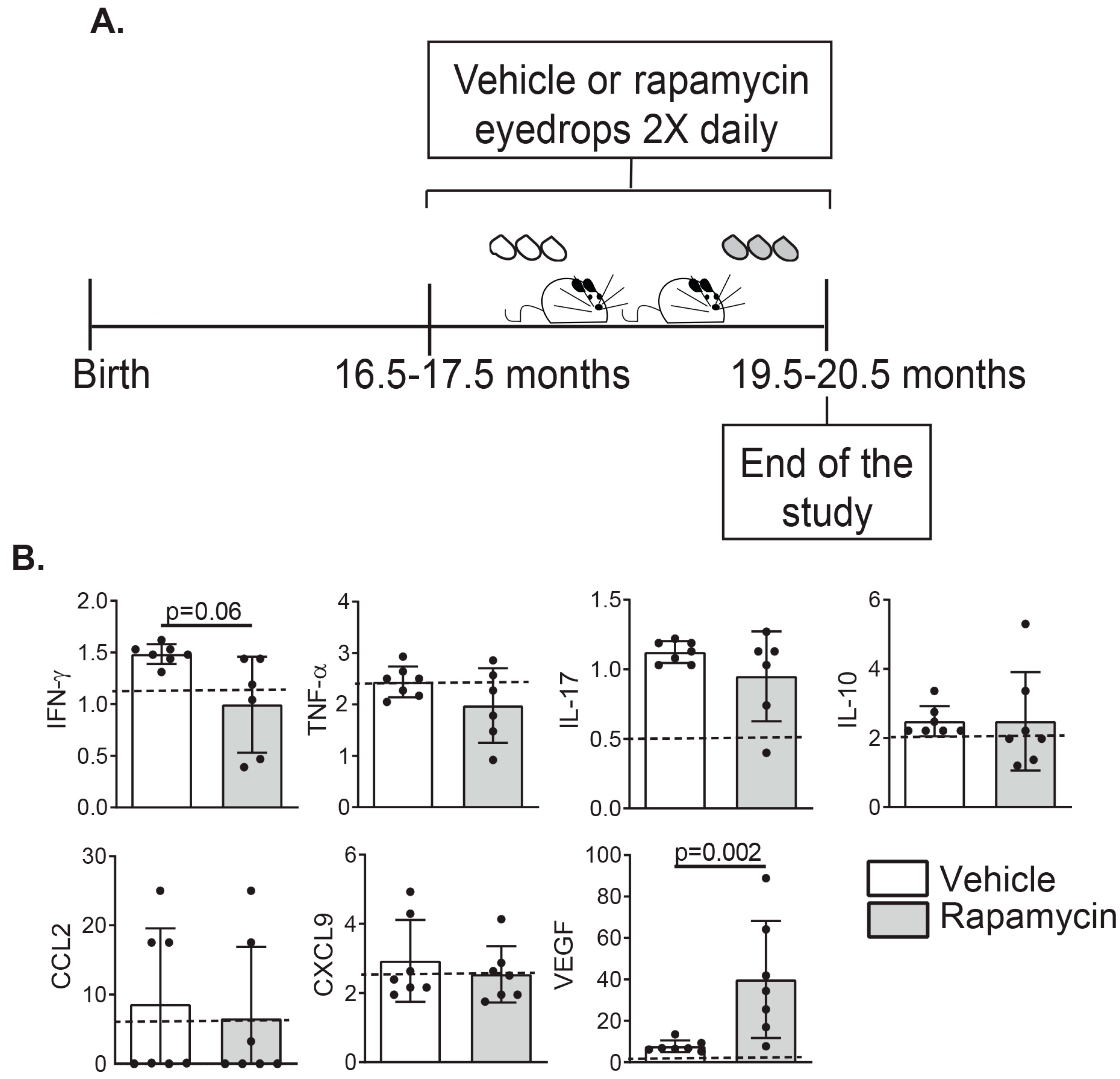
2.1. Rapamycin Eyedrops Can Decrease the Concentration of Effector Lymphocyte Cytokines and Angiogenesis Mediators in Tears of Aged Mice
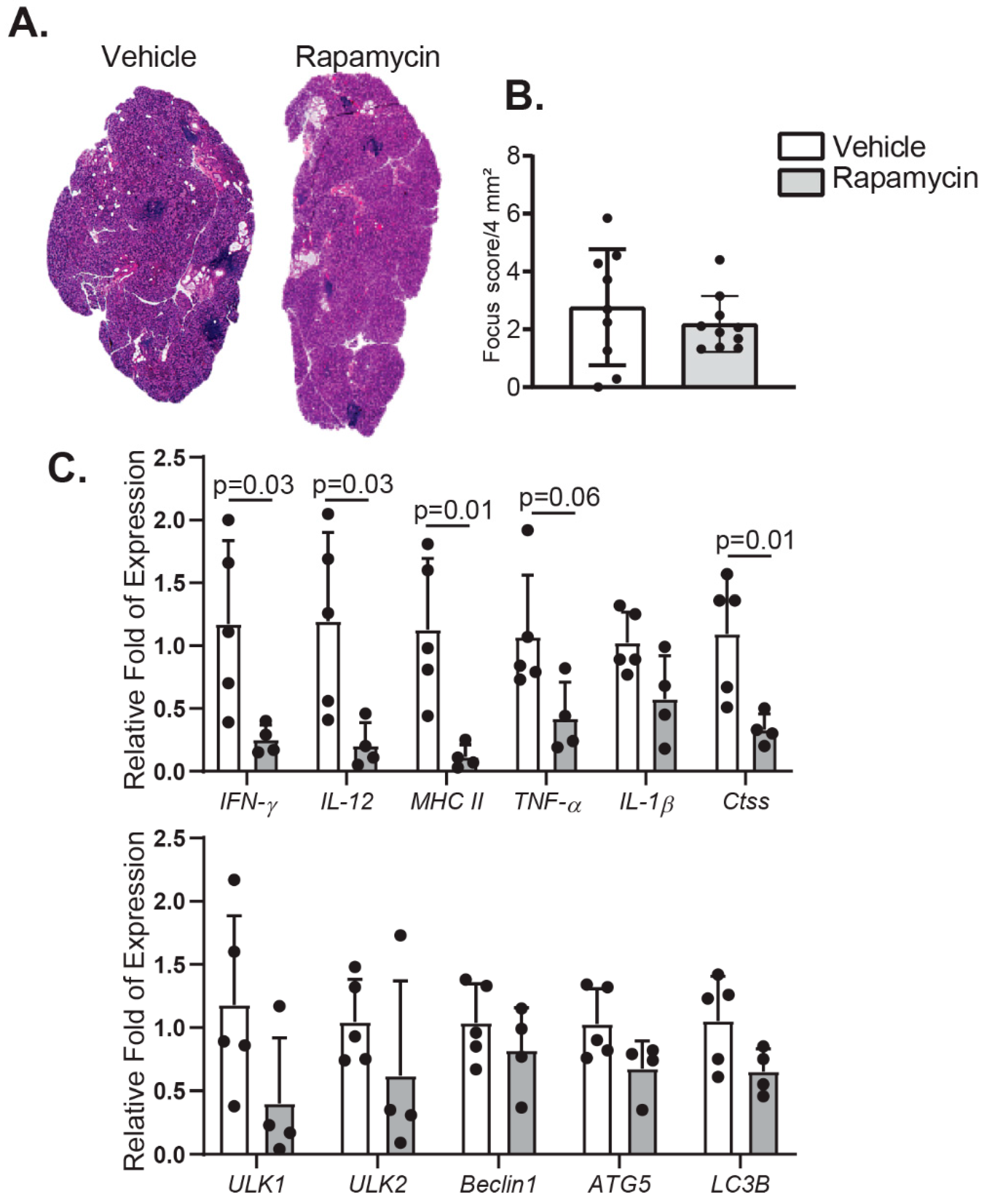
2.2. Rapamycin Eyedrops Decrease Pro-Inflammatory Markers in the Lacrimal Gland of Aged Mice with Minor Impact in Lymphocyte Infiltration and Autophagy
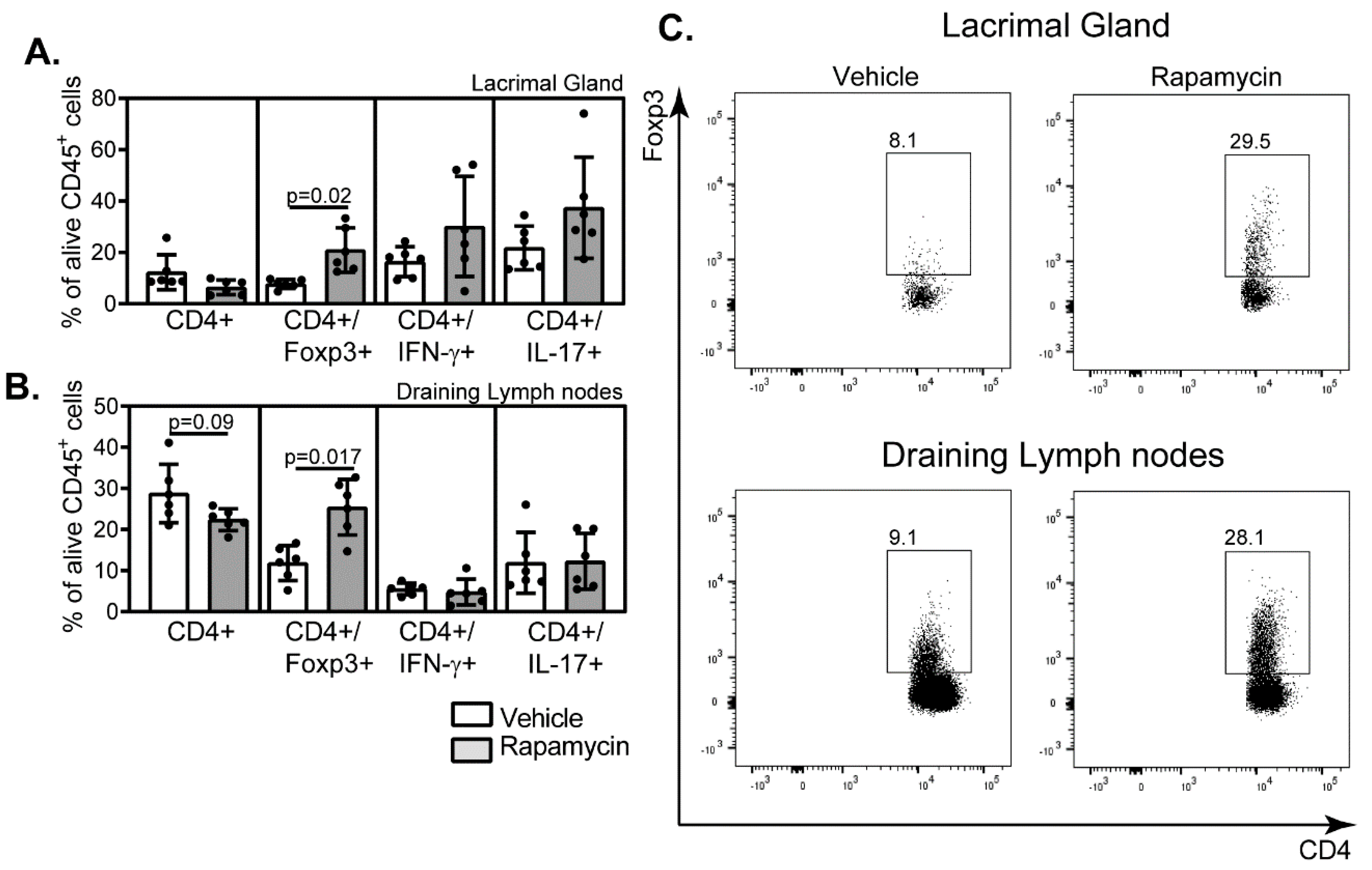
2.3. Rapamycin Eyedrops Skewed the Effector Adaptive Immune Response Phenotype in the Aged Eye
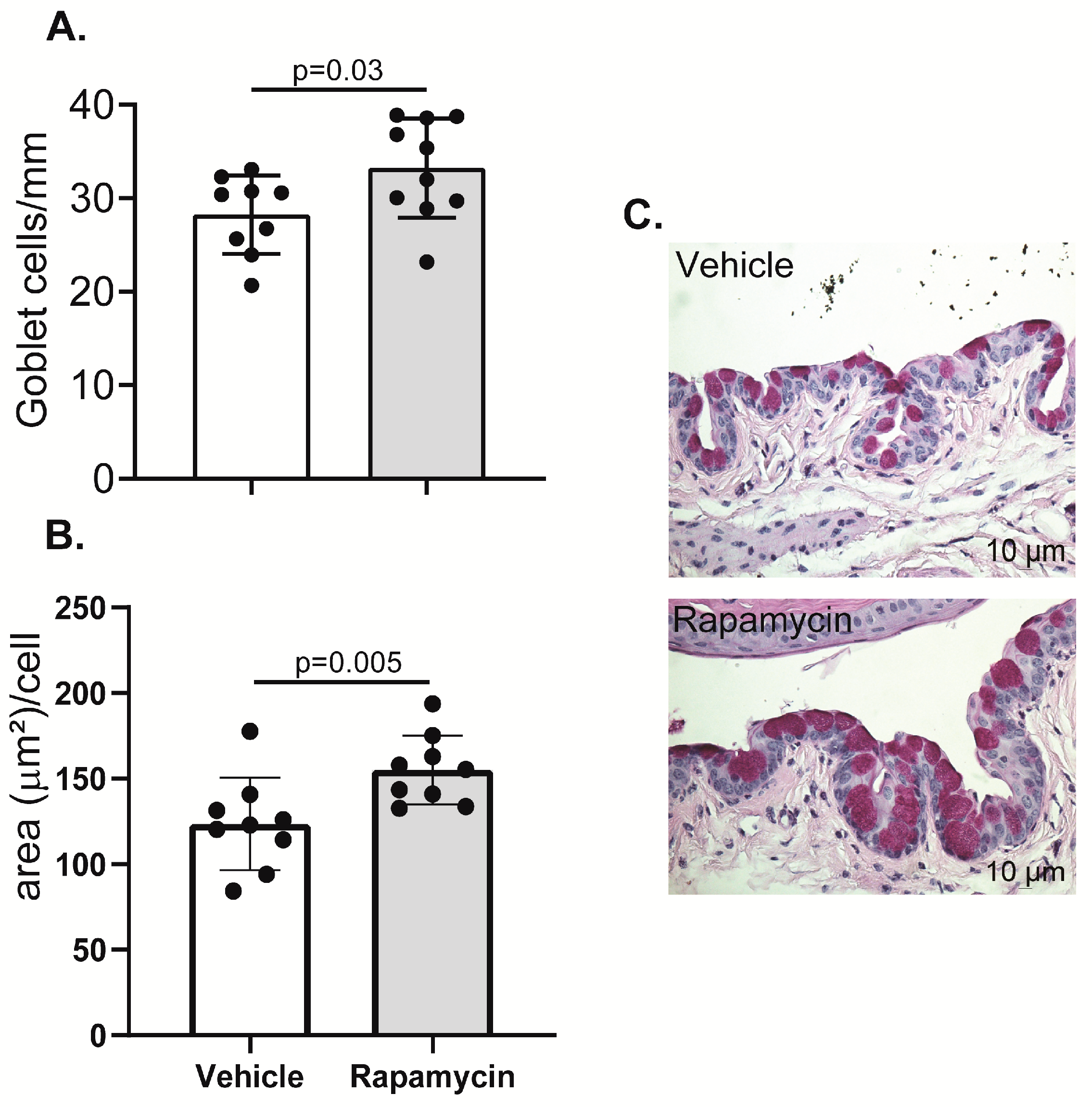
2.4. Goblet Cell Density Increased in the Conjunctiva of Rapamycin-Treated Aged Mice
3. Discussion
4. Materials and Methods
4.1. Reagents
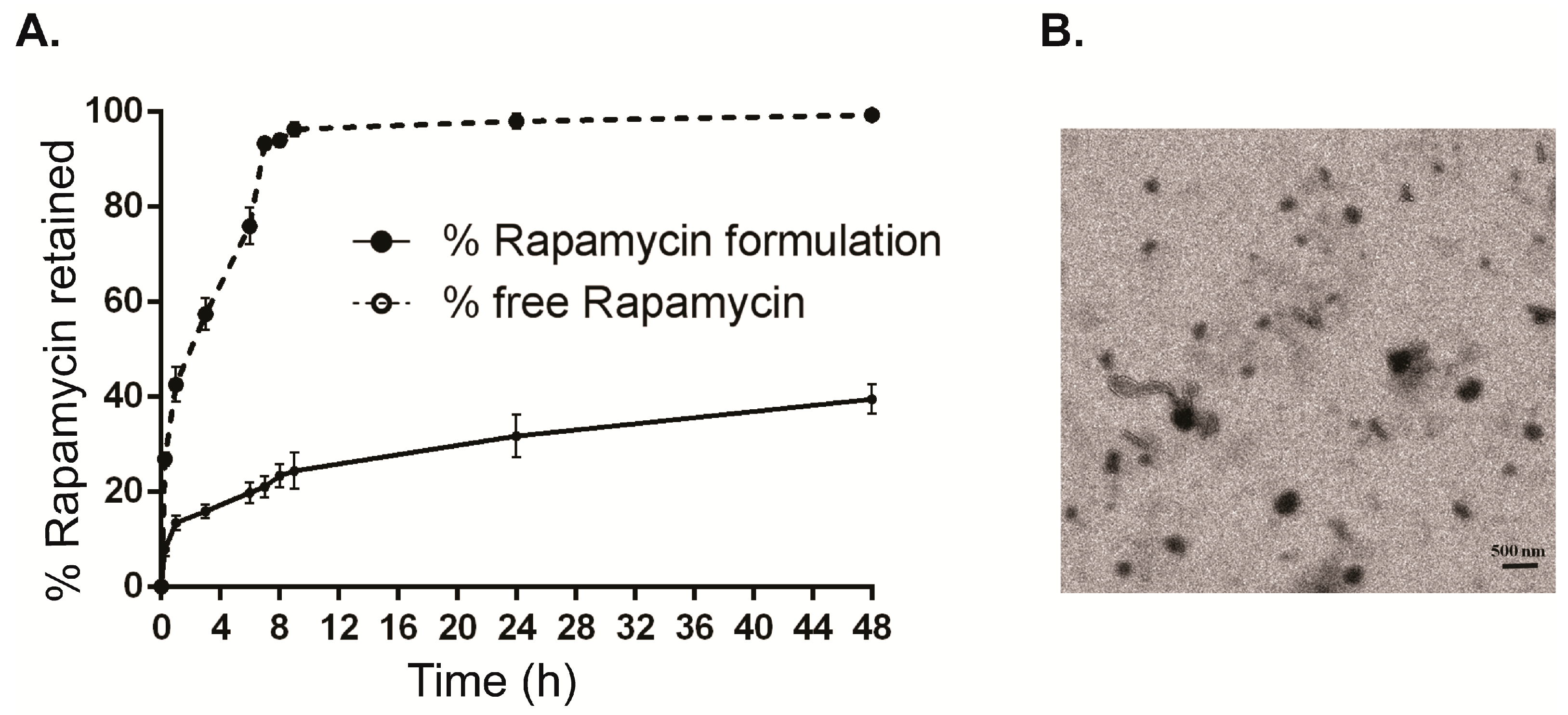
4.2. Preparation and Characterization of Rapamycin Eyedrop Formulation
4.3. Rapamycin Release Studies
4.4. Animals
4.5. Rapamycin or Vehicle Eyedrops Dosing Regime
4.6. Tear Washings and Multiplex Cytokine Immunobead Assay
4.7. RNA Isolation and Real-Time PCR
4.8. Histology, PAS Staining, and Quantification of Focus Score in Lacrimal Glands
4.9. Flow Cytometry Analysis
4.10. Statistical Analyses
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| DED | Dry Eye Disease |
| Th | T helper |
| mTORC1 | Mammalian target of rapamycin complex 1 |
| NOD | Non-obese diabetic |
| TEM | Transmission electron microscopy |
| HPLC | High performance liquid chromatography |
| EE | Encapsulation Efficiency |
| IFN-γ | Interferon gamma |
| TNF-α | Tumor Necrosis Factor alpha |
| IL | Interleukin |
| CCL2 | C-C motif chemokine ligand 2 |
| CXCL9 | Chemokine (C-X-C motif) ligand 9 |
| VEGF-A | Vascular endothelial growth factor-A |
| ULK1 | Unc-51 Like Autophagy Activating Kinase 1 |
| PAS | Periodic Acid Schiff |
| LC3 | Microtubule-associated protein 1A/1B-light chain 3 |
References
- Di Zazzo, A.; Micera, A.; Coassin, M.; Varacalli, G.; Foulsham, W.; De Piano, M.; Bonini, S. InflammAging at Ocular Surface: Clinical and Biomolecular Analyses in Healthy Volunteers. Investig. Ophthalmol. Vis. Sci. 2019, 60, 1769–1775. [Google Scholar] [CrossRef] [Green Version]
- Micera, A.; Di Zazzo, A.; Esposito, G.; Longo, R.; Foulsham, W.; Sacco, R.; Sgrulletta, R.; Bonini, S. Age-Related Changes to Human Tear Composition. Investig. Ophthalmol. Vis. Sci. 2018, 59, 2024–2031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ventura, M.T.; Casciaro, M.; Gangemi, S.; Buquicchio, R. Immunosenescence in aging: Between immune cells depletion and cytokines up-regulation. Clin. Mol. Allergy CMA 2017, 15, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matossian, C.; McDonald, M.; Donaldson, K.E.; Nichols, K.K.; MacIver, S.; Gupta, P.K. Dry Eye Disease: Consideration for Women’s Health. J. Women’s Health 2019, 28, 502–514. [Google Scholar] [CrossRef] [PubMed]
- Mashaghi, A.; Hong, J.; Chauhan, S.K.; Dana, R. Ageing and ocular surface immunity. Br. J. Ophthalmol. 2017, 101, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Schaumburg, C.S.; Siemasko, K.F.; De Paiva, C.S.; Wheeler, L.A.; Niederkorn, J.Y.; Pflugfelder, S.C.; Stern, M.E. Ocular surface APCs are necessary for autoreactive T cell-mediated experimental autoimmune lacrimal keratoconjunctivitis. J. Immunol. 2011, 187, 3653–3662. [Google Scholar] [CrossRef] [Green Version]
- De Paiva, C.S.; Villarreal, A.L.; Corrales, R.M.; Rahman, H.T.; Chang, V.Y.; Farley, W.J.; Stern, M.E.; Niederkorn, J.Y.; Li, D.Q.; Pflugfelder, S.C. Dry eye-induced conjunctival epithelial squamous metaplasia is modulated by interferon-gamma. Investig. Ophthalmol. Vis. Sci. 2007, 48, 2553–2560. [Google Scholar] [CrossRef] [PubMed]
- De Paiva, C.S.; Chotikavanich, S.; Pangelinan, S.B.; Pitcher, J.D., III; Fang, B.; Zheng, X.; Ma, P.; Farley, W.J.; Siemasko, K.F.; Niederkorn, J.Y.; et al. IL-17 disrupts corneal barrier following desiccating stress. Mucosal Immunol. 2009, 2, 243–253. [Google Scholar] [CrossRef] [Green Version]
- Coursey, T.G.; Bian, F.; Zaheer, M.; Pflugfelder, S.C.; Volpe, E.A.; de Paiva, C.S. Age-related spontaneous lacrimal keratoconjunctivitis is accompanied by dysfunctional T regulatory cells. Mucosal Immunol. 2017, 10, 743–756. [Google Scholar] [CrossRef]
- McClellan, A.J.; Volpe, E.A.; Zhang, X.; Darlington, G.J.; Li, D.Q.; Pflugfelder, S.C.; de Paiva, C.S. Ocular surface disease and dacryoadenitis in aging C57BL/6 mice. Am. J. Pathol. 2014, 184, 631–643. [Google Scholar] [CrossRef] [Green Version]
- Volpe, E.A.; Henriksson, J.T.; Wang, C.; Barbosa, F.L.; Zaheer, M.; Zhang, X.; Pflugfelder, S.C.; de Paiva, C.S. Interferon-gamma deficiency protects against aging-related goblet cell loss. Oncotarget 2016, 7, 64605–64614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Souza, R.G.; de Paiva, C.S.; Alves, M.R. Age-related Autoimmune Changes in Lacrimal Glands. Immune Netw. 2019, 19, e3. [Google Scholar] [CrossRef] [PubMed]
- Bian, F.; Xiao, Y.; Barbosa, F.L.; de Souza, R.G.; Hernandez, H.; Yu, Z.; Pflugfelder, S.C.; de Paiva, C.S. Age-associated antigen-presenting cell alterations promote dry-eye inducing Th1 cells. Mucosal Immunol. 2019, 12, 897–908. [Google Scholar] [CrossRef] [PubMed]
- De Silva, M.E.H.; Hill, L.J.; Downie, L.E.; Chinnery, H.R. The Effects of Aging on Corneal and Ocular Surface Homeostasis in Mice. Investig. Ophthalmol. Vis. Sci. 2019, 60, 2705–2715. [Google Scholar] [CrossRef] [Green Version]
- Lamming, D.W.; Ye, L.; Katajisto, P.; Goncalves, M.D.; Saitoh, M.; Stevens, D.M.; Davis, J.G.; Salmon, A.B.; Richardson, A.; Ahima, R.S.; et al. Rapamycin-induced insulin resistance is mediated by mTORC2 loss and uncoupled from longevity. Science 2012, 335, 1638–1643. [Google Scholar] [CrossRef] [Green Version]
- Martel, R.R.; Klicius, J.; Galet, S. Inhibition of the immune response by rapamycin, a new antifungal antibiotic. Can. J. Physiol. Pharmacol. 1977, 55, 48–51. [Google Scholar] [CrossRef]
- Lui, S.L.; Yung, S.; Tsang, R.; Zhang, F.; Chan, K.W.; Tam, S.; Chan, T.M. Rapamycin prevents the development of nephritis in lupus-prone NZB/W F1 mice. Lupus 2008, 17, 305–313. [Google Scholar] [CrossRef]
- Granger, D.K.; Cromwell, J.W.; Chen, S.C.; Goswitz, J.J.; Morrow, D.T.; Beierle, F.A.; Sehgal, S.N.; Canafax, D.M.; Matas, A.J. Prolongation of renal allograft survival in a large animal model by oral rapamycin monotherapy. Transplantation 1995, 59, 183–186. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, F.; Zou, S.; Qu, L. Rapamycin: A Bacteria-Derived Immunosuppressant That Has Anti-atherosclerotic Effects and Its Clinical Application. Front. Pharmacol. 2018, 9, 1520. [Google Scholar] [CrossRef]
- Schinaman, J.M.; Rana, A.; Ja, W.W.; Clark, R.I.; Walker, D.W. Rapamycin modulates tissue aging and lifespan independently of the gut microbiota in Drosophila. Sci. Rep. 2019, 9, 7824. [Google Scholar] [CrossRef] [Green Version]
- Swindell, W.R. Meta-Analysis of 29 Experiments Evaluating the Effects of Rapamycin on Life Span in the Laboratory Mouse. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2017, 72, 1024–1032. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Shen, L.; Yu, J.; Wan, H.; Guo, A.; Chen, J.; Long, Y.; Zhao, J.; Pei, G. Rapamycin and other longevity-promoting compounds enhance the generation of mouse induced pluripotent stem cells. Aging Cell 2011, 10, 908–911. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Singh, S.; Tripathi, V.K.; Bissoyi, A.; Garg, G.; Rizvi, S.I. Rapamycin Confers Neuroprotection Against Aging-Induced Oxidative Stress, Mitochondrial Dysfunction, and Neurodegeneration in Old Rats Through Activation of Autophagy. Rejuvenation Res. 2019, 22, 60–70. [Google Scholar] [CrossRef] [PubMed]
- Kolosova, N.G.; Muraleva, N.A.; Zhdankina, A.A.; Stefanova, N.A.; Fursova, A.Z.; Blagosklonny, M.V. Prevention of age-related macular degeneration-like retinopathy by rapamycin in rats. Am. J. Pathol. 2012, 181, 472–477. [Google Scholar] [CrossRef]
- Stanojlovic, S.; Schlickeiser, S.; Appelt, C.; Vogt, K.; Schmitt-Knosalla, I.; Haase, S.; Ritter, T.; Sawitzki, B.; Pleyer, U. Influence of combined treatment of low dose rapamycin and cyclosporin A on corneal allograft survival. Graefe’s Arch. Clin. Exp. Ophthalmol. 2010, 248, 1447–1456. [Google Scholar] [CrossRef]
- Birnbaum, F.; Reis, A.; Bohringer, D.; Sokolowska, Y.; Mayer, K.; Voiculescu, A.; Oellerich, M.; Sundmacher, R.; Reinhard, T. An open prospective pilot study on the use of rapamycin after penetrating high-risk keratoplasty. Transplantation 2006, 81, 767–772. [Google Scholar] [CrossRef]
- Gidfar, S.; Milani, F.Y.; Milani, B.Y.; Shen, X.; Eslani, M.; Putra, I.; Huvard, M.J.; Sagha, H.; Djalilian, A.R. Rapamycin Prolongs the Survival of Corneal Epithelial Cells in Culture. Sci. Rep. 2017, 7, 40308. [Google Scholar] [CrossRef] [Green Version]
- Johnson, S.C.; Kaeberlein, M. Rapamycin in aging and disease: Maximizing efficacy while minimizing side effects. Oncotarget 2016, 7, 44876–44878. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Truong, T.; Ladiges, W.; Goh, J. Rapamycin increases breast tumor burden in young wheel-running mice. Pathobiol. Aging Age Relat. Dis. 2019, 9, 1647746. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Wu, X.; Duan, J.; Hinrichs, D.; Wegmann, K.; Zhang, G.L.; Hall, M.; Rosenbaum, J.T. Low dose rapamycin exacerbates autoimmune experimental uveitis. PLoS ONE 2012, 7, e36589. [Google Scholar] [CrossRef] [Green Version]
- Sutter, D.; Dzhonova, D.V.; Prost, J.C.; Bovet, C.; Banz, Y.; Rahnfeld, L.; Leroux, J.C.; Rieben, R.; Vogelin, E.; Plock, J.A.; et al. Delivery of Rapamycin Using In Situ Forming Implants Promotes Immunoregulation and Vascularized Composite Allograft Survival. Sci. Rep. 2019, 9, 9269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thapa, R.K.; Nguyen, H.T.; Jeong, J.H.; Kim, J.R.; Choi, H.G.; Yong, C.S.; Kim, J.O. Progressive slowdown/prevention of cellular senescence by CD9-targeted delivery of rapamycin using lactose-wrapped calcium carbonate nanoparticles. Sci. Rep. 2017, 7, 43299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lesovaya, E.A.; Kirsanov, K.I.; Antoshina, E.E.; Trukhanova, L.S.; Gorkova, T.G.; Shipaeva, E.V.; Salimov, R.M.; Belitsky, G.A.; Blagosklonny, M.V.; Yakubovskaya, M.G.; et al. Rapatar, a nanoformulation of rapamycin, decreases chemically-induced benign prostate hyperplasia in rats. Oncotarget 2015, 6, 9718–9727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Xu, L.; Chen, H.; Li, X. Rapamycin-loaded poly(epsilon-caprolactone)-poly(ethylene glycol)-poly(epsilon-caprolactone) nanoparticles: Preparation, characterization and potential application in corneal transplantation. J. Pharm. Pharmacol. 2014, 66, 557–563. [Google Scholar] [CrossRef]
- Shah, M.; Edman, M.C.; Janga, S.R.; Shi, P.; Dhandhukia, J.; Liu, S.; Louie, S.G.; Rodgers, K.; Mackay, J.A.; Hamm-Alvarez, S.F. A rapamycin-binding protein polymer nanoparticle shows potent therapeutic activity in suppressing autoimmune dacryoadenitis in a mouse model of Sjogren’s syndrome. J. Control. Rel. Off. J. Control. Release Soc. 2013, 171, 269–279. [Google Scholar] [CrossRef] [Green Version]
- Shah, M.; Edman, M.C.; Reddy Janga, S.; Yarber, F.; Meng, Z.; Klinngam, W.; Bushman, J.; Ma, T.; Liu, S.; Louie, S.; et al. Rapamycin Eye Drops Suppress Lacrimal Gland Inflammation In a Murine Model of Sjogren’s Syndrome. Investig. Ophthalmol. Vis. Sci. 2017, 58, 372–385. [Google Scholar] [CrossRef]
- Wang, S.; Wang, M.; Liu, Y.; Hu, D.; Gu, L.; Fei, X.; Zhang, J. Effect of Rapamycin Microspheres in Sjogren Syndrome Dry Eye: Preparation and Outcomes. Ocul. Immunol. Inflamm. 2019, 27, 1357–1364. [Google Scholar] [CrossRef]
- Blagosklonny, M.V. Rapamycin for longevity: Opinion article. Aging 2019, 11, 8048–8067. [Google Scholar] [CrossRef]
- Lam, H.; Bleiden, L.; de Paiva, C.S.; Farley, W.; Stern, M.E.; Pflugfelder, S.C. Tear cytokine profiles in dysfunctional tear syndrome. Am. J. Ophthalmol. 2009, 147, 198–205.e191. [Google Scholar] [CrossRef] [Green Version]
- Enriquez-de-Salamanca, A.; Castellanos, E.; Stern, M.E.; Fernandez, I.; Carreno, E.; Garcia-Vazquez, C.; Herreras, J.M.; Calonge, M. Tear cytokine and chemokine analysis and clinical correlations in evaporative-type dry eye disease. Mol. Vis. 2010, 16, 862–873. [Google Scholar]
- Dodd, K.M.; Yang, J.; Shen, M.H.; Sampson, J.R.; Tee, A.R. mTORC1 drives HIF-1α and VEGF-A signalling via multiple mechanisms involving 4E-BP1, S6K1 and STAT3. Oncogene 2015, 34, 2239–2250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocha, E.M.; Alves, M.; Rios, J.D.; Dartt, D.A. The aging lacrimal gland: Changes in structure and function. Ocul. Surf. 2008, 6, 162–174. [Google Scholar] [CrossRef] [Green Version]
- Obata, H.; Yamamoto, S.; Horiuchi, H.; Machinami, R. Histopathologic study of human lacrimal gland. Statistical analysis with special reference to aging. Ophthalmology 1995, 102, 678–686. [Google Scholar] [CrossRef]
- Zachari, M.; Ganley, I.G. The mammalian ULK1 complex and autophagy initiation. Essays Biochem. 2017, 61, 585–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, C.H.; Ryu, J.S.; Hwang, H.S.; Kim, M.K. Comparative Analysis of Age-Related Changes in Lacrimal Glands and Meibomian Glands of a C57BL/6 Male Mouse Model. Int. J. Mol. Sci. 2020, 21, 4169. [Google Scholar] [CrossRef] [PubMed]
- Contreras-Ruiz, L.; Ghosh-Mitra, A.; Shatos, M.A.; Dartt, D.A.; Masli, S. Modulation of conjunctival goblet cell function by inflammatory cytokines. Mediat. Inflamm. 2013, 2013, 636812. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Y.; de Paiva, C.S.; Yu, Z.; de Souza, R.G.; Li, D.Q.; Pflugfelder, S.C. Goblet cell-produced retinoic acid suppresses CD86 expression and IL-12 production in bone marrow-derived cells. Int. Immunol. 2018, 30, 457–470. [Google Scholar] [CrossRef] [PubMed]
- Alam, J.; de Paiva, C.S.; Pflugfelder, S.C. Immune—Goblet cell interaction in the conjunctiva. Ocul. Surf. 2020, 18, 236–334. [Google Scholar] [CrossRef]
- De Paiva, C.S.; Raince, J.K.; McClellan, A.J.; Shanmugam, K.P.; Pangelinan, S.B.; Volpe, E.A.; Corrales, R.M.; Farley, W.J.; Corry, D.B.; Li, D.Q.; et al. Homeostatic control of conjunctival mucosal goblet cells by NKT-derived IL-13. Mucosal Immunol. 2011, 4, 397–408. [Google Scholar] [CrossRef]
- Majumder, S.; Caccamo, A.; Medina, D.X.; Benavides, A.D.; Javors, M.A.; Kraig, E.; Strong, R.; Richardson, A.; Oddo, S. Lifelong rapamycin administration ameliorates age-dependent cognitive deficits by reducing IL-1beta and enhancing NMDA signaling. Aging Cell 2012, 11, 326–335. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Wu, K.; Edman, M.; Schenke-Layland, K.; MacVeigh-Aloni, M.; Janga, S.R.; Schulz, B.; Hamm-Alvarez, S.F. Increased expression of cathepsins and obesity-induced proinflammatory cytokines in lacrimal glands of male NOD mouse. Investig. Ophthalmol. Vis. Sci. 2010, 51, 5019–5029. [Google Scholar] [CrossRef] [PubMed]
- Kopf, H.; de la Rosa, G.M.; Howard, O.M.; Chen, X. Rapamycin inhibits differentiation of Th17 cells and promotes generation of FoxP3+ T regulatory cells. Int. Immunopharmacol. 2007, 7, 1819–1824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basu, S.; Golovina, T.; Mikheeva, T.; June, C.H.; Riley, J.L. Cutting edge: Foxp3-mediated induction of pim 2 allows human T regulatory cells to preferentially expand in rapamycin. J. Immunol. 2008, 180, 5794–5798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Battaglia, M.; Stabilini, A.; Migliavacca, B.; Horejs-Hoeck, J.; Kaupper, T.; Roncarolo, M.G. Rapamycin promotes expansion of functional CD4+CD25+FOXP3+ regulatory T cells of both healthy subjects and type 1 diabetic patients. J. Immunol. 2006, 177, 8338–8347. [Google Scholar] [CrossRef] [Green Version]
- Turnquist, H.R.; Raimondi, G.; Zahorchak, A.F.; Fischer, R.T.; Wang, Z.; Thomson, A.W. Rapamycin-conditioned dendritic cells are poor stimulators of allogeneic CD4+ T cells, but enrich for antigen-specific Foxp3+ T regulatory cells and promote organ transplant tolerance. J. Immunol. 2007, 178, 7018–7031. [Google Scholar] [CrossRef] [Green Version]
- Mushaben, E.M.; Kramer, E.L.; Brandt, E.B.; Khurana Hershey, G.K.; Le Cras, T.D. Rapamycin attenuates airway hyperreactivity, goblet cells, and IgE in experimental allergic asthma. J. Immunol. 2011, 187, 5756–5763. [Google Scholar] [CrossRef] [Green Version]
- Sampson, L.L.; Davis, A.K.; Grogg, M.W.; Zheng, Y. mTOR disruption causes intestinal epithelial cell defects and intestinal atrophy postinjury in mice. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2016, 30, 1263–1275. [Google Scholar] [CrossRef] [Green Version]
- Alcalde, I.; Íñigo-Portugués, A.; González-González, O.; Almaraz, L.; Artime, E.; Morenilla-Palao, C.; Gallar, J.; Viana, F.; Merayo-Lloves, J.; Belmonte, C. Morphological and functional changes in TRPM8-expressing corneal cold thermoreceptor neurons during aging and their impact on tearing in mice. J. Comp. Neurol. 2018, 526, 1859–1874. [Google Scholar] [CrossRef]
- Harpaz, I.; Bhattacharya, U.; Elyahu, Y.; Strominger, I.; Monsonego, A. Old Mice Accumulate Activated Effector CD4 T Cells Refractory to Regulatory T Cell-Induced Immunosuppression. Front. Immunol. 2017, 8, 283. [Google Scholar] [CrossRef] [Green Version]
- Tanida, I.; Ueno, T.; Kominami, E. LC3 and Autophagy. Meth. Mol. Biol. 2008, 445, 77–88. [Google Scholar] [CrossRef]
- Cho, B.J.; Hwang, J.S.; Shin, Y.J.; Kim, J.W.; Chung, T.Y.; Hyon, J.Y. Rapamycin Rescues Endoplasmic Reticulum Stress-Induced Dry Eye Syndrome in Mice. Investig. Ophthalmol. Vis. Sci. 2019, 60, 1254–1264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Chen, W.; Gao, Q.; Yang, J.; Yan, X.; Zhao, H.; Su, L.; Yang, M.; Gao, C.; Yao, Y.; et al. Rapamycin directly activates lysosomal mucolipin TRP channels independent of mTOR. PLoS Biol. 2019, 17, e3000252. [Google Scholar] [CrossRef] [PubMed]
- Guaiquil, V.H.; Pan, Z.; Karagianni, N.; Fukuoka, S.; Alegre, G.; Rosenblatt, M.I. VEGF-B selectively regenerates injured peripheral neurons and restores sensory and trophic functions. Proc. Natl. Acad. Sci. USA 2014, 111, 17272–17277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Burns, A.R.; Han, L.; Rumbaut, R.E.; Smith, C.W. IL-17 and VEGF are necessary for efficient corneal nerve regeneration. Am. J. Pathol. 2011, 178, 1106–1116. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; Guo, H.; Klinngam, W.; Janga, S.R.; Yarber, F.; Peddi, S.; Edman, M.C.; Tiwari, N.; Liu, S.; Louie, S.G.; et al. Berunda Polypeptides: Biheaded Rapamycin Carriers for Subcutaneous Treatment of Autoimmune Dry Eye Disease. Mol. Pharm. 2019, 16, 3024–3039. [Google Scholar] [CrossRef] [PubMed]
- Ju, Y.; Guo, H.; Yarber, F.; Edman, M.C.; Peddi, S.; Janga, S.R.; MacKay, J.A.; Hamm-Alvarez, S.F. Molecular Targeting of Immunosuppressants Using a Bifunctional Elastin-Like Polypeptide. Bioconjug. Chem. 2019, 30, 2358–2372. [Google Scholar] [CrossRef]
- Aliper, A.; Jellen, L.; Cortese, F.; Artemov, A.; Karpinsky-Semper, D.; Moskalev, A.; Swick, A.G.; Zhavoronkov, A. Towards natural mimetics of metformin and rapamycin. Aging 2017, 9, 2245–2268. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Maksoud, M.S.; El-Gamal, M.I.; Benhalilou, D.R.; Ashraf, S.; Mohammed, S.A.; Oh, C.H. Mechanistic/mammalian target of rapamycin: Recent pathological aspects and inhibitors. Med. Res. Rev. 2019, 39, 631–664. [Google Scholar] [CrossRef]
- National Research Council Committee for the Update of the Guide for the Care; Use of Laboratory Animals. The National Academies Collection: Reports funded by National Institutes of Health. In Guide for the Care and Use of Laboratory Animals; National Academies Press (US): Washington, DC, USA, 2011. [Google Scholar] [CrossRef]
- De Paiva, C.S.; Yoon, K.C.; Pangelinan, S.B.; Pham, S.; Puthenparambil, L.M.; Chuang, E.Y.; Farley, W.J.; Stern, M.E.; Li, D.Q.; Pflugfelder, S.C. Cleavage of functional IL-2 receptor alpha chain (CD25) from murine corneal and conjunctival epithelia by MMP-9. J. Inflamm. 2009, 6, 31. [Google Scholar] [CrossRef] [Green Version]
- Coursey, T.G.; Bohat, R.; Barbosa, F.L.; Pflugfelder, S.C.; de Paiva, C.S. Desiccating stress-induced chemokine expression in the epithelium is dependent on upregulation of NKG2D/RAE-1 and release of IFN-gamma in experimental dry eye. J. Immunol. 2014, 193, 5264–5272. [Google Scholar] [CrossRef] [Green Version]
- Zaheer, M.; Wang, C.; Bian, F.; Yu, Z.; Hernandez, H.; de Souza, R.G.; Simmons, K.T.; Schady, D.; Swennes, A.G.; Pflugfelder, S.C.; et al. Protective role of commensal bacteria in Sjogren Syndrome. J. Autoimmun. 2018, 93, 45–56. [Google Scholar] [CrossRef] [PubMed]
- You, I.C.; Bian, F.; Volpe, E.A.; de Paiva, C.S.; Pflugfelder, S.C. Age-Related Conjunctival Disease in the C57BL/6.NOD-Aec1Aec2 Mouse Model of Sjogren Syndrome Develops Independent of Lacrimal Dysfunction. Investig. Ophthalmol. Vis. Sci. 2015, 56, 2224–2233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Zaheer, M.; Bian, F.; Quach, D.; Swennes, A.G.; Britton, R.A.; Pflugfelder, S.C.; de Paiva, C.S. Sjogren-Like Lacrimal Keratoconjunctivitis in Germ-Free Mice. Int. J. Mol. Sci. 2018, 19, 565. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trujillo-Vargas, C.M.; Kutlehria, S.; Hernandez, H.; de Souza, R.G.; Lee, A.; Yu, Z.; Pflugfelder, S.C.; Singh, M.; de Paiva, C.S. Rapamycin Eyedrops Increased CD4+Foxp3+ Cells and Prevented Goblet Cell Loss in the Aged Ocular Surface. Int. J. Mol. Sci. 2020, 21, 8890. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21238890
Trujillo-Vargas CM, Kutlehria S, Hernandez H, de Souza RG, Lee A, Yu Z, Pflugfelder SC, Singh M, de Paiva CS. Rapamycin Eyedrops Increased CD4+Foxp3+ Cells and Prevented Goblet Cell Loss in the Aged Ocular Surface. International Journal of Molecular Sciences. 2020; 21(23):8890. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21238890
Chicago/Turabian StyleTrujillo-Vargas, Claudia M., Shallu Kutlehria, Humberto Hernandez, Rodrigo G. de Souza, Andrea Lee, Zhiyuan Yu, Stephen C. Pflugfelder, Mandip Singh, and Cintia S. de Paiva. 2020. "Rapamycin Eyedrops Increased CD4+Foxp3+ Cells and Prevented Goblet Cell Loss in the Aged Ocular Surface" International Journal of Molecular Sciences 21, no. 23: 8890. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21238890