Coffee Extends Yeast Chronological Lifespan through Antioxidant Properties

 , , and
, , and
Abstract
:1. Introduction
2. Results
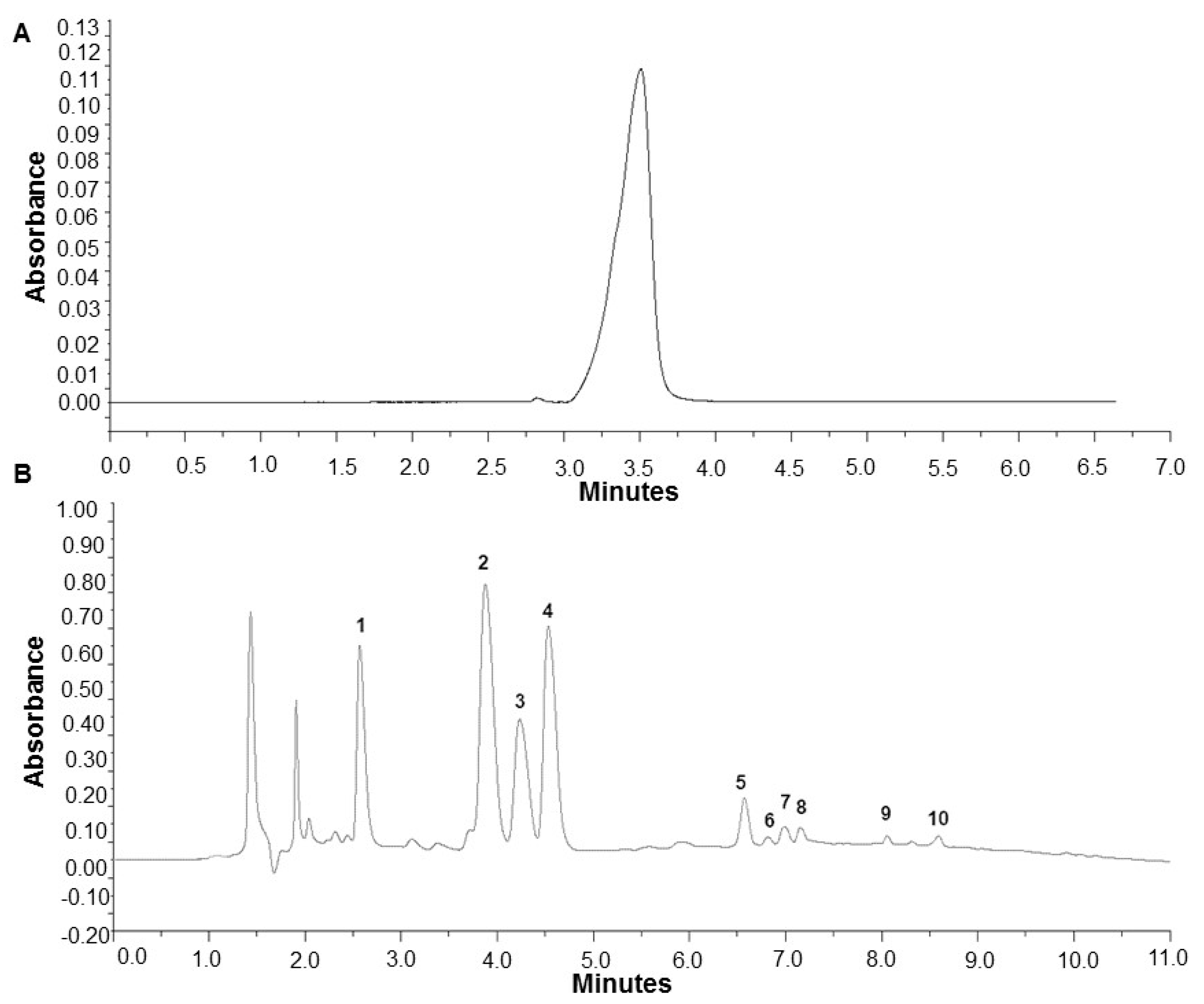
2.1. Analysis of Caffeine and Polyphenols in Specialty Coffee Infusions
2.2. Antioxidant Properties of Coffee Infusions
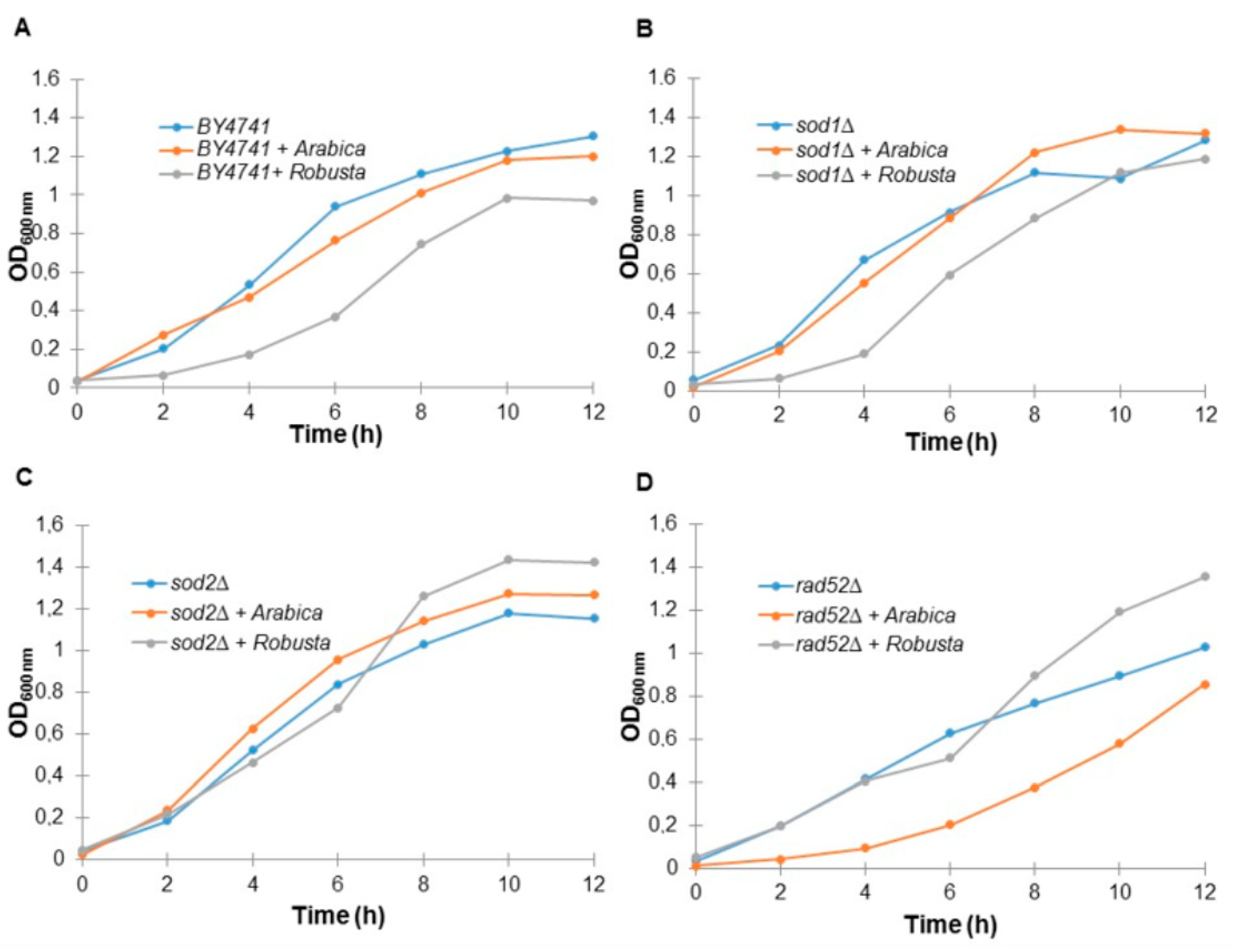
2.3. Coffee Infusion Reduces Yeast Cell Growth
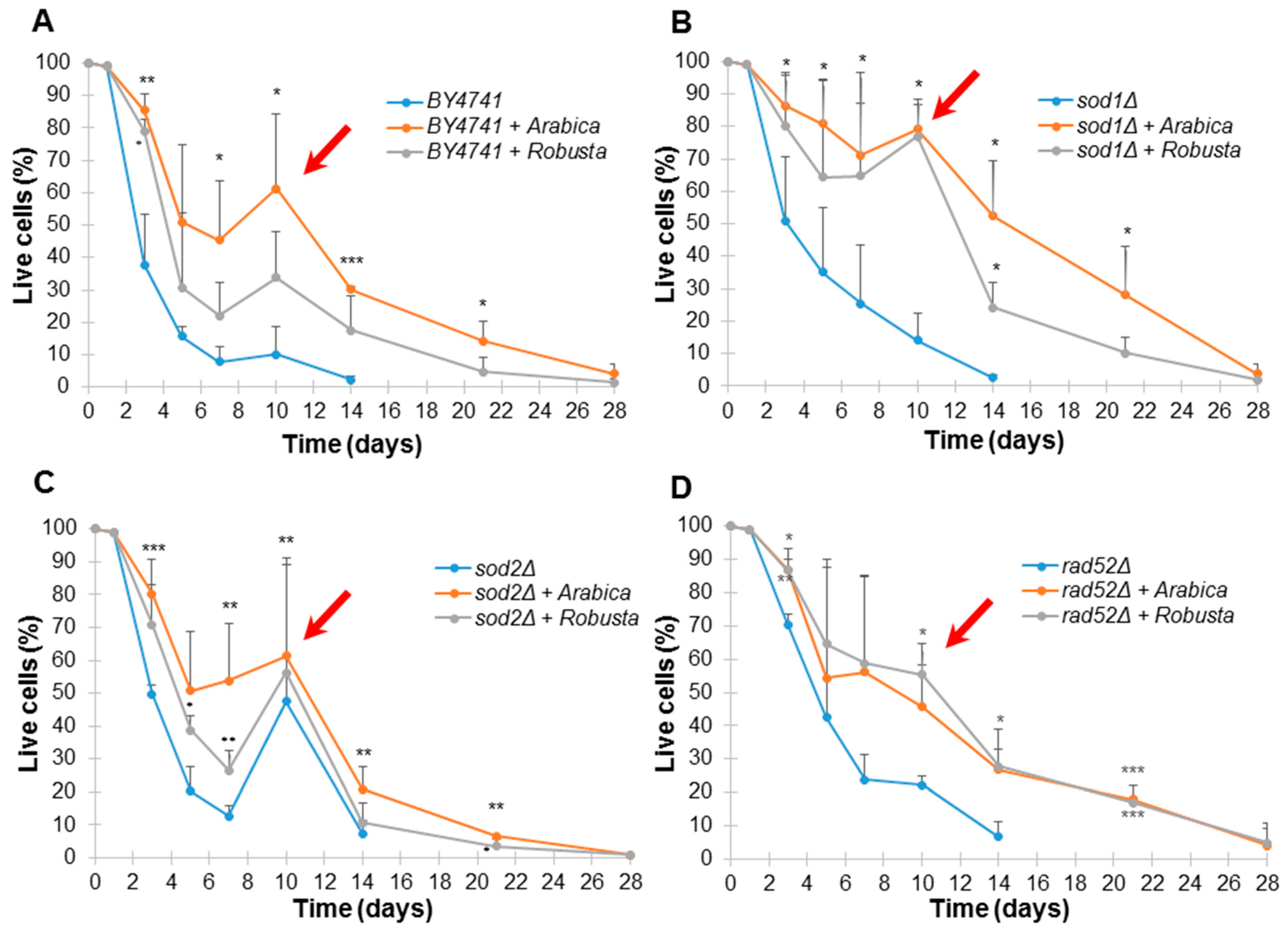
2.4. Impact of Coffee on Saccharomyces Cerevisiae Yeast Aging
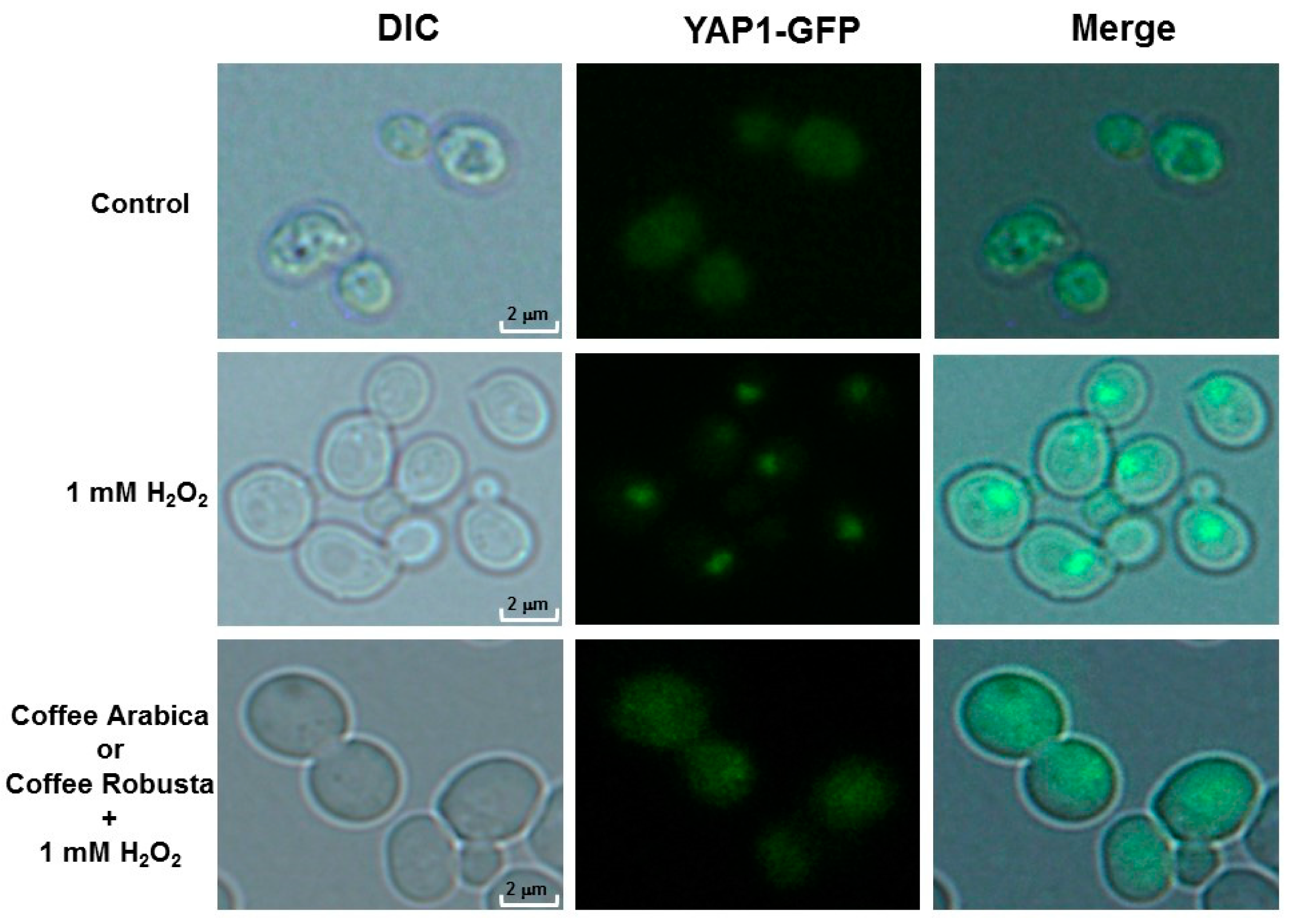
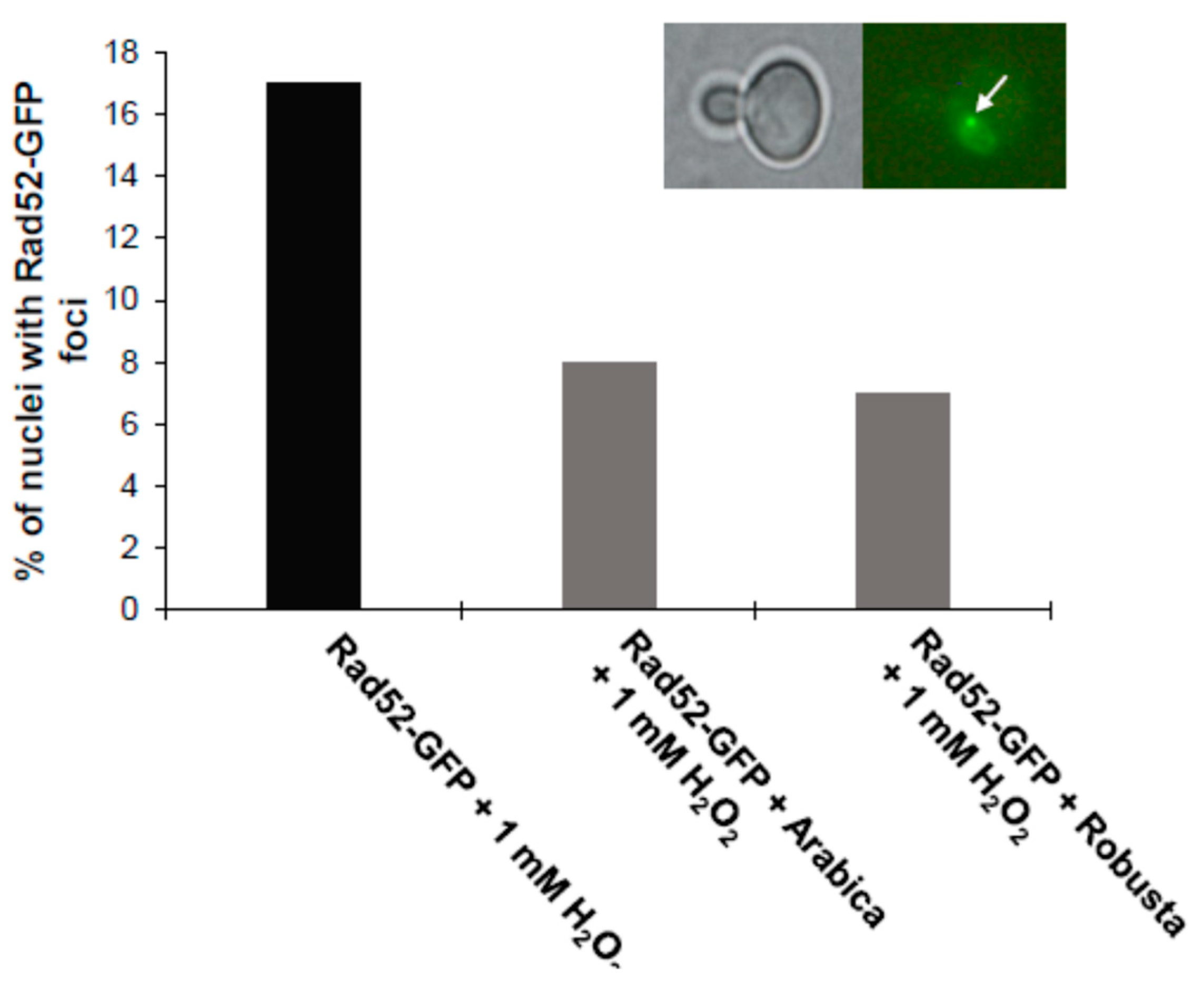
2.5. Coffee Protects Cells from Oxidative Stress and DNA Damage
2.6. Impact of Coffee on Sensitivity to Stress Factors
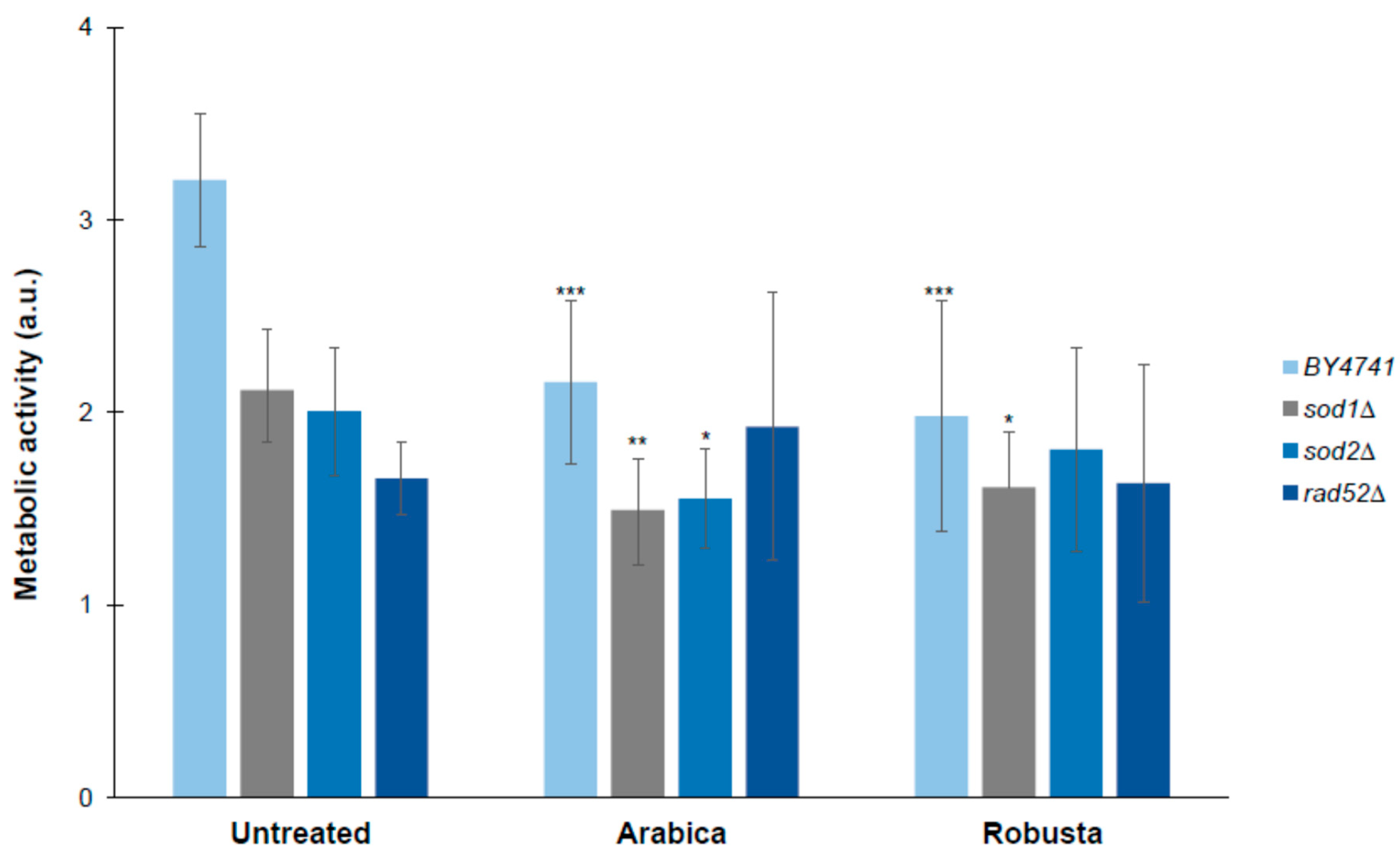
2.7. Impact of Coffee on the Metabolic Activity of Yeast
3. Discussion
4. Materials and Methods
4.1. Preparation of Coffee Infusions
4.2. Preparing Infusion with a Dripper
4.3. Determination of Caffeine and Polyphenols Concentration in Coffee Infusions Using HPLC
4.4. Determination by the ABTS Radical Method
4.5. Determination by the FRAP Assay
4.6. Determination of DPPH Free Radical Scavenging Activity
4.7. Total Phenolic Content
4.8. Strains and Growth Conditions
4.9. Kinetics of the Growth Assay
4.10. Chronological Life Span (CLS) Assays
4.11. Cell Viability
4.12. Phenotypic Analysis—A Spot Test for Sensitivity to Congo Red, Calcofluor White, Methyl Methanesulfonate (MMS), Sodium Chloride (NaCl) and Acetic Acid
4.13. Measurement of Cell Metabolic Activity
4.14. Cellular Localization of the Yap1-GFP and Rad52-GFP Proteins
4.15. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Gunter, M.J.; Murphy, N.; Cross, A.J.; Dossus, L.; Dartois, L.; Fagherazzi, G.; Kaaks, R.; Kuhn, T.; Boeing, H.; Aleksandrova, K.; et al. Coffee Drinking and Mortality in 10 European Countries A Multinational Cohort Study. Ann. Intern. Med. 2017, 167, 236–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Privat, I.; Foucrier, S.; Prins, A.; Epalle, T.; Eychenne, M.; Kandalaft, L.; Caillet, V.; Lin, C.W.; Tanksley, S.; Foyer, C.; et al. Differential regulation of grain sucrose accumulation and metabolism in Coffea arabica (Arabica) and Coffea canephora (Robusta) revealed through gene expression and enzyme activity analysis. New Phytol. 2008, 178, 781–797. [Google Scholar] [CrossRef] [PubMed]
- Perrois, C.; Strickler, S.R.; Mathieu, G.; Lepelley, M.; Bedon, L.; Michaux, S.; Husson, J.; Mueller, L.; Privat, I. Differential regulation of caffeine metabolism in Coffea arabica (Arabica) and Coffea canephora (Robusta). Planta 2015, 241, 179–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarini, L.; Gilli, R.; Gombac, V.; Abatangelo, A.; Bosco, M.; Toffanin, R. Polysaccharides from hot water extracts of roasted Coffea arabica beans: Isolation and characterization. Carbohydr. Polym. 1999, 40, 71–811999. [Google Scholar] [CrossRef]
- Kang, D.E.; Lee, H.U.; Davaatseren, M.; Chung, M.S. Comparison of acrylamide and furan concentrations, antioxidant activities, and volatile profiles in cold or hot brew coffees. Food Sci. Biotechnol. 2020, 29, 141–148. [Google Scholar] [CrossRef]
- Spencer, M.; Sage, E.; Velez, M.; Guinard, J.X. Using Single Free Sorting and Multivariate Exploratory Methods to Design a New Coffee Taster’s Flavor Wheel. J. Food Sci. 2016, 81, S2997–S3005. [Google Scholar] [CrossRef] [Green Version]
- Heckman, M.A.; Weil, J.; de Mejia, E.G. Caffeine (1, 3, 7-trimethylxanthine) in Foods: A Comprehensive Review on Consumption, Functionality, Safety, and Regulatory Matters. J. Food Sci. 2010, 75, R77–R87. [Google Scholar] [CrossRef]
- Frischknecht, P.M.; Ulmerdufek, J.; Baumann, T.W. Purine Alkaloid Formation in Buds and Developing Leaflets of Coffea-arabica—Expression of an Optimal Defense Strategy. Phytochemistry 1986, 25, 613–616. [Google Scholar] [CrossRef]
- Ricci, E.; Vigano, P.; Cipriani, S.; Somigliana, E.; Chiaffarino, F.; Bulfoni, A.; Parazzini, F. Coffee and caffeine intake and male infertility: A systematic review. Nutr. J. 2017, 16. [Google Scholar] [CrossRef] [Green Version]
- Nehlig, A. Interindividual Differences in Caffeine Metabolism and Factors Driving Caffeine Consumption. Pharmacol. Rev. 2018, 70, 384–411. [Google Scholar] [CrossRef] [Green Version]
- Yang, A.; Palmer, A.; de Wit, H. Genetics of caffeine consumption and responses to caffeine. Psychopharmacology 2010, 211, 245–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kot, M.; Daniel, W.A. Caffeine as a marker substrate for testing cytochrome P450 activity in human and rat. Pharmacol. Rep. 2008, 60, 789–797. [Google Scholar] [PubMed]
- Cappelletti, S.; Daria, P.; Sani, G.; Aromatario, M. Caffeine: Cognitive and Physical Performance Enhancer or Psychoactive Drug? Curr. Neuropharmacol. 2015, 13, 71–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DePaula, J.; Farah, A. Caffeine Consumption through Coffee: Content in the Beverage, Metabolism, Health Benefits and Risks. Beverages 2019, 5, 37. [Google Scholar] [CrossRef] [Green Version]
- Agostoni, C.; Canani, R.B.; Fairweather-Tait, S.; Heinonen, M.; Korhonen, H.; La Vieille, S.; Marchelli, R.; Martin, A.; Naska, A.; Neuhauser-Berthold, M.; et al. Scientific Opinion on the safety of caffeine. EFSA J. 2015, 13. [Google Scholar] [CrossRef] [Green Version]
- Ferruzzi, M.G. The influence of beverage composition on delivery of phenolic compounds from coffee and tea. Physiol. Behav. 2010, 100, 33–41. [Google Scholar] [CrossRef]
- Takahashi, K.; Ishigami, A. Anti-aging effects of coffee. Aging Us 2017, 9, 1863–1864. [Google Scholar] [CrossRef] [Green Version]
- Buldak, R.J.; Hejmo, T.; Osowski, M.; Buldak, L.; Kukla, M.; Polaniak, R.; Birkner, E. The Impact of Coffee and Its Selected Bioactive Compounds on the Development and Progression of Colorectal Cancer In Vivo and In Vitro. Molecules 2018, 23, 3309. [Google Scholar] [CrossRef] [Green Version]
- Welsh, E.J.; Bara, A.; Barley, E.; Cates, C.J. Caffeine for asthma. Cochrane Database Syst. Rev. 2010. [Google Scholar] [CrossRef] [Green Version]
- Roehrs, T.; Roth, T. Caffeine: Sleep and daytime sleepiness. Sleep Med. Rev. 2008, 12, 153–162. [Google Scholar] [CrossRef]
- Yadegari, M.; Khazaei, M.; Anvari, M.; Eskandari, M. Prenatal Caffeine Exposure Impairs Pregnancy in Rats. Int. J. Fertil. Steril. 2016, 9, 558–562. [Google Scholar] [PubMed]
- Sengpiel, V.; Elind, E.; Bacelis, J.; Nilsson, S.; Grove, J.; Myhre, R.; Haugen, M.; Meltzer, H.M.; Alexander, J.; Jacobsson, B.; et al. Maternal caffeine intake during pregnancy is associated with birth weight but not with gestational length: Results from a large prospective observational cohort study. BMC Med. 2013, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harman, D. Aging—A Theory Based on Free-Radical and Radiation-Chemistry. J. Gerontol. 1956, 11, 298–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaeberlein, M. Lessons on longevity from budding yeast. Nature 2010, 464, 513–519. [Google Scholar] [CrossRef] [Green Version]
- Fabrizio, P.; Longo, V.D. The chronological life span of Saccharomyces cerevisiae. Aging Cell 2003, 2, 73–81. [Google Scholar] [CrossRef]
- Fabrizio, P.; Pletcher, S.D.; Minois, N.; Vaupel, J.W.; Longo, V.D. Chronological aging-independent replicative life span regulation by Msn2/Msn4 and Sod2 in Saccharomyces cerevisiae. Febs Lett. 2004, 557, 136–142. [Google Scholar] [CrossRef] [Green Version]
- Longo, V.D.; Shadel, G.S.; Kaeberlein, M.; Kennedy, B. Replicative and Chronological Aging in Saccharomyces cerevisiae. Cell Metab. 2012, 16, 18–31. [Google Scholar] [CrossRef] [Green Version]
- Loomis, D.; Guyton, K.Z.; Grosse, Y.; Lauby-Secretan, B.; El Ghissassi, F.; Bouvard, V.; Benbrahim-Tallaa, L.; Guha, N.; Mattock, H.; Straif, K.; et al. Carcinogenicity of drinking coffee, mate, and very hot beverages. Lancet Oncol. 2016, 17, 877–878. [Google Scholar] [CrossRef]
- Crippa, A.; Discacciati, A.; Larsson, S.C.; Wolk, A.; Orsini, N. Coffee Consumption and Mortality from All Causes, Cardiovascular Disease, and Cancer: A Dose-Response Meta-Analysis. Am. J. Epidemiol. 2014, 180, 763–775. [Google Scholar] [CrossRef] [Green Version]
- Von Zglinicki, T. Role of oxidative stress in telomere length regulation and replicative senescence. Mol. Cell. Gerontol. 2000, 908, 99–110. [Google Scholar] [CrossRef]
- Yashin, A.; Yashin, Y.; Wang, J.Y.; Nemzer, B. Antioxidant and Antiradical Activity of Coffee. Antioxidants 2013, 2, 230–245. [Google Scholar] [CrossRef] [PubMed]
- Janda, K.; Jakubczyk, K.; Baranowska-Bosiacka, I.; Kapczuk, P.; Kochman, J.; Rebacz-Maron, E.; Gutowska, I. Mineral Composition and Antioxidant Potential of Coffee Beverages Depending on the Brewing Method. Foods 2020, 9, 121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acidri, R.; Sawai, Y.; Sugimoto, Y.; Handa, T.; Sasagawa, D.; Masunaga, T.; Yamamoto, S.; Nishihara, E. Phytochemical Profile and Antioxidant Capacity of Coffee Plant Organs Compared to Green and Roasted Coffee Beans. Antioxidants 2020, 9, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorecki, M.; Hallmann, E. The Antioxidant Content of Coffee and Its In Vitro Activity as an Effect of Its Production Method and Roasting and Brewing Time. Antioxidants 2020, 9, 308. [Google Scholar] [CrossRef] [Green Version]
- Pokorna, J.; Venskutonis, P.R.; Kraujalyte, V.; Kraujalis, P.; Dvorak, P.; Tremlova, B.; Kopriva, V.; Ostadalova, M. Comparison of Different Methods of Antioxidant Activity Evaluation of Green and Roast C. arabica and C. robusta Coffee Beans. Acta Aliment. 2015, 44, 454–460. [Google Scholar] [CrossRef] [Green Version]
- Vignoli, J.A.; Viegas, M.C.; Bassoli, D.G.; Benassi, M.D. Roasting process affects differently the bioactive compounds and the antioxidant activity of arabica and robusta coffees. Food Res. Int. 2014, 61, 279–285. [Google Scholar] [CrossRef] [Green Version]
- Stalmach, A.; Mullen, W.; Nagai, C.; Crozier, A. On-line HPLC analysis of the antioxidant activity of phenolic compounds in brewed, paper-filtered coffee. Braz. J. Plant Physiol. 2006, 18, 253–262. [Google Scholar] [CrossRef] [Green Version]
- Belguidoum, K.; Amira-Guebailia, H.; Boulmokh, Y.; Houache, O. HPLC coupled to UV-vis detection for quantitative determination of phenolic compounds and caffeine in different brands of coffee in the Algerian market. J. Taiwan Inst. Chem. Eng. 2014, 45, 1314–1320. [Google Scholar] [CrossRef]
- Wanke, V.; Cameroni, E.; Uotila, A.; Piccolis, M.; Urban, J.; Loewith, R.; De Virgilio, C. Caffeine extends yeast lifespan by targeting TORC1. Mol. Microbiol. 2008, 69, 277–285. [Google Scholar] [CrossRef] [Green Version]
- Molon, M.; Zadrag-Tecza, R.; Bilinski, T. The longevity in the yeast Saccharomyces cerevisiae: A comparison of two approaches for assessment the lifespan. Biochem. Biophys. Res. Commun. 2015, 460, 651–656. [Google Scholar] [CrossRef]
- Hwangbo, D.S.; Lee, H.Y.; Abozaid, L.S.; Min, K.J. Mechanisms of Lifespan Regulation by Calorie Restriction and Intermittent Fasting in Model Organisms. Nutrients 2020, 12, 1194. [Google Scholar] [CrossRef] [PubMed]
- Selvarani, R.; Mohammed, S.; Richardson, A. Effect of rapamycin on aging and age-related diseases-past and future. Geroscience 2020. [Google Scholar] [CrossRef] [PubMed]
- Shukitt-Hale, B.; Miller, M.G.; Chu, Y.F.; Lyle, B.J.; Joseph, J.A. Coffee, but not caffeine, has positive effects on cognition and psychomotor behavior in aging. Age 2013, 35, 2183–2192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, C.H.; Wang, L.; Lin, X.Y.; Mamcarz, M.; Zhang, C.; Bai, G.; Nong, J.; Sussman, S.; Arendash, G. Caffeine Synergizes with Another Coffee Component to Increase Plasma GCSF: Linkage to Cognitive Benefits in Alzheimer’s Mice. J. Alzheimers Dis. 2011, 25, 323–335. [Google Scholar] [CrossRef] [Green Version]
- Toone, W.M.; Jones, N. AP-1 transcription factors in yeast. Curr. Opin. Genet. Dev. 1999, 9, 55–61. [Google Scholar] [CrossRef]
- Yan, C.; Lee, L.H.; Davis, L.I. Crm1p mediates regulated nuclear export of a yeast AP-1-life transcription factor. Embo Jl 1998, 17, 7416–7429. [Google Scholar] [CrossRef]
- Gulshan, K.; Rovinsky, S.A.; Coleman, S.T.; Moye-Rowley, W.S. Oxidant-specific folding of Yap1p regulates both transcriptional activation and nuclear localization. J. Biol. Chem. 2005, 280, 40524–40533. [Google Scholar] [CrossRef] [Green Version]
- Chu, Y.F.; Brown, P.H.; Lyle, B.J.; Chen, Y.M.; Black, R.M.; Williams, C.E.; Lin, Y.C.; Hsu, C.W.; Cheng, I.H. Roasted Coffees High in Lipophilic Antioxidants and Chlorogenic Acid Lactones Are More Neuroprotective than Green Coffees. J. Agric. Food Chem. 2009, 57, 9801–9808. [Google Scholar] [CrossRef]
- Azam, S.; Hadi, N.; Khan, N.U.; Hadi, S.M. Antioxidant and prooxidant properties of caffeine, theobromine and xanthine. Med Sci. Monit. Int. Med. J. Exp. Clin. Res. 2003, 9, BR325–BR330. [Google Scholar]
- Li, H.M.; Roxo, M.; Cheng, X.L.; Zhang, S.X.; Cheng, H.R.; Wink, M. Pro-oxidant and lifespan extension effects of caffeine and related methylxanthines in Caenorhabditis elegans. Food Chem. X 2019, 1. [Google Scholar] [CrossRef]
- Bridi, J.C.; Barros, A.G.D.; Sampaio, L.R.; Ferreira, J.C.D.; Soares, F.A.A.; Romano-Silva, M.A. Lifespan Extension Induced by Caffeine in Caenorhabditis elegans is Partially Dependent on Adenosine Signaling. Front. Aging Neurosci. 2015, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sutphin, G.L.; Bishop, E.; Yanos, M.E.; Moller, R.M.; Kaeberlein, M. Caffeine extends life span, improves healthspan, and delays age-associated pathology in Caenorhabditis elegans. Longev. Healthspan 2012, 1, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peixoto, H.; Roxo, M.; Krstin, S.; Roehrig, T.; Richling, E.; Wink, M. An Anthocyanin-Rich Extract of Acai (Euterpe precatoria Mart.) Increases Stress Resistance and Retards Aging-Related Markers in Caenorhabditis elegans. J. Agric. Food Chem. 2016, 64, 1283–1290. [Google Scholar] [CrossRef] [PubMed]
- Lisby, M.; Rothstein, R.; Mortensen, U.H. Rad52 forms DMA repair and recombination centers during S phase. Proc. Natl. Acad. Sci. USA 2001, 98, 8276–8282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molon, M.; Szajwaj, M.; Tchorzewski, M.; Skoczowski, A.; Niewiadomska, E.; Zadrag-Tecza, R. The rate of metabolism as a factor determining longevity of the Saccharomyces cerevisiae yeast. Age 2016, 38. [Google Scholar] [CrossRef] [PubMed]
- Mirisola, M.G.; Longo, V.D. Acetic acid and acidification accelerate chronological and replicative aging in yeast. Cell Cycle 2012, 11, 3532–3533. [Google Scholar] [CrossRef]
- Burhans, W.C.; Weinberger, M. Acetic acid effects on aging in budding yeast. Cell Cycle 2009, 8, 2300–2302. [Google Scholar] [CrossRef] [Green Version]
- Yamagata, K. Do Coffee Polyphenols Have a Preventive Action on Metabolic Syndrome Associated Endothelial Dysfunctions? An Assessment of the Current Evidence. Antioxidants 2018, 7, 26. [Google Scholar] [CrossRef] [Green Version]
- Liang, N.J.; Kitts, D.D. Antioxidant Property of Coffee Components: Assessment of Methods that Define Mechanisms of Action. Molecules 2014, 19, 19180–19208. [Google Scholar] [CrossRef] [Green Version]
- Patay, E.B.; Bencsik, T.; Papp, N. Phytochemical overview and medicinal importance of Coffea species from the past until now. Asian Pac. J. Trop. Med. 2016, 9, 1127–1135. [Google Scholar] [CrossRef] [Green Version]
- Gasscht, F.; Dicato, M.; Diederich, M. Coffee provides a natural multitarget pharmacopeia against the hallmarks of cancer. Genes Nutr. 2015, 10. [Google Scholar] [CrossRef]
- Martini, D.; Del Bo, C.; Tassotti, M.; Riso, P.; Del Rio, D.; Brighenti, F.; Porrini, M. Coffee Consumption and Oxidative Stress: A Review of Human Intervention Studies. Molecules 2016, 21, 979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotyczka, C.; Boettler, U.; Lang, R.; Stiebitz, H.; Bytof, G.; Lantz, I.; Hofmann, T.; Marko, D.; Somoza, V. Dark roast coffee is more effective than light roast coffee in reducing body weight, and in restoring red blood cell vitamin E and glutathione concentrations in healthy volunteers. Mol. Nutr. Food Res. 2011, 55, 1582–1586. [Google Scholar] [CrossRef] [PubMed]
- Steinkellner, H.; Hoelzl, C.; Uhl, M.; Cavin, C.; Haidinger, G.; Gsur, A.; Schmid, R.; Kundi, M.; Bichler, J.; Knasmuller, S. Coffee consumption induces GSTP in plasma and protects lymphocytes against (+/-)-anti-benzo a pyrene-7,8-dihydrodiol-9,10-epoxide induced DNA-damage: Results of controlled human intervention trials. Mutat. Res Fundam. Mol. Mech. Mutagen. 2005, 591, 264–275. [Google Scholar] [CrossRef]
- Yukawa, G.S.; Mune, M.; Otani, H.; Tone, Y.; Liang, X.M.; Iwahashi, H.; Sakamoto, W. Effects of coffee consumption on oxidative susceptibility of low-density lipoproteins and serum lipid levels in humans. Biochem. Mosc. 2004, 69, 70–74. [Google Scholar] [CrossRef]
- Cardin, R.; Piciocchi, M.; Martines, D.; Scribano, L.; Petracco, M.; Farinati, F. Effects of coffee consumption in chronic hepatitis C: A randomized controlled trial. Dig. Liver Dis. 2013, 45, 499–504. [Google Scholar] [CrossRef]
- Fajara, B.E.P.; Susanti, H. HPLC determination of caffeine in coffee beverage. Conf. Ser. Mater. Sci. Eng. 2017, 259, 012011. [Google Scholar]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of ‘‘antioxidant power’’: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Blois, M.S. Antioxidant determinations by the use of a stable free radical. Nature 1958, 181, 1199–1200. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A., Jr. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Viticult. 1965, 16, 144–158. [Google Scholar]
- Kwolek-Mirek, M.; Zadrag-Tecza, R. Comparison of methods used for assessing the viability and vitality of yeast cells. FEMS Yeast Res. 2014, 14, 1068–1079. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Average Retention Time (min) | Quantity | |
|---|---|---|---|
| Arabica | Robusta | ||
| Caffeine (mg/mL) | |||
| caffeine | 3.5 | 1.63 | 2.55 |
| Polyphenols (μg/mL) | |||
| neochlorogenic acid | 2.56 | 79.07 | 167.21 |
| cryptochlorogenic acid | 3.87 | 224.34 | 277.41 |
| chlorogenic acid | 4.22 | 225.21 | 150.49 |
| ferulic acid | 6.58 | 35.13 | 110.18 |
| dicaffeoylquinic or feruloylquinic acids isomers | 6.82 | 5.06 | 7.28 |
| 6.99 | 12.85 | 22.72 | |
| 7.16 | 11.06 | 16.93 | |
| 8.07 | 3.66 | 11.68 | |
| 8.33 | 1.61 | 7.16 | |
| 8.60 | 7.19 | 25.98 | |
| Method | Robusta Coffee | Arabica Coffee | Caffeine 1 mg/mL | Caffeine 10 mg/mL |
|---|---|---|---|---|
| TPC (mg GAE/mL) | 3.11 ± 0.22 b | 2.08 ± 0.07 a | 0 | 0 |
| ABTS (μmol Trolox/g of sample) | 413.49 ± 11.26 b | 258.41 ± 5.56 a | 0 | 0 |
| FRAP (μmol Trolox/g of sample) | 392.87 ± 17.75 b | 262.6 ± 6.73 a | 0 | 0 |
| DPPH (%) | 46.09 ± 2.93 b | 27.97 ± 3.06 a | 1.73 ± 0.28 c | 2.40 ± 0.27 d |
| Strain | Growth Conditions | |||||
|---|---|---|---|---|---|---|
| Congo Red 15 μg/mL | Congo Red 15 µg/mL + Arabica | Congo Red 15 µg/mL + Robusta | Calcofluor White 20 µg/mL | Calcofluor White 20 µg/mL + Arabica | Calcofluor White 20 µg/mL + Robusta | |
| BY4741 | ++++ | ++++ | +++ | ++++ | ++++ | ++++ |
| sod1Δ | ++ | +++ | ++++ | ++ | ++++ | ++++ |
| sod2Δ | ++++ | ++++ | +++ | ++++ | ++++ | ++++ |
| rad52Δ | +++ | ++++ | ++++ | +++ | +++ | ++++ |
| Strain | NaCl 0.5 M | NaCl 0.5 M + Arabica | NaCl 0.5 M + Robusta | CH3COOH 40 mM | CH3COOH 40 mM + Arabica | CH3COOH 40 mM + Robusta |
| BY4741 | ++++ | +++ | ++ | +++ | +++ | ++ |
| sod1Δ | ++++ | +++ | + | ++ | ++ | + |
| sod2Δ | ++++ | +++ | +++ | +++ | ++++ | ++ |
| rad52Δ | ++++ | +++ | ++ | +++ | ++++ | +++ |
| Strain | MMS 0.03% | MMS 0.03% + Arabica | MMS 0.03% + Robusta | Control | ||
| BY4741 | +++ | ++++ | ++++ | ++++ | ||
| sod1Δ | ++ | ++++ | +++ | ++++ | ||
| sod2Δ | ++++ | ++++ | ++++ | ++++ | ||
| rad52Δ | - | - | - | ++++ | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Czachor, J.; Miłek, M.; Galiniak, S.; Stępień, K.; Dżugan, M.; Mołoń, M. Coffee Extends Yeast Chronological Lifespan through Antioxidant Properties. Int. J. Mol. Sci. 2020, 21, 9510. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21249510
Czachor J, Miłek M, Galiniak S, Stępień K, Dżugan M, Mołoń M. Coffee Extends Yeast Chronological Lifespan through Antioxidant Properties. International Journal of Molecular Sciences. 2020; 21(24):9510. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21249510
Chicago/Turabian StyleCzachor, Jadwiga, Michał Miłek, Sabina Galiniak, Karolina Stępień, Małgorzata Dżugan, and Mateusz Mołoń. 2020. "Coffee Extends Yeast Chronological Lifespan through Antioxidant Properties" International Journal of Molecular Sciences 21, no. 24: 9510. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21249510