Possible Role of Crystal-Bearing Cells in Tomato Fertility and Formation of Seedless Fruits

 ,
, 
 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Characteristics of Flowers, Anthers and Pollen Structure of Transgenic and WT Plants
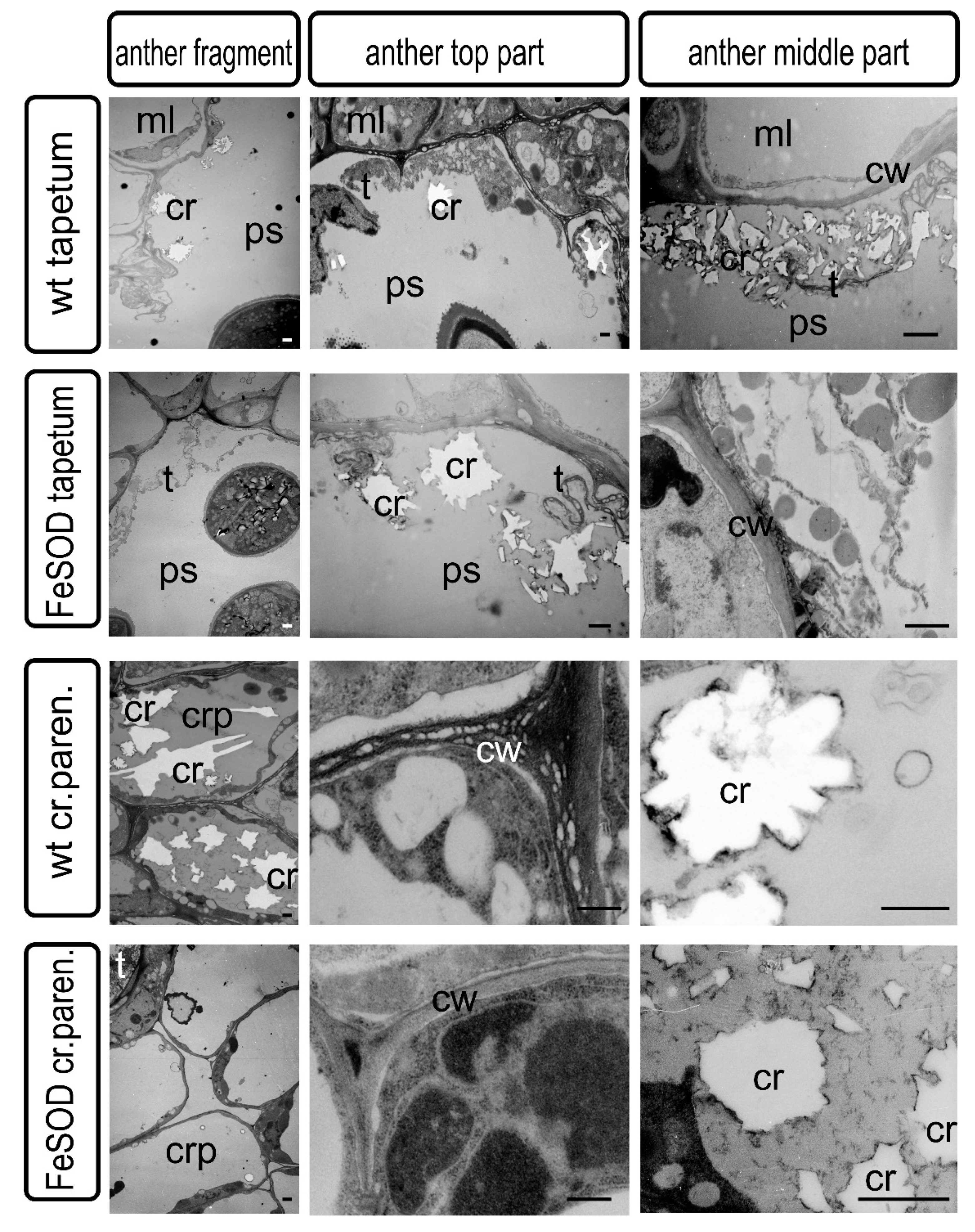
2.2. Features of the Ultrastructure of Pollen Sacs and Crystal-Bearing Parenchymatous Cells of the Connective Tissue
2.3. Morphological Features of Anthers of FeSOD-Transgenic and WT Tomato Plants
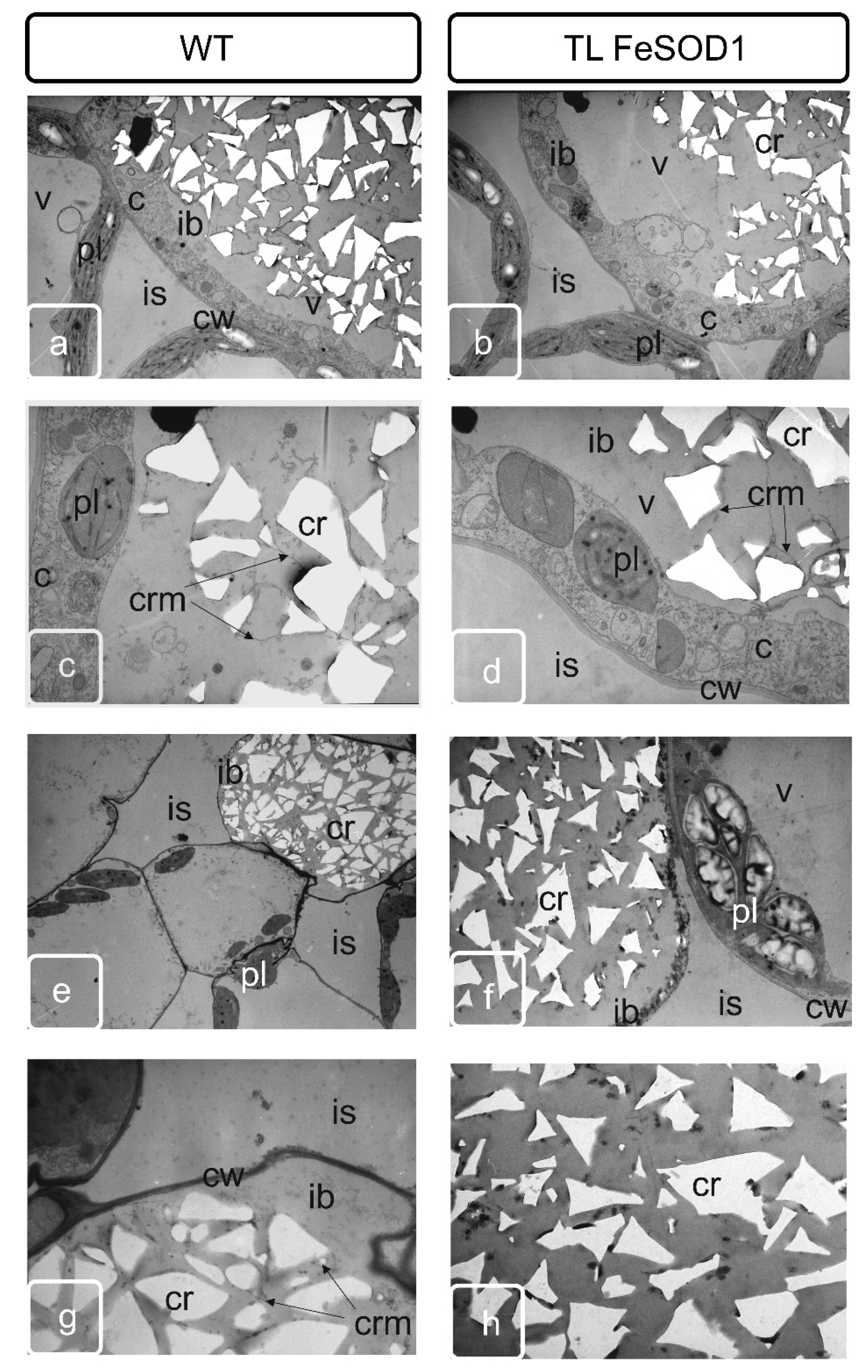
2.4. Comparative Study of Idioblasts of WT and FeSOD-Transgenic Plants
2.5. Transmission Microscopy of Cross Sections of Idioblasts Containing Spongy Parenchyma Tissue from Mature Leaf of Tomato Plants
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Sampling for Scanning and Transmission Electron Microscopy
4.3. Sample Preparation for Scanning Microscopy
4.4. Sample Preparation for Transmission Electron Microscopy
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- White, P.J.; Broadley, M.R. Calcium in plants. Ann. Bot. 2003, 92, 487–511. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, V.R.; Nakata, P.A. Calcium oxalate in plants: Formation and function. Annu. Rev. Plant Biol. 2005, 56, 41–71. [Google Scholar] [CrossRef] [PubMed]
- Prasad, R.; Shivay, Y.S. Oxalic acid/oxalates in plants: From self-defence to phytoremediation. Curr. Sci. 2017, 112, 110–112. [Google Scholar] [CrossRef]
- Libert, B.; Franceschi, V.P. Oxalate in crop plants. J. Agric. Food Chem. 1987, 35, 926–938. [Google Scholar] [CrossRef]
- Franceschi, V.R.; Horner, H.T. Calcium oxalate crystals in plants. Bot. Rev. 1980, 46, 361–427. [Google Scholar] [CrossRef]
- McLaughlin, S.B.; Wimmer, R. Tansley review no. 104: Calcium physiology and terrestrial ecosystem processes. New Phytol. 1999, 142, 373–417. [Google Scholar] [CrossRef]
- Hara, T.; Kobayashi, E.; Ohtsubo, K.; Kumada, S.; Kanazawa, M.; Abe, T.; Itoh, R.D.; Fujiwara, M.T. Organ-level analysis of idioblast patterning in Egeria densa Planch. leaves. PLoS ONE 2015, 10, e0118965. [Google Scholar] [CrossRef]
- Cuellar-Cruz, M.; Perez, K.S.; Mendoza, M.E.; Moreno, A. Biocrystals in plants: A short review on biomineralization processes and the role of phototropins into the uptake of calcium. Crystals 2020, 10, 591. [Google Scholar] [CrossRef]
- Marina, M.; Romero, F.M.; Villarreal, N.M.; Medina, A.J.; Gárriz, A.; Rossi, F.R.; Martinez, G.A.; Pieckenstain, F.L. Mechanisms of plant protection against two oxalate-producing fungal pathogens by oxalotrophic strains of Stenotrophomonas spp. Plant Mol. Biol. 2019, 100, 659–674. [Google Scholar] [CrossRef]
- Mazen, A.M.A.; El Maghraby, O.M.O. Accumulation of cadmium, lead and strontium, and a role of calcium oxalate in water hyacinth tolerance. Biol. Plant. 1998, 40, 411–417. [Google Scholar]
- Mazen, A.M.A. Calcium oxalate deposits in leaves of Corchorus olitotius as related to accumulation of toxic metals. Russ. J. Plant Physiol. 2004, 51, 281–285. [Google Scholar] [CrossRef]
- Leitenmaier, B.; Küpper, H. Compartmentation and complexation of metals in hyperaccumulator plants. Front. Plant Sci. 2013, 4, 374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korth, K.L.; Doege, S.J.; Park, S.-H.; Goggin, F.L.; Wang, Q.; Gomez, S.K.; Liu, G.; Jia, L.; Nakata, P.A. Medicago truncatula mutants demonstrate the role of plant calcium oxalate crystals as an effective defense against chewing insects. Plant Physiol. 2006, 141, 188–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, M.M.; Abdullah, R.B.; Wan Khadijah, W.E. A review of oxalate poisoning in domestic animals: Tolerance and performance aspects. J. Anim. Physiol. Anim. Nutr. 2013, 97, 605–614. [Google Scholar] [CrossRef]
- Helper, P.K.; Wayne, R.O. Calcium and plant development. Annu. Rev. Plant Physiol. 1985, 36, 397–439. [Google Scholar]
- Oryol, L.I.; Zhakova, M.A. The mechanism of anther dehiscence of tomato Lycopersicon esculentum Mill. (Solanaceae). Bot. Zhurnal 1977, 62, 1720–1739. (In Russian) [Google Scholar]
- Bonner, L.J.; Dickinson, H.G. Anther dehiscence in Lycopersicon esculentum Mill. I. Structural aspects. New Phytol. 1989, 113, 97–115. [Google Scholar] [CrossRef]
- Pacini, E. Relationships between tapetum, loculus, and pollen during development. Int. J. Plant Sci. 2010, 171, 1–11. [Google Scholar] [CrossRef]
- Burrieza, H.P.; López-Fernández, M.P.; Láinez, V.; Montenegro, T.; Maldonado, S. On the nature and origin of the oxalate package in Solanum sisymbriifolium anthers. Protoplasma 2010, 247, 45–56. [Google Scholar] [CrossRef]
- Wilson, Z.A.; Song, J.; Taylor, B.; Yang, C. The final split: The regulation of anther dehiscence. J. Exp. Bot. 2011, 62, 1633–1649. [Google Scholar] [CrossRef] [Green Version]
- Burchi, G.; Bauchan, G.R.; Murphy, C.; Roh, M.S. Characterization of calcium crystals in Abelia spp. using X-ray diffraction and electron microscopy. J. Hort. Sci. Biotech. 2014, 89, 61–68. [Google Scholar] [CrossRef]
- Du, M.; Zhou, K.; Liu, Y.; Deng, L.; Zhang, X.; Lin, L.; Zhou, M.; Zhao, W.; Wen, C.-L.; Xing, J.; et al. A biotechnology-based male-sterility system for hybrid seed production in tomato. Plant J. 2020, 102, 1090–1100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Ginkel, M.; Flipphi, R.C. Why self-fertilizing plants still exist in wild populations: Diversity assurance through stress-induced male sterility may promote selective outcrossing and recombination. Agronomy 2020, 10, 349. [Google Scholar] [CrossRef] [Green Version]
- Carroll, T.W.; Mayhew, D.E. Anther and pollen infection in relation to the pollen and seed transmissibility of two strains of barley stripe mosaic virus in barley. Can. J. Bot. 1976, 54, 1604–1621. [Google Scholar] [CrossRef]
- Hunter, D.G.; Bowyer, J.W. Cytopathology of developing anthers and pollen mother cells from lettuce plants infected by lettuce mosaic potyvirus. J. Phytopathol. 1997, 145, 521–524. [Google Scholar] [CrossRef]
- Stikić, R.; Jovanović, Z.; Vucelić-Radović, B.; Marjanović, M.; Savić, S. Tomato: A model species for fruit growth and development studies. Bot. Serbica 2015, 39, 95–102. [Google Scholar]
- Bohner, J.; Bangerth, F. Cell number, cell size and hormone level in semi-isogenic mutants of Lycopersicon pimpinellifolium differing in fruit size. Physiol. Plant. 1988, 72, 316–320. [Google Scholar] [CrossRef]
- Pesaresi, P.; Mizzotti, C.; Colombo, M.; Masiero, S. Genetic regulation and structural changes during tomato fruit development and ripening. Front. Plant Sci. 2014, 5, 124. [Google Scholar] [CrossRef] [Green Version]
- Gorguet, B.; Van Heusden, A.W.; Lindhout, P. Parthenocarpic fruit development in tomato. Plant Biol. 2005, 7, 131–139. [Google Scholar] [CrossRef]
- Serenko, E.K.; Baranova, E.N.; Balakhnina, T.I.; Kurenina, L.V.; Gulevich, A.A.; Kosobruhov, A.A.; Maysurian, A.N.; Polyakov, V.Y. Structural organization of chloroplast of tomato plants Solanum lycopersicum transformed by Fe-containing superoxide dismutase. Biochem. (Mosc.) Suppl. Ser. A Membr. Cell Biol. 2011, 5, 177–184. [Google Scholar] [CrossRef]
- Ekici, N.; Dane, F. Calcium oxalate crystals in vegetative and floral organs of Galanthus sp. (Amaryllidaceae). Asian J. Plant Sci. 2007, 6, 508–512. [Google Scholar]
- Ekici, N.; Feruza, D.A.N.E. Calcium oxalate crystals during development of male and female gametophyte in Leucojum aestivum (Amaryllidaceae). J. Appl. Biol. Sci. 2009, 3, 13–16. [Google Scholar]
- Baranova, E.N.; Kurenina, L.V.; Smirnov, A.N.; Beloshapkina, O.O.; Gulevich, A.A. Formation of the hypersensitivity response due to the expression of FeSOD1 gene in tomato when it is inoculated with Phytophthora infestans. Russ. Agric. Sci. 2017, 43, 15–21. [Google Scholar] [CrossRef]
- Bogoutdinova, L.R.; Lazareva, E.M.; Chaban, I.A.; Kononenko, N.V.; Dilovarova, T.A.; Khaliluev, M.R.; Kurenina, L.V.; Gulevich, A.A.; Smirnova, E.A.; Baranova, E.N. Salt stress-induced structural changes are mitigated in transgenic tomato plants over-expressing superoxide dismutase. Biology 2020, 9, 297. [Google Scholar] [CrossRef]
- Van Doorn, W.G.; Beers, E.P.; Dangl, J.L.; Franklin-Tong, V.E.; Gallois, P.; Hara-Nishimura, I.; Jones, A.M.; Kawai-Yamada, M.; Lam, E.; Mundy, J.; et al. Morphological classification of plant cell deaths. Cell Death Diff. 2011, 18, 1241–1246. [Google Scholar] [CrossRef] [Green Version]
- Khaliluev, M.R.; Chaban, I.A.; Kononenko, N.V.; Baranova, E.N.; Dolgov, S.V.; Kharchenko, P.N.; Polyakov, V.Y. Abnormal floral meristem development in transgenic tomato plants do not depend on the expression of genes encoding defense-related PR-proteins and antimicrobial peptides. Rus. J. Dev. Biol. 2014, 45, 22–33. [Google Scholar] [CrossRef]
- Katayama, H.; Fujibayashi, S.; Sugimura, Y. Cell wall sheath surrounding calcium oxalate crystals in mulberry idioblasts. Protoplasma 2007, 231, 245–248. [Google Scholar] [CrossRef]
- Gębura, J.; Winiarczyk, K. A study on calcium oxalate crystals in Tinantia anomala (Commelinaceae) with special reference to ultrastructural changes during anther development. J. Plant Res. 2016, 129, 685–695. [Google Scholar] [CrossRef]
- Borchert, R. Calcium acetate induces calcium uptake and formation of calcium-oxalate crystals in isolated leaflets of Gleditsia triacanthos L. Planta 1986, 168, 571–578. [Google Scholar] [CrossRef]
- Fink, S. Comparative microscopical studies on the patterns of calcium oxalate distribution in the needles of various conifer species. Plant Biol. 1991, 104, 306–315. [Google Scholar] [CrossRef]
- Horner, H.T.; Kausch, A.P.; Wagner, B.L. Ascorbic acid: A precursor of oxalate in crystal idioblasts of Yucca torreyi in liquid root culture. Int. J. Plant Sci. 2000, 161, 861–868. [Google Scholar] [CrossRef]
- Kostman, T.A.; Tarlin, N.M.; Franceschi, V.R. Autoradiography utilizing labeled ascorbic acid reveals biochemical and morphological details in diverse calcium oxalate crystal-forming species. Funct. Plant Biol. 2007, 34, 339–342. [Google Scholar] [CrossRef] [PubMed]
- Arnott, H.J.; Pautard, F.G.E. (Eds.) Calcification in Plants. In Biological Calcification: Cellular and Molecular Aspects; Springer: Boston, MA, USA, 1970; pp. 375–446. [Google Scholar]
- Kausch, A.P.; Horner, H.T. The development of mucilaginous raphide crystal idioblasts in young leaves of Typha angustifolia L. (Typhaceae). Am. J. Bot. 1983, 70, 691–705. [Google Scholar] [CrossRef]
- Yang, X.; Yang, J.; Wang, Y.; He, H.; Niu, L.; Guo, D.; Xing, G.; Zhao, Q.; Zhong, X.; Sui, L.; et al. Enhanced resistance to sclerotinia stem rot in transgenic soybean that overexpresses a wheat oxalate oxidase. Transgenic Res. 2019, 28, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Paiva, E.A.S. Are calcium oxalate crystals a dynamic calcium store in plants? New Phytol. 2019, 223, 1707–1711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ledbetter, M.C.; Porter, K.R. Introduction to the Fine Structure of Plant Cells; Springer: Berlin/Heidelberg, Germany, 2012; p. 188. [Google Scholar]
- Meric, C.; Dane, F. Calcium oxalate crystals in floral organs of Helianthus annuus L. and H. tuberosus L. (Asteraceae). Acta Biol. Szeged. 2004, 48, 19–23. [Google Scholar]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baranova, E.N.; Chaban, I.A.; Kurenina, L.V.; Konovalova, L.N.; Varlamova, N.V.; Khaliluev, M.R.; Gulevich, A.A. Possible Role of Crystal-Bearing Cells in Tomato Fertility and Formation of Seedless Fruits. Int. J. Mol. Sci. 2020, 21, 9480. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21249480
Baranova EN, Chaban IA, Kurenina LV, Konovalova LN, Varlamova NV, Khaliluev MR, Gulevich AA. Possible Role of Crystal-Bearing Cells in Tomato Fertility and Formation of Seedless Fruits. International Journal of Molecular Sciences. 2020; 21(24):9480. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21249480
Chicago/Turabian StyleBaranova, Ekaterina N., Inna A. Chaban, Ludmila V. Kurenina, Ludmila N. Konovalova, Natalia V. Varlamova, Marat R. Khaliluev, and Alexander A. Gulevich. 2020. "Possible Role of Crystal-Bearing Cells in Tomato Fertility and Formation of Seedless Fruits" International Journal of Molecular Sciences 21, no. 24: 9480. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21249480