Species-Specific and Distance-Dependent Dispersive Behaviour of Forisomes in Different Legume Species


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
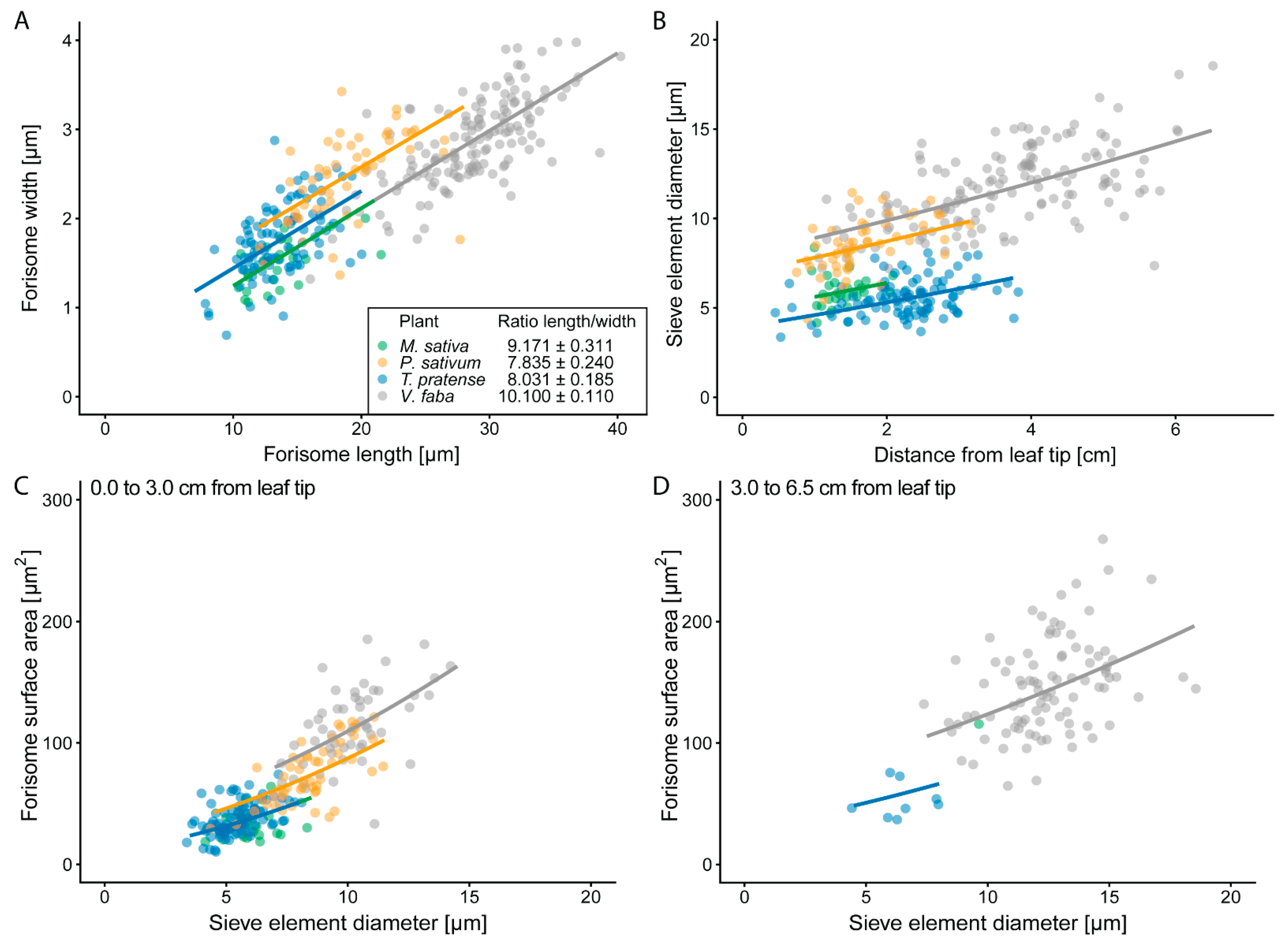
2.1. Forisome Surface Area (AF) Increases with SE Diameter
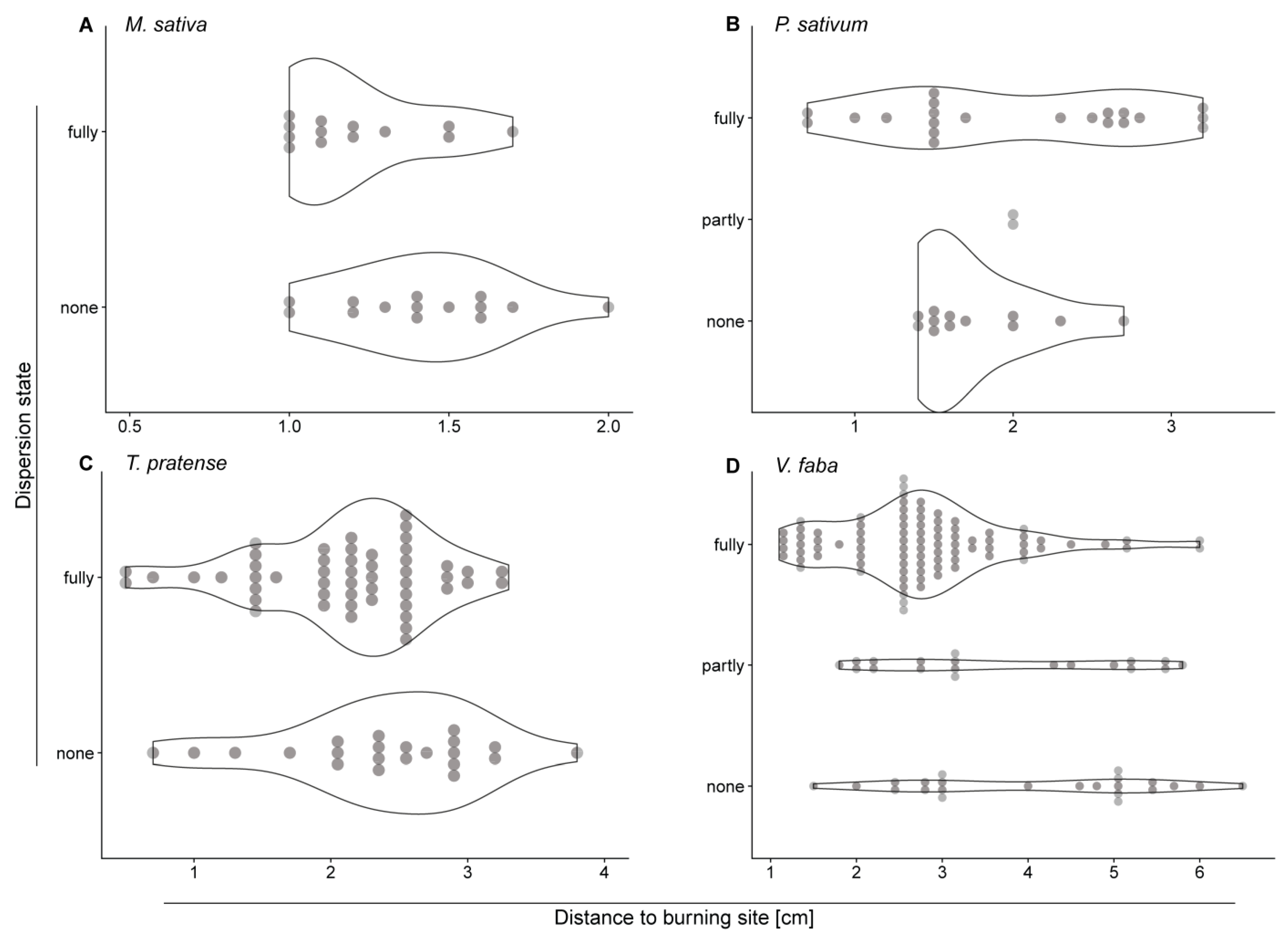
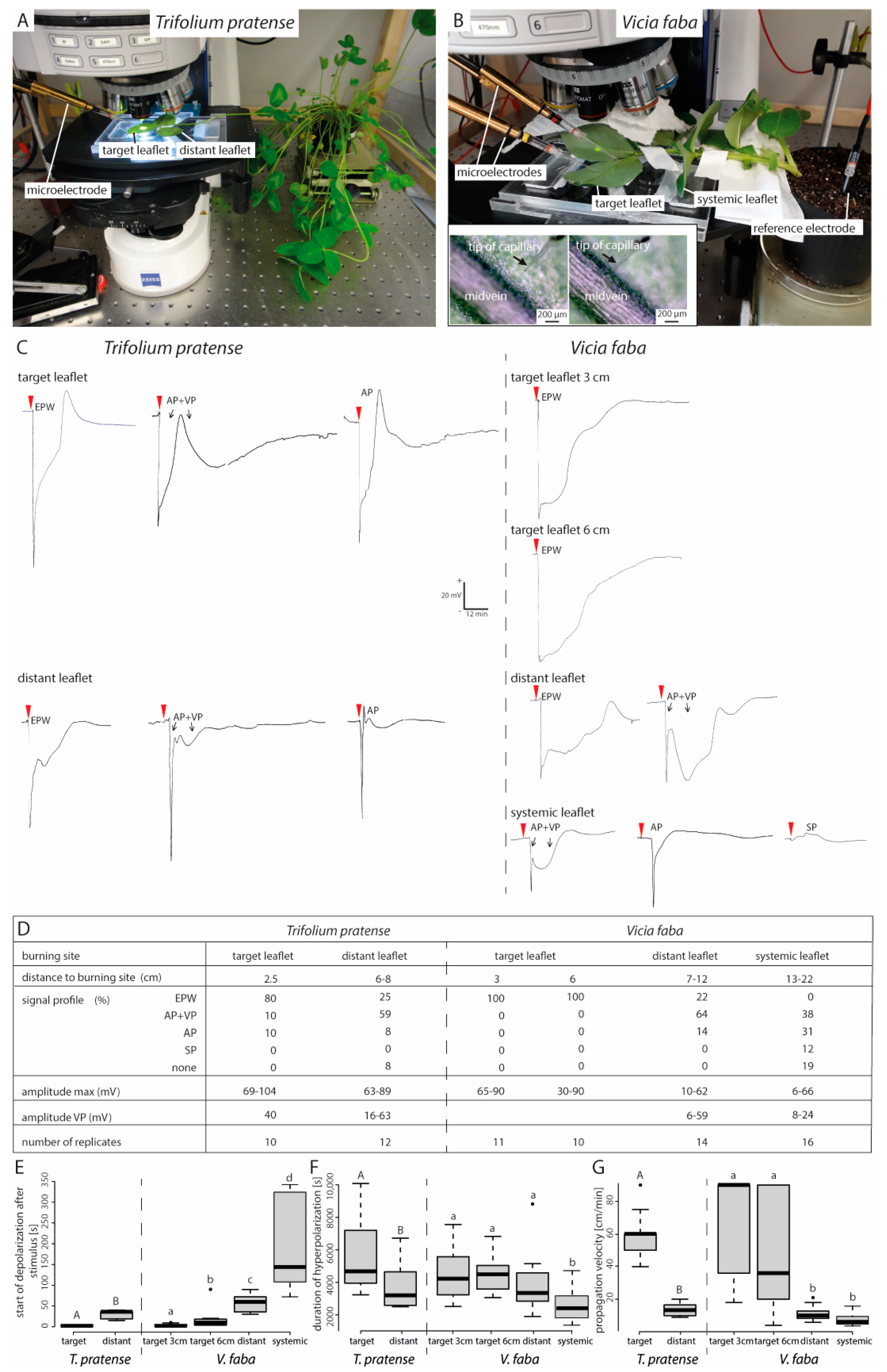
2.2. The Closer to the Stimulus Site the Higher is the Chance of Full Forisome Dispersion
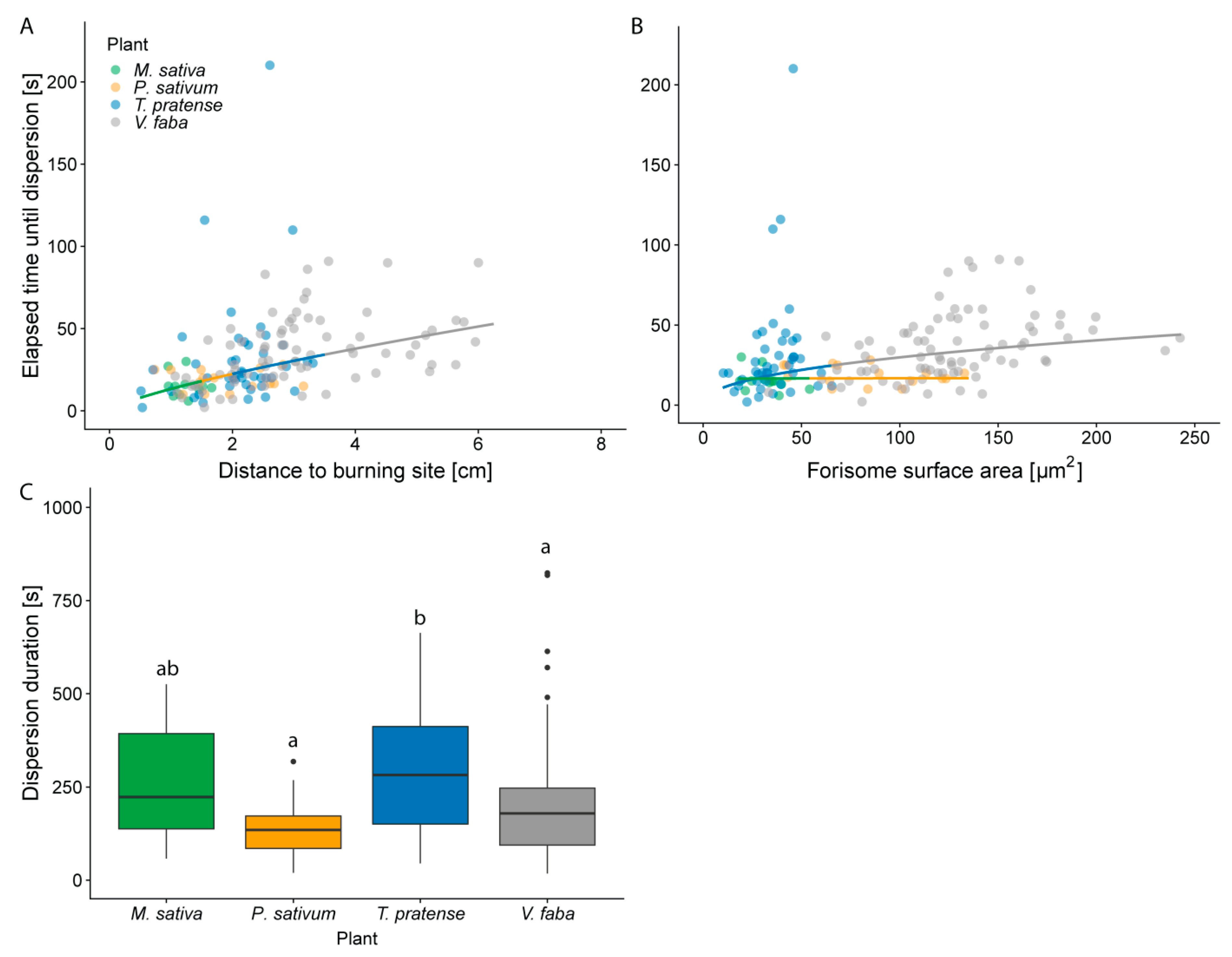
2.3. The Time Lapse until Full Forisome Dispersion Increases with Distance to Stimulus Site
2.4. Plants Differ in the Duration of Forisome Dispersion
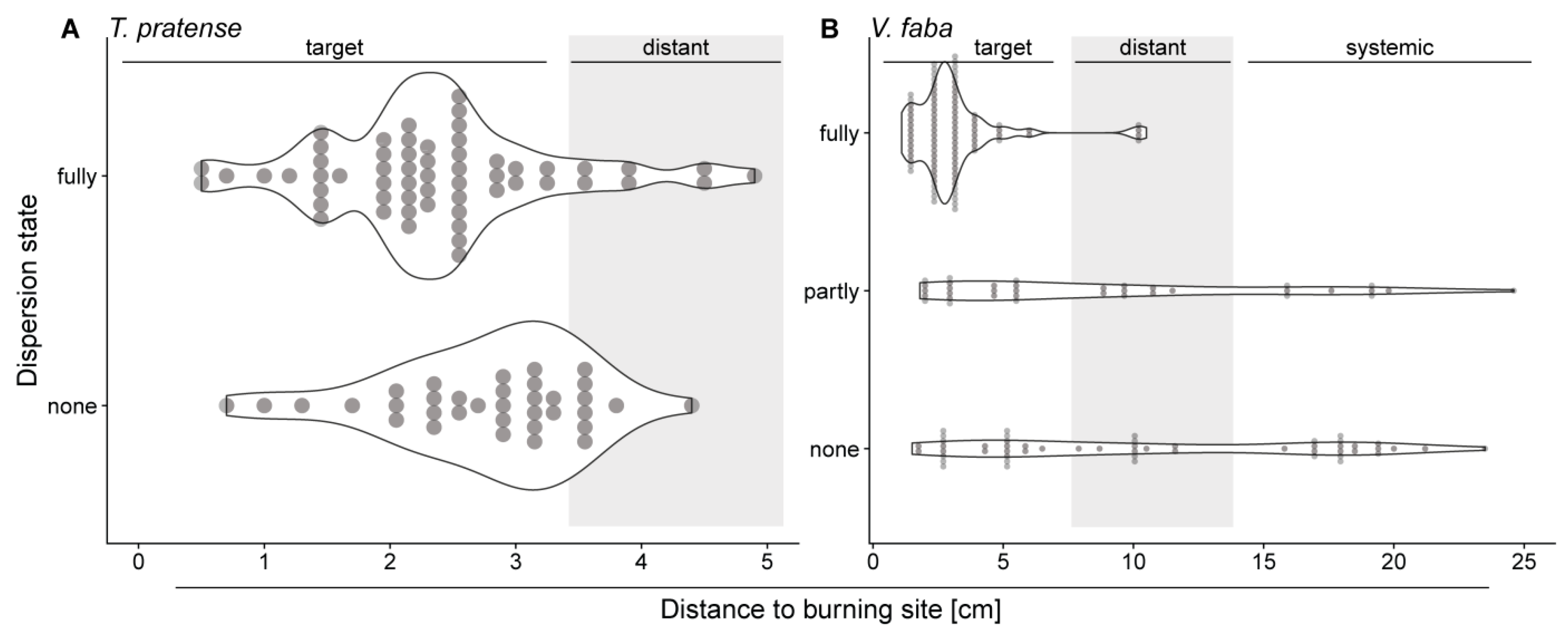
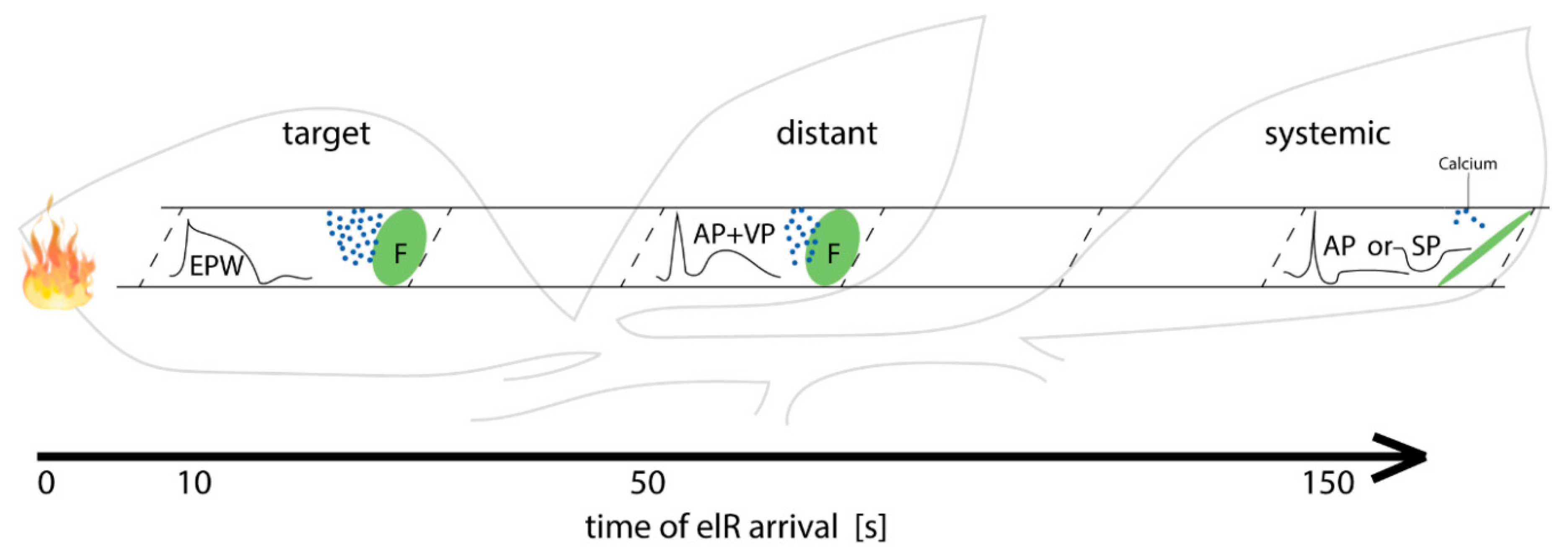
2.5. Distant and Systemic Stimuli Barely Induce Forisome Responses
2.6. Forisome Dispersion is Only Triggered by a Variation Potential
3. Discussion
3.1. Forisome Dimensions are Related to the SE Diameters: Structure Relates to Function
3.2. Forisome Dispersion is Linked to Forisome Dimensions
3.3. Ca2+-Transmembrane Movements in Response to Stimuli May Differ between Legume Species
3.4. Increasing Distance from the Stimulus Site Impacts the Type of the Passing elR
4. Material and Methods
4.1. Plant Cultivation
4.2. General Experimental Set-Up
4.3. Preparation of Intact Plants for Microscopy
4.4. Preparation of Intact Plants for Electrophysiological Measurements
4.5. Calculation of the Approximate Forisome Surface Area in the Condensed State
4.6. Illustration of the Vascular Connectivity
4.7. Statistics
4.8. Software
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AF | forisome surface area |
| AM | lateral surface area |
| AP | action potential |
| CC | companion cell |
| cv. | cultivar |
| elR | electrophysiological reaction |
| EPW | electropotential wave |
| fl | forisome lenght |
| fw | forisome width |
| h | Height of the cone |
| no. | number |
| r | base radius |
| SE | sieve element |
| SEO | sieve element occlusion |
| SEOR | sieve element occlusion related |
| SEO-F | sieve element occlusion by forisome |
| SP | system potential |
| VP | variation potential |
References
- Sukhov, V.; Sukhova, E.; Vodeneev, V. Long-distance electrical signals as a link between the local action of stressors and the systemic physiological responses in higher plants. Prog. Biophys. Mol. Biol. 2019, 146, 63–84. [Google Scholar] [CrossRef] [PubMed]
- Benning, U.F.; Tamot, B.; Guelette, B.S.; Hoffmann-Benning, S. New aspects of phloem-mediated long-distance lipid signaling in plants. Front. Plant Sci. 2012, 3, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lacombe, B.; Achard, P. Long-distance transport of phytohormones through the plant vascular system. Curr. Opin. Plant Biol. 2016, 34, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Van Bel, A.J.E.; Furch, A.C.U.; Hafke, J.B.; Knoblauch, M.; Patrick, J.W. (Questions)n on phloem biology. 2. Mass flow, molecular hopping, distribution patterns and macromolecular signalling. Plant Sci. 2011, 181, 325–330. [Google Scholar] [CrossRef]
- Van Bel, A.J.E.; Musetti, R. Sieve element biology provides leads for research on phytoplasma lifestyle in plant hosts. J. Exp. Bot. 2019, 70, 3737–3755. [Google Scholar] [CrossRef]
- Lucas, W.J.; Groover, A.; Lichtenberger, R.; Furuta, K.; Yadav, S.R.; Helariutta, Y.; He, X.Q.; Fukuda, H.; Kang, J.; Brady, S.M.; et al. The plant vascular system: Evolution, development and functions. J. Integr. Plant Biol. 2013, 55, 294–388. [Google Scholar] [CrossRef]
- Heo, J.O.; Blob, B.; Helariutta, Y. Differentiation of conductive cells: A matter of life and death. Curr. Opin. Plant Biol. 2017, 35, 23–29. [Google Scholar] [CrossRef] [Green Version]
- Otero, S.; Helariutta, Y. Companion cells: A diamond in the rough. J. Exp. Bot. 2017, 68, 71–78. [Google Scholar] [CrossRef] [Green Version]
- Van Bel, A.J.E.; Kempers, R. The pore/plasmodesm unit; key element in the interplay between sieve element and companion cell. In Progress in Botany; Behnke, H.-D., Lüttge, U., Esser, K., Kadereit, J.W., Runge, M., Eds.; Springer: Berlin/Heidelberg, Germany, 1997; Volume 58. [Google Scholar]
- Kalmbach, L.; Helariutta, Y. Sieve plate pores in the phloem and the unknowns of their formation. Plants 2019, 8, 25. [Google Scholar] [CrossRef] [Green Version]
- Furch, A.C.U.; van Bel, A.J.E.; Fricker, M.D.; Felle, H.H.; Fuchs, M.; Hafke, J.B. Sieve element Ca2+ channels as relay stations between remote stimuli and sieve tube occlusion in Vicia faba. Plant Cell 2009, 21, 2118–2132. [Google Scholar] [CrossRef] [Green Version]
- Van Bel, A.J.E.; Knoblauch, M.; Furch, A.C.U.; Hafke, J.B. (Questions)n on phloem biology. 1. Electropotential waves, Ca2+ fluxes and cellular cascades along the propagation pathway. Plant Sci. 2011, 181, 210–218. [Google Scholar] [CrossRef] [PubMed]
- Van Bel, A.J.E.; Furch, A.C.U.; Will, T.; Buxa, S.V.; Musetti, R.; Hafke, J.B. Spread the news: Systemic dissemination and local impact of Ca2+ signals along the phloem pathway. J. Exp. Bot. 2014, 65, 1761–1787. [Google Scholar] [CrossRef]
- Chen, X.Y.; Kim, J.Y. Callose synthesis in higher plants. Plant Signal. Behav. 2009, 4, 489–492. [Google Scholar] [CrossRef] [Green Version]
- Schneider, R.; Hanak, T.; Persson, S.; Voigt, C.A. Cellulose and callose synthesis and organization in focus, what’s new? Curr. Opin. Plant Biol. 2016, 34, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Knoblauch, M.; Peters, W.S.; van Bel, A.J.E. Reversible calcium-regulated stopcocks in legume sieve tubes. Plant Cell 2001, 13, 1221–1230. [Google Scholar]
- Knoblauch, M.; van Bel, A.J.E. Sieve tubes in action. Plant Cell 1998, 10, 35–50. [Google Scholar]
- Pagliari, L.; Buoso, S.; Santi, S.; Furch, A.C.U.; Martini, M.; Degola, F.; Loschi, A.; van Bel, A.J.E.; Musetti, R. Filamentous sieve element proteins are able to limit phloem mass flow, but not phytoplasma spread. J. Exp. Bot. 2017, 68, 3673–3688. [Google Scholar] [CrossRef]
- Anderson, R.; Cronshaw, J. Sieve-plate pores in tobacco and bean. Planta 1970, 91, 173–180. [Google Scholar] [CrossRef]
- Batailler, B.; Lemaitre, T.; Vilaine, F.; Sanchez, C.; Renard, D.; Cayla, T.; Beneteau, J.; Dinant, S. Soluble and filamentous proteins in Arabidopsis sieve elements. Plant Cell Environ. 2012, 35, 1258–1273. [Google Scholar] [CrossRef]
- Cronshaw, J.; Sabnis, D.D. Phloem proteins. In Sieve Elements; Behnke, H.-D., Sjolund, R.D., Eds.; Springer: Berlin/Heidelberg, Germany, 1990; pp. 257–283. [Google Scholar]
- Rüping, B.; Ernst, A.M.; Jekat, S.B.; Nordzieke, S.; Reineke, A.R.; Müller, B.; Bornberg-Bauer, E.; Prüfer, D.; Noll, G.A. Molecular and phylogenetic characterization of the sieve element occlusion gene family in Fabaceae and non-Fabaceae plants. BMC Plant Biol. 2010, 10, 219. [Google Scholar] [CrossRef] [Green Version]
- Anstead, J.A.; Froelich, D.R.; Knoblauch, M.; Thompson, G.A. Arabidopsis P-protein filament formation requires both AtSEOR1 and AtSEOR2. Plant Cell Physiol. 2012, 53, 1033–1042. [Google Scholar] [CrossRef] [Green Version]
- Lawton, D.M. Ultrastructural comparison of the tailed and tailless P-protein crystals respectively of runner bean (Phaseolus multiflorus) and garden pea (Pisum sativum) with tilting stage electron microscopy. Protoplasma 1978, 97, 1–11. [Google Scholar] [CrossRef]
- Lawton, D.M. P-protein crystals do not disperse in uninjured sieve elements in roots of runner bean (Phaseolus multiflorus) fixed with glutaraldehyde. Ann. Bot. 1978, 42, 353–361. [Google Scholar] [CrossRef]
- Knoblauch, M.; Noll, G.A.; Müller, T.; Prüfer, D.; Schneider-Huther, I.; Scharner, D.; van Bel, A.J.E.; Peters, W.S. ATP-independent contractile proteins from plants. Nat. Mater. 2003, 2, 600–603. [Google Scholar] [CrossRef]
- Mullendore, D.L.; Ross-Elliott, T.; Liu, Y.; Hellmann, H.H.; Roalson, E.H.; Peters, W.S.; Knoblauch, M. Non-dispersive phloem-protein bodies (NPBs) of Populus trichocarpa consist of a SEOR protein and do not respond to cell wounding and Ca2+. PeerJ 2018, 6, e4665. [Google Scholar] [CrossRef] [Green Version]
- Peters, W.S.; Haffer, D.; Hanakam, C.B.; van Bel, A.J.E.; Knoblauch, M. Legume phylogeny and the evolution of a unique contractile apparatus that regulates phloem transport. Am. J. Bot. 2010, 97, 797–808. [Google Scholar] [CrossRef]
- Ernst, A.M.; Rüping, B.; Jekat, S.B.; Nordzieke, S.; Reineke, A.R.; Müller, B.; Bornberg-Bauer, E.; Prüfer, D.; Noll, G.A. The sieve element occlusion gene family in dicotyledonous plants. Plant Signal. Behav. 2011, 6, 151–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, B.; Groscurth, S.; Menzel, M.; Rüping, B.A.; Twyman, R.M.; Prüfer, D.; Noll, G.A. Molecular and ultrastructural analysis of forisome subunits reveals the principles of forisome assembly. Ann. Bot. 2014, 113, 1121–1137. [Google Scholar] [CrossRef] [Green Version]
- Pelissier, H.C.; Peters, W.S.; Collier, R.; van Bel, A.J.E.; Knoblauch, M. GFP tagging of sieve element occlusion (SEO) proteins results in green fluorescent forisomes. Plant Cell Physiol. 2008, 49, 1699–1710. [Google Scholar] [CrossRef]
- Froelich, D.R.; Mullendore, D.L.; Jensen, K.H.; Ross-Elliott, T.J.; Anstead, J.A.; Thompson, G.A.; Pelissier, H.C.; Knoblauch, M. Phloem ultrastructure and pressure flow: Sieve-element-occlusion-related agglomerations do not affect translocation. Plant Cell 2011, 23, 4428–4445. [Google Scholar] [CrossRef] [Green Version]
- Furch, A.C.U.; Hafke, J.B.; Schulz, A.; van Bel, A.J.E. Ca2+-mediated remote control of reversible sieve tube occlusion in Vicia faba. J. Exp. Bot. 2007, 58, 2827–2838. [Google Scholar] [CrossRef] [Green Version]
- Knoblauch, M.; Oparka, K. The structure of the phloem—Still more questions than answers. Plant J. 2012, 70, 147–156. [Google Scholar] [CrossRef] [Green Version]
- Furch, A.C.U.; van Bel, A.J.E.; Will, T. Aphid salivary proteases are capable of degrading sieve-tube proteins. J. Exp. Bot. 2015, 66, 533–539. [Google Scholar] [CrossRef] [Green Version]
- Gilroy, S.; Bialasek, M.; Suzuki, N.; Gorecka, M.; Devireddy, A.R.; Karpinski, S.; Mittler, R. ROS, calcium, and electric signals: Key mediators of rapid systemic signaling in plants. Plant Physiol. 2016, 171, 1606–1615. [Google Scholar] [CrossRef]
- Hafke, J.B.; Furch, A.C.U.; Fricker, M.D.; van Bel, A.J.E. Forisome dispersion in Vicia faba is triggered by Ca2+ hotspots created by concerted action of diverse Ca2+ channels in sieve elements. Plant Signal. Behav. 2009, 4, 968–972. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, M.R.; Mithöfer, A. Electrical long-distance signaling in plants. In Long-Distance Systemic Signaling and Communication in Plants; Baluska, F., Ed.; Springer: Berlin/Heidelberg, Germany, 2013; Volume 19, pp. 291–308. [Google Scholar]
- Felle, H.H.; Zimmermann, M.R. Systemic signalling in barley through action potentials. Planta 2007, 226, 203–214. [Google Scholar] [CrossRef]
- Furch, A.C.U.; Zimmermann, M.R.; Will, T.; Hafke, J.B.; van Bel, A.J.E. Remote-controlled stop of phloem mass flow by biphasic occlusion in Cucurbita maxima. J. Exp. Bot. 2010, 61, 3697–3708. [Google Scholar] [CrossRef]
- Hafke, J.B.; Ehlers, K.; Foller, J.; Holl, S.R.; Becker, S.; van Bel, A.J.E. Involvement of the sieve element cytoskeleton in electrical responses to cold shocks. Plant Physiol. 2013, 162, 707–719. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, M.R.; Felle, H.H. Dissection of heat-induced systemic signals: Superiority of ion fluxes to voltage changes in substomatal cavities. Planta 2009, 229, 539–547. [Google Scholar] [CrossRef]
- Knoblauch, M.; Stubenrauch, M.; van Bel, A.J.E.; Peters, W.S. Forisome performance in artificial sieve tubes. Plant Cell Environ. 2012, 35, 1419–1427. [Google Scholar] [CrossRef]
- Peters, W.S.; Knoblauch, M.; Warmann, S.A.; Pickard, W.F.; Shen, A.Q. Anisotropic contraction in forisomes: Simple models won’t fit. Cell Motil. Cytoskelet. 2008, 65, 368–378. [Google Scholar] [CrossRef]
- Medina-Ortega, K.J.; Walker, G.P. Faba bean forisomes can function in defence against generalist aphids. Plant Cell Environ. 2015, 38, 1167–1177. [Google Scholar] [CrossRef] [Green Version]
- Wojciechowski, M.F.; Lavin, M.; Sanderson, M.J. A phyolgeny of legumes (Leguminosae) based on analysis of the plastid matK gene resolves many well-supported subclades within the family. Am. J. Bot. 2004, 91, 1846–1862. [Google Scholar] [CrossRef]
- Koenen, E.J.M.; Ojeda, D.I.; Steeves, R.; Migliore, J.; Bakker, F.T.; Wieringa, J.J.; Kidner, C.; Hardy, O.J.; Pennington, R.T.; Bruneau, A.; et al. Large-scale genomic sequence data resolve the deepest divergences in the legume phylogeny and support a near-simultaneous evolutionary origin of all six subfamilies. New Phytol. 2020, 225, 1355–1369. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Peters, W.S.; Froelich, D.R.; Howell, A.H.; Mooney, S.; Evans, J.E.; Hellmann, H.A.; Knoblauch, M. Aspartate residues in a forisome-forming SEO protein are critical for protein body assembly and Ca2+-responsiveness. Plant Cell Physiol. 2020, 61, 1699–1710. [Google Scholar] [CrossRef]
- Rose, J.; Visser, F.; Müller, B.; Senft, M.; Groscurth, S.; Sicking, K.F.; Twyman, R.M.; Prüfer, D.; Noll, G.A. Identification and molecular analysis of interaction sites in the MtSEO-F1 protein involved in forisome assembly. Int. J. Biol. Macromol. 2020, 144, 603–614. [Google Scholar] [CrossRef]
- Peters, W.S.; van Bel, A.J.E.; Knoblauch, M. The geometry of the forisome-sieve element-sieve plate complex in the phloem of Vicia faba L. leaflets. J. Exp. Bot. 2006, 57, 3091–3098. [Google Scholar] [CrossRef]
- Tuteja, N.; Umate, P.; van Bel, A.J.E. Forisomes: Calcium-powered protein complexes with potential as ‘smart’ biomaterials. Trends Biotechnol. 2010, 28, 102–110. [Google Scholar] [CrossRef]
- Srivastava, V.K.; Tuteja, R.; Tuteja, N. Calcium-energized motor protein forisome controls damage in phloem: Potential applications as biomimetic “smart” material. Crit. Rev. Biotechnol. 2013, 35, 173–183. [Google Scholar] [CrossRef]
- Groscurth, S.; Müller, B.; Schwan, S.; Menzel, M.; Diekstall, F.; Senft, M.; Kendall, A.; Kommor, B.A.; Neumann, U.; Kalischuk, M.; et al. Artificial forisomes are ideal models of forisome assembly and activity that allow the development of technical devices. Biomacromolecules 2012, 13, 3076–3086. [Google Scholar] [CrossRef]
- Knoblauch, M.; Peters, W.S. Forisomes, a novel type of Ca2+-dependent contractile protein motor. Cell Motil. Cytoskelet. 2004, 58, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Schwan, S.; Fritzsche, M.; Cismak, A.; Heilmann, A.; Spohn, U. In vitro investigation of the geometric contraction behavior of chemo-mechanical P-protein aggregates (forisomes). Biophys. Chem. 2007, 125, 444–452. [Google Scholar] [CrossRef] [PubMed]
- Kudla, J.; Batistic, O.; Hashimoto, K. Calcium signals: The lead currency of plant information processing. Plant Cell 2010, 22, 541–563. [Google Scholar] [CrossRef]
- Huda, K.M.; Banu, M.S.; Tuteja, R.; Tuteja, N. Global calcium transducer P-type Ca2+-ATPases open new avenues for agriculture by regulating stress signalling. J. Exp. Bot. 2013, 64, 3099–3109. [Google Scholar] [CrossRef] [PubMed]
- Kiep, V.; Vadassery, J.; Lattke, J.; Maass, J.P.; Boland, W.; Peiter, E.; Mithöfer, A. Systemic cytosolic Ca2+ elevation is activated upon wounding and herbivory in Arabidopsis. New Phytol. 2015, 207, 996–1004. [Google Scholar] [CrossRef]
- Mousavi, S.A.; Chauvin, A.; Pascaud, F.; Kellenberger, S.; Farmer, E.E. Glutamate receptor-like genes mediate leaf-to-leaf wound signalling. Nature 2013, 500, 422–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmermann, M.R.; Mithöfer, A.; Will, T.; Felle, H.H.; Furch, A.C.U. Herbivore-triggered electrophysiological reactions: Candidates for systemic signals in higher plants and the challenge of their identification. Plant Physiol. 2016, 170, 2407–2419. [Google Scholar] [CrossRef] [Green Version]
- Hedrich, R.; Salvador-Recatalà, V.; Dreyer, I. Electrical wiring and long-distance plant communication. Trends Plant Sci. 2016, 21, 376–387. [Google Scholar] [CrossRef]
- Ju, J.; Kim, K.; Lee, K.J.; Lee, W.H.; Ju, H.J. Localization of Barley yellow dwarf virus movement protein modulating programmed cell death in Nicotiana benthamiana. Plant Pathol. J. 2017, 33, 53–65. [Google Scholar] [CrossRef] [Green Version]
- Rinaldelli, E.; Luvisi, A.; Panattoni, A. Virus interference with trans-plasma membrane activity in infected grapevine leaves. Acta Physiol. Plant. 2014, 36, 3345–3349. [Google Scholar] [CrossRef]
- Rhodes, J.D.; Thain, J.F.; Wildon, D.C. The pathway for systemic electrical signal conduction in the wounded tomato plant. Planta 1996, 200, 50–57. [Google Scholar] [CrossRef]
- Furch, A.C.U.; Hafke, J.B.; van Bel, A.J.E. Plant- and stimulus-specific variations in remote-controlled sieve-tube occlusion. Plant Signal. Behav. 2008, 3, 858–861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vodeneev, V.; Akinchits, E.; Sukhov, V. Variation potential in higher plants: Mechanisms of generation and propagation. Plant Signal. Behav. 2015, 10, e1057365. [Google Scholar] [CrossRef]
- Stahlberg, R.; Cleland, R.E.; Van Volkenburgh, E. Slow wave potentials—A propagating electrical signal unique to higher plants. In Communication in Plants, 1st ed.; Baluska, F., Mancuso, S., Volkmann, D., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; pp. 291–308. [Google Scholar]
- Kurenda, A.; Nguyen, C.T.; Chetelat, A.; Stolz, S.; Farmer, E.E. Insect-damaged Arabidopsis moves like wounded Mimosa pudica. Proc. Natl. Acad. Sci. USA 2019, 116, 26066–26071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farmer, E.E.; Gao, Y.Q.; Lenzoni, G.; Wolfender, J.L.; Wu, Q. Wound- and mechanostimulated electrical signals control hormone responses. New Phytol. 2020, 227, 1037–1050. [Google Scholar] [CrossRef] [PubMed]
- Eschrich, W. Bidirectional transport. In Transport in Plants I: Phloem Transport; Zimmermann, M.H., Milburn, J.A., Eds.; Springer: Berlin/Heidelberg, Germany, 1975; pp. 245–255. [Google Scholar]
- Furch, A.C.U.; Paulmann, M.K.; Wegner, L.; Kunert, G.; Van Bel, A.J.E. Comparison of intracellular location and stimulus reaction times of forisomes in sieve tubes of four legume species. Plant Signal. Behav. 2018, 13, e1503493. [Google Scholar] [CrossRef] [Green Version]
- Crawley, M.J. The R Book; John Wiley & Sons Ltd.: West Sussex, UK, 2013. [Google Scholar]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; R Core Team. nlme: Linear and Nonlinear Mixed Effects Models, R Package Version 3.1-148. Available online: https://CRAN.R-project.org/package=nlme (accessed on 28 May 2020).
- Zuur, A.F.; Ieno, E.N.; Walker, N.J.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Springer: New York, NY, USA, 2009. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 1 August 2018).







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paulmann, M.K.; Zimmermann, M.R.; Wegner, L.; van Bel, A.J.E.; Kunert, G.; Furch, A.C.U. Species-Specific and Distance-Dependent Dispersive Behaviour of Forisomes in Different Legume Species. Int. J. Mol. Sci. 2021, 22, 492. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020492
Paulmann MK, Zimmermann MR, Wegner L, van Bel AJE, Kunert G, Furch ACU. Species-Specific and Distance-Dependent Dispersive Behaviour of Forisomes in Different Legume Species. International Journal of Molecular Sciences. 2021; 22(2):492. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020492
Chicago/Turabian StylePaulmann, Maria K., Matthias R. Zimmermann, Linus Wegner, Aart J. E. van Bel, Grit Kunert, and Alexandra C. U. Furch. 2021. "Species-Specific and Distance-Dependent Dispersive Behaviour of Forisomes in Different Legume Species" International Journal of Molecular Sciences 22, no. 2: 492. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020492