Influence of Microgravity on Apoptosis in Cells, Tissues, and Other Systems In Vivo and In Vitro


 , , , ,
, , , , 
Abstract
:1. Introduction
2. Spaceflight Options and Ground-Based Techniques Suitable for Microgravity-Based Research
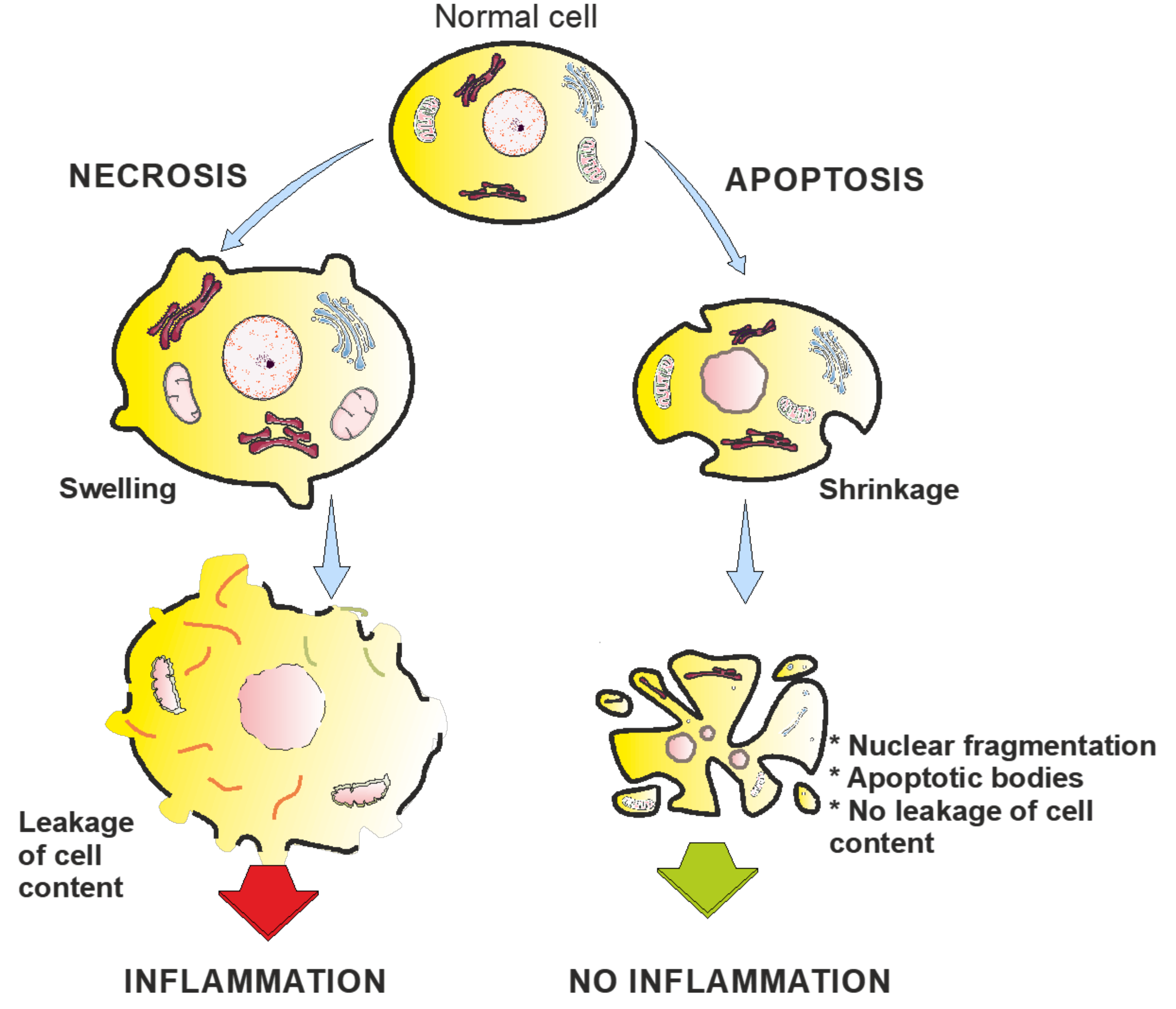
3. Apoptosis
4. Apoptosis Research Using the HS Rat Model
5. Murine Experiments in the Space Environment
6. Impact of Microgravity on Apoptosis in Different Cell Types
6.1. Benign Specialized Cells
6.1.1. Immune Cells
6.1.2. (Cardio)Vascular System
6.1.3. Brain and Eye Cells
6.1.4. Chondrocytes and Bone Cells
6.1.5. Gastrointestinal, Gastric, Liver, and Pancreatic Cells
6.2. Cancer Cells

{kind=link}
{kind=link}
| Tumor/Cell Line | Microgravity Conditions | Apoptosis Findings | Ref. |
|---|---|---|---|
| Thyroid cancer | |||
| ML-1 | RPM, 24 h | p53↑, Fas↑, Bax↑, Bcl-2↓ | [6] |
| FTC-133 | RPM, 72 h | No apoptosis detected | [165] |
| ONCO-DG1 | CN, 24 h | Bax↑, activated caspase-3, Bcl-2↓, TUNEL positive | [151] |
| Breast cancer | |||
| MCF-7 | RPM, 24 h | p53↑, Fas↑, Bax↑, caspase-8↑ | [166] |
| RPM, 48 h | No apoptosis detected | [163] | |
| MDA-MB-231 | RPM, 2 h | TUNEL negative | [167] |
| RPM, 72 h | Bax↑, Bcl-2↓ | [152] | |
| RCCS, 7 d | Bcl-2↓, MMP9↓, cyclin D3↑ | [160] | |
| Colorectal cancer | |||
| DLD-1 | RCCS, 48 h | PTEN↑, FOXO3↑, AKT↓ | [161] |
| MIP-101 | real microgravity, 10/12 d | EGFR↓, TGF-α↓, TGF-β↓ | [159] |
| Gastric cancer | |||
| SGC-7901 | CN, 72 h | TUNEL positive | [149] |
| Lung cancer | |||
| CRL-5889 | RPM, 72 h | TUNEL positive | [164] |
| Hepatoblastoma | |||
| HepG2 | 3D-CN, 72 h | BAX↓, CDKN1A↓, PTEN↑, DRAM1↑, PRKAA1↑ | [168] |
| Glioma | |||
| U251 | CN, 72h | p21↑, IGFBP-2↓ | [124] |
| CN, 24 h | Bcl-2↓, Bnip3↓, cleaved caspases 3/9↑ | [169] | |
| C6 | RPM, 1-24 h | Translocation of Bax, Bcl-2 | [170] |
| Melanoma | |||
| BL6-10 | CN, 24 h | Bcl-2↓, Bnip3↓, caspases 3/7/8↑ | [171] |
7. The Impact of Cosmic Radiation on Apoptosis
8. Summary and Perspectives
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 2D | Two-dimensional |
| 3D | Three-dimensional |
| 5-LOX | Arachidonate 5-lipoxygenase |
| AEM | Animal Enclosure Module |
| Apaf-1 | Apoptosis protease activating factor-1 |
| APO-1 | Apoptosis antigen 1 |
| BAECs | Bovine aortic endothelial cells |
| DISC | Death-inducing signalling complex |
| DLR | Deutsches Zentrum für Luft- und Raumfahrt |
| ESA | European Space Agency |
| FADD | Fas-associated protein with a death domain |
| HDBR | Head-down bed rest |
| HS | Hindlimb suspension |
| HOB | Human osteoblastic |
| HUVEC | human umbilical vein endothelial cells |
| HZE | High (H) atomic number (Z) and energy (E) |
| IL | Interleukin |
| ISS | International Space Station |
| LET | Linear energy transfer |
| MCS | Multicellular spheroid(s) |
| MDA-MB231 | M.D. Anderson-Metastasis Breast cancer cell line |
| MDS | Mice Drawer System |
| mRNA | messenger RNA |
| miRNA | microRNA |
| mTOR | Mammalian target of rapamycin |
| NASA | National Aeronautics and Space Administration |
| NOX2 | Nicotinamide adenine dinucleotide phosphate oxidase 2 |
| NK | Natural killer |
| NF-κB | Nuclear factor kappa B |
| OPN | Osteopontin |
| PARP | Poly (ADP-ribose) polymerase |
| PFA | Paraformaldehyde |
| PBLs | Peripheral blood lymphocytes |
| qPCR | Quantitative polymerase chain reaction |
| RPM | Random positioning machine |
| RANKL | Receptor Activator of NF-κB Ligand |
| RCCS-HARV | Rotational Cell Culture System-High Aspect Ratio Vessel |
| ROS | Reactive oxygen species |
| RWV | Rotating-wall vessels |
| RIP | Receptor-interacting protein |
| RPE | Retinal pigment epithelial |
| RPM | Random positioning machine |
| SRF | Serum Response Factor |
| SOD | Superoxide dismutase |
| SODD | Silencer of death domain |
| STS | Space transportation system |
| TNBC | Triple-negative breast cancer |
| TUNEL | Terminal deoxynucleotidyl transferase-mediated nick end-labeling |
| TNF | Tumor necrosis factor |
| TRADD | TNFR-associated protein with a death domain |
| TS | Tubular structure |
| WHO | World Health Organization |
| XIAP | X-linked inhibitor of apoptosis |
References
- White, R.J.; Averner, M. Humans in space. Nature 2001, 409, 1115–1118. [Google Scholar] [CrossRef] [PubMed]
- Grimm, D.; Wehland, M.; Corydon, T.J.; Richter, P.; Prasad, B.; Bauer, J.; Egli, M.; Kopp, S.; Lebert, M.; Krüger, M. The effects of microgravity on differentiation and cell growth in stem cells and cancer stem cells. Stem. Cells Transl. Med. 2020, 9, 882–894. [Google Scholar] [CrossRef] [PubMed]
- Strauch, S.M.; Grimm, D.; Corydon, T.J.; Krüger, M.; Bauer, J.; Lebert, M.; Wise, P.; Infanger, M.; Richter, P. Current Knowledge about the Impact of Microgravity on the Proteome. Expert Rev. Proteom. 2018. [Google Scholar] [CrossRef] [PubMed]
- Dang, B.; Yang, Y.; Zhang, E.; Li, W.; Mi, X.; Meng, Y.; Yan, S.; Wang, Z.; Wei, W.; Shao, C.; et al. Simulated microgravity increases heavy ion radiation-induced apoptosis in human B lymphoblasts. Life Sci. 2014, 97, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Gridley, D.S.; Mao, X.W.; Stodieck, L.S.; Ferguson, V.L.; Bateman, T.A.; Moldovan, M.; Cunningham, C.E.; Jones, T.A.; Slater, J.M.; Pecaut, M.J. Changes in mouse thymus and spleen after return from the STS-135 mission in space. PLoS ONE 2013, 8, e75097. [Google Scholar] [CrossRef]
- Grimm, D.; Bauer, J.; Kossmehl, P.; Shakibaei, M.; Schöberger, J.; Pickenhahn, H.; Schulze-Tanzil, G.; Vetter, R.; Eilles, C.; Paul, M.; et al. Simulated microgravity alters differentiation and increases apoptosis in human follicular thyroid carcinoma cells. Faseb J. 2002, 16, 604–606. [Google Scholar] [CrossRef] [PubMed]
- Lewis, M.L.; Reynolds, J.L.; Cubano, L.A.; Hatton, J.P.; Lawless, B.D.; Piepmeier, E.H. Spaceflight alters microtubules and increases apoptosis in human lymphocytes (Jurkat). Faseb J. 1998, 12, 1007–1018. [Google Scholar] [CrossRef]
- Ohnishi, T.; Takahashi, A.; Wang, X.; Ohnishi, K.; Ohira, Y.; Nagaoka, S. Accumulation of a tumor suppressor p53 protein in rat muscle during a space flight. Mutat. Res. 1999, 430, 271–274. [Google Scholar] [CrossRef]
- Radugina, E.A.; Almeida, E.A.C.; Blaber, E.; Poplinskaya, V.A.; Markitantova, Y.V.; Grigoryan, E.N. Exposure to microgravity for 30 days onboard Bion M1 caused muscle atrophy and impaired regeneration in murine femoral Quadriceps. Life Sci. Space Res. 2018, 16, 18–25. [Google Scholar] [CrossRef]
- Uva, B.M.; Masini, M.A.; Sturla, M.; Prato, P.; Passalacqua, M.; Giuliani, M.; Tagliafierro, G.; Strollo, F. Clinorotation-induced weightlessness influences the cytoskeleton of glial cells in culture. Brain Res. 2002, 934, 132–139. [Google Scholar] [CrossRef]
- Infanger, M.; Kossmehl, P.; Shakibaei, M.; Baatout, S.; Witzing, A.; Grosse, J.; Bauer, J.; Cogoli, A.; Faramarzi, S.; Derradji, H.; et al. Induction of three-dimensional assembly and increase in apoptosis of human endothelial cells by simulated microgravity: Impact of vascular endothelial growth factor. Apoptosis 2006, 11, 749–764. [Google Scholar] [CrossRef] [PubMed]
- Schoenberger, J.; Bauer, J.; Moosbauer, J.; Eilles, C.; Grimm, D. Innovative strategies in in vivo apoptosis imaging. CMC 2008, 15, 187–194. [Google Scholar] [CrossRef]
- Riwaldt, S.; Monici, M.; Graver Petersen, A.; Birk Jensen, U.; Evert, K.; Pantalone, D.; Utpatel, K.; Evert, M.; Wehland, M.; Krüger, M.; et al. Preparation of A Spaceflight: Apoptosis Search in Sutured Wound Healing Models. Int. J. Med. Sci. 2017, 18, 2604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kerr, J.F.; Wyllie, A.H.; Currie, A.R. Apoptosis: A basic biological phenomenon with wide-ranging implications in tissue kinetics. Br. J. Cancer 1972, 26, 239–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hemmersbach, R.; Häder, D.-P.; Braun, M. Methods for Gravitational Biology Research. In Gravitational Biology I; Braun, M., Böhmer, M., Häder, D.-P., Hemmersbach, R., Palme, K., Eds.; Springer International Publishing: Cham, Switherlands, 2018; pp. 13–26. ISBN 978-3-319-93893-6. [Google Scholar]
- Sabbatini, M. Introduction. In ESA User Guide to Low Gravity Platforms, 3rd ed.; Sentse, N., Oldenburg, K., Eds.; European Space Agency: Noordwijk, The Netherlands, 2014. [Google Scholar]
- Amselem, S. Remote Controlled Autonomous Microgravity Lab Platforms for Drug Research in Space. Pharm. Res. 2019, 36, 183. [Google Scholar] [CrossRef] [PubMed]
- Prasad, B.; Richter, P.; Vadakedath, N.; Mancinelli, R.; Krüger, M.; Strauch, S.M.; Grimm, D.; Darriet, P.; Chapel, J.-P.; Cohen, J.; et al. Exploration of space to achieve scientific breakthroughs. Biotechnol. Adv. 2020, 43, 107572. [Google Scholar] [CrossRef]
- Eigenbrod, C. Drop Tower Bremen—User’s guide. Zarm Drop Tower Bremen. User Mannual. 2011. Available online: http://www.zarm.uni-bremen.de/drop-tower/downloads/ (accessed on 26 October 2020).
- Von Kampen, P.; Kaczmarczik, U.; Rath, H.J. The new Drop Tower catapult system. Acta Astronaut. 2006, 59, 278–283. [Google Scholar] [CrossRef]
- Pletser, V.; Rouquette, S.; Friedrich, U.; Clervoy, J.-F.; Gharib, T.; Gai, F.; Mora, C. The First European Parabolic Flight Campaign with the Airbus A310 ZERO-G. Microgravity Sci. Technol. 2016, 28, 587–601. [Google Scholar] [CrossRef]
- Sabbatini, M. Parabolic flights. In ESA User Guide to Low Gravity Platforms, 3rd ed.; Sentse, N., Oldenburg, K., Eds.; European Space Agency: Noordwijk, The Netherlands, 2014. [Google Scholar]
- Sabbatini, M. Sounding rockets. In ESA User Guide to Low Gravity Platforms, 3rd ed.; Sentse, N., Oldenburg, K., Eds.; European Space Agency: Noordwijk, The Netherlands, 2014. [Google Scholar]
- Horn, E.R.; Dournon, C.; Frippiat, J.-P.; Marco, R.; Böser, S.; Kirschnick, U. Development of neuronal and sensorimotor systems in the absence of gravity: Neurobiological research on four soyuz taxi flights to the international space station. Microgravity Sci. Technol. 2007, 19, 164–169. [Google Scholar] [CrossRef]
- Ou, X.; Long, L.; Wu, Y.; Yu, Y.; Lin, X.; Qi, X.; Liu, B. Spaceflight-induced genetic and epigenetic changes in the rice (Oryza sativa L.) genome are independent of each other. Genome 2010, 53, 524–532. [Google Scholar] [CrossRef]
- Preu, P.; Braun, M. German SIMBOX on Chinese mission Shenzhou-8: Europe’s first bilateral cooperation utilizing China’s Shenzhou programme. Acta Astronaut. 2014, 94, 584–591. [Google Scholar] [CrossRef]
- Sabbatini, M. International Space Station. In ESA User Guide to Low Gravity Platforms, 3rd ed.; Sentse, N., Oldenburg, K., Eds.; European Space Agency: Noordwijk, The Netherlands, 2014. [Google Scholar]
- Krüger, M.; Pietsch, J.; Bauer, J.; Kopp, S.; Carvalho, D.T.O.; Baatout, S.; Moreels, M.; Melnik, D.; Wehland, M.; Egli, M.; et al. Growth of Endothelial Cells in Space and in Simulated Microgravity—a Comparison on the Secretory Level. Cell. Physiol. Biochem. 2019, 52, 1039–1060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wuest, S.L.; Richard, S.; Kopp, S.; Grimm, D.; Egli, M. Simulated Microgravity: Critical Review on the Use of Random Positioning Machines for Mammalian Cell Culture. Biomed. Res. Int. 2015, 2015, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herranz, R.; Anken, R.; Boonstra, J.; Braun, M.; Christianen, P.C.M.; de Geest, M.; Hauslage, J.; Hilbig, R.; Hill, R.J.A.; Lebert, M.; et al. Ground-based facilities for simulation of microgravity: Organism-specific recommendations for their use, and recommended terminology. Astrobiology 2013, 13, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morey-Holton, E.; Globus, R.K.; Kaplansky, A.; Durnova, G. The Hindlimb Unloading Rat Model: Literature Overview, Technique Update and Comparison with Space Flight Data. In Experimentation with Animal Models in Space, 1st ed.; Sonnenfeld, G., Ed.; Elsevier: Amsterdam, The Netherlands, 2005; pp. 7–40. ISBN 9780444519078. [Google Scholar]
- Sabbatini, M. Ground Based Facilities. In ESA User Guide to Low Gravity Platforms, 3rd ed.; Sentse, N., Oldenburg, K., Eds.; European Space Agency: Noordwijk, The Netherlands, 2014. [Google Scholar]
- Schwarz, R.P.; Goodwin, T.J.; Wolf, D.A. Cell culture for three-dimensional modeling in rotating-wall vessels: An application of simulated microgravity. J. Tissue Cult. Methods 1992, 14, 51–57. [Google Scholar] [CrossRef]
- Globus, R.K.; Morey-Holton, E. Hindlimb unloading: Rodent analog for microgravity. J. Appl. Physiol. 2016, 120, 1196–1206. [Google Scholar] [CrossRef]
- Pavy-Le Traon, A.; Heer, M.; Narici, M.V.; Rittweger, J.; Vernikos, J. From space to Earth: Advances in human physiology from 20 years of bed rest studies (1986–2006). Eur. J. Appl. Physiol. 2007, 101, 143–194. [Google Scholar] [CrossRef]
- Little, G.H.; Flores, A. Inhibition of programmed cell death by catalase and phenylalanine methyl ester. Comp. Biochem. Physiol. A Physiol. 1993, 105, 79–83. [Google Scholar] [CrossRef]
- Horvitz, H.R. Genetic control of programmed cell death in the nematode Caenorhabditis elegans. Cancer Res. 1999, 59, 1701s–1706s. [Google Scholar]
- Norbury, C.J.; Hickson, I.D. Cellular responses to DNA damage. Annu. Rev. Pharm. Toxicol. 2001, 41, 367–401. [Google Scholar] [CrossRef] [Green Version]
- Cao, X.; Wen, P.; Fu, Y.; Gao, Y.; Qi, X.; Chen, B.; Tao, Y.; Wu, L.; Xu, A.; Lu, H.; et al. Radiation induces apoptosis primarily through the intrinsic pathway in mammalian cells. Cell Signal 2019, 62, 109337. [Google Scholar] [CrossRef] [PubMed]
- Galluzzi, L.; Vitale, I.; Aaronson, S.A.; Abrams, J.M.; Adam, D.; Agostinis, P.; Alnemri, E.S.; Altucci, L.; Amelio, I.; Andrews, D.W.; et al. Molecular mechanisms of cell death: Recommendations of the Nomenclature Committee on Cell Death 2018. Cell Death Differ. 2018, 25, 486–541. [Google Scholar] [CrossRef] [PubMed]
- Böhm, I.; Schild, H. Apoptosis: The Complex Scenario for a Silent Cell Death. Mol. Imaging Biol. 2003, 5, 2–14. [Google Scholar] [CrossRef]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Saelens, X.; Festjens, N.; Vande Walle, L.; van Gurp, M.; van Loo, G.; Vandenabeele, P. Toxic proteins released from mitochondria in cell death. Oncogene 2004, 23, 2861–2874. [Google Scholar] [CrossRef] [Green Version]
- Nagata, S. Apoptosis by Death Factor. Cell 1997, 88, 355–365. [Google Scholar] [CrossRef] [Green Version]
- Strasser, A.; Jost, P.J.; Nagata, S. The many roles of FAS receptor signaling in the immune system. Immunity 2009, 30, 180–192. [Google Scholar] [CrossRef] [Green Version]
- Maher, S.; Toomey, D.; Condron, C.; Bouchier-Hayes, D. Activation-induced cell death: The controversial role of Fas and Fas ligand in immune privilege and tumour counterattack. Immunol. Cell Biol. 2002, 80, 131–137. [Google Scholar] [CrossRef]
- Green, D.R.; Ferguson, T.A. The role of Fas ligand in immune privilege. Nat. Rev. Mol. Cell Biol. 2001, 2, 917–924. [Google Scholar] [CrossRef]
- Igney, F.H.; Krammer, P.H. Tumor counterattack: Fact or fiction? Cancer Immunol. Immunother. 2005, 54, 1127–1136. [Google Scholar] [CrossRef]
- Cabahug-Zuckerman, P.; Frikha-Benayed, D.; Majeska, R.J.; Tuthill, A.; Yakar, S.; Judex, S.; Schaffler, M.B. Osteocyte Apoptosis Caused by Hindlimb Unloading is Required to Trigger Osteocyte RANKL Production and Subsequent Resorption of Cortical and Trabecular Bone in Mice Femurs. J. Bone Min. Res. 2016, 31, 1356–1365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshihara, T.; Yamamoto, Y.; Shibaguchi, T.; Miyaji, N.; Kakigi, R.; Naito, H.; Goto, K.; Ohmori, D.; Yoshioka, T.; Sugiura, T. Dietary astaxanthin supplementation attenuates disuse-induced muscle atrophy and myonuclear apoptosis in the rat soleus muscle. J. Physiol. Sci. 2017, 67, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Hu, N.-F.; Chang, H.; Du, B.; Zhang, Q.-W.; Arfat, Y.; Dang, K.; Gao, Y.-F. Tetramethylpyrazine ameliorated disuse-induced gastrocnemius muscle atrophy in hindlimb unloading rats through suppression of Ca2+/ROS-mediated apoptosis. Appl. Physiol. Nutr. Metab. 2017, 42, 117–127. [Google Scholar] [CrossRef] [PubMed]
- Farley, A.; Gnyubkin, V.; Vanden-Bossche, A.; Laroche, N.; Neefs, M.; Baatout, S.; Baselet, B.; Vico, L.; Mastrandrea, C. Unloading-Induced Cortical Bone Loss is Exacerbated by Low-Dose Irradiation During a Simulated Deep Space Exploration Mission. Calcif. Tissue Int. 2020, 107, 170–179. [Google Scholar] [CrossRef]
- Aguirre, J.I.; Plotkin, L.I.; Stewart, S.A.; Weinstein, R.S.; Parfitt, A.M.; Manolagas, S.C.; Bellido, T. Osteocyte apoptosis is induced by weightlessness in mice and precedes osteoclast recruitment and bone loss. J. Bone Min. Res. 2006, 21, 605–615. [Google Scholar] [CrossRef]
- Plotkin, L.I.; Gortazar, A.R.; Davis, H.M.; Condon, K.W.; Gabilondo, H.; Maycas, M.; Allen, M.R.; Bellido, T. Inhibition of osteocyte apoptosis prevents the increase in osteocytic receptor activator of nuclear factor κB ligand (RANKL) but does not stop bone resorption or the loss of bone induced by unloading. J. Biol. Chem. 2015, 290, 18934–18942. [Google Scholar] [CrossRef] [Green Version]
- Horie, K.; Kudo, T.; Yoshinaga, R.; Akiyama, N.; Sasanuma, H.; Kobayashi, T.J.; Shimbo, M.; Jeon, H.; Miyao, T.; Miyauchi, M.; et al. Long-term hindlimb unloading causes a preferential reduction of medullary thymic epithelial cells expressing autoimmune regulator (Aire). Biochem. Biophys. Res. Commun. 2018, 501, 745–750. [Google Scholar] [CrossRef]
- Gaignier, F.; Schenten, V.; de Carvalho Bittencourt, M.; Gauquelin-Koch, G.; Frippiat, J.-P.; Legrand-Frossi, C. Three weeks of murine hindlimb unloading induces shifts from B to T and from th to tc splenic lymphocytes in absence of stress and differentially reduces cell-specific mitogenic responses. PLoS ONE 2014, 9, e92664. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.X.; Shi, Y.; Denhardt, D.T. Osteopontin regulates hindlimb-unloading-induced lymphoid organ atrophy and weight loss by modulating corticosteroid production. Proc. Natl. Acad. Sci. USA 2007, 104, 14777–14782. [Google Scholar] [CrossRef] [Green Version]
- Wise, K.C.; Manna, S.K.; Yamauchi, K.; Ramesh, V.; Wilson, B.L.; Thomas, R.L.; Sarkar, S.; Kulkarni, A.D.; Pellis, N.R.; Ramesh, G.T. Activation of nuclear transcription factor-kappaB in mouse brain induced by a simulated microgravity environment. Vitr. Cell. Dev. Biol. Anim. 2005, 41, 118–123. [Google Scholar] [CrossRef]
- Kulikova, E.A.; Kulikov, V.A.; Sinyakova, N.A.; Kulikov, A.V.; Popova, N.K. The effect of long-term hindlimb unloading on the expression of risk neurogenes encoding elements of serotonin-, dopaminergic systems and apoptosis; comparison with the effect of actual spaceflight on mouse brain. Neurosci. Lett. 2017, 640, 88–92. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, T.; Hayashi, M.; Fukunaga, T.; Kurata, K.; Oh-Hora, M.; Feng, J.Q.; Bonewald, L.F.; Kodama, T.; Wutz, A.; Wagner, E.F.; et al. Evidence for osteocyte regulation of bone homeostasis through RANKL expression. Nat. Med. 2011, 17, 1231–1234. [Google Scholar] [CrossRef] [PubMed]
- Allen, D.L.; Roy, R.R.; Edgerton, V.R. Myonuclear domains in muscle adaptation and disease. Muscle Nerve 1999, 22, 1350–1360. [Google Scholar] [CrossRef]
- Stein, T.P. Weight, muscle and bone loss during space flight: Another perspective. Eur. J. Appl. Physiol. 2013, 113, 2171–2181. [Google Scholar] [CrossRef]
- Roberts, D.R.; Albrecht, M.H.; Collins, H.R.; Asemani, D.; Chatterjee, A.R.; Spampinato, M.V.; Zhu, X.; Chimowitz, M.I.; Antonucci, M.U. Effects of Spaceflight on Astronaut Brain Structure as Indicated on MRI. New Engl. J. Med. 2017, 377, 1746–1753. [Google Scholar] [CrossRef]
- Vernice, N.A.; Meydan, C.; Afshinnekoo, E.; Mason, C.E. Long-term spaceflight and the cardiovascular system. Precis. Clin. Med. 2020, pbaa022. [Google Scholar] [CrossRef]
- Cancedda, R.; Liu, Y.; Ruggiu, A.; Tavella, S.; Biticchi, R.; Santucci, D.; Schwartz, S.; Ciparelli, P.; Falcetti, G.; Tenconi, C.; et al. The Mice Drawer System (MDS) experiment and the space endurance record-breaking mice. PLoS ONE 2012, 7, e32243. [Google Scholar] [CrossRef] [Green Version]
- Moyer, E.L.; Dumars, P.M.; Sun, G.-S.; Martin, K.J.; Heathcote, D.G.; Boyle, R.D.; Skidmore, M.G. Evaluation of rodent spaceflight in the NASA animal enclosure module for an extended operational period (up to 35 days). Npj Microgravity 2016, 2, 16002. [Google Scholar] [CrossRef]
- Mader, T.H.; Gibson, C.R.; Pass, A.F.; Kramer, L.A.; Lee, A.G.; Fogarty, J.; Tarver, W.J.; Dervay, J.P.; Hamilton, D.R.; Sargsyan, A.; et al. Optic disc edema, globe flattening, choroidal folds, and hyperopic shifts observed in astronauts after long-duration space flight. Ophthalmology 2011, 118, 2058–2069. [Google Scholar] [CrossRef] [Green Version]
- Philpott, D.E.; Corbett, R.; Turnbill, C.; Harrison, G.; Leaffer, D.; Black, S.; Sapp, W.; Klein, G.; Savik, L.F. Cosmic ray effects on the eyes of rats flown on Cosmos No. 782, experimental K-007. Aviat. Space Env. Med. 1978, 49, 19–28. [Google Scholar]
- Mao, X.W.; Pecaut, M.J.; Stodieck, L.S.; Ferguson, V.L.; Bateman, T.A.; Bouxsein, M.; Jones, T.A.; Moldovan, M.; Cunningham, C.E.; Chieu, J.; et al. Spaceflight Environment Induces Mitochondrial Oxidative Damage in Ocular Tissue. Radiat. Res. 2013, 180, 340–350. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.W.; Nishiyama, N.C.; Pecaut, M.J.; Campbell-Beachler, M.; Gifford, P.; Haynes, K.E.; Becronis, C.; Gridley, D.S. Simulated Microgravity and Low-Dose/Low-Dose-Rate Radiation Induces Oxidative Damage in the Mouse Brain. Radiat. Res. 2016, 185, 647–657. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.W.; Pecaut, M.J.; Stodieck, L.S.; Ferguson, V.L.; Bateman, T.A.; Bouxsein, M.L.; Gridley, D.S. Biological and metabolic response in STS-135 space-flown mouse skin. Free Radic. Res. 2014, 48, 890–897. [Google Scholar] [CrossRef] [PubMed]
- Horie, K.; Kato, T.; Kudo, T.; Sasanuma, H.; Miyauchi, M.; Akiyama, N.; Miyao, T.; Seki, T.; Ishikawa, T.; Takakura, Y.; et al. Impact of spaceflight on the murine thymus and mitigation by exposure to artificial gravity during spaceflight. Sci. Rep. 2019, 9, 19866. [Google Scholar] [CrossRef] [PubMed]
- Gridley, D.S.; Mao, X.W.; Tian, J.; Cao, J.D.; Perez, C.; Stodieck, L.S.; Ferguson, V.L.; Bateman, T.A.; Pecaut, M.J. Genetic and Apoptotic Changes in Lungs of Mice Flown on the STS-135 Mission in Space. Vivo 2015, 29, 423–433. [Google Scholar]
- Hammond, T.G.; Allen, P.L.; Birdsall, H.H. Effects of Space Flight on Mouse Liver versus Kidney: Gene Pathway Analyses. Int. J. Med. Sci. 2018, 19, 4106. [Google Scholar] [CrossRef] [Green Version]
- Lewis, M.L. The cytoskeleton, apoptosis, and gene expression in T lymphocytes and other mammalian cells exposed to altered gravity. Adv. Space Biol. Med. 2002, 8, 77–128. [Google Scholar]
- Cubano, L.A.; Lewis, M.L. Fas/APO-1 protein is increased in spaceflown lymphocytes (Jurkat)☆. Exp. Gerontol. 2000, 35, 389–400. [Google Scholar] [CrossRef]
- Sokolovskaya, A.; Korneeva, E.; Zaichenko, D.; Virus, E.; Kolesov, D.; Moskovtsev, A.; Kubatiev, A. Changes in the Surface Expression of Intercellular Adhesion Molecule 3, the Induction of Apoptosis, and the Inhibition of Cell-Cycle Progression of Human Multidrug-Resistant Jurkat/A4 Cells Exposed to a Random Positioning Machine. Int. J. Med. Sci. 2020, 21, 855. [Google Scholar] [CrossRef] [Green Version]
- Bakos, A.; Varkonyi, A.; Minarovits, J.; Batkai, L. Effect of simulated microgravity on human lymphocytes. J. Gravit. Physiol. 2001, 8, P69–P70. [Google Scholar]
- Maccarrone, M.; Battista, N.; Meloni, M.; Bari, M.; Galleri, G.; Pippia, P.; Cogoli, A.; Finazzi-Agrò, A. Creating conditions similar to those that occur during exposure of cells to microgravity induces apoptosis in human lymphocytes by 5-lipoxygenase-mediated mitochondrial uncoupling and cytochrome c release. J. Leukoc. Biol. 2003, 73, 472–481. [Google Scholar] [CrossRef] [PubMed]
- Battista, N.; Meloni, M.A.; Bari, M.; Mastrangelo, N.; Galleri, G.; Rapino, C.; Dainese, E.; Agrò, A.F.; Pippia, P.; Maccarrone, M. 5-Lipoxygenase-dependent apoptosis of human lymphocytes in the International Space Station: Data from the ROALD experiment. Faseb J. 2012, 26, 1791–1798. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Wang, C.; Feng, M.; Zhao, Y. Microgravity inhibits resting T cell immunity in an exposure time-dependent manner. Int. J. Med. Sci. 2014, 11, 87–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Risin, D.; Pellis, N.R. Modeled microgravity inhibits apoptosis in peripheral blood lymphocytes1. Febs Lett. 2001, 37, 66. [Google Scholar] [CrossRef] [Green Version]
- Kumari, R.; Singh, K.P.; Dumond, J.W. Simulated microgravity decreases DNA repair capacity and induces DNA damage in human lymphocytes. J. Cell Biochem. 2009, 107, 723–731. [Google Scholar] [CrossRef]
- Girardi, C.; de Pittà, C.; Casara, S.; Calura, E.; Romualdi, C.; Celotti, L.; Mognato, M. Integration analysis of microRNA and mRNA expression profiles in human peripheral blood lymphocytes cultured in modeled microgravity. Biomed. Res. Int. 2014, 2014, 296747. [Google Scholar] [CrossRef] [Green Version]
- Smyth, M.J.; Cretney, E.; Kelly, J.M.; Westwood, J.A.; Street, S.E.A.; Yagita, H.; Takeda, K.; van Dommelen, S.L.H.; Degli-Esposti, M.A.; Hayakawa, Y. Activation of NK cell cytotoxicity. Mol. Immunol. 2005, 42, 501–510. [Google Scholar] [CrossRef]
- Li, Q.; Mei, Q.; Huyan, T.; Xie, L.; Che, S.; Yang, H.; Zhang, M.; Huang, Q. Effects of simulated microgravity on primary human NK cells. Astrobiology 2013, 13, 703–714. [Google Scholar] [CrossRef]
- Congdon, C.C.; Allebban, Z.; Gibson, L.A.; Kaplansky, A.; Strickland, K.M.; Jago, T.L.; Johnson, D.L.; Lange, R.D.; Ichiki, A.T. Lymphatic tissue changes in rats flown on Spacelab Life Sciences-2. J. Appl. Physiol. 1996, 81, 172–177. [Google Scholar] [CrossRef]
- Novoselova, E.G.; Lunin, S.M.; Khrenov, M.O.; Parfenyuk, S.B.; Novoselova, T.V.; Shenkman, B.S.; Fesenko, E.E. Changes in immune cell signalling, apoptosis and stress response functions in mice returned from the BION-M1 mission in space. Immunobiology 2015, 220, 500–509. [Google Scholar] [CrossRef]
- Wei, L.X.; Zhou, J.N.; Roberts, A.I.; Shi, Y.F. Lymphocyte reduction induced by hindlimb unloading: Distinct mechanisms in the spleen and thymus. Cell Res. 2003, 13, 465–471. [Google Scholar] [CrossRef] [PubMed]
- Hur, E.M.; Youssef, S.; Haws, M.E.; Zhang, S.Y.; Sobel, R.A.; Steinman, L. Osteopontin-induced relapse and progression of autoimmune brain disease through enhanced survival of activated T cells. Nat. Immunol. 2007, 8, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Delp, M.D.; Charvat, J.M.; Limoli, C.L.; Globus, R.K.; Ghosh, P. Apollo Lunar Astronauts Show Higher Cardiovascular Disease Mortality: Possible Deep Space Radiation Effects on the Vascular Endothelium. Sci. Rep. 2016, 6, 29901. [Google Scholar] [CrossRef] [Green Version]
- Carlsson, S.I.M.; Bertilaccio, M.T.S.; Ballabio, E.; Maier, J.A.M. Endothelial stress by gravitational unloading: Effects on cell growth and cytoskeletal organization. Biochim. Biophys. Acta 2003, 1642, 173–179. [Google Scholar] [CrossRef] [Green Version]
- Versari, S.; Villa, A.; Bradamante, S.; Maier, J.A.M. Alterations of the actin cytoskeleton and increased nitric oxide synthesis are common features in human primary endothelial cell response to changes in gravity. Biochim. Biophys. Acta 2007, 1773, 1645–1652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanford, G.L.; Ellerson, D.; Melhado-Gardner, C.; Sroufe, A.E.; Harris-Hooker, S. Three-dimensional growth of endothelial cells in the microgravity-based rotating wall vessel bioreactor. Vitr. Cell Dev. Biol. Anim. 2002, 38, 493–504. [Google Scholar] [CrossRef]
- Cotrupi, S.; Ranzani, D.; Maier, J.A.M. Impact of modeled microgravity on microvascular endothelial cells. Biochim. Biophys. Acta 2005, 1746, 163–168. [Google Scholar] [CrossRef] [Green Version]
- Mariotti, M.; Maier, J.A.M. Gravitational unloading induces an anti-angiogenic phenotype in human microvascular endothelial cells. J. Cell Biochem. 2008, 104, 129–135. [Google Scholar] [CrossRef]
- Morbidelli, L.; Monici, M.; Marziliano, N.; Cogoli, A.; Fusi, F.; Waltenberger, J.; Ziche, M. Simulated hypogravity impairs the angiogenic response of endothelium by up-regulating apoptotic signals. Biochem. Biophys. Res. Commun. 2005, 334, 491–499. [Google Scholar] [CrossRef]
- Pan, Y.-K.; Li, C.-F.; Gao, Y.; Wang, Y.-C.; Sun, X.-Q. Effect of miR-27b-5p on apoptosis of human vascular endothelial cells induced by simulated microgravity. Apoptosis 2020, 25, 73–91. [Google Scholar] [CrossRef] [Green Version]
- Locatelli, L.; Cazzaniga, A.; de Palma, C.; Castiglioni, S.; Maier, J.A.M. Mitophagy contributes to endothelial adaptation to simulated microgravity. Faseb J. 2020, 34, 1833–1845. [Google Scholar] [CrossRef] [Green Version]
- Li, C.-F.; Pan, Y.-K.; Gao, Y.; Shi, F.; Wang, Y.-C.; Sun, X.-Q. Autophagy protects HUVECs against ER stress-mediated apoptosis under simulated microgravity. Apoptosis 2019, 24, 812–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, D.; Guo, Y.-B.; Zhang, M.; SUN, Y. The subsequent biological effects of simulated microgravity on endothelial cell growth in HUVECs. Chin. J. Traumatol. 2018, 21, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Kang, C.-Y.; Zou, L.; Yuan, M.; Wang, Y.; Li, T.-Z.; Zhang, Y.; Wang, J.-F.; Li, Y.; Deng, X.-W.; Liu, C.-T. Impact of simulated microgravity on microvascular endothelial cell apoptosis. Eur. J. Appl. Physiol. 2011, 111, 2131–2138. [Google Scholar] [CrossRef] [PubMed]
- Tang, N.-P.; Hui, T.-T.; Ma, J.; Mei, Q.-B. Effects of miR-503-5p on apoptosis of human pulmonary microvascular endothelial cells in simulated microgravity. J. Cell Biochem. 2019, 120, 727–737. [Google Scholar] [CrossRef] [Green Version]
- Cotrupi, S.; Maier, J.A.M. Is HSP70 upregulation crucial for cellular proliferative response in simulated microgravity? J. Gravit. Physiol. 2004, 11, P173–P176. [Google Scholar]
- Cazzaniga, A.; Locatelli, L.; Castiglioni, S.; Maier, J.A.M. The dynamic adaptation of primary human endothelial cells to simulated microgravity. Faseb J. 2019, 33, 5957–5966. [Google Scholar] [CrossRef]
- Dittrich, A.; Grimm, D.; Sahana, J.; Bauer, J.; Krüger, M.; Infanger, M.; Magnusson, N.E. Key Proteins Involved in Spheroid Formation and Angiogenesis in Endothelial Cells After Long-Term Exposure to Simulated Microgravity. Cell. Physiol. Biochem. 2018, 45, 429–445. [Google Scholar] [CrossRef]
- Pietsch, J.; Gass, S.; Nebuloni, S.; Echegoyen, D.; Riwaldt, S.; Baake, C.; Bauer, J.; Corydon, T.J.; Egli, M.; Infanger, M.; et al. Three-dimensional growth of human endothelial cells in an automated cell culture experiment container during the SpaceX CRS-8 ISS space mission—The SPHEROIDS project. Biomaterials 2017, 124, 126–156. [Google Scholar] [CrossRef]
- Tombran-Tink, J.; Barnstable, C.J. Space flight environment induces degeneration in the retina of rat neonates. Adv. Exp. Med. Biol. 2006, 572, 417–424. [Google Scholar] [CrossRef]
- Mader, C.T.H.; Gibson, C.R.; Caputo, M.; Hunter, N.; Taylor, G.; Charles, J.; Meehan, R.T. Intraocular Pressure and Retinal Vascular Changes During Transient Exposure to Microgravity. Am. J. Ophthalmol. 1993, 115, 347–350. [Google Scholar] [CrossRef]
- Uva, B.M.; Masini, M.A.; Sturla, M.; Tagliafierro, G.; Strollo, F. Microgravity-induced programmed cell death in astrocytes. J. Gravit. Physiol. 2002, 9, P275–P276. [Google Scholar]
- Mao, X.W.; Green, L.M.; Mekonnen, T.; Lindsey, N.; Gridley, D.S. Gene expression analysis of oxidative stress and apoptosis in proton-irradiated rat retina. Vivo 2010, 24, 425–430. [Google Scholar]
- Cingolani, C.; Rogers, B.; Lu, L.; Kachi, S.; Shen, J.; Campochiaro, P.A. Retinal degeneration from oxidative damage. Free Radic. Biol. Med. 2006, 40, 660–669. [Google Scholar] [CrossRef] [PubMed]
- Roberts, J.E.; Kukielczak, B.M.; Chignell, C.F.; Sik, B.H.; Hu, D.-N.; Principato, M.A. Simulated microgravity induced damage in human retinal pigment epithelial cells. Mol. Vis. 2006, 12, 633–638. [Google Scholar]
- Corydon, T.J.; Mann, V.; Slumstrup, L.; Kopp, S.; Sahana, J.; Askou, A.L.; Magnusson, N.E.; Echegoyen, D.; Bek, T.; Sundaresan, A.; et al. Reduced Expression of Cytoskeletal and Extracellular Matrix Genes in Human Adult Retinal Pigment Epithelium Cells Exposed to Simulated Microgravity. Cell. Physiol. Biochem. 2016, 40, 1–17. [Google Scholar] [CrossRef]
- Mao, X.W.; Byrum, S.; Nishiyama, N.C.; Pecaut, M.J.; Sridharan, V.; Boerma, M.; Tackett, A.J.; Shiba, D.; Shirakawa, M.; Takahashi, S.; et al. Impact of Spaceflight and Artificial Gravity on the Mouse Retina: Biochemical and Proteomic Analysis. Int. J. Med. Sci. 2018, 19, 2546. [Google Scholar] [CrossRef] [Green Version]
- Lulli, M.; Cialdai, F.; Vignali, L.; Monici, M.; Luzzi, S.; Cicconi, A.; Cacchione, S.; Magi, A.; Balsamo, M.; Vukich, M.; et al. The Coenzyme Q10 as an antiapoptotic countermeasure for retinal lesions onboard the International Space Station. Front. Physiol. 2018, 9, 36. [Google Scholar] [CrossRef]
- Brancato, R.; Schiavone, N.; Siano, S.; Lapucci, A.; Papucci, L.; Donnini, M.; Formigli, L.; Orlandini, S.Z.; Carella, G.; Carones, F.; et al. Prevention of corneal keratocyte apoptosis after argon fluoride excimer laser irradiation with the free radical scavenger ubiquinone Q10. Eur. J. Ophthalmol. 2000, 10, 32–38. [Google Scholar] [CrossRef]
- Brancato, R.; Fiore, T.; Papucci, L.; Schiavone, N.; Formigli, L.; Orlandini, S.Z.; Gobbi, P.G.; Carones, F.; Donnini, M.; Lapucci, A.; et al. Concomitant effect of topical ubiquinone Q10 and vitamin E to prevent keratocyte apoptosis after excimer laser photoablation in rabbits. J. Refract. Surg. 2002, 18, 135–139. [Google Scholar]
- Krasnov, I.B. Chapter 4 Gravitational Neuromorphology; Elsevier: Amsterdam, The Netherlands, 1994; pp. 85–110. ISBN 9781559384117. [Google Scholar]
- Bondar, R.L. The Neurolab Spacelab Mission: Neuroscience Research in Space: Results From the STS-90, Neurolab Spacelab Mission. Arch. Neurol. 2005, 62, 1314. [Google Scholar] [CrossRef]
- Mariggiò, M.; Illic, G.F. The effects of simulated microgravity on the human nervous system: The proposal of a three-dimensional glia–neuron co-culture cell model. Sci. Proc. 2015, 2, e892. [Google Scholar] [CrossRef] [Green Version]
- Uva, B.M.; Masini, M.A.; Sturla, M.; Bruzzone, F.; Giuliani, M.; Tagliafierro, G.; Strollo, F. Microgravity-induced apoptosis in cultured glial cells. Eur. J. Histochem. 2002, 46, 209–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pani, G.; Verslegers, M.; Quintens, R.; Samari, N.; de Saint-Georges, L.; van Oostveldt, P.; Baatout, S.; Benotmane, M.A. Combined Exposure to Simulated Microgravity and Acute or Chronic Radiation Reduces Neuronal Network Integrity and Survival. PLoS ONE 2016, 11, e0155260. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Ma, H.; Wu, L.; Cao, L.; Yang, Q.; Dong, H.; Wang, Z.; Ma, J.; Li, Z. The influence of simulated microgravity on proliferation and apoptosis in U251 glioma cells. Vitr. Cell. Dev. Biol. Anim. 2017, 53, 744–751. [Google Scholar] [CrossRef] [PubMed]
- Andreev-Andrievskiy, A.; Popova, A.; Boyle, R.; Alberts, J.; Shenkman, B.; Vinogradova, O.; Dolgov, O.; Anokhin, K.; Tsvirkun, D.; Soldatov, P.; et al. Mice in Bion-M 1 space mission: Training and selection. PLoS ONE 2014, 9, e104830. [Google Scholar] [CrossRef] [PubMed]
- Naumenko, V.S.; Kulikov, A.V.; Kondaurova, E.M.; Tsybko, A.S.; Kulikova, E.A.; Krasnov, I.B.; Shenkman, B.S.; Sychev, V.N.; Bazhenova, E.Y.; Sinyakova, N.A.; et al. Effect of actual long-term spaceflight on BDNF, TrkB, p75, BAX and BCL-XL genes expression in mouse brain regions. Neuroscience 2015, 284, 730–736. [Google Scholar] [CrossRef]
- Grimm, D.; Grosse, J.; Wehland, M.; Mann, V.; Reseland, J.E.; Sundaresan, A.; Corydon, T.J. The impact of microgravity on bone in humans. Bone 2016, 87, 44–56. [Google Scholar] [CrossRef]
- Blaber, E.; Marçal, H.; Burns, B.P. Bioastronautics: The influence of microgravity on astronaut health. Astrobiology 2010, 10, 463–473. [Google Scholar] [CrossRef]
- Bucaro, M.A.; Zahm, A.M.; Risbud, M.V.; Ayyaswamy, P.S.; Mukundakrishnan, K.; Steinbeck, M.J.; Shapiro, I.M.; Adams, C.S. The effect of simulated microgravity on osteoblasts is independent of the induction of apoptosis. J. Cell. Biochem. 2007, 102, 483–495. [Google Scholar] [CrossRef]
- Blaber, E.A.; Dvorochkin, N.; Lee, C.; Alwood, J.S.; Yousuf, R.; Pianetta, P.; Globus, R.K.; Burns, B.P.; Almeida, E.A.C. Microgravity induces pelvic bone loss through osteoclastic activity, osteocytic osteolysis, and osteoblastic cell cycle inhibition by CDKN1a/p21. PLoS ONE 2013, 8, e61372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bucaro, M.A.; Fertala, J.; Adams, C.S.; Steinbeck, M.; Ayyaswamy, P.; Mukundakrishnan, K.; Shapiro, I.M.; Risbud, M.V. Bone cell survival in microgravity: Evidence that modeled microgravity increases osteoblast sensitivity to apoptogens. Ann. N. Y. Acad. Sci. 2004, 1027, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Chatani, M.; Morimoto, H.; Takeyama, K.; Mantoku, A.; Tanigawa, N.; Kubota, K.; Suzuki, H.; Uchida, S.; Tanigaki, F.; Shirakawa, M.; et al. Acute transcriptional up-regulation specific to osteoblasts/osteoclasts in medaka fish immediately after exposure to microgravity. Sci. Rep. 2016, 6, 39545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumei, Y.; Shimokawa, H.; Ohya, K.; Katano, H.; Akiyama, H.; Hirano, M.; Morita, S. Small GTPase Ras and Rho expression in rat osteoblasts during spaceflight. Ann. N. Y. Acad. Sci. 2007, 1095, 292–299. [Google Scholar] [CrossRef]
- Kumei, Y.; Morita, S.; Katano, H.; Akiyama, H.; Hirano, M.; Oyha, K.; Shimokawa, H. Microgravity signal ensnarls cell adhesion, cytoskeleton, and matrix proteins of rat osteoblasts: Osteopontin, CD44, osteonectin, and alpha-tubulin. Ann. N. Y. Acad. Sci. 2006, 1090, 311–317. [Google Scholar] [CrossRef]
- Kumei, Y.; Morita, S.; Shimokawa, H.; Ohya, K.; Akiyama, H.; Hirano, M.; Sams, C.F.; Whitson, P.A. Inhibition of HSP70 and a collagen-specific molecular chaperone (HSP47) expression in rat osteoblasts by microgravity. Ann. N. Y. Acad. Sci. 2003, 1010, 476–480. [Google Scholar] [CrossRef]
- Kumei, Y.; Morita, S.; Nakamura, H.; Akiyama, H.; Hirano, M.; Shimokawa, H.; Ohya, K. Coinduction of GTP cyclohydrolase I and inducible NO synthase in rat osteoblasts during space flight: Apoptotic and self-protective response? Ann. N. Y. Acad. Sci. 2003, 1010, 481–485. [Google Scholar] [CrossRef]
- Nakamura, H.; Kumei, Y.; Morita, S.; Shimokawa, H.; Ohya, K.; Shinomiya, K. Antagonism between apoptotic (Bax/Bcl-2) and anti-apoptotic (IAP) signals in human osteoblastic cells under vector-averaged gravity condition. Ann. N. Y. Acad. Sci. 2003, 1010, 143–147. [Google Scholar] [CrossRef]
- Rucci, N.; Rufo, A.; Alamanou, M.; Teti, A. Modeled microgravity stimulates osteoclastogenesis and bone resorption by increasing osteoblast RANKL/OPG ratio. J. Cell. Biochem. 2007, 100, 464–473. [Google Scholar] [CrossRef]
- Sato, A.; Hamazaki, T.; Oomura, T.; Osada, H.; Kakeya, M.; Watanabe, M.; Nakamura, T.; Nakamura, Y.; Koshikawa, N.; Yoshizaki, I.; et al. Effects of microgravity on c-fos gene expression in osteoblast-like MC3T3-E1 cells. Adv. Space Res. 1999, 24, 807–813. [Google Scholar] [CrossRef]
- Chatziravdeli, V.; Katsaras, G.N.; Lambrou, G.I. Gene Expression in Osteoblasts and Osteoclasts Under Microgravity Conditions: A Systematic Review. Curr. Genom. 2019, 20, 184–198. [Google Scholar] [CrossRef] [PubMed]
- Freed, L.E.; Langer, R.; Martin, I.; Pellis, N.R.; Vunjak-Novakovic, G. Tissue engineering of cartilage in space. Proc. Natl. Acad. Sci. USA 1997, 94, 13885–13890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klement, B.; Spooner, B. Mineralization and growth of cultured embryonic skeletal tissue in microgravity. Bone 1999, 24, 349–359. [Google Scholar] [CrossRef]
- Duke, P.J.; Daane, E.L.; Montufar-Solis, D. Studies of chondrogenesis in rotating systems. J. Cell. Biochem. 1993, 51, 274–282. [Google Scholar] [CrossRef] [PubMed]
- Montufar-Solis, D.; Duke, P.J. Gravitational changes affect tibial growth plates according to Hert’s curve. Aviat. Space Env. Med. 1999, 70, 245–249. [Google Scholar]
- Wehland, M.; Aleshcheva, G.; Schulz, H.; Saar, K.; Hübner, N.; Hemmersbach, R.; Braun, M.; Ma, X.; Frett, T.; Warnke, E.; et al. Differential gene expression of human chondrocytes cultured under short-term altered gravity conditions during parabolic flight maneuvers. Cell Commun. Signal 2015, 13, 18. [Google Scholar] [CrossRef] [Green Version]
- Ulbrich, C.; Westphal, K.; Pietsch, J.; Winkler, H.D.F.; Leder, A.; Bauer, J.; Kossmehl, P.; Grosse, J.; Schoenberger, J.; Infanger, M.; et al. Characterization of human chondrocytes exposed to simulated microgravity. Cell. Physiol. Biochem. 2010, 25, 551–560. [Google Scholar] [CrossRef]
- Whiteman, M.; Armstrong, J.S.; Cheung, N.S.; Siau, J.-L.; Rose, P.; Schantz, J.-T.; Jones, D.P.; Halliwell, B. Peroxynitrite mediates calcium-dependent mitochondrial dysfunction and cell death via activation of calpains. Faseb J. 2004, 18, 1395–1397. [Google Scholar] [CrossRef]
- Pisanu, M.E.; Noto, A.; de Vitis, C.; Masiello, M.G.; Coluccia, P.; Proietti, S.; Giovagnoli, M.R.; Ricci, A.; Giarnieri, E.; Cucina, A.; et al. Lung cancer stem cell lose their stemness default state after exposure to microgravity. Biomed. Res. Int. 2014, 2014, 470253. [Google Scholar] [CrossRef]
- Zhu, M.; Jin, X.W.; Wu, B.Y.; Nie, J.L.; Li, Y.H. Effects of simulated weightlessness on cellular morphology and biological characteristics of cell lines SGC-7901 and HFE-145. Genet. Mol. Res. 2014, 13, 6060–6069. [Google Scholar] [CrossRef]
- Chen, Z.-Y.; Jiang, N.; Guo, S.; Li, B.-B.; Yang, J.-Q.; Chai, S.-B.; Yan, H.-F.; Sun, P.-M.; Zhang, T.; Sun, H.-W.; et al. Effect of simulated microgravity on metabolism of HGC-27 gastric cancer cells. Oncol. Lett. 2020, 19, 3439–3450. [Google Scholar] [CrossRef] [PubMed]
- Kossmehl, P.; Shakibaei, M.; Cogoli, A.; Infanger, M.; Curcio, F.; Schönberger, J.; Eilles, C.; Bauer, J.; Pickenhahn, H.; Schulze-Tanzil, G.; et al. Weightlessness induced apoptosis in normal thyroid cells and papillary thyroid carcinoma cells via extrinsic and intrinsic pathways. Endocrinology 2003, 144, 4172–4179. [Google Scholar] [CrossRef] [Green Version]
- Masiello, M.G.; Cucina, A.; Proietti, S.; Palombo, A.; Coluccia, P.; D’Anselmi, F.; Dinicola, S.; Pasqualato, A.; Morini, V.; Bizzarri, M. Phenotypic switch induced by simulated microgravity on MDA-MB-231 breast cancer cells. Biomed. Res. Int. 2014, 2014, 652434. [Google Scholar] [CrossRef] [PubMed]
- Du, F.; Ding, Y.; Zou, J.; Li, Z.; Tian, J.; She, R.; Wang, D.; Wang, H.; Lv, D.; Chang, L. Morphology and Molecular Mechanisms of Hepatic Injury in Rats under Simulated Weightlessness and the Protective Effects of Resistance Training. PLoS ONE 2015, 10, e0127047. [Google Scholar] [CrossRef]
- Blaber, E.A.; Pecaut, M.J.; Jonscher, K.R. Spaceflight Activates Autophagy Programs and the Proteasome in Mouse Liver. Int. J. Med. Sci. 2017, 18, 2062. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Duan, X.-Q.; Li, X.; Han, L.-O.; Xu, P.; Song, C.-F.; Jin, L.-H. Experimental study of rat beta islet cells cultured under simulated microgravity conditions. Acta Biochim. Biophys. Sin. (Shanghai) 2004, 36, 47–50. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Wei, Z.; Song, C.; Xie, S.; Feng, J.; Fan, J.; Zhang, Z.; Shi, Y. Simulated microgravity combined with polyglycolic acid scaffold culture conditions improves the function of pancreatic islets. Biomed. Res. Int. 2013, 2013, 150739. [Google Scholar] [CrossRef]
- Tobin, B.W.; Leeper-Woodford, S.K.; Hashemi, B.B.; Smith, S.M.; Sams, C.F. Altered TNF-alpha, glucose, insulin, and amino acids in islets of Langerhans cultured in a microgravity model system. Am. J. Physiol. Endocrinol. Metab. 2001, 280, E92–E102. [Google Scholar] [CrossRef] [Green Version]
- Rutzky, L.P.; Bilinski, S.; Kloc, M.; Phan, T.; Zhang, H.; Katz, S.M.; Stepkowski, S.M. Microgravity culture condition reduces immunogenicity and improves function of pancreatic islets1. Transplantation 2002, 74, 13–21. [Google Scholar] [CrossRef]
- Jessup, J.M.; Frantz, M.; Sonmez-Alpan, E.; Locker, J.; Skena, K.; Waller, H.; Battle, P.; Nachman, A.; Weber, M.E.; Thomas, D.A.; et al. Microgravity culture reduces apoptosis and increases the differentiation of a human colorectal carcinoma cell line. Nat. Med. 2000, 36, 367. [Google Scholar] [CrossRef]
- Jiang, N.; Chen, Z.; Li, B.; Guo, S.; Li, A.; Zhang, T.; Fu, X.; Si, S.; Cui, Y. Effects of rotary cell culture system-simulated microgravity on the ultrastructure and biological behavior of human MDA-MB-231 breast cancer cells. Prec. Radiat. Oncol. 2019, 3, 87–93. [Google Scholar] [CrossRef] [Green Version]
- Arun, R.P.; Sivanesan, D.; Vidyasekar, P.; Verma, R.S. PTEN/FOXO3/AKT pathway regulates cell death and mediates morphogenetic differentiation of Colorectal Cancer Cells under Simulated Microgravity. Sci. Rep. 2017, 7, 5952. [Google Scholar] [CrossRef] [PubMed]
- Vidyasekar, P.; Shyamsunder, P.; Arun, R.; Santhakumar, R.; Kapadia, N.K.; Kumar, R.; Verma, R.S. Genome Wide Expression Profiling of Cancer Cell Lines Cultured in Microgravity Reveals Significant Dysregulation of Cell Cycle and MicroRNA Gene Networks. PLoS ONE 2015, 10, e0135958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coinu, R.; Chiaviello, A.; Galleri, G.; Franconi, F.; Crescenzi, E.; Palumbo, G. Exposure to modeled microgravity induces metabolic idleness in malignant human MCF-7 and normal murine VSMC cells. Febs Lett. 2006, 580, 2465–2470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dietz, C.; Infanger, M.; Romswinkel, A.; Strube, F.; Kraus, A. Apoptosis Induction and Alteration of Cell Adherence in Human Lung Cancer Cells under Simulated Microgravity. Int. J. Med. Sci. 2019, 20, 3601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melnik, D.; Sahana, J.; Corydon, T.J.; Kopp, S.; Nassef, M.Z.; Wehland, M.; Infanger, M.; Grimm, D.; Krüger, M. Dexamethasone Inhibits Spheroid Formation of Thyroid Cancer Cells Exposed to Simulated Microgravity. Cells 2020, 9, 367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kopp, S.; Sahana, J.; Islam, T.; Petersen, A.G.; Bauer, J.; Corydon, T.J.; Schulz, H.; Saar, K.; Huebner, N.; Slumstrup, L.; et al. The role of NFκB in spheroid formation of human breast cancer cells cultured on the Random Positioning Machine. Sci. Rep. 2018, 8, 921. [Google Scholar] [CrossRef]
- Nassef, M.Z.; Kopp, S.; Melnik, D.; Corydon, T.J.; Sahana, J.; Krüger, M.; Wehland, M.; Bauer, T.J.; Liemersdorf, C.; Hemmersbach, R.; et al. Short-Term Microgravity Influences Cell Adhesion in Human Breast Cancer Cells. Int. J. Med. Sci. 2019, 20, 5730. [Google Scholar] [CrossRef] [Green Version]
- Fukazawa, T.; Tanimoto, K.; Shrestha, L.; Imura, T.; Takahashi, S.; Sueda, T.; Hirohashi, N.; Hiyama, E.; Yuge, L. Simulated microgravity enhances CDDP-induced apoptosis signal via p53-independent mechanisms in cancer cells. PLoS ONE 2019, 14, e0219363. [Google Scholar] [CrossRef] [Green Version]
- Deng, B.; Liu, R.; Tian, X.; Han, Z.; Chen, J. Simulated microgravity inhibits the viability and migration of glioma via FAK/RhoA/Rock and FAK/Nek2 signaling. Vitr. Cell. Dev. Biol. Anim. 2019, 55, 260–271. [Google Scholar] [CrossRef]
- Bonfiglio, T.; Biggi, F.; Bassi, A.M.; Ferrando, S.; Gallus, L.; Loiacono, F.; Ravera, S.; Rottigni, M.; Scarfì, S.; Strollo, F.; et al. Simulated microgravity induces nuclear translocation of Bax and BCL-2 in glial cultured C6 cells. Heliyon 2019, 5, e01798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, T.; Tang, X.; Umeshappa, C.S.; Ma, H.; Gao, H.; Deng, Y.; Freywald, A.; Xiang, J. Simulated Microgravity Promotes Cell Apoptosis Through Suppressing Uev1A/TICAM/TRAF/NF-κB-Regulated Anti-Apoptosis and p53/PCNA- and ATM/ATR-Chk1/2-Controlled DNA-Damage Response Pathways. J. Cell Biochem. 2016, 117, 2138–2148. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.-Y.; Guo, S.; Li, B.-B.; Jiang, N.; Li, A.; Yan, H.-F.; Yang, H.-M.; Zhou, J.-L.; Li, C.-L.; Cui, Y. Effect of Weightlessness on the 3D Structure Formation and Physiologic Function of Human Cancer Cells. Biomed. Res. Int. 2019, 2019, 4894083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ambrosini, G.; Adida, C.; Altieri, D.C. A novel anti-apoptosis gene, survivin, expressed in cancer and lymphoma. Nat. Med. 1997, 3, 917–921. [Google Scholar] [CrossRef]
- Ma, X.; Pietsch, J.; Wehland, M.; Schulz, H.; Saar, K.; Hübner, N.; Bauer, J.; Braun, M.; Schwarzwälder, A.; Segerer, J.; et al. Differential gene expression profile and altered cytokine secretion of thyroid cancer cells in space. Faseb J. 2014, 28, 813–835. [Google Scholar] [CrossRef]
- Shin, S.; Sung, B.J.; Cho, Y.S.; Kim, H.J.; Ha, N.C.; Hwang, J.I.; Chung, C.W.; Jung, Y.K.; Oh, B.H. An anti-apoptotic protein human survivin is a direct inhibitor of caspase-3 and -7. Biochemistry 2001, 40, 1117–1123. [Google Scholar] [CrossRef]
- Liu, T.; Brouha, B.; Grossman, D. Rapid induction of mitochondrial events and caspase-independent apoptosis in Survivin-targeted melanoma cells. Oncogene 2004, 23, 39–48. [Google Scholar] [CrossRef] [Green Version]
- Furukawa, S.; Nagamatsu, A.; Nenoi, M.; Fujimori, A.; Kakinuma, S.; Katsube, T.; Wang, B.; Tsuruoka, C.; Shirai, T.; Nakamura, A.J.; et al. Space Radiation Biology for “Living in Space”. Biomed Res. Int. 2020, 2020, 4703286. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, A.R. Biological Effects of Space Radiation and Development of Effective Countermeasures. Life Sci. Space Res. 2014, 1, 10–43. [Google Scholar] [CrossRef] [Green Version]
- Hellweg, C.E.; Gerzer, R.; Reitz, G. A new chapter in doctoral candidate training: The Helmholtz Space Life Sciences Research School (SpaceLife). Acta Astronaut. 2011, 68, 1620–1627. [Google Scholar] [CrossRef]
- Li, Z.; Jella, K.K.; Jaafar, L.; Li, S.; Park, S.; Story, M.D.; Wang, H.; Wang, Y.; Dynan, W.S. Exposure to galactic cosmic radiation compromises DNA repair and increases the potential for oncogenic chromosomal rearrangement in bronchial epithelial cells. Sci. Rep. 2018, 8, 11038. [Google Scholar] [CrossRef] [PubMed]
- Smart, D.F.; Shea, M.A. The local time dependence of the anisotropic solar cosmic ray flux. Adv. Space Res. 2003, 32, 109–114. [Google Scholar] [CrossRef]
- Abe, K.; Takahashi, H.; Suge, H. Graviresponding sites in shoots of normal and ‘lazy’ rice seedlings. Physiol. Plant 1994, 92, 371–374. [Google Scholar] [CrossRef]
- Takahashi, A.; Ikeda, H.; Yoshida, Y. Role of High-Linear Energy Transfer Radiobiology in Space Radiation Exposure Risks. Int. J. Part. Ther. 2018, 5, 151–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikjoo, H.; O’Neill, P.; Wilson, W.E.; Goodhead, D.T. Computational Approach for Determining the Spectrum of DNA Damage Induced by Ionizing Radiation. Radiat. Res. 2001, 156, 577–583. [Google Scholar] [CrossRef]
- Friedland, W.; Dingfelder, M.; Kundrát, P.; Jacob, P. Track structures, DNA targets and radiation effects in the biophysical Monte Carlo simulation code PARTRAC. Mutat. Res. 2011, 711, 28–40. [Google Scholar] [CrossRef]
- Cucinotta, F.A.; Manuel, F.K.; Jones, J.; Iszard, G.; Murrey, J.; Djojonegro, B.; Wear, M. Space Radiation and Cataracts in Astronauts. Radiat. Res. 2001, 156, 460–466. [Google Scholar] [CrossRef]
- Cucinotta, F.A.; Durante, M. Cancer risk from exposure to galactic cosmic rays: Implications for space exploration by human beings. Lancet Oncol. 2006, 7, 431–435. [Google Scholar] [CrossRef] [Green Version]
- Arena, C.; de Micco, V.; Macaeva, E.; Quintens, R. Space radiation effects on plant and mammalian cells. Acta Astronaut. 2014, 104, 419–431. [Google Scholar] [CrossRef]
- Danho, S.; Thorgrimson, J.; Saary, J. Effects of Space Radiation on Mammalian Cells. In Handbook of Space Pharmaceuticals; Pathak, Y., Araújo dos Santos, M., Zea, L., Eds.; Springer International Publishing: Cham, Switherlands, 2018; pp. 1–18. ISBN 978-3-319-50909-9. [Google Scholar]
- Horneck, G.; Klaus, D.M.; Mancinelli, R.L. Space Microbiology. Microbiol. Mol. Biol. Rev. 2010, 74, 121–156. [Google Scholar] [CrossRef] [Green Version]
- De Micco, V.; Arena, C.; Pignalosa, D.; Durante, M. Effects of sparsely and densely ionizing radiation on plants. Radiat. Env. Biophys. 2011, 50, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Gonzalo, R.; Baatout, S.; Moreels, M. Impact of Particle Irradiation on the Immune System: From the Clinic to Mars. Front. Immunol. 2017, 8, 177. [Google Scholar] [CrossRef] [PubMed]
- Barcellos-Hoff, M.H.; Blakely, E.A.; Burma, S.; Fornace, A.J.; Gerson, S.; Hlatky, L.; Kirsch, D.G.; Luderer, U.; Shay, J.; Wang, Y.; et al. Concepts and challenges in cancer risk prediction for the space radiation environment. Life Sci. Space Res. 2015, 6, 92–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cucinotta, F.A. A new approach to reduce uncertainties in space radiation cancer risk predictions. PLoS ONE 2015, 10, e0120717. [Google Scholar] [CrossRef] [PubMed]
- Parihar, V.K.; Allen, B.; Tran, K.K.; Macaraeg, T.G.; Chu, E.M.; Kwok, S.F.; Chmielewski, N.N.; Craver, B.M.; Baulch, J.E.; Acharya, M.M.; et al. What happens to your brain on the way to Mars. Sci. Adv. 2015, 1, e1400256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cherry, J.D.; Liu, B.; Frost, J.L.; Lemere, C.A.; Williams, J.P.; Olschowka, J.A.; O’Banion, M.K. Galactic cosmic radiation leads to cognitive impairment and increased aβ plaque accumulation in a mouse model of Alzheimer’s disease. PLoS ONE 2012, 7, e53275. [Google Scholar] [CrossRef]
- Cucinotta, F.A.; Alp, M.; Sulzman, F.M.; Wang, M. Space radiation risks to the central nervous system. Life Sci. Space Res. 2014, 2, 54–69. [Google Scholar] [CrossRef] [Green Version]
- Zhou, B.B.; Elledge, S.J. The DNA damage response: Putting checkpoints in perspective. Nature 2000, 408, 433–439. [Google Scholar] [CrossRef]
- Elledge, S.J. Cell cycle checkpoints: Preventing an identity crisis. Science 1996, 274, 1664–1672. [Google Scholar] [CrossRef]
- Hirao, A.; Cheung, A.; Duncan, G.; Girard, P.-M.; Elia, A.J.; Wakeham, A.; Okada, H.; Sarkissian, T.; Wong, J.A.; Sakai, T.; et al. Chk2 is a tumor suppressor that regulates apoptosis in both an ataxia telangiectasia mutated (ATM)-dependent and an ATM-independent manner. Mol. Cell. Biol. 2002, 22, 6521–6532. [Google Scholar] [CrossRef] [Green Version]
- Manda, K.; Ueno, M.; Anzai, K. Memory impairment, oxidative damage and apoptosis induced by space radiation: Ameliorative potential of alpha-lipoic acid. Behav. Brain Res. 2008, 187, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Manda, K.; Ueno, M.; Anzai, K. Space radiation-induced inhibition of neurogenesis in the hippocampal dentate gyrus and memory impairment in mice: Ameliorative potential of the melatonin metabolite, AFMK. J. Pineal Res. 2008, 45, 430–438. [Google Scholar] [CrossRef] [PubMed]
- Manda, K.; Ueno, M.; Anzai, K. Melatonin mitigates oxidative damage and apoptosis in mouse cerebellum induced by high-LET 56Fe particle irradiation. J. Pineal Res. 2008, 44, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Limoli, C.L.; Giedzinski, E.; Rola, R.; Otsuka, S.; Palmer, T.D.; Fike, J.R. Radiation response of neural precursor cells: Linking cellular sensitivity to cell cycle checkpoints, apoptosis and oxidative stress. Radiat. Res. 2004, 161, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Limoli, C.L.; Giedzinski, E.; Baure, J.; Rola, R.; Fike, J.R. Redox changes induced in hippocampal precursor cells by heavy ion irradiation. Radiat. Env. Biophys. 2007, 46, 167–172. [Google Scholar] [CrossRef]
- Yamada, T.; Ohyama, H. Radiation-induced interphase death of rat thymocytes is internally programmed (apoptosis). Int. J. Radiat. Biol. Relat. Stud. Phys. Chem. Med. 1988, 53, 65–75. [Google Scholar] [CrossRef]
- Mori, N.; Okumoto, M.; Morimoto, J.; Imai, S.; Matsuyama, T.; Takamori, Y.; Yagasaki, O. Genetic analysis of susceptibility to radiation-induced apoptosis of thymocytes in mice. Int. J. Radiat. Biol. 1992, 62, 153–159. [Google Scholar] [CrossRef]
- Mori, N.; Okumoto, M.; van der Valk, M.A.; Imai, S.; Haga, S.; Esaki, K.; Hart, A.A.; Demant, P. Genetic dissection of susceptibility to radiation-induced apoptosis of thymocytes and mapping of Rapop1, a novel susceptibility gene. Genomics 1995, 25, 609–614. [Google Scholar] [CrossRef]
- Potten, C.S. The significance of spontaneous and induced apoptosis in the gastrointestinal tract of mice. Cancer Metastasis Rev. 1992, 11, 179–195. [Google Scholar] [CrossRef]
- Potten, C.S.; Grant, H.K. The relationship between ionizing radiation-induced apoptosis and stem cells in the small and large intestine. Br. J. Cancer 1998, 78, 993–1003. [Google Scholar] [CrossRef] [Green Version]
- Mori, N.; van Wezel, T.; van der Valk, M.; Yamate, J.; Sakuma, S.; Okumoto, M.; Demant, P. Genetics of susceptibility to radiation-induced apoptosis in colon: Two loci on chromosomes 9 and 16. Mamm. Genome 1998, 9, 377–380. [Google Scholar] [CrossRef] [PubMed]
- Weil, M.M.; Amos, C.I.; Mason, K.A.; Stephens, L.C. Genetic Basis of Strain Variation in Levels of Radiation-Induced Apoptosis of Thymocytes. Radiat. Res. 1996, 146, 646. [Google Scholar] [CrossRef]
- Weil, M.M.; Stephens, L.C.; Amos, C.I.; Ruifrok, A.C.; Mason, K.A. Strain difference in jejunal crypt cell susceptibility to radiation-induced apoptosis. Int. J. Radiat. Biol. 1996, 70, 579–585. [Google Scholar] [CrossRef] [PubMed]
- Mori, N.; Okumoto, M.; Hart, A.A.; Demant, P. Apoptosis susceptibility genes on mouse chromosome 9 (Rapop2) and chromosome 3 (Rapop3). Genomics 1995, 30, 553–557. [Google Scholar] [CrossRef] [PubMed]
- Ristic-Fira, A.; Nikolic, D.; Petrovic, I.; Ruzdijic, S.; Raffaele, L.; Sabini, M.G.; Cirrone, G.A.; Cuttone, G.; Farruggia, G.; Masotti, L.; et al. The late effects of proton irradiation on cell growth, cell cycle arrest and apoptosis in a human melanoma cell line. J. Exp. Clin. Cancer Res. 2001, 20, 135–143. [Google Scholar]
- Sanzari, J.K.; Wambi, C.; Lewis-Wambi, J.S.; Kennedy, A.R. Antioxidant dietary supplementation in mice exposed to proton radiation attenuates expression of programmed cell death-associated genes. Radiat. Res. 2011, 175, 650–656. [Google Scholar] [CrossRef] [Green Version]
- Di Pietro, C.; Piro, S.; Tabbì, G.; Ragusa, M.; Di Pietro, V.; Zimmitti, V.; Cuda, F.; Anello, M.; Consoli, U.; Salinaro, E.T.; et al. Cellular and molecular effects of protons: Apoptosis induction and potential implications for cancer therapy. Apoptosis 2006, 11, 57–66. [Google Scholar] [CrossRef]
- Lee, K.B.; Lee, J.S.; Park, J.W.; Huh, T.L.; Lee, Y.M. Low energy proton beam induces tumor cell apoptosis through reactive oxygen species and activation of caspases. Exp. Mol. Med. 2008, 40, 118–129. [Google Scholar] [CrossRef]
- Finnberg, N.; Wambi, C.; Ware, J.H.; Kennedy, A.R.; El-Deiry, W.S. Gamma-radiation (GR) triggers a unique gene expression profile associated with cell death compared to proton radiation (PR) in mice in vivo. Cancer Biol. Ther. 2008, 7, 2023–2033. [Google Scholar] [CrossRef] [Green Version]
- Petrović, I.; Ristić-Fira, A.; Todorović, D.; Valastro, L.; Cirrone, P.; Cuttone, G. Radiobiological analysis of human melanoma cells on the 62 MeV CATANA proton beam. Int. J. Radiat. Biol. 2006, 82, 251–265. [Google Scholar] [CrossRef]
- Green, L.M.; Murray, D.K.; Tran, D.T.; Bant, A.M.; Kazarians, G.; Moyers, M.F.; Nelson, G.A. Response of Thyroid Follicular Cells to Gamma Irradiation Compared to Proton Irradiation. I. Initial Characterization of DNA Damage, Micronucleus Formation, Apoptosis, Cell Survival, and Cell Cycle Phase Redistribution. Radiat. Res. 2001, 155, 32–42. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prasad, B.; Grimm, D.; Strauch, S.M.; Erzinger, G.S.; Corydon, T.J.; Lebert, M.; Magnusson, N.E.; Infanger, M.; Richter, P.; Krüger, M. Influence of Microgravity on Apoptosis in Cells, Tissues, and Other Systems In Vivo and In Vitro. Int. J. Mol. Sci. 2020, 21, 9373. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21249373
Prasad B, Grimm D, Strauch SM, Erzinger GS, Corydon TJ, Lebert M, Magnusson NE, Infanger M, Richter P, Krüger M. Influence of Microgravity on Apoptosis in Cells, Tissues, and Other Systems In Vivo and In Vitro. International Journal of Molecular Sciences. 2020; 21(24):9373. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21249373
Chicago/Turabian StylePrasad, Binod, Daniela Grimm, Sebastian M. Strauch, Gilmar Sidnei Erzinger, Thomas J. Corydon, Michael Lebert, Nils E. Magnusson, Manfred Infanger, Peter Richter, and Marcus Krüger. 2020. "Influence of Microgravity on Apoptosis in Cells, Tissues, and Other Systems In Vivo and In Vitro" International Journal of Molecular Sciences 21, no. 24: 9373. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21249373