MicroRNAs and Sepsis-Induced Cardiac Dysfunction: A Systematic Review

 ,
,  and
and
Abstract
:1. Introduction
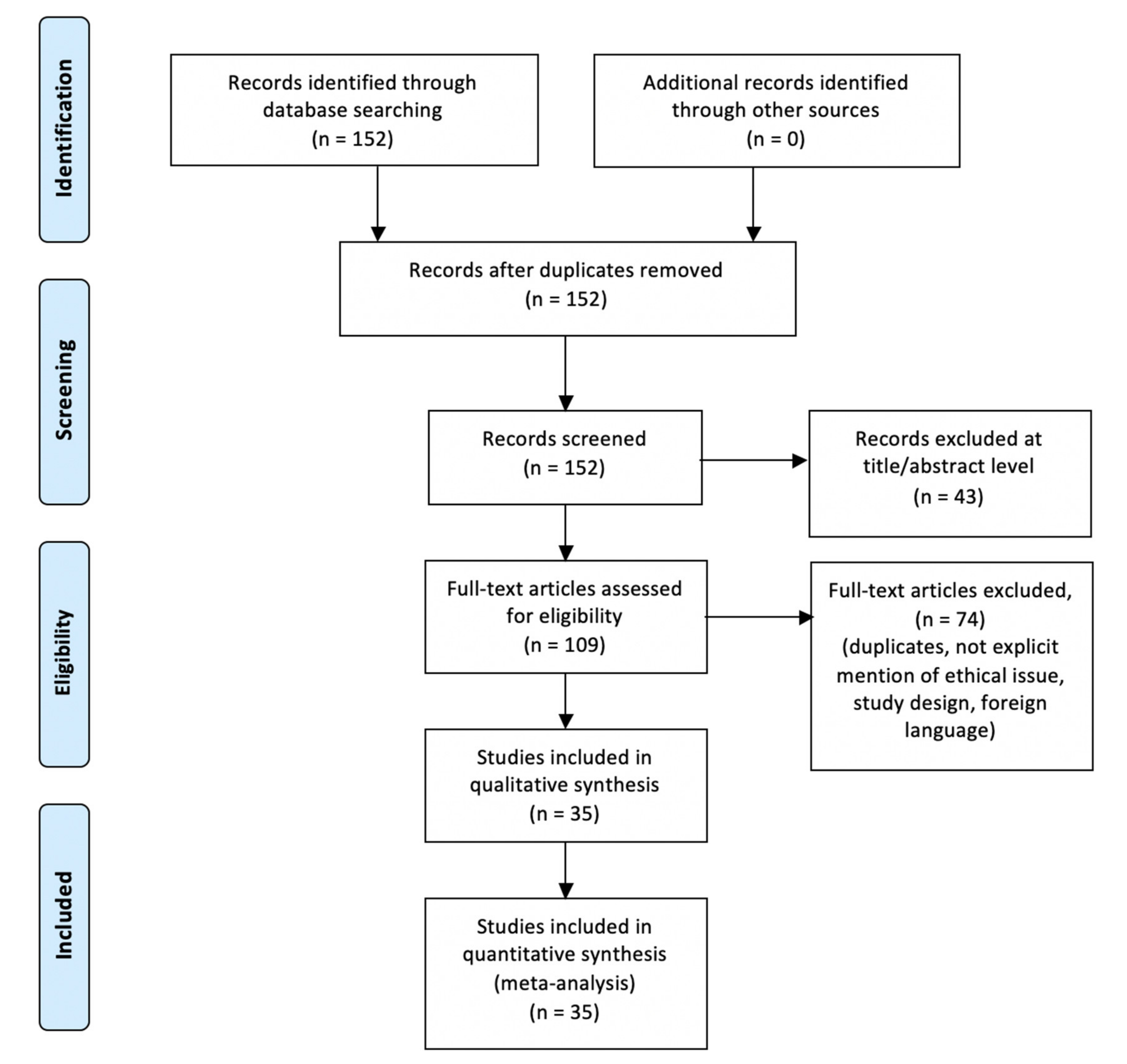
2. Materials and Methods
3. Results
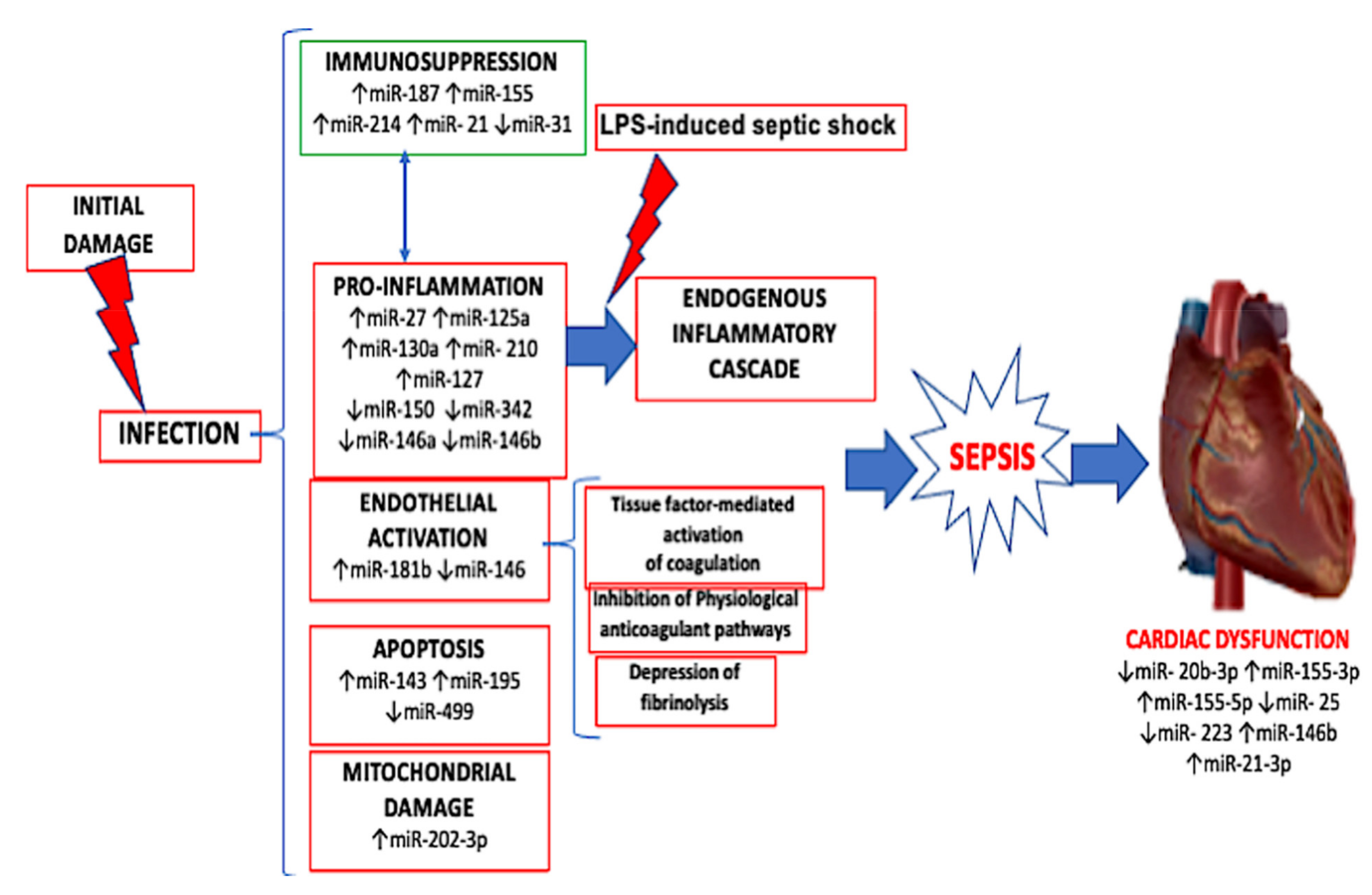
miRNA Expression in Sepsis-Related Cardiac Dysfunction
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Singer, M.; Deutschman, C.S.; Seymour, C.W.; Shankar-Hari, M.; Annane, D.; Bauer, M.; Bellomo, R.; Bernard, G.R.; Chiche, J.D.; Coopersmith, C.M.; et al. The Third International Consensus Definitions for Sepsis and Septic Shock (Sepsis-3). JAMA 2016, 315, 801–810. [Google Scholar] [CrossRef]
- Bone, R.C.; Sibbald, W.J.; Sprung, C.L. The ACCP-SCCM consensus conference on sepsis and organ failure. Chest 1992, 101, 1481–1483. [Google Scholar] [CrossRef] [Green Version]
- Angus, D.C.; van der Poll, T. Severe sepsis and septic shock. N. Engl. J. Med. 2013, 369, 840–851. [Google Scholar] [CrossRef]
- Iskander, K.N.; Osuchowski, M.F.; Stearns-Kurosawa, D.J.; Kurosawa, S.; Stepien, D.; Valentine, C.; Remick, D.G. Sepsis: Multiple abnormalities, heterogeneous responses, and evolving understanding. Physiol. Rev. 2013, 93, 1247–1288. [Google Scholar] [CrossRef] [Green Version]
- Angus, D.C.; Linde-Zwirble, W.T.; Lidicker, J.; Clermont, G.; Carcillo, J.; Pinsky, M.R. Epidemiology of severe sepsis in the United States: Analysis of incidence, outcome, and associated costs of care. Crit. Care Med. 2001, 29, 1303–1310. [Google Scholar] [CrossRef] [PubMed]
- Romero-Bermejo, F.J.; Ruiz-Bailen, M.; Gil-Cebrian, J.; Huertos-Ranchal, M.J. Sepsis-induced cardiomyopathy. Curr. Cardiol. Rev. 2011, 7, 163–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hochstadt, A.; Meroz, Y.; Landesberg, G. Myocardial dysfunction in severe sepsis and septic shock: More questions than answers? J. Cardiothorac. Vasc. Anesth. 2011, 25, 526–535. [Google Scholar] [CrossRef] [PubMed]
- Sanfilippo, F.; Corredor, C.; Fletcher, N.; Landesberg, G.; Benedetto, U.; Foex, P.; Cecconi, M. Diastolic dysfunction and mortality in septic patients: A systematic review and meta-analysis. Intensive Care Med. 2015, 41, 1004–1013. [Google Scholar] [CrossRef] [PubMed]
- Arefian, H.; Heublein, S.; Scherag, A.; Brunkhorst, F.M.; Younis, M.Z.; Moerer, O.; Fischer, D.; Hartmann, M. Hospital-related cost of sepsis: A systematic review. J. Infect. 2017, 74, 107–117. [Google Scholar] [CrossRef]
- Sergi, C.; Shen, F.; Lim, D.W.; Liu, W.; Zhang, M.; Chiu, B.; Anand, V.; Sun, Z. Cardiovascular dysfunction in sepsis at the dawn of emerging mediators. Biomed. Pharmacother. 2017, 95, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Piletič, K.; Kunej, T. MicroRNA epigenetic signatures in human disease. Arch. Toxicol. 2016, 90, 2405–2419. [Google Scholar] [CrossRef] [PubMed]
- Krol, J.; Loedige, I.; Filipowicz, W. The widespread regulation of microRNA biogenesis, function and decay. Nat. Rev. Genet. 2010, 11, 597–610. [Google Scholar] [CrossRef] [PubMed]
- Vasudevan, S.; Tong, Y.; Steitz, J.A. Switching from repression to activation: microRNAs can up-regulate translation. Science 2007, 318, 1931–1934. [Google Scholar] [CrossRef] [Green Version]
- Ardekani, A.M.; Naeini, M.M. The Role of MicroRNAs in Human Diseases. Avicenna J. Med. Biotechnol. 2010, 2, 161–179. [Google Scholar]
- Wendlandt, E.B.; Graff, J.W.; Gioannini, T.L.; McCaffrey, A.P.; Wilson, M.E. The role of microRNAs miR-200b and miR-200c in TLR4 signaling and NF-κB activation. Innate Immun. 2012, 18, 846–855. [Google Scholar] [CrossRef] [Green Version]
- Xu, G.; Zhang, Z.; Xing, Y.; Wei, J.; Ge, Z.; Liu, X.; Zhang, Y.; Huang, X. MicroRNA-149 negatively regulates TLR-triggered inflammatory response in macrophages by targeting MyD88. J. Cell Biochem. 2014, 115, 919–927. [Google Scholar] [CrossRef]
- Wei, J.; Huang, X.; Zhang, Z.; Jia, W.; Zhao, Z.; Zhang, Y.; Liu, X.; Xu, G. MyD88 as a target of microRNA-203 in regulation of lipopolysaccharide or Bacille Calmette-Guerin induced inflammatory response of macrophage RAW264.7 cells. Mol. Immunol. 2013, 55, 303–309. [Google Scholar] [CrossRef]
- Essandoh, K.; Fan, G.C. Role of extracellular and intracellular microRNAs in sepsis. Biochim. Biophys. Acta 2014, 1842, 2155–2162. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Ba, Y.; Ma, L.; Cai, X.; Yin, Y.; Wang, K.; Guo, J.; Zhang, Y.; Chen, J.; Guo, X.; et al. Characterization of microRNAs in serum: A novel class of biomarkers for diagnosis of cancer and other diseases. Cell Res. 2008, 18, 997–1006. [Google Scholar] [CrossRef] [Green Version]
- Vasilescu, C.; Rossi, S.; Shimizu, M.; Tudor, S.; Veronese, A.; Ferracin, M.; Nicoloso, M.S.; Barbarotto, E.; Popa, M.; Stanciulea, O.; et al. MicroRNA fingerprints identify miR-150 as a plasma prognostic marker in patients with sepsis. PLoS ONE 2009, 4, e7405. [Google Scholar] [CrossRef] [PubMed]
- Roderburg, C.; Luedde, M.; Vargas Cardenas, D.; Vucur, M.; Scholten, D.; Frey, N.; Koch, A.; Trautwein, C.; Tacke, F.; Luedde, T. Circulating microRNA-150 serum levels predict survival in patients with critical illness and sepsis. PLoS ONE 2013, 8, e54612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.F.; Yu, M.L.; Yu, G.; Bian, J.J.; Deng, X.M.; Wan, X.J.; Zhu, K.M. Serum miR-146a and miR-223 as potential new biomarkers for sepsis. Biochem. Biophys. Res. Commun. 2010, 394, 184–188. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Vilanova, D.; Atalar, K.; Delfour, O.; Edgeworth, J.; Ostermann, M.; Hernandez-Fuentes, M.; Razafimahatratra, S.; Michot, B.; Persing, D.H.; et al. Genome-wide sequencing of cellular microRNAs identifies a combinatorial expression signature diagnostic of sepsis. PLoS ONE 2013, 8, e75918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Meng, K.; jun Chen, W.; Feng, D.; Jia, Y.; Xie, L. Serum miR-574-5p: A prognostic predictor of sepsis patients. Shock 2012, 37, 263–267. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Icli, B.; Wara, A.K.; Belkin, N.; He, S.; Kobzik, L.; Hunninghake, G.M.; Vera, M.P.; Blackwell, T.S.; Baron, R.M.; et al. MicroRNA-181b regulates NF-κB-mediated vascular inflammation. J. Clin. Investig. 2012, 122, 1973–1990. [Google Scholar] [CrossRef]
- Li, Y.; Dalli, J.; Chiang, N.; Baron, R.M.; Quintana, C.; Serhan, C.N. Plasticity of leukocytic exudates in resolving acute inflammation is regulated by MicroRNA and proresolving mediators. Immunity 2013, 39, 885–898. [Google Scholar] [CrossRef] [Green Version]
- Tacke, F.; Roderburg, C.; Benz, F.; Cardenas, D.V.; Luedde, M.; Hippe, H.J.; Frey, N.; Vucur, M.; Gautheron, J.; Koch, A.; et al. Levels of circulating miR-133a are elevated in sepsis and predict mortality in critically ill patients. Crit. Care Med. 2014, 42, 1096–1104. [Google Scholar] [CrossRef] [Green Version]
- Benz, F.; Roy, S.; Trautwein, C.; Roderburg, C.; Luedde, T. Circulating MicroRNAs as Biomarkers for Sepsis. Int. J. Mol. Sci. 2016, 17, 78. [Google Scholar] [CrossRef] [Green Version]
- Shankar-Hari, M.; Lord, G.M. How might a diagnostic microRNA signature be used to speed up the diagnosis of sepsis? Expert Rev. Mol. Diagn. 2014, 14, 249–251. [Google Scholar] [CrossRef] [Green Version]
- Liberati, A.; Altman, D.G.; Tetzla, J.; Mulrow, C.; Gøtzsche, P.C.; Ioannidis, J.P.; Clarke, M.; Devereaux, P.J.; Kleijnen, J.; Moher, D. The PRISMA statement for reporting systematic reviews and meta-analyses of studies that evaluate health care interventions: Explanation and elaboration. Ann. Intern. Med. 2009, 62, e1–e34. [Google Scholar]
- Wang, X.; Huang, W.; Yang, Y.; Wang, Y.; Peng, T.; Chang, J.; Caldwell, C.C.; Zingarelli, B.; Fan, G.C. Loss of duplex miR-223 (5p and 3p) aggravates myocardial depression and mortality in polymicrobial sepsis. Biochim. Biophys. Acta 2014, 1842, 701–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Zhang, P.; Chen, W.; Feng, D.; Jia, Y.; Xie, L. Serum microRNA signatures identified by Solexa sequencing predict sepsis patients’ mortality: A prospective observational study. PLoS ONE 2012, 7, e38885. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Wang, Z.; Wang, H.; Ren, F.; Li, Y.; Zou, S.; Xu, J.; Xie, L. The protective role of miR-223 in sepsis-induced mortality. Sci. Rep. 2020, 10, 17691. [Google Scholar] [CrossRef]
- Fourdinier, O.; Schepers, E.; Metzinger-Le Meuth, V.; Glorieux, G.; Liabeuf, S.; Verbeke, F.; Vanholder, R.; Brigant, B.; Pletinck, A.; Diouf, M.; et al. Serum levels of miR-126 and miR-223 and outcomes in chronic kidney disease patients. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef] [Green Version]
- M’baya-Moutoula, E.; Louvet, L.; Molinié, R.; Guerrera, I.C.; Cerutti, C.; Fourdinier, O.; Nourry, V.; Gutierrez, L.; Morlière, P.; Mesnard, F.; et al. A multi-omics analysis of the regulatory changes induced by miR-223 in a monocyte/macrophage cell line. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 2664–2678. [Google Scholar] [CrossRef]
- Xue, W.L.; Bai, X.; Zhang, L. rhTNFR:Fc increases Nrf2 expression via miR-27a mediation to protect myocardium against sepsis injury. Biochem. Biophy.s Res. Commun. 2015, 464, 855–861. [Google Scholar] [CrossRef]
- Kensler, T.W.; Wakabayashi, N.; Biswal, S. Cell survival responses to environmental stresses via the Keap1-Nrf2-ARE pathway. Annu. Rev. Pharmacol. Toxicol. 2007, 47, 89–116. [Google Scholar] [CrossRef]
- Gao, M.; Wang, X.; Zhang, X.; Ha, T.; Ma, H.; Liu, L.; Kalbfleisch, J.H.; Gao, X.; Kao, R.L.; Williams, D.L.; et al. Attenuation of Cardiac Dysfunction in Polymicrobial Sepsis by MicroRNA-146a Is Mediated via Targeting of IRAK1 and TRAF6 Expression. J. Immunol. 2015, 195, 672–682. [Google Scholar] [CrossRef] [Green Version]
- An, R.; Feng, J.; Xi, C.; Xu, J.; Sun, L. miR-146a Attenuates Sepsis-Induced Myocardial Dysfunction by Suppressing IRAK1 and TRAF6 via Targeting ErbB4 Expression. Oxid. Med. Cell. Longev. 2018, 2018, 7163057. [Google Scholar] [CrossRef] [Green Version]
- Xie, J.; Zhang, L.; Fan, X.; Dong, X.; Zhang, Z.; Fan, W. MicroRNA-146a improves sepsis-induced cardiomyopathy by regulating the TLR-4/NF-κB signaling pathway. Exp. Ther. Med. 2019, 18, 779–785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Yu, Y. MiR-146b protect against sepsis induced mice myocardial injury through inhibition of Notch1. J. Mol. Histol. 2018, 49, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Wang, X.; Ha, T.; Gao, M.; Liu, L.; Wang, R.; Yu, K.; Kalbfleisch, J.H.; Kao, R.L.; Williams, D.L.; et al. MicroRNA-125b Prevents Cardiac Dysfunction in Polymicrobial Sepsis by Targeting TRAF6-Mediated Nuclear Factor κB Activation and p53-Mediated Apoptotic Signaling. J. Infect. Dis. 2016, 214, 1773–1783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Wang, X.; Yan, X.; Cheng, X.; He, X.; Zheng, W. LncRNA MALAT1 regulates sepsis-induced cardiac inflammation and dysfunction via interaction with miR-125b and p38 MAPK/NFκB. Int. Immunopharmacol. 2018, 55, 69–76. [Google Scholar] [CrossRef]
- Wei, S.; Liu, Q. Long noncoding RNA MALAT1 modulates sepsis-induced cardiac inflammation through the miR-150-5p/NF-κB axis. Int. J. Clin. Exp. Pathol. 2019, 12, 3311–3319. [Google Scholar]
- Zhu, X.G.; Zhang, T.N.; Wen, R.; Liu, C.F. Overexpression of miR-150-5p Alleviates Apoptosis in Sepsis-Induced Myocardial Depression. Biomed. Res. Int. 2020, 2020, 3023186. [Google Scholar] [CrossRef]
- Wang, H.; Bei, Y.; Shen, S.; Huang, P.; Shi, J.; Zhang, J.; Sun, Q.; Chen, Y.; Yang, Y.; Xu, T.; et al. miR-21-3p controls sepsis-associated cardiac dysfunction via regulating SORBS2. J. Mol. Cell Cardiol. 2016, 94, 43–53. [Google Scholar] [CrossRef]
- Wang, H.; Bei, Y.; Huang, P.; Zhou, Q.; Shi, J.; Sun, Q.; Zhong, J.; Li, X.; Kong, X.; Xiao, J. Inhibition of miR-155 Protects Against LPS-induced Cardiac Dysfunction and Apoptosis in Mice. Mol. Ther. Nucleic Acids 2016, 5, e374. [Google Scholar] [CrossRef] [Green Version]
- Diao, X.; Sun, S. PMicroRNA-124a regulates LPS-induced septic cardiac dysfunction by targeting STX2. Biotechnol. Lett. 2017, 39, 1335–1342. [Google Scholar] [CrossRef]
- Zheng, G.; Pan, M.; Jin, W.; Jin, G.; Huang, Y. MicroRNA-135a is up-regulated and aggravates myocardial depression in sepsis via regulating p38 MAPK/NF-κB pathway. Int. Immunopharmacol. 2017, 45, 6–12. [Google Scholar] [CrossRef]
- Zhou, Y.; Song, Y.; Shaikh, Z.; Li, H.; Zhang, H.; Caudle, Y.; Zheng, S.; Yan, H.; Hu, D.; Stuart, C.; et al. MicroRNA-155 attenuates late sepsis-induced cardiac dysfunction through JNK and β-arrestin 2. Oncotarget 2017, 8, 47317–47329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ge, C.; Liu, J.; Dong, S. miRNA-214 Protects Sepsis-Induced Myocardial Injury. Shock 2018, 50, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Sang, Z.; Zhang, P.; Wei, Y.; Dong, S. miR-214-3p Attenuates Sepsis-Induced Myocardial Dysfunction in Mice by Inhibiting Autophagy through PTEN/AKT/mTOR Pathway. Biomed. Res. Int. 2020, 2020, 1409038. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Hu, J.; Wang, Z.; Zong, H.; Zhang, L.; Zhang, R.; Sun, L. LncRNA H19 functions as an Aquaporin 1 competitive endogenous RNA to regulate microRNA-874 expression in LPS sepsis. Biomed. Pharmacother. 2018, 105, 1183–1191. [Google Scholar] [CrossRef]
- Wang, W.; Li, F.; Sun, Y.; Lei, L.; Zhou, H.; Lei, T.; Xia, Y.; Verkman, A.S.; Yang, B. Aquaporin-1 retards renal cyst development in polycystic kidney disease by inhibition of Wnt signaling. FASEB J. 2015, 29, 1551–1563. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Yan, M.; Jiang, H.; Wang, Q.; Guan, X.; Chen, J.; Wang, C. Protective effects of puerarin on acute lung and cerebrum injury induced by hypobaric hypoxia via the regulation of aquaporin (AQP) via NF-κB signaling pathway. Int. Immunopharmacol. 2016, 40, 300–309. [Google Scholar] [CrossRef]
- Tomita, Y.; Dorward, H.; Yool, A.J.; Smith, E.; Townsend, A.R.; Price, T.J.; Hardingham, J.E. Role of Aquaporin 1 Signalling in Cancer Development and Progression. Int. J. Mol. Sci. 2017, 18, 299. [Google Scholar] [CrossRef] [Green Version]
- Tang, B.; Xuan, L.; Tang, M.; Wang, H.; Zhou, J.; Liu, J.; Wu, S.; Li, M.; Wang, X.; Zhang, H. miR-93-3p alleviates lipopolysaccharide-induced inflammation and apoptosis in H9c2 cardiomyocytes by inhibiting toll-like receptor 4. Pathol. Res. Pract. 2018, 214, 1686–1693. [Google Scholar] [CrossRef]
- Yao, Y.; Sun, F.; Lei, M. miR-25 inhibits sepsis-induced cardiomyocyte apoptosis by targetting PTEN. Biosci. Rep. 2018, 38, BSR20171511. [Google Scholar] [CrossRef] [Green Version]
- Wu, P.; Kong, L.; Li, J. MicroRNA-494-3p protects rat cardiomyocytes against septic shock via PTEN. Exp. Ther. Med. 2019, 17, 1706–1716. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Caudle, Y.; Shaikh, A.; Yao, B.; Yin, D. Inhibition of microRNA-23b prevents polymicrobial sepsis-induced cardiac dysfunction by modulating TGIF1 and PTEN. Biomed. Pharmacother. 2018, 103, 869–878. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Liu, Q.; Xiao, J.; Lei, J.; Liu, Y.; Xu, H.; Hong, Z. Molecular validation of the precision-cut kidney slice (PCKS) model of renal fibrosis through assessment of TGF-β1-induced Smad and p38/ERK signaling. Int. Immunopharmacol. 2016, 34, 32–36. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.K.; Quaggin, S.E.; Vaughan, D.E. Molecular basis of organ fibrosis: Potential therapeutic approaches. Exp. Biol. Med. (Maywood) 2013, 238, 461–481. [Google Scholar] [CrossRef] [PubMed]
- Cao, C.; Zhang, Y.; Chai, Y.; Wang, L.; Yin, C.; Shou, S.; Jin, H. Attenuation of Sepsis-Induced Cardiomyopathy by Regulation of MicroRNA-23b Is Mediated Through Targeting of MyD88-Mediated NF-κB Activation. Inflammation 2019, 42, 973–986. [Google Scholar] [CrossRef]
- Guo, H.; Tang, L.; Xu, J.; Lin, C.; Ling, X.; Lu, C.; Liu, Z. MicroRNA-495 serves as a diagnostic biomarker in patients with sepsis and regulates sepsis-induced inflammation and cardiac dysfunction. Eur. J. Med. Res. 2019, 24, 37. [Google Scholar] [CrossRef]
- Zhu, J.; Lin, X.; Yan, C.; Yang, S.; Zhu, Z. microRNA-98 protects sepsis mice from cardiac dysfunction, liver and lung injury by negatively regulating HMGA2 through inhibiting NF-κB signaling pathway. Cell Cycle 2019, 18, 1948–1964. [Google Scholar] [CrossRef]
- Ouyang, H.; Tan, Y.; Li, Q.; Xia, F.; Xiao, X.; Zheng, S.; Lu, J.; Zhong, J.; Hu, Y. MicroRNA-208-5p regulates myocardial injury of sepsis mice via targeting SOCS2-mediated NF-κB/HIF-1α pathway. Int. Immunopharmacol. 2020, 81, 106204. [Google Scholar] [CrossRef]
- Sun, B.; Luan, C.; Guo, L.; Zhang, B.; Liu, Y. Low expression of microRNA-328 can predict sepsis and alleviate sepsis-induced cardiac dysfunction and inflammatory response. Braz. J. Med. Biol. Res. 2020, 53, e9501. [Google Scholar] [CrossRef]
- Zhang, T.N.; Yang, N.; Goodwin, J.E.; Mahrer, K.; Li, D.; Xia, J.; Wen, R.; Zhou, H.; Zhang, T.; Song, W.L.; et al. Characterization of Circular RNA and microRNA Profiles in Septic Myocardial Depression: A Lipopolysaccharide-Induced Rat Septic Shock Model. Inflammation 2019, 42, 1990–2002. [Google Scholar] [CrossRef]
- Hansen, T.B.; Jensen, T.I.; Clausen, B.H.; Bramsen, J.B.; Finsen, B.; Damgaard, C.K.; Kjems, J. Natural RNA circles function as efficient microRNA sponges. Nature 2013, 495, 384–388. [Google Scholar] [CrossRef]
- Zhu, Y.; Sun, A.; Meng, T.; Li, H. Protective role of long noncoding RNA CRNDE in myocardial tissues from injury caused by sepsis through the microRNA-29a/SIRT1 axis. Life Sci. 2020, 255, 117849. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.X.; Ou, Y.M.; Zhou, J.Y. Gracillin inhibits apoptosis and inflammation induced by lipopolysaccharide (LPS) to alleviate cardiac injury in mice via improving miR-29a. Biochem. Biophys. Res. Commun. 2020, 523, 580–587. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Zhu, C.; Ye, C. LncRNA CYTOR attenuates sepsis-induced myocardial injury via regulating miR-24/XIAP. Cell Biochem. Funct. 2020. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.; Yuan, W.; Wu, H.; Chen, G.; Sun, Y.; Yuan, L.; Zhang, W.; Lei, M. LncRNA KCNQ1OT1 attenuates sepsis-induced myocardial injury via regulating miR-192-5p/XIAP axis. Exp. Biol. Med. (Maywood) 2020, 245, 620–630. [Google Scholar] [CrossRef] [Green Version]
- Wei, J.L.; Wu, C.J.; Chen, J.J.; Shang, F.T.; Guo, S.G.; Zhang, X.C.; Liu, H. LncRNA NEAT1 promotes the progression of sepsis-induced myocardial cell injury by sponging miR-144-3p. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 851–861. [Google Scholar]
- Xing, P.C.; An, P.; Hu, G.Y.; Wang, D.L.; Zhou, M.J. LncRNA MIAT Promotes Inflammation and Oxidative Stress in Sepsis-Induced Cardiac Injury by Targeting miR-330-5p/TRAF6/NF-κB Axis. Biochem. Genet. 2020. [Google Scholar] [CrossRef]
- Han, Y.; Cai, Y.; Lai, X.; Wang, Z.; Wei, S.; Tan, K.; Xu, M.; Xie, H. lncRNA RMRP Prevents Mitochondrial Dysfunction and Cardiomyocyte Apoptosis via the miR-1-5p/hsp70 Axis in LPS-Induced Sepsis Mice. Inflammation 2020, 43, 605–618. [Google Scholar] [CrossRef]
- Hammond, S.M. An overview of microRNAs. Adv. Drug Deliv. Rev. 2015, 87, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Boscaino, V.; Fiannaca, A.; La Paglia, L.; La Rosa, M.; Rizzo, R.; Urso, A. MiRNA therapeutics based on logic circuits of biological pathways. BMC Bioinform. 2019, 20 (Suppl. 9), 344. [Google Scholar] [CrossRef]
- Morris, K.V.; Mattick, J.S. The rise of regulatory RNA. Nat. Rev. Genet. 2014, 15, 423–437. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wang, L.; Chen, C.; Chu, X. New insights into the regulatory role of microRNA in tumor angiogenesis and clinical implications. Mol. Cancer 2018, 17, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tahamtan, A.; Teymoori-Rad, M.; Nakstad, B.; Salimi, V. Anti-Inflammatory MicroRNAs and Their Potential for Inflammatory Diseases Treatment. Front. Immunol. 2018, 9, 1377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aghabozorgi, A.S.; Ahangari, N.; Eftekhaari, T.E.; Torbati, P.N.; Bahiraee, A.; Ebrahimi, R.; Pasdar, A. Circulating exosomal miRNAs in cardiovascular disease pathogenesis: New emerging hopes. J. Cell Physiol. 2019, 234, 21796–21809. [Google Scholar] [CrossRef] [PubMed]
- Precone, V.; Stornaiuolo, G.; Amato, A.; Brancaccio, G.; Nardiello, S.; Gaeta, G.B. Different changes in mitochondrial apoptotic pathway in lymphocytes and granulocytes in cirrhotic patients with sepsis. Liver Int. 2013, 33, 834–842. [Google Scholar] [CrossRef] [PubMed]
- Marques-Rocha, J.L.; Samblas, M.; Milagro, F.I.; Bressan, J.; Martínez, J.A.; Marti, A. Noncoding RNAs, cytokines, and inflammation-related diseases. FASEB J. 2015, 29, 595–3611. [Google Scholar] [CrossRef] [Green Version]
- Brudecki, L.; Ferguson, D.A.; McCall, C.E.; El Gazzar, M. MicroRNA-146a and RBM4 form a negative feed-forward loop that disrupts cytokine mRNA translation following TLR4 responses in human THP-1 monocytes. Immunol. Cell Biol. 2013, 91, 532–540. [Google Scholar] [CrossRef]
- Zheng, D.; Yu, Y.; Li, M.; Wang, G.; Chen, R.; Fan, G.C.; Martin, C.; Xiong, S.; Peng, T. Inhibition of MicroRNA 195 Prevents Apoptosis and Multiple-Organ Injury in Mouse Models of Sepsis. J. Infect. Dis. 2016, 213, 1661–1670. [Google Scholar] [CrossRef] [Green Version]
- O’Connell, R.M.; Rao, D.S.; Baltimore, D. microRNA regulation of inflammatory responses. Annu. Rev. Immunol. 2012, 30, 295–312. [Google Scholar] [CrossRef]
- Hashemian, S.M.; Pourhanifeh, M.H.; Fadaei, S.; Velayati, A.A.; Mirzaei, H.; Hamblin, M.R. Non-coding RNAs and Exosomes: Their Role in the Pathogenesis of Sepsis. Mol. Ther. Nucleic Acids 2020, 21, 51–74. [Google Scholar] [CrossRef]
- Wu, J.T.; Kral, J.G. The NF-kappaB/IkappaB signaling system: A molecular target in breast cancer therapy. J. Surg. Res. 2005, 123, 158–169. [Google Scholar] [CrossRef]
- Feinman, R.; Siegel, D.S.; Berenson, J. Regulation of NF-kB in multiple myeloma: Therapeutic implications. Clin. Adv. Hematol. Oncol. 2004, 2, 162–166. [Google Scholar] [PubMed]
- Cao, C.; Yin, C.; Chai, Y.; Jin, H.; Wang, L.; Shou, S. Ulinastatin mediates suppression of regulatory T cells through TLR4/NF-κB signaling pathway in murine sepsis. Int. Immunopharmacol. 2018, 64, 411–423. [Google Scholar] [CrossRef] [PubMed]
- Hobai, I.A.; Edgecomb, J.; LaBarge, K.; Colucci, W.S. Dysregulation of intracellular calcium transporters in animal models of sepsis-induced cardiomyopathy. Shock 2015, 43, 3–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Epstein, S.E.; Zhou, Y.F.; Zhu, J. Infection and atherosclerosis: Emerging mechanistic paradigms. Circulation 1999, 100, e20–e28. [Google Scholar] [CrossRef]
- O’Connor, S.; Taylor, C.; Campbell, L.A.; Epstein, S.; Libby, P. Potential infectious etiologies of atherosclerosis: A multifactorial perspective. Emerg. Infect. Dis. 2001, 7, 780–788. [Google Scholar] [CrossRef]
- Maitrias, P.; Metzinger-Le Meuth, V.; Nader, J.; Reix, T.; Caus, T.; Metzinger, L. The Involvement of miRNA in Carotid-Related Stroke. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 1608–1617. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Author and Reference | Year of Publication | Sepsis Model | Brief Description of miRNAs in Sepsis-Induced Cardiac Injury |
|---|---|---|---|
| Wang et al. [32] | 2014 | In vivo | miR-223 (both 3p and 5p) down regulated, its absence enhances myocardial inflammation |
| Xue et al. [37] | 2015 | In vivo + in vitro | miR-27a up regulated, its overexpression enhances myocardial inflammation |
| Gao et al. [39] | 2015 | In vivo + in vitro | miR146a up regulated, its overexpression attenuates myocardial disfunction |
| Ma et al. [43] | 2016 | In vivo + in vitro | miR-125b down regulated, its overexpression attenuates myocardial disfunction |
| Wang et al. [47] | 2016 | In vivo + in vitro | miR-21-3p up regulated, its down regulation attenuates myocardial inflammation, while its overexpression worsens it. |
| Wang et al. [48] | 2016 | In vivo | miR-155 up regulated, its inhibition attenuates myocardial apoptosis, its overexpression worsens it. |
| Diao et al. [49] | 2017 | In vivo + in vitro | miR-124a down regulated, its overexpression attenuates myocardial damage, its down regulation worsens it. |
| Zheng et al. [50] | 2017 | In vivo + in vitro | MiR-135a up regulated, its overexpression worsens myocardial inflammation |
| Zhou et al. [51] | 2017 | In vivo | MiR-155 overexpression attenuates myocardial damage |
| An et al. [40] | 2018 | In vitro | miR-146a up regulated, its overexpression attenuates myocardial inflammation |
| Chen et al. [44] | 2018 | In vivo + in vitro | MiR-125b down regulated, its overexpression attenuates myocardial damage |
| Ge et al. [52] | 2018 | In vivo | MiR-214 up regulated, its overexpression attenuates myocardial apoptosis and damage |
| Fang et al. [54] | 2018 | In vivo + in vitro | MiR-874 up regulated, its inhibition attenuates myocardial dysfunction |
| Wang et al. [42] | 2018 | In vivo + in vitro | MiR-146b up regulated, its overexpression attenuates myocardial inflammation |
| Zhang et al. [61] | 2018 | In vivo + in vitro | MiR-23b up regulated, its inhibition attenuates myocardial dysfunction and fibrosis; |
| Tang et al. [58] | 2018 | In vitro | MiR-93-3p down regulated, its overexpression attenuates inflammation and apoptosis |
| Yao et al. [59] | 2018 | In vivo + in vitro | MiR-25 down regulated, its overexpression attenuates apoptosis |
| Wu et al. [60] | 2018 | In vivo + in vitro | MiR-494-3p down regulated and correlated with myocardial damage, its overexpression attenuates myocardial injury and apoptosis |
| Cao et al. [64] | 2019 | In vivo + in vitro | MiR-23b up regulated, its overexpression attenuates myocardial inflammation and apoptosis |
| Guo et al. [65] | 2019 | In vivo | MiR-495 down regulated, its overexpression attenuates myocardial dysfunction |
| Xie et al. [41] | 2019 | In vivo | MiR-146a up regulated, its overexpression attenuates myocardial inflammation and apoptosis |
| Zhu et al. [66] | 2019 | In vivo | MiR-98 down regulated, its overexpression attenuates myocardial damage, apoptosis, and inflammation |
| Zhang et al. [69] | 2019 | In vivo | 78 miRNAs differently expressed in myocardium during sepsis |
| Wei et Liu [45] | 2019 | In vitro | MiR-150-5p down regulated, its overexpression attenuates inflammation |
| Chen et al. [73] | 2020 | In vivo + in vitro | MiR-24 up regulated, its inhibition attenuates myocardial apoptosis |
| Sun et al. [74] | 2020 | In vivo + in vitro | MiR-192-5p up regulated, its inhibition attenuates myocardial apoptosis |
| Ouyang et al. [67] | 2020 | In vivo | MiR-208a-5p up regulated, its inhibition attenuates myocardial damage |
| Sun et al. [68] | 2020 | In vivo | MiR-328 up regulated, its inhibition attenuates myocardial inflammation |
| Zhu et al. [46] | 2020 | In vivo + in vitro | MiR-150-5p down regulated, its overexpression attenuates myocardial apoptosis |
| Zhu et al. [71] | 2020 | In vivo + in vitro | MiR29a up regulated, its inhibition attenuates myocardial damage |
| Wei et al. [75] | 2020 | In vitro | MiR-144-3p down regulated, its overexpression attenuates myocardial damage |
| Xing et al. [76] | 2020 | In vivo + in vitro | MiR-330-5p down regulated, its overexpression attenuates inflammation and oxidative stress |
| Han et al. [77] | 2020 | In vivo + in vitro | MiR-1-5p enhances myocardial damage |
| Song et al. [72] | 2020 | In vivo + in vitro | MiR-29a attenuates myocardial damage |
| Sang et al. [53] | 2020 | In vivo | MiR-214-3p up regulated, its overexpression attenuates myocardial damage and autophagy |
| MiRNA | Expression in Sepsis (Myocardium and/or Serum) | Target Genes | Reference |
|---|---|---|---|
| MiR-1-5p | its inhibition is protective | HSPA4 | Han et al. 2020 [77] |
| MiR-7a-5p | ↑ | - | Zhang et al. 2019 [69] |
| MiR-20b-3p | ↓ | - | Zhang et al. 2019 [69] |
| MiR-21-3p | ↑ Its up-regulation worsens inflammation | SORBS2 | Wang et al. 2016 [47] |
| MiR-21-5p | Zhang et al. 2019 [69] | ||
| MiR-23b | ↑ its inhibition is protective | TGIF1 PTEN TRAF6 IκκB | Zhang et al. 2018 [61] |
| its up-regulation is protective | Cao et al. 2019 [64] | ||
| MiR-24 | ↑ its inhibition is protective | XIAP | Chen et al. 2020 [73] |
| MiR-24-1-5p | ↑ | - | Zhang et al. 2019 [69] |
| MiR-24-2-5p | ↑ | - | Zhang et al. 2019 [69] |
| MiR-24-3p | ↑ | - | Zhang et al. 2019 [69] |
| MiR-25 | ↓ its up-regulation is protective | PTEN | Yao et al. 2018 [59] |
| MiR-27a | ↑ | Nrf2 | Xue et al. 2015 [37] |
| MiR-27a-5p | - | Zhang et al. 2019 [69] | |
| MiR-29a | ↑ its inhibition is protective | SIRT1 | Zhu et al. 2020 [71] |
| ↓ its up-regulation is protective | Song et al. 2020 [72] | ||
| MiR-30c-5p | ↓ | - | Zhang et al. 2019 [69] |
| MiR-30d-3p | ↓ | - | Zhang et al. 2019 [69] |
| MiR-92a-1-5p | ↑ | - | Zhang et al. 2019 [69] |
| MiR-93-5p | ↓ | - | Zhang et al. 2019 [69] |
| MiR-93-3p | ↓ its up-regulation is protective | TLR4 | Tang et al. 2018 [58] |
| MiR-98 | ↓ its up-regulation is protective | HMGA2 | Zhu et al. 2019 [66] |
| MiR-99a-5p | ↑ | - | Zhang et al. 2019 [69] |
| MiR-122-5p | ↑ | - | Zhang et al. 2019 [69] |
| MiR-124a | ↓ its up-regulation is protective | STX2 | Diao et al. 2017 [49] |
| MiR-125b | ↓ its up-regulation is protective | TRAF6 | Ma et al. 2016 [43] |
| Chen et al. 2018 [44] | |||
| MiR-126a-3p | ↑ | - | Zhang et al. 2019 [69] |
| MiR-128-3p | ↑ | - | Zhang et al. 2019 [69] |
| MiR-132-5p/3p | ↑ | - | Zhang et al. 2019 [69] |
| MiR-133a-3p/5p | ↓ | - | Zhang et al. 2019 [69] |
| MiR-135a | ↑ Its up-regulation worsens inflammation | - | Zheng et al. 2017 [50] |
| MiR-143-3p | ↑ | - | Zhang et al. 2019 [69] |
| MiR-144-3p | ↓ its up-regulation is protective | - | Wei et al. 2020 [75] |
| MiR-145-5p | ↓ | - | Zhang et al. 2019 [69] |
| MiR-146a | ↑ its up-regulation is protective | IRAK1 TRAF6 ErbB4 | Gao et al. 2015 [39] |
| An et al. 2018 [40] | |||
| Xie et al. 2019 [41] | |||
| MiR-146b | ↑ its up-regulation is protective | Notch1 | Wang et al. 2018 [42] |
| MiR-146b-5p/-3p | Zhang et al. 2019 [69] | ||
| MiR150-5p | ↓ its up-regulation is protective | Akt2 | Wei et Liu 2019 [45] |
| Zhu et al. 2020 [46] | |||
| MiR-150-3p | ↓ | Zhang et al. 2019 [69] | |
| MiR-155 | ↑ its up-regulation is protective | Pea15a Arrb2 | Wang et al. 2016 [48] |
| Zhou et al. 2017 [51] | |||
| MiR-155-3p | ↑ | - | Zhang et al. 2019 [69] |
| MiR-181b-1-3p | ↑ | - | Zhang et al. 2019 [69] |
| MiR-181b-5p | ↓ | - | Zhang et al. 2019 [69] |
| MiR-192-5p | ↑ its inhibition is protective | XIAP | Sun et al. 2020 [74] |
| MiR-195-3p | ↑ | - | Zhang et al. 2019 [69] |
| MiR-200a-5p | ↑ | - | Zhang et al. 2019 [69] |
| MiR-201-5p | ↓ | - | Zhang et al. 2019 [69] |
| MiR-208a-5p | ↑ its inhibition is protective | SOCS2 | Ouyang et al. 2020 [67] |
| MiR-210-3p | ↓ | - | Zhang et al. 2019 [69] |
| MiR-214 | ↑ its up-regulation is protective | PTEN | Ge et al. 2018 [52] |
| MiR-214-3p | Sang et al. 2020 [53] | ||
| MiR-218a-5p | ↑ | - | Zhang et al. 2019 [69] |
| MiR-219a-1-3p | ↑ | - | Zhang et al. 2019 [69] |
| MiR-223-5p | ↓ | Sema3A | Wang et al. 2014 [32] |
| MiR-233-3p | ↑ | - | Zhang et al. 2019 [69] |
| MiR-233-5p | ↑ | - | Zhang et al. 2019 [69] |
| MiR-322-5p | ↓ | - | Zhang et al. 2019 [69] |
| MiR-328 | ↑ its inhibition is protective | - | Sun et al. 2020 [68] |
| MiR-330-5p | ↑ | - | Zhang et al. 2019 [69] |
| ↓ its up-regulation is protective | TRAF6 | Xing et al. 2020 [74] | |
| MiR-339-3p | ↑ | - | Zhang et al. 2019 [69] |
| MiR-340-3p | ↑ | - | Zhang et al. 2019 [69] |
| MiR-362-5p | ↓ | - | Zhang et al. 2019 [69] |
| MiR-369-5p | ↑ | - | Zhang et al. 2019 [69] |
| MiR-378a-5p | ↓ | - | Zhang et al. 2019 [69] |
| MiR-379-5p | ↑ | - | Zhang et al. 2019 [69] |
| MiR-380-3p | ↑ | - | Zhang et al. 2019 [69] |
| MiR-409a-3p | ↑ | - | Zhang et al. 2019 [69] |
| MiR-425-3p | ↓ | - | Zhang et al. 2019 [69] |
| MiR-434-5p | ↑ | - | Zhang et al. 2019 [69] |
| MiR-466b-5p | ↑ | - | Zhang et al. 2019 [69] |
| MiR-490-5p | ↑ | - | Zhang et al. 2019 [69] |
| MiR-494-3p | ↓ its up-regulation is protective | PTEN | Wu et al. 2018 [60] |
| MiR-495 | ↓ its up-regulation is protective | - | Guo et al. 2019 [65] |
| MiR-503-5p | ↓ | - | Zhang et al. 2019 [69] |
| MiR-674-3p | ↑ | - | Zhang et al. 2019 [69] |
| MiR-708-3p | ↑ | - | Zhang et al. 2019 [69] |
| MiR-708-5p | ↓ | - | |
| MiR-874 | ↑ its inhibition is protective | AQP1 | Fang et al. 2018 [54] |
| MiR-3557-3p | ↓ | - | Zhang et al. 2019 [69] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manetti, A.C.; Maiese, A.; Paolo, M.D.; De Matteis, A.; La Russa, R.; Turillazzi, E.; Frati, P.; Fineschi, V. MicroRNAs and Sepsis-Induced Cardiac Dysfunction: A Systematic Review. Int. J. Mol. Sci. 2021, 22, 321. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22010321
Manetti AC, Maiese A, Paolo MD, De Matteis A, La Russa R, Turillazzi E, Frati P, Fineschi V. MicroRNAs and Sepsis-Induced Cardiac Dysfunction: A Systematic Review. International Journal of Molecular Sciences. 2021; 22(1):321. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22010321
Chicago/Turabian StyleManetti, Alice Chiara, Aniello Maiese, Marco Di Paolo, Alessandra De Matteis, Raffaele La Russa, Emanuela Turillazzi, Paola Frati, and Vittorio Fineschi. 2021. "MicroRNAs and Sepsis-Induced Cardiac Dysfunction: A Systematic Review" International Journal of Molecular Sciences 22, no. 1: 321. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22010321