Extracellular RNAs in Bacterial Infections: From Emerging Key Players on Host-Pathogen Interactions to Exploitable Biomarkers and Therapeutic Targets


Abstract
:1. Introduction
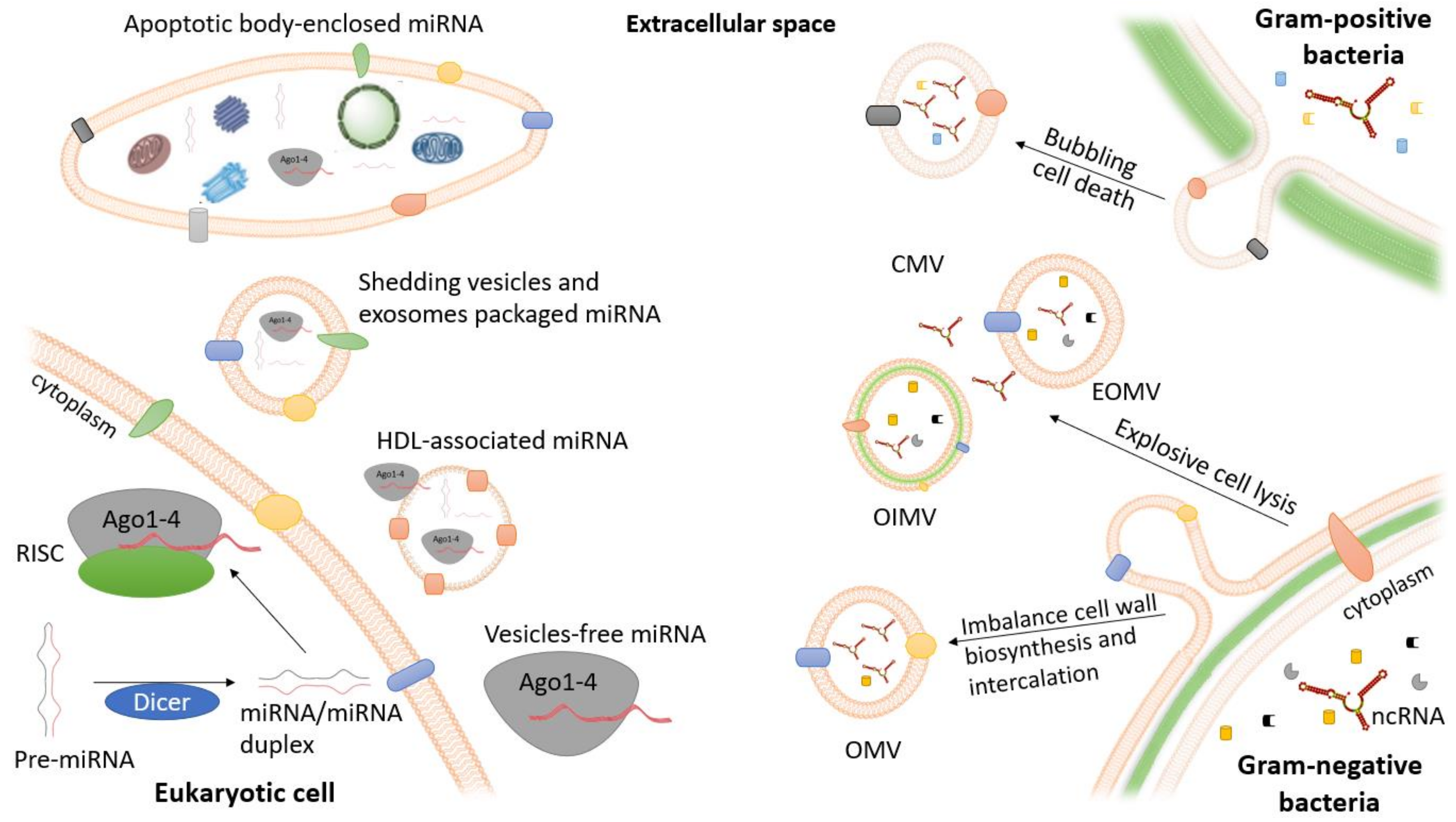
2. Extracellular ncRNAs in Eukaryotes: Release Mechanisms and Relevance in Bacterial Infections
3. Extracellular ncRNAs in Bacteria
3.1. OMVs Biogenesis
3.2. OMVs Composition and RNA Secretion
3.3. Specific Examples of Bacterial Secreted Micro-Size RNAs
3.3.1. Escherichia coli
3.3.2. Pseudomonas aeruginosa
3.3.3. Helicobacter pylori
3.3.4. Listeria monocytogenes
3.3.5. Salmonella sp.
3.3.6. Mycobacterium tuberculosis
3.3.7. Periodontal Pathogens: Aggregatibacter actinomycetemcomitans, Porphyromonas gingivalis, Treponema denticola and Streptococcus sanguinis
3.3.8. Additional Examples
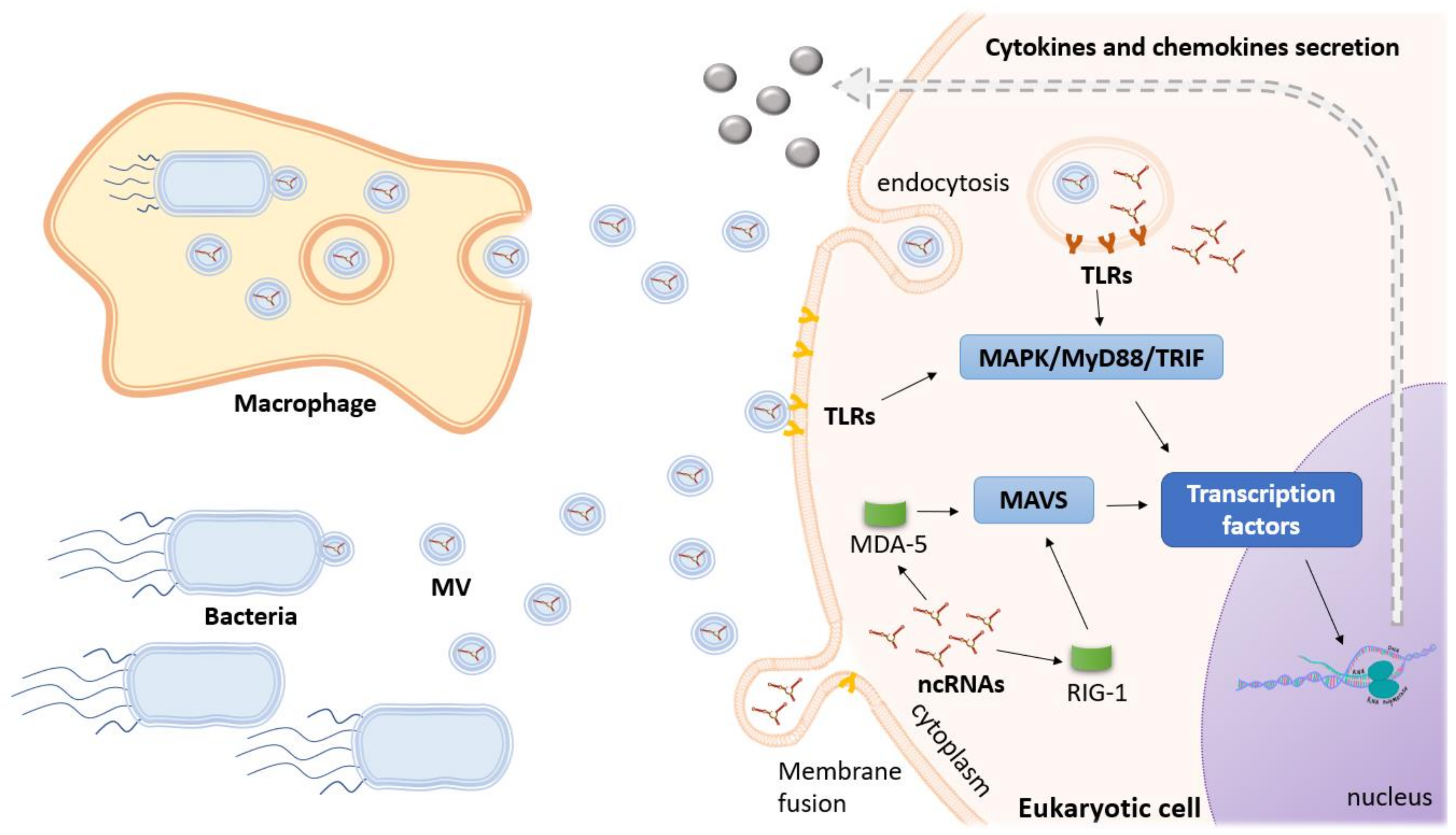
4. seRNAs in Novel Diagnostic and Therapeutic Approaches
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ledda, B.; Ottaggio, L.; Izzotti, A.; Sukkar, S.G.; Miele, M. Small RNAs in eucaryotes: New clues for amplifying microRNA benefits. Cell Biosci. 2020, 10, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carthew, R.W.; Sontheimer, E.J. Origins and Mechanisms of miRNAs and siRNAs. Cell 2009, 136, 642–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, Y.; Fullwood, M.J. Roles, Functions, and Mechanisms of Long Non-coding RNAs in Cancer. Genom. Proteom. Bioinforma. 2016, 14, 42–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fehlmann, T.; Backes, C.; Pirritano, M.; Laufer, T.; Galata, V.; Kern, F.; Kahraman, M.; Gasparoni, G.; Ludwig, N.; Lenhof, H.-P.; et al. The sncRNA Zoo: A repository for circulating small noncoding RNAs in animals. Nucleic Acids Res. 2019, 47, 4431–4441. [Google Scholar] [CrossRef]
- Lu, J.; Shen, Y.; Wu, Q.; Kumar, S.; He, B.; Shi, S.; Carthew, R.W.; Wang, S.M.; Wu, C.-I. The birth and death of microRNA genes in Drosophila. Nat. Genet. 2008, 40, 351–355. [Google Scholar] [CrossRef]
- Axtell, M.J. Classification and Comparison of Small RNAs from Plants. Annu. Rev. Plant Biol. 2013, 64, 137–159. [Google Scholar] [CrossRef] [Green Version]
- Mello, C.C.; Conte, D. Revealing the world of RNA interference. Nat. Cell Biol. 2004, 431, 338–342. [Google Scholar] [CrossRef]
- Biswas, S.; Chakrabarti, S. Increased Extracellular Matrix Protein Production in Chronic Diabetic Complications: Implications of Non-Coding RNAs. Non-Coding RNA 2019, 5, 30. [Google Scholar] [CrossRef] [Green Version]
- Gutschner, T.; Diederichs, S. The hallmarks of cancer. RNA Biol. 2012, 9, 703–719. [Google Scholar] [CrossRef] [Green Version]
- Mikulík, K. Structure and functional properties of prokaryotic small noncoding RNAs. Folia Microbiol. 2003, 48, 443–468. [Google Scholar] [CrossRef]
- Hindley, J. Fractionation of 32P-labelled ribonucleic acids on polyacrylamide gels and their characterization by fingerprinting. J. Mol. Biol. 1967, 30, 125–136. [Google Scholar] [CrossRef]
- Nitzan, M.; Rehani, R.; Margalit, H. Integration of Bacterial Small RNAs in Regulatory Networks. Annu. Rev. Biophys. 2017, 46, 131–148. [Google Scholar] [CrossRef] [PubMed]
- Pita, T.; Feliciano, J.R.; Leitão, J.H. Small Noncoding Regulatory RNAs from Pseudomonas aeruginosa and Burkholderia cepacia Complex. Int. J. Mol. Sci. 2018, 19, 3759. [Google Scholar] [CrossRef] [Green Version]
- Papenfort, K.; Pfeiffer, V.; Mika, F.; Lucchini, S.; Hinton, J.C.D.; Vogel, J. ΣE-dependent small RNAs of Salmonella respond to membrane stress by accelerating global omp mRNA decay. Mol. Microbiol. 2006, 62, 1674–1688. [Google Scholar] [CrossRef] [Green Version]
- Klein, G.; Raina, S. Small regulatory bacterial RNAs regulating the envelope stress response. Biochem. Soc. Trans. 2017, 45, 417–425. [Google Scholar] [CrossRef] [Green Version]
- Michaux, C.; Verneuil, N.; Hartke, A.; Giard, J.-C. Physiological roles of small RNA molecules. Microbiology 2014, 160, 1007–1019. [Google Scholar] [CrossRef] [PubMed]
- Brantl, S. Chapter 4 Small Regulatory RNAs (SRNAs): Key Players in Prokaryotic Metabolism, Stress Response, and Virulence. In Regulatory RNAs; Mallick, B., Ed.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 73–108. [Google Scholar]
- Park, H.; Yoon, Y.; Suk, S.; Lee, J.Y.; Lee, Y. Effects of different target sites on antisense RNA-mediated regulation of gene expression. BMB Rep. 2014, 47, 619–624. [Google Scholar] [CrossRef] [PubMed]
- Figueroa-Bossi, N.; Valentini, M.; Malleret, L.; Bossi, L. Caught at its own game: Regulatory small RNA inactivated by an inducible transcript mimicking its target. Genes Dev. 2009, 23, 2004–2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azam, M.S.; Vanderpool, C.K. Talk among yourselves: RNA sponges mediate cross talk between functionally related messenger RNA s. EMBO J. 2015, 34, 1436–1438. [Google Scholar] [CrossRef] [Green Version]
- Malabirade, A.; Morgado-Brajones, J.; Trépout, S.; Wien, F.; Marquez, I.; Seguin, J.; Marco, S.; Velez, M.; Arluison, V. Membrane association of the bacterial riboregulator Hfq and functional perspectives. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef]
- Brennan, R.G.; Link, T.M. Hfq structure, function and ligand binding. Curr. Opin. Microbiol. 2007, 10, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Feliciano, J.R.; Grilo, A.M.; I Guerreiro, S.; Sousa, S.A.; Leitão, J.H. Hfq: A multifaceted RNA chaperone involved in virulence. Futur. Microbiol. 2016, 11, 137–151. [Google Scholar] [CrossRef] [PubMed]
- Feliciano, J.R.; Seixas, A.M.; Pita, T.; Leitão, J.H. Comparative Genomics and Evolutionary Analysis of RNA-Binding Proteins of Burkholderia cenocepacia J2315 and Other Members of the B. cepacia Complex. Genes 2020, 11, 231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smirnov, A.; Foerstner, K.U.; Holmqvist, E.; Otto, A.; Günster, R.; Becher, D.; Reinhardt, R.; Vogel, J. Grad-seq guides the discovery of ProQ as a major small RNA-binding protein. Proc. Natl. Acad. Sci. USA 2016, 113, 11591–11596. [Google Scholar] [CrossRef] [Green Version]
- Cajal, S.R.y.; Segura, M.F.; Hümmer, S. Interplay between ncRNAs and Cellular Communication: A Proposal for Understanding Cell-Specific Signaling Pathways. Front. Genet. 2019, 10, 281. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Li, X. Recent advances in extracellular vesicles enriched with non-coding RNAs related to cancers. Genes Dis. 2018, 5, 36–42. [Google Scholar] [CrossRef]
- Turchinovich, A.; Weiz, L.; Burwinkel, B. Extracellular miRNAs: The mystery of their origin and function. Trends Biochem. Sci. 2012, 37, 460–465. [Google Scholar] [CrossRef]
- Filipowicz, W.; Bhattacharyya, S.N.; Sonenberg, N. Mechanisms of post-transcriptional regulation by microRNAs: Are the answers in sight? Nat. Rev. Genet. 2008, 9, 102–114. [Google Scholar] [CrossRef]
- Arroyo, J.D.; Chevillet, J.R.; Kroh, E.M.; Ruf, I.K.; Pritchard, C.C.; Gibson, D.F.; Mitchell, P.S.; Bennett, C.F.; Pogosova-Agadjanyan, E.L.; Stirewalt, D.L.; et al. Argonaute2 complexes carry a population of circulating microRNAs independent of vesicles in human plasma. Proc. Natl. Acad. Sci. USA 2011, 108, 5003–5008. [Google Scholar] [CrossRef] [Green Version]
- Vickers, K.C.; Palmisano, B.T.; Shoucri, B.M.; Shamburek, R.D.; Remaley, A.T. MicroRNAs are transported in plasma and delivered to recipient cells by high-density lipoproteins. Nat. Cell Biol. 2011, 13, 423–433. [Google Scholar] [CrossRef] [Green Version]
- Niculescu, L.S.; Simionescu, N.; Sanda, G.M.; Carnuta, M.G.; Stancu, C.S.; Popescu, A.C.; Popescu, M.R.; Vlad, A.; Dimulescu, D.R.; Simionescu, M.; et al. MiR-486 and miR-92a Identified in Circulating HDL Discriminate between Stable and Vulnerable Coronary Artery Disease Patients. PLoS ONE 2015, 10, e0140958. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; El Andaloussi, S.; Wood, M.J.A. Exosomes and microvesicles: Extracellular vesicles for genetic information transfer and gene therapy. Hum. Mol. Genet. 2012, 21, R125–R134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mause, S.F.; Weber, C. Microparticles. Circ. Res. 2010, 107, 1047–1057. [Google Scholar] [CrossRef] [PubMed]
- Sohel, M.H. Extracellular/Circulating MicroRNAs: Release Mechanisms, Functions and Challenges. Achiev. Life Sci. 2016, 10, 175–186. [Google Scholar] [CrossRef] [Green Version]
- Turiák, L.; Misják, P.; Szabó, T.G.; Aradi, B.; Pálóczi, K.; Ozohanics, O.; Drahos, L.; Kittel, A.; Falus, A.; Buzás, E.I.; et al. Proteomic characterization of thymocyte-derived microvesicles and apoptotic bodies in BALB/c mice. J. Proteom. 2011, 74, 2025–2033. [Google Scholar] [CrossRef] [PubMed]
- Hergenreider, E.; Heydt, S.; Tréguer, K.; Boettger, T.; Horrevoets, A.J.G.; Zeiher, A.M.; Scheffer, M.P.; Frangakis, A.S.; Yin, X.; Mayr, M.; et al. Atheroprotective communication between endothelial cells and smooth muscle cells through miRNAs. Nat. Cell Biol. 2012, 14, 249–256. [Google Scholar] [CrossRef]
- Kourembanas, S. Exosomes: Vehicles of Intercellular Signaling, Biomarkers, and Vectors of Cell Therapy. Annu. Rev. Physiol. 2015, 77, 13–27. [Google Scholar] [CrossRef] [Green Version]
- Villarroya-beltri, C.; Gutie, C.; Martin-cofreces, N.; Martinez-herrera, D.J.; Pascual-montano, A. Sumoylated hnRNPA2B1 controls the sorting of miRNAs into exosomes through binding to specific motifs. Nat. Commun. 2013, 4, 2980. [Google Scholar] [CrossRef] [Green Version]
- McKenzie, A.J.; Hoshino, D.; Hong, N.H.; Cha, D.J.; Franklin, J.L.; Coffey, R.J.; Patton, J.G.; Weaver, A.M. KRAS-MEK Signaling Controls Ago2 Sorting into Exosomes. Cell Rep. 2016, 15, 978–987. [Google Scholar] [CrossRef] [Green Version]
- Shurtleff, M.J.; Temoche-Diaz, M.M.; Karfilis, K.V.; Ri, S.; Schekman, R. Y-box protein 1 is required to sort microRNAs into exosomes in cells and in a cell-free reaction. eLife 2016, 5, e19276. [Google Scholar] [CrossRef]
- Santangelo, L.; Giurato, G.; Cicchini, C.; Alonzi, T.; Weisz, A.; Tripodi, M.; Santangelo, L.; Giurato, G.; Cicchini, C.; Montaldo, C.; et al. The RNA-Binding Protein SYNCRIP Is a Component of the Hepatocyte Exosomal Machinery Controlling MicroRNA Sorting. Cell Rep. 2016, 17, 799–808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, J.; Lv, L.; Wan, Y.; Cao, Y.; Li, G.; Lin, H.; Zhou, R. Vps4A functions as a tumor suppressor by regulating the secretion and uptake of exosomal microRNAs in human hepatoma cells. Hepatology 2015, 61, 1284–1294. [Google Scholar] [CrossRef] [PubMed]
- Squadrito, M.L.; Maderna, C.; Gilfillan, G.D.; Lyle, R.; Baer, C.; Ibberson, M.; Palma, M.D. Endogenous RNAs Modulate MicroRNA Sorting to Exosomes and Transfer to Acceptor Cells. Cell Rep. 2014, 8, 1432–1446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toyofuku, M.; Nomura, N.; Eberl, L. Types and origins of bacterial membrane vesicles. Nat. Rev. Microbiol. 2019, 17, 13–24. [Google Scholar] [CrossRef]
- Toyofuku, M.; Cárcamo-Oyarce, G.; Yamamoto, T.; Eisenstein, F.; Hsiao, C.-C.; Kurosawa, M.; Gademann, K.; Pilhofer, M.; Nomura, N.; Eberl, L. Prophage-triggered membrane vesicle formation through peptidoglycan damage in Bacillus subtilis. Nat. Commun. 2017, 8, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Eulalio, A.; Schulte, L.; Vogel, J.; Eulalio, A.; Schulte, L.N.; Vogel, J. The mammalian microRNA response to bacterial infections. RNA Biol. 2012, 9, 742–750. [Google Scholar] [CrossRef]
- Kumar, M.; Kundu, M.; Basu, J. The Role of MicroRNAs in Bacterial Infections. In AGO-Driven Non-Coding RNAs; Mallick, B., Ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 57–73. [Google Scholar]
- Lai, F.W.; Lichty, B.D.; Bowdish, D.M.E. Microvesicles: Ubiquitous contributors to infection and immunity. J. Leukoc. Biol. 2014, 97, 237–245. [Google Scholar] [CrossRef]
- Fleming, A.; Sampey, G.; Chung, M.-C.; Bailey, C.; Van Hoek, M.L.; Kashanchi, F.; Hakami, R.M. The carrying pigeons of the cell: Exosomes and their role in infectious diseases caused by human pathogens. Pathog. Dis. 2014, 71, 109–120. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Jiang, X.; Bao, J.; Wang, Y.; Liu, H.; Tang, L. Exosomes in Pathogen Infections: A Bridge to Deliver Molecules and Link Functions. Front. Immunol. 2018, 9, 90. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Q.; Chen, H.; Yang, Y.; Fu, Y.; Yi, Z. miR-18a promotes Mycobacterial survival in macrophages via inhibiting autophagy by down-regulation of ATM. J. Cell. Mol. Med. 2019, 24, 2004–2012. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Deng, Z.; Wang, Z.; Wu, J.; Gu, T.; Jiang, Y.; Li, G. MicroRNA-155 in exosomes secreted from Helicobacter pylori infection macrophages immunomodulates inflammatory response. Am. J. Transl. Res. 2016, 8, 3700–3709. [Google Scholar] [PubMed]
- Alexander, M.; Hu, R.; Runtsch, M.C.; Kagele, D.A.; Mosbruger, T.L.; Tolmachova, T.; Seabra, M.C.; Round, J.L.; Ward, D.M.; O’Connell, R.M. Exosome-delivered microRNAs modulate the inflammatory response to endotoxin. Nat. Commun. 2015, 6, 7321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulp, A.; Kuehn, M.J. Biological Functions and Biogenesis of Secreted Bacterial Outer Membrane Vesicles. Annu. Rev. Microbiol. 2010, 64, 163–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tashiro, Y.; Ichikawa, S.; Shimizu, M.; Toyofuku, M.; Takaya, N.; Nakajima-Kambe, T.; Uchiyama, H.; Nomura, N. Variation of Physiochemical Properties and Cell Association Activity of Membrane Vesicles with Growth Phase in Pseudomonas aeruginosa. Appl. Environ. Microbiol. 2010, 76, 3732–3739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, J.; I Kim, S.; Ryu, S.; Yoon, H. Identification and Characterization of Outer Membrane Vesicle-Associated Proteins in Salmonella enterica Serovar Typhimurium. Infect. Immun. 2014, 82, 4001–4010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, C.-W.; Park, E.C.; Yun, S.H.; Lee, S.-Y.; Lee, Y.G.; Hong, Y.; Park, K.R.; Kim, S.-H.; Kim, G.-H.; Kim, S.I. Proteomic Characterization of the Outer Membrane Vesicle of Pseudomonas putida KT2440. J. Proteome Res. 2014, 13, 4298–4309. [Google Scholar] [CrossRef] [PubMed]
- Mashburn, L.M.; Whiteley, M. Membrane vesicles traffic signals and facilitate group activities in a prokaryote. Nat. Cell Biol. 2005, 437, 422–425. [Google Scholar] [CrossRef]
- Jan, A.T. Outer Membrane Vesicles (OMVs) of Gram-negative Bacteria: A Perspective Update. Front. Microbiol. 2017, 8, 1053. [Google Scholar] [CrossRef]
- Lee, E.-Y.; Choi, D.-Y.; Kim, D.-K.; Kim, J.-W.; Park, J.O.; Kim, S.; Kim, S.-H.; Desiderio, D.M.; Kim, Y.-K.; Kim, K.P.; et al. Gram-positive bacteria produce membrane vesicles: Proteomics-based characterization of Staphylococcus aureus-derived membrane vesicles. Proteomics 2009, 9, 5425–5436. [Google Scholar] [CrossRef]
- Brown, L.; Wolf, J.M.; Prados-Rosales, R.; Casadevall, A. Through the wall: Extracellular vesicles in Gram-positive bacteria, mycobacteria and fungi. Nat. Rev. Genet. 2015, 13, 620–630. [Google Scholar] [CrossRef] [Green Version]
- Bos, M.P.; Robert, V.; Tommassen, J. Biogenesis of the Gram-Negative Bacterial Outer Membrane. Annu. Rev. Microbiol. 2007, 61, 191–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulkarni, H.M.; Jagannadham, M.V. Biogenesis and multifaceted roles of outer membrane vesicles from Gram-negative bacteria. Microbiology 2014, 160, 2109–2121. [Google Scholar] [CrossRef] [Green Version]
- Qing, G.; Gong, N.; Chen, X.; Chen, J.; Zhang, H.; Wang, Y.; Wang, R.; Zhang, S. Natural and engineered bacterial outer membrane vesicles. Biophys. Rep. 2019, 5, 184–198. [Google Scholar] [CrossRef] [Green Version]
- Schertzer, J.W.; Whiteley, M. A Bilayer-Couple Model of Bacterial Outer Membrane Vesicle. MBio 2012, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roier, S.; Zingl, F.G.; Cakar, F.; Durakovic, S.; Kohl, P.; Eichmann, T.O.; Klug, L.; Gadermaier, B.; Weinzerl, K.; Prassl, R.; et al. A novel mechanism for the biogenesis of outer membrane vesicles in Gram-negative bacteria. Nat. Commun. 2016, 7, 10515. [Google Scholar] [CrossRef] [Green Version]
- Nevermann, J.; Silva, A.; Otero, C.; Oyarzún, D.P.; Barrera, B.; Gil, F.; Calderón, I.L.; Fuentes, J.A. Identification of Genes Involved in Biogenesis of Outer Membrane Vesicles (OMVs) in Salmonella enterica Serovar Typhi. Front. Microbiol. 2019, 10, 104. [Google Scholar] [CrossRef]
- Malabirade, A.; Habier, J.; Heintz-Buschart, A.; May, P.; Godet, J.; Halder, R.; Etheridge, A.; Galas, D.; Wilmes, P.; Fritz, J.V. The RNA Complement of Outer Membrane Vesicles from Salmonella enterica Serovar Typhimurium Under Distinct Culture Conditions. Front. Microbiol. 2018, 9, 2015. [Google Scholar] [CrossRef]
- Tan, K.; Li, R.; Huang, X.; Liu, Q. Outer Membrane Vesicles: Current Status and Future Direction of These Novel Vaccine Adjuvants. Front. Microbiol. 2018, 9, 783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kieft, J.S.; Rabe, J.L.; Chapman, E.G. RNA: Conservation, Folding, and Host Adaptation. RNA Biol. 2015, 12, 1169–1177. [Google Scholar] [CrossRef] [Green Version]
- Benmoussa, A.; Lee, C.H.C.; Laffont, B.; Savard, P.; Laugier, J.; Boilard, E.; Gilbert, C.; Fliss, I.; Provost, P. Commercial Dairy Cow Milk microRNAs Resist Digestion under Simulated Gastrointestinal Tract Conditions. J. Nutr. 2016, 146, 2206–2215. [Google Scholar] [CrossRef]
- Dauros-Singorenko, P.; Blenkiron, C.; Phillips, A.; Swift, S. The functional RNA cargo of bacterial membrane vesicles. FEMS Microbiol. Lett. 2018, 365, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koeppen, K.; Hampton, T.H.; Jarek, M.; Scharfe, M.; Gerber, S.A.; Mielcarz, D.W.; Demers, E.G.; Dolben, E.L.; Hammond, J.H.; Hogan, D.A.; et al. A Novel Mechanism of Host-Pathogen Interaction through sRNA in Bacterial Outer Membrane Vesicles. PLOS Pathog. 2016, 12, e1005672. [Google Scholar] [CrossRef]
- Kang, S.; Choi, J.-W.; Lee, Y.; Hong, S.-H.; Lee, H.-J. Identification of microRNA-Size, Small RNAs in Escherichia coli. Curr. Microbiol. 2013, 67, 609–613. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-J.; Hong, S.-H. Analysis of microRNA-size, small RNAs in Streptococcus mutans by deep sequencing. FEMS Microbiol. Lett. 2011, 326, 131–136. [Google Scholar] [CrossRef]
- Ghosal, A.; Upadhyaya, B.B.; Fritz, J.V.; Heintz-Buschart, A.; Desai, M.S.; Yusuf, D.; Huang, D.; Baumuratov, A.; Wang, K.; Galas, D.; et al. The extracellular RNA complement of Escherichia coli. Microbiology 2015, 4, 252–266. [Google Scholar] [CrossRef]
- Ghosal, A. Secreted bacterial RNA: An unexplored avenue. FEMS Microbiol. Lett. 2018, 365, 2017–2019. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhang, Y.; Song, Z.; Li, R.; Ruan, H.; Liu, Q.; Huang, X. sncRNAs packaged by Helicobacter pylori outer membrane vesicles attenuate IL-8 secretion in human cells. Int. J. Med. Microbiol. 2020, 310, 151356. [Google Scholar] [CrossRef]
- Pagliuso, A.; Tham, T.N.; Allemand, E.; Robertin, S.; Dupuy, B.; Bertrand, Q.; Bécavin, C.; Koutero, M.; Najburg, V.; Nahori, M.-A.; et al. An RNA-Binding Protein Secreted by a Bacterial Pathogen Modulates RIG-I Signaling. Cell Host Microbe 2019, 26, 823–835.e11. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.; Schorey, J.S. Extracellular vesicles deliver Mycobacterium RNA to promote host immunity and bacterial killing. EMBO Rep. 2019, 20, e46613. [Google Scholar] [CrossRef]
- Fritz, J.V.; Heintz-Buschart, A.; Ghosal, A.; Wampach, L.; Etheridge, A.; Galas, D.J.; Wilmes, P. Sources and Functions of Extracellular Small RNAs in Human Circulation. Annu. Rev. Nutr. 2016, 36, 301–336. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Li, H.; Yuan, Y.; Etheridge, A.; Zhou, Y.; Huang, D.; Wilmes, P.; Galas, D.J. The Complex Exogenous RNA Spectra in Human Plasma: An Interface with Human Gut Biota? PLoS ONE 2012, 7, e51009. [Google Scholar] [CrossRef] [PubMed]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Tvall, J.O.L.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorward, D.W.; Garon, C.F.; Judd, R.C. Export and intercellular transfer of DNA via membrane blebs of Neisseria gonorrhoeae. J. Bacteriol. 1989, 171, 2499–2505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heroven, A.K.; Sest, M.; Pisano, F.; Scheb-Wetzel, M.; Steinmann, R.; Böhme, K.; Klein, J.; Münch, R.; Schomburg, D.; Dersch, P. Crp Induces Switching of the CsrB and CsrC RNAs in Yersinia pseudotuberculosis and Links Nutritional Status to Virulence. Front. Cell. Infect. Microbiol. 2012, 2, 158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blenkiron, C.; Simonov, D.; Muthukaruppan, A.; Tsai, P.; Dauros, P.; Green, S.; Hong, J.; Print, C.G.; Swift, S.; Phillips, A.R. Uropathogenic Escherichia coli Releases Extracellular Vesicles That Are Associated with RNA. PLoS ONE 2016, 11, e0160440. [Google Scholar] [CrossRef]
- Diggle, S.P.; Whiteley, M. Microbe Profile: Pseudomonas aeruginosa: Opportunistic pathogen and lab rat. Microbiology 2020, 166, 30–33. [Google Scholar] [CrossRef]
- Turnbull, L.; Toyofuku, M.; Hynen, A.L.; Kurosawa, M.; Pessi, G.; Petty, N.K.; Osvath, S.R.; Cárcamo-Oyarce, G.; Gloag, E.S.; Shimoni, R.; et al. Explosive cell lysis as a mechanism for the biogenesis of bacterial membrane vesicles and biofilms. Nat. Commun. 2016, 7, 11220. [Google Scholar] [CrossRef] [Green Version]
- Sonnleitner, E.; González, N.; Sorger-Domenigg, T.; Heeb, S.; Richter, A.S.; Backofen, R.; Williams, P.; Hüttenhofer, A.; Haas, D.; Bläsi, U. The small RNA PhrS stimulates synthesis of the Pseudomonas aeruginosa quinolone signal. Mol. Microbiol. 2011, 80, 868–885. [Google Scholar] [CrossRef]
- Turner, L.; Bitto, N.J.; Steer, D.L.; Lo, C.; D’Costa, K.; Ramm, G.; Shambrook, M.; Hill, A.F.; Ferrero, R.L.; Kaparakis-Liaskos, M. Helicobacter pylori Outer Membrane Vesicle Size Determines Their Mechanisms of Host Cell Entry and Protein Content. Front. Immunol. 2018, 9, 1466. [Google Scholar] [CrossRef] [Green Version]
- Yonezawa, H.; Osaki, T.; Kurata, S.; Fukuda, M.; Kawakami, H.; Ochiai, K.; Hanawa, T.; Kamiya, S. Outer Membrane Vesicles of Helicobacter pylori TK1402 are Involved in Biofilm Formation. BMC Microbiol. 2009, 9, 197. [Google Scholar] [CrossRef] [Green Version]
- Jarzab, M.; Posselt, G.; Meisner-Kober, N.; Wessler, S. Helicobacter pylori-Derived Outer Membrane Vesicles (OMVs): Role in Bacterial Pathogenesis? Microorganisma 2020, 8, 1328. [Google Scholar] [CrossRef] [PubMed]
- Heras, A.D.L.; Cain, R.J.; Bielecka, M.K.; Vazquez-Boland, J. Regulation of Listeria virulence: PrfA master and commander. Curr. Opin. Microbiol. 2011, 14, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Vdovikova, S.; Luhr, M.; Szalai, P.; Skalman, L.N.; Francis, M.K.; Lundmark, R.; Engedal, N.; Johansson, J.; Wai, S.N. A Novel Role of Listeria monocytogenes Membrane Vesicles in Inhibition of Autophagy and Cell Death. Front. Cell. Infect. Microbiol. 2017, 7, 154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdullah, Z.; Schlee, M.; Roth, S.; Abu Mraheil, M.; Barchet, W.; Böttcher, J.; Hain, T.; Geiger, S.; Hayakawa, Y.; Fritz, J.H.; et al. RIG-I detects infection with live Listeria by sensing secreted bacterial nucleic acids. EMBO J. 2012, 31, 4153–4164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Connell, R.M.; Saha, S.K.; Vaidya, S.A.; Bruhn, K.W.; Miranda, G.A.; Zarnegar, B.; Perry, A.K.; Nguyen, B.O.; Lane, T.F.; Taniguchi, T.; et al. Type I Interferon Production Enhances Susceptibility to Listeria monocytogenes Infection. J. Exp. Med. 2004, 200, 437–445. [Google Scholar] [CrossRef] [Green Version]
- Frantz, R.; Teubner, L.; Schultze, T.; La Pietra, L.; Müller, C.; Gwozdzinski, K.; Pillich, H.; Hain, T.; Weber-Gerlach, M.; Panagiotidis, G.-D.; et al. The secRNome of Listeria monocytogenes Harbors Small Noncoding RNAs That Are Potent Inducers of Beta Interferon. mBio 2019, 10, e01223-19. [Google Scholar] [CrossRef] [Green Version]
- Gu, H.; Zhao, C.; Zhang, T.; Liang, H.; Wang, X.-M.; Pan, Y.; Chen, X.; Zhao, Q.; Li, D.; Liu, F.; et al. Salmonella produce microRNA-like RNA fragment Sal-1 in the infected cells to facilitate intracellular survival. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef]
- Kaplan, G.; Post, F.A.; Moreira, A.L.; Wainwright, H.; Kreiswirth, B.N.; Tanverdi, M.; Mathema, B.; Ramaswamy, S.V.; Walther, G.; Steyn, L.M.; et al. Mycobacterium tuberculosis Growth at theCavity Surface: A Microenvironment with FailedImmunity. Infect. Immun. 2003, 71, 7099–7108. [Google Scholar] [CrossRef] [Green Version]
- Furuse, Y.; Finethy, R.; Saka, H.A.; Xet-Mull, A.M.; Sisk, D.M.; Smith, K.L.J.; Lee, S.; Coers, J.; Valdivia, R.H.; Tobin, D.M.; et al. Search for MicroRNAs Expressed by Intracellular Bacterial Pathogens in Infected Mammalian Cells. PLoS ONE 2014, 9, e106434. [Google Scholar] [CrossRef]
- Obregón-Henao, A.; Duque-Correa, M.A.; Rojas, M.; García, L.F.; Brennan, P.J.; Ortiz, B.L.; Belisle, J.T. Stable Extracellular RNA Fragments of Mycobacterium tuberculosis Induce Early Apoptosis in Human Monocytes via a Caspase-8 Dependent Mechanism. PLoS ONE 2012, 7, e29970. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.-W.; Kim, S.-C.; Hong, S.-H.; Lee, H.-J. Secretable Small RNAs via Outer Membrane Vesicles in Periodontal Pathogens. J. Dent. Res. 2017, 96, 458–466. [Google Scholar] [CrossRef] [PubMed]
- Ho, M.-H.; Chen, C.-H.; Goodwin, J.S.; Wang, B.-Y.; Xie, H. Functional Advantages of Porphyromonas gingivalis Vesicles. PLoS ONE 2015, 10, e0123448. [Google Scholar] [CrossRef]
- Han, E.-C.; Choi, S.-Y.; Lee, Y.; Park, J.-W.; Hong, S.-H.; Lee, H.-J. Extracellular RNAs in periodontopathogenic outer membrane vesicles promote TNF-α production in human macrophages and cross the blood–brain barrier in mice. FASEB J. 2019, 33, 13412–13422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, J.-W.; Kwon, T.-Y.; Hong, S.-H.; Lee, H.-J. Isolation and Characterization of a microRNA-size Secretable Small RNA in Streptococcus sanguinis. Cell Biophys. 2016, 76, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Sjöström, A.E.; Sandblad, L.; Uhlin, B.E.; Wai, S.N. Membrane vesicle-mediated release of bacterial RNA. Sci. Rep. 2015, 5, 15329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moriano-Gutierrez, S.; Bongrand, C.; Essock-Burns, T.; Wu, L.; McFall-Ngai, M.; Ruby, E. The noncoding small RNA SsrA is released by Vibrio fischeri and modulates critical host responses. PLoS Biol. 2020, 18, e3000934. [Google Scholar] [CrossRef] [PubMed]
- Singorenko, P.D.; Chang, V.; Whitcombe, A.; Simonov, D.; Hong, J.; Phillips, A.; Swift, S.; Blenkiron, C. Isolation of membrane vesicles from prokaryotes: A technical and biological comparison reveals heterogeneity. J. Extracell. Vesicles 2017, 6, 1324731. [Google Scholar] [CrossRef]
- Resch, U.; Tsatsaronis, J.A.; Le Rhun, A.; Stuebiger, G.; Rohde, M.; Kasvandik, S.; Holzmeister, S.; Tinnefeld, P.; Wai, S.N.; Charpentier, E. A Two-Component Regulatory System Impacts Extracellular Membrane-Derived Vesicle Production in Group A Streptococcus. mBio 2016, 7, e00207-16. [Google Scholar] [CrossRef] [Green Version]
- Malge, A.; Ghai, V.; Reddy, P.J.; Baxter, D.; Kim, T.-K.; Moritz, R.L.; Wang, K. mRNA transcript distribution bias between Borrelia burgdorferi bacteria and their outer membrane vesicles. FEMS Microbiol. Lett. 2018, 365, 1–9. [Google Scholar] [CrossRef]
- Domenech, M.; Pedrero-Vega, E.; Prieto, A.; García, E. Evidence of the presence of nucleic acids and β-glucan in the matrix of non-typeable Haemophilus influenzae in vitro biofilms. Sci. Rep. 2016, 6, 36424. [Google Scholar] [CrossRef] [Green Version]
- Biller, S.J.; Schubotz, F.; Roggensack, S.E.; Thompson, A.W.; Summons, R.E.; Chisholm, S.W. Bacterial Vesicles in Marine Ecosystems. Science 2014, 343, 183–186. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, B.V.; Kuehn, M.J. Staphylococcus aureus secretes immunomodulatory RNA and DNA via membrane vesicles. Sci. Rep. 2020, 10, 1–22. [Google Scholar] [CrossRef]
- Anfossi, S.; Babayan, A.; Pantel, K.; Calin, G.A. Clinical utility of circulating non-coding RNAs—an update. Nat. Rev. Clin. Oncol. 2018, 15, 541–563. [Google Scholar] [CrossRef] [PubMed]
- Correia, C.N.; Nalpas, N.C.; McLoughlin, K.E.; Browne, J.A.; Gordon, S.V.; MacHugh, D.E.; Shaughnessy, R.G. Circulating microRNAs as Potential Biomarkers of Infectious Disease. Front. Immunol. 2017, 8, 118. [Google Scholar] [CrossRef] [Green Version]
- Sarshar, M.; Scribano, D.; Ambrosi, C.; Palamara, A.T.; Masotti, A. Fecal microRNAs as Innovative Biomarkers of Intestinal Diseases and Effective Players in Host-Microbiome Interactions. Cancers 2020, 12, 2174. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Aswath, K.; Schroeder, S.G.; Lippolis, J.D.; Reinhardt, T.A.; Sonstegard, T.S. MicroRNA expression profiles of bovine milk exosomes in response to Staphylococcus aureus infection. BMC Genom. 2015, 16, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Galvanin, A.; Dostert, G.; Ayadi, L.; Marchand, V.; Velot, E.; Motorin, Y. Diversity and heterogeneity of extracellular RNA in human plasma. Biochimie 2019, 164, 22–36. [Google Scholar] [CrossRef]
- Beatty, M.; Guduric-Fuchs, J.; Brown, E.; Bridgett, S.; Chakravarthy, U.; Hogg, R.E.; Simpson, D. Small RNAs from plants, bacteria and fungi within the order Hypocreales are ubiquitous in human plasma. BMC Genom. 2014, 15, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Semenov, D.V.; Baryakin, D.N.; Brenner, E.V.; Kurilshikov, A.; Vasiliev, G.V.; Bryzgalov, L.A.; Chikova, E.D.; Filippova, J.A.; Kuligina, E.V.; Richter, V.A. Unbiased approach to profile the variety of small non-coding RNA of human blood plasma with massively parallel sequencing technology. Expert Opin. Biol. Ther. 2012, 12, 1–9. [Google Scholar] [CrossRef]
- Setten, R.L.; Rossi, J.J.; Han, S.-P. The current state and future directions of RNAi-based therapeutics. Nat. Rev. Drug Discov. 2019, 18, 421–446. [Google Scholar] [CrossRef]
- Gilligan, K.E.; Dwyer, R.M. Extracellular Vesicles for Cancer Therapy: Impact of Host Immune Response. Cells 2020, 9, 224. [Google Scholar] [CrossRef] [Green Version]
- Hong, W.; Zeng, J.; Xie, J. Antibiotic drugs targeting bacterial RNAs. Acta Pharm. Sin. B 2014, 4, 258–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheah, H.-L.; Raabe, C.A.; Lee, L.-P.; Rozhdestvensky, T.S.; Citartan, M.; Ahmed, S.A.; Tang, T.H. Bacterial regulatory RNAs: Complexity, function, and putative drug targeting. Crit. Rev. Biochem. Mol. Biol. 2018, 53, 335–355. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Bacterial Species | Sequenced Biological Samples | seRNAs Features | Host Cell | Remarks | References |
|---|---|---|---|---|---|
| Aggregatibacter actinomycetemcomitans (ATCC 33384) | Outer OMV-associated secreted vs. cell small RNAs. | ncRNAs sizing 15 to 28 nt, some derived from degraded products, ribosomal RNA (rRNA) and transfer RNA (tRNA) | Jurkat T-cells Human macrophage-like cells (U937) | msRNA A.A_20050 reduce anti-inflammatory cytokines Interleukin (IL)–5, IL-13, and IL-15 in vitro. Cytoplasmic delivery and activity of microbial EV-derived small seRNAs in macrophages. Enhanced TNF-α production via TLR-8 and NF-κB signalling pathways. | [103,105] |
| Borrelia burgdorferi B31 | Outer OMV-associated secreted vs. cell RNA. | Reduced content of rRNA. The majority of small ncRNAs found on OMVs is derived from chromosomes. | N/A | Most of small ncRNAs are enriched in OMVs. Enrichment on plasmid-encoded transcripts in the OMVs, suggesting an involvement in genomic rearrangement | [111] |
| Chlamydia trachomatis LGV-L2 434/Bu | Secreted small RNAs inside host cells vs. cell small RNAs. | ncRNA of 15 to 43 nt size. Among the ten most abundant bacterial small RNAs 9 were originated of ORF regions and just one from non-coding regions. | HeLa CCL-2 cells | [101] | |
| Escherichia coli 536 | Outer OMV-associated secreted vs. cell RNA. | rRNA, tRNAs, other small RNAs. Undegraded mRNAs. Abundant small size RNA (15–50 nt) | 5637 bladder epithelial cells | 1% of MV RNA cargo delivered into cultured cells, including sRNA csrC | [87] |
| Escherichia coli K-12 substrain MG1655 | Outer free and OMV-associated secreted vs. cell RNA (<200 nt) | Selective exportation of 15 to 40 nt long cleavage products of tRNAs, 4.5S RNA, 6S RNA and tmRNA. | N/A | [77] | |
| Helicobacter pylori J99 | Outer OMV-associated secreted vs. cell RNA. | 59 ncRNAs present in OMVs. 45 ncRNAs enriched in OMVs. | Human gastric adenocarcinoma cells | seRNAs are delivered to host cells. seRNAs sR-2509025 and sR-989262 reduce interleukin 8 (IL-8) secretion | [79] |
| Legionella pneumophila CR39 | Secreted small RNAs inside host cells vs. cell small RNAs. | ncRNA of 15 to 43 nt size. Among the ten most abundant bacterial small RNAs 7 were originated from tRNA and 3 from ORF regions. | Murine macrophage cell line RAW264.7 | [101] | |
| Listeria monocytogenes EGD-e | Outer free and OMV-associated secreted vs. cell RNA. | seRNAs are mainly of short size (<200 nt), especially in OMVs | BMDM, P388D1 macrophages, and HEK293 cells | seRNAs highly induce IFN-β. ncRNA rli32-induced IFN- β in a RIG-I dependent way, leading to an inhibition of influenza virus replication. rli32 are involved in L. monocytogenes grow ability inside host cells. The RBP Zea bind seRNAs and RIG-I. | [80,98] |
| Mycobacterium marinum M | Secreted small RNAs inside host cells vs. cell small RNAs. | ncRNA of 15 to 43 nt size. Among the ten most abundant bacterial small RNAs 9 were originated of ORF regions and just one from non-coding regions. | Murine macrophage cell line RAW264.7 | The small RNA MM-H presented features of a miRNA was able to repress the host mRNA-target MM-H-specific RLuc reporter. | [101] |
| Mycobacterium smegmatis | Secreted small RNAs inside host cells vs. cell small RNAs. | ncRNA of 15 to 43 nt size. The ten most abundant bacterial small RNAs were originated from coding regions. | Murine macrophage cell line RAW264.7 | [101,109] | |
| Mycobacterium tuberculosis | Secreted small RNAs inside host cells vs. cell small RNAs. | ncRNA of 15 to 43 nt size. Among the ten most abundant bacterial small RNAs 9 were originated of ORF regions and just one from non-coding regions. | Murine macrophage cell line RAW264.7 | seRNA compromise human monocytes’ ability to control infection. seRNA secretion through sec2A secretory system. EVs released from M. tuberculosis infected macrophages enhance host defences. | [81,101,102] |
| non-typeable Haemophilus influenzae | N/A | RNA present in biofilm matrix | N/A | Extracellular RNA appears to be required for the initial attachment of a biofilm | [112] |
| Porphyromonas gingivalis (ATCC 33277) | Outer OMV-associated secreted vs. cell small RNAs. | ncRNAs sizing 15 to 50 nt, some derived from degraded products, ribosomal RNA (rRNA) and transfer RNA (tRNA). mRNAs of virulence factors detected by qRT-PCR | Jurkat T-cells, human gingival fibroblasts, human oral keratinocytes and human umbilical vein endothelial cells | msRNA P.G_45033 reduce anti-inflammatory cytokines Interleukin (IL)–5, IL-13, and IL-15 in vitro. OMVs can penetrate host cells. Vesicle-mediated RNA transference between P. gingivalis strains. | [103,104] |
| Pseudomonas aeruginosa PA14 | Outer OMV-associated secreted vs. cell RNA. | 64 ncRNAs present in OMVs. 52 ncRNAs enriched in OMVs. | Human bronchial epithelial (HBE) cells | sRNA52320 reaches HBE cells, reduces IL-8 secretion by cultured HBE cells and attenuate KC cytokine secretion and neutrophil infiltration in mouse lung | [74] |
| Pseudomonas aeruginosa PAO1 | Outer MV-associated secreted vs. cell RNA. | Intact 16S and 23S rRNAs. mRNAs related to SOS response (oxidative stress) enriched in cell lysis derived MVs | N/A | Reduced small RNA PA3305.1 (PhrS) content in MVs, suggesting an explosive cell lysis mediated MV production independent of PQS. | [89] |
| Salmonella enterica serovar Typhimurium LT2 | Outer OMV-associated secreted vs. cell RNA. | Mainly rRNA. mRNA and ncRNA were specifically enriched in OMVs from different growth conditions. | N/A | RNA is protected from degradation in OMVS. RNA cargo in OMVs is dependent of growth conditions and content abundance differs from the cell | [69] |
| Salmonella enteritidis SE2472 | Secreted RNA inside host cells vs. cell RNA. | RNA secretion inside host cells. ncRNA Sal-1 assume its mature form by host cell Argonaute 2 (AGO2) processing. Sal-1 increases infection rate in mice and the cell survival rate of Salmonella | [99] | ||
| Streptococcus pyogenes (GAS) | Outer OMV-associated secreted vs. cell RNA. | Mainly mRNAs, rRNAs and tRNAs | N/A | Different from most gram-negative bacteria, GAS membrane vesicles is enriched in intragenic RNA | [110] |
| Streptococcus sanguinis | N/A | msRNA detected in OMVs by qRT-PCR | N/A | msRNA are found in inside bacteria and are secreted through OMVs | [106] |
| Treponema denticola (ATCC 35405) | Outer OMV-associated secreted vs. cell small RNAs. | ncRNAs sizing 15 to 28 nt, some derived from degraded products, ribosomal RNA (rRNA) and transfer RNA (tRNA) | Jurkat T-cells | msRNA T.D_2161 reduce anti-inflammatory cytokines Interleukin (IL)–5, IL-13, and IL-15 in vitro | [103] |
| Vibrio cholerae A1552 | Outer OMV-associated secreted vs. cell RNA. | Abundant content of ncRNAs | N/A | RNA from the bacterial cells in association with OMVs are not growth phase dependent | [107] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pita, T.; Feliciano, J.R.; Leitão, J.H. Extracellular RNAs in Bacterial Infections: From Emerging Key Players on Host-Pathogen Interactions to Exploitable Biomarkers and Therapeutic Targets. Int. J. Mol. Sci. 2020, 21, 9634. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21249634
Pita T, Feliciano JR, Leitão JH. Extracellular RNAs in Bacterial Infections: From Emerging Key Players on Host-Pathogen Interactions to Exploitable Biomarkers and Therapeutic Targets. International Journal of Molecular Sciences. 2020; 21(24):9634. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21249634
Chicago/Turabian StylePita, Tiago, Joana R. Feliciano, and Jorge H. Leitão. 2020. "Extracellular RNAs in Bacterial Infections: From Emerging Key Players on Host-Pathogen Interactions to Exploitable Biomarkers and Therapeutic Targets" International Journal of Molecular Sciences 21, no. 24: 9634. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21249634