The Role of Endogenous Antimicrobial Peptides in Modulating Innate Immunity of the Ocular Surface in Dry Eye Diseases


Abstract
:1. Introduction
2. Antimicrobial Peptides
3. Distribution/Expression of AMPs in the Ocular Surface
4. Human Beta Defensins and Cathelicidin
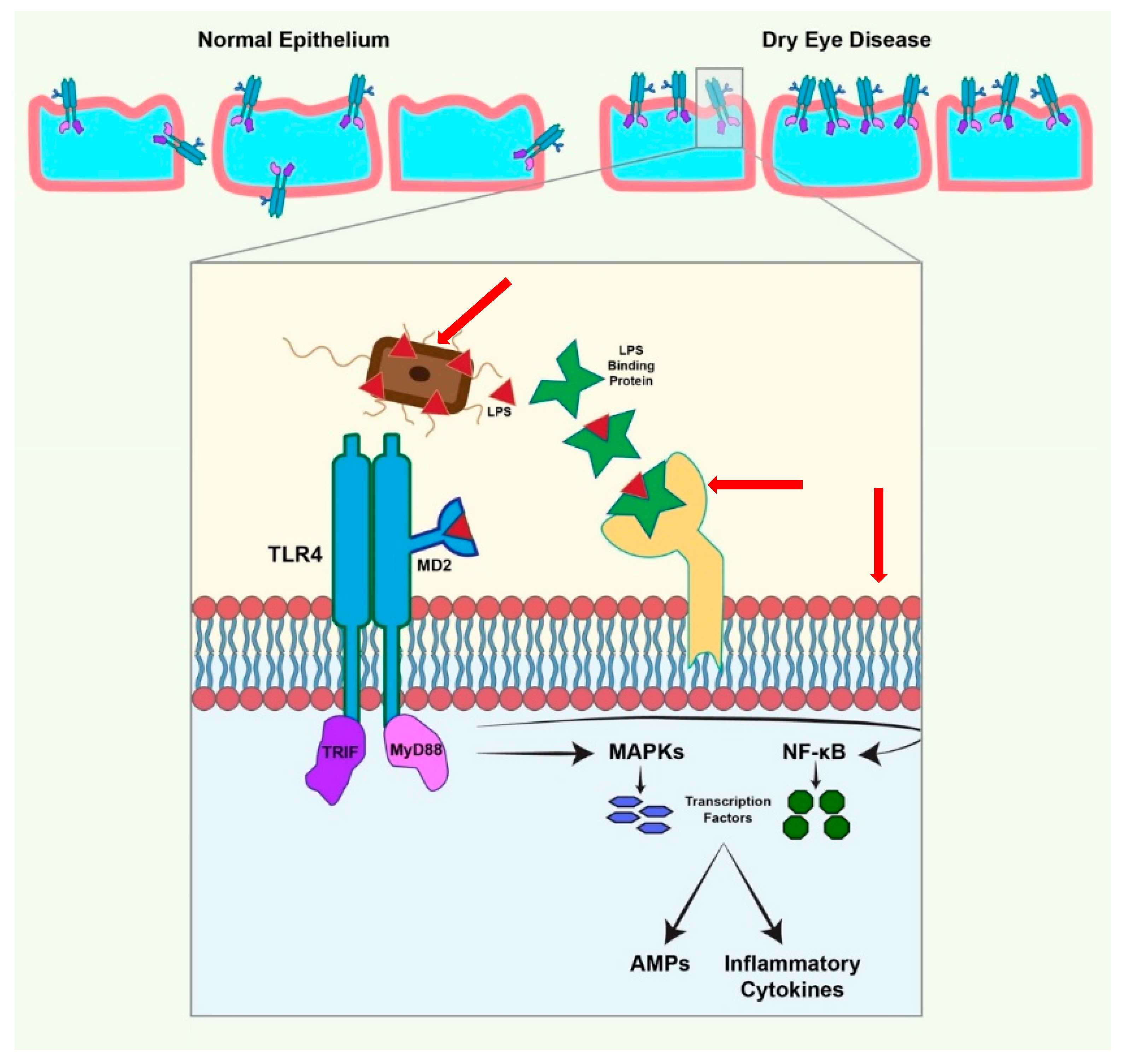
5. Toll-Like Receptor-Mediated Expression of Defensins
6. TLR Expression on the Ocular Surface
7. Thymosin Beta
8. Histatins
9. Lacritin
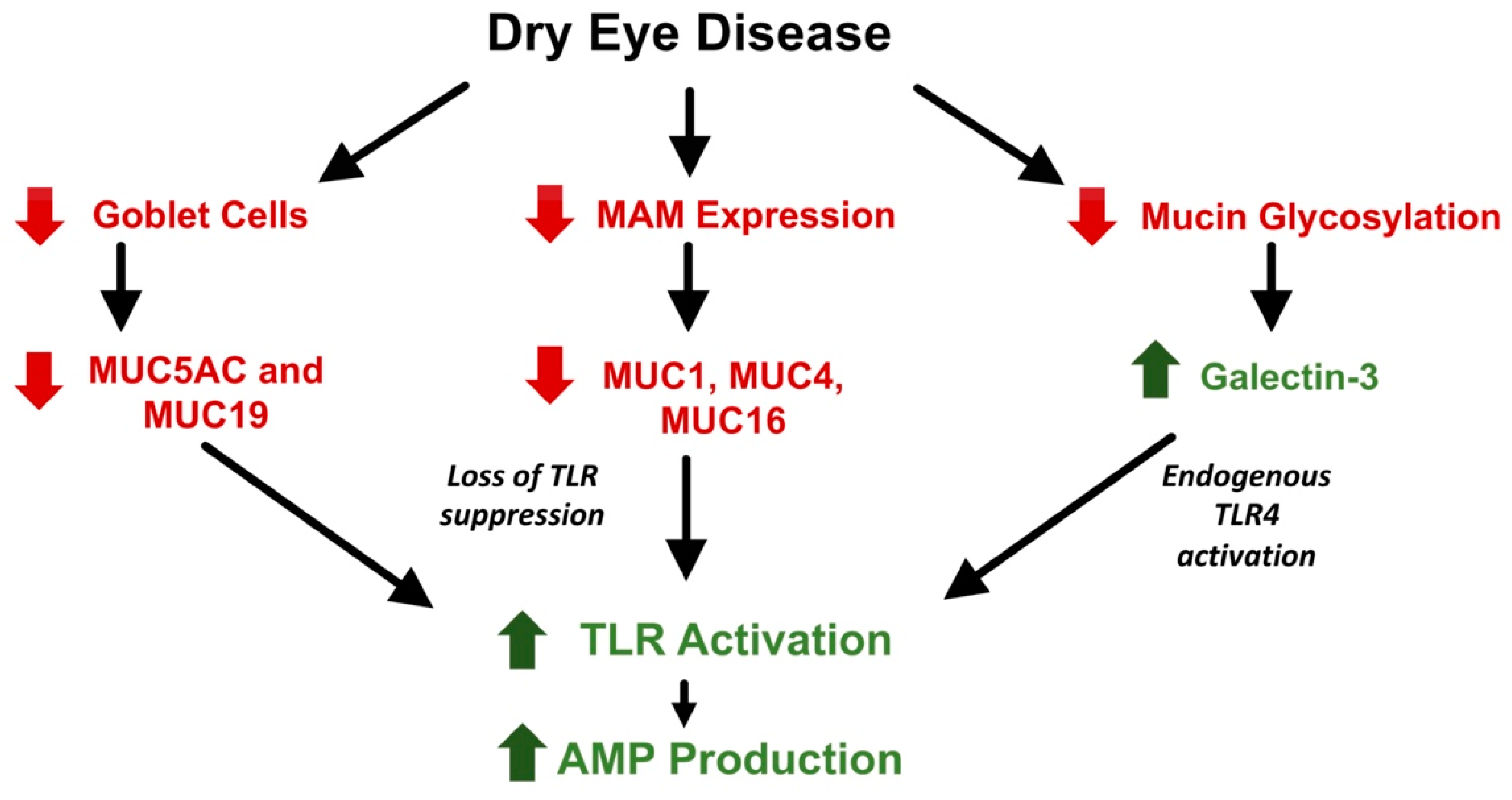
10. Role of AMPs in Dry Eye Diseases
11. AMPs in Other OSDs and Therapeutic Avenues
12. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- De Paiva, C.S.; Pflugfelder, S.C. Rationale for anti-inflammatory therapy in dry eye syndrome. Arq. Bras. Oftalmol. 2009, 71, 89–95. [Google Scholar] [CrossRef] [Green Version]
- Bron, A.J.; de Paiva, C.S.; Chauhan, S.K.; Bonini, S.; Gabison, E.E.; Jain, S.; Knop, E.; Markoulli, M.; Ogawa, Y.; Perez, V.; et al. TFOS DEWS II pathophysiology report. Ocul. Surf. 2017, 15, 438–510. [Google Scholar] [CrossRef] [PubMed]
- Gipson, I.K. The ocular surface: The challenge to enable and protect vision. The Friedenwald lecture. Investig. Ophthalmol. Vis. Sci. 2007, 48, 4390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doan, T.; Akileswaran, L.; Andersen, D.; Johnson, B.; Ko, N.; Shrestha, A.; Shestopalov, V.; Lee, C.S.; Lee, A.Y.; Van Gelder, R.N. Paucibacterial microbiome and resident DNA virome of the healthy conjunctiva. Investig. Ophthalmol. Vis. Sci. 2016, 57, 5116–5126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simmons, K.T.; Xiao, Y.; Pflugfelder, S.C.; de Paiva, C.S. Inflammatory response to lipopolysaccharide on the ocular surface in a murine dry eye model. Investig. Ophthalmol. Vis. Sci. 2016, 57, 2443. [Google Scholar] [CrossRef] [Green Version]
- Redfern, R.L.; Patel, N.; Hanlon, S.; Farley, W.; Gondo, M.; Pflugfelder, S.C.; McDermott, A.M. Toll-like receptor expression and activation in mice with experimental dry eye. Investig. Ophthalmol. Vis. Sci. 2013, 54, 1554–1563. [Google Scholar] [CrossRef]
- Redfern, R.L.; Barabino, S.; Baxter, J.; Lema, C.; McDermott, A.M. Dry eye modulates the expression of toll-like receptors on the ocular surface. Exp. Eye Res. 2015, 134, 80–89. [Google Scholar] [CrossRef] [Green Version]
- Craig, J.P.; Nichols, K.K.; Akpek, E.K.; Caffery, B.; Dua, H.S.; Joo, C.K.; Liu, Z.; Nelson, J.D.; Nichols, J.J.; Tsubota, K.; et al. TFOS DEWS II Definition and Classification Report. Ocul. Surf. 2017, 15, 276–283. [Google Scholar] [CrossRef]
- Li, D.Q.; Chen, Z.; Song, X.J.; Luo, L.; Pflugfelder, S.C. Stimulation of matrix metalloproteinases by hyperosmolarity via a JNK pathway in human corneal epithelial cells. Investig. Ophthalmol. Vis. Sci. 2004, 45, 4302–4311. [Google Scholar] [CrossRef]
- Sambursky, R.; OBrien, T.P. MMP-9 and the perioperative management of LASIK surgery. Curr. Opin. Ophthalmol. 2011, 22, 294–303. [Google Scholar] [CrossRef] [Green Version]
- Shah, D.; Ali, M.; Shukla, D.; Jain, S.; Aakalu, V.K. Effects of histatin-1 peptide on human corneal epithelial cells. PLoS ONE 2017, 12, e0178030. [Google Scholar] [CrossRef] [PubMed]
- Shah, D.; Son, K.N.; Kalmodia, S.; Lee, B.S.; Ali, M.; Balasubramaniam, A.; Shukla, D.; Aakalu, V.K. Wound Healing Properties of Histatin-5 and Identification of a Functional Domain Required for Histatin-5-Induced Cell Migration. Mol. Ther. Methods Clin. Dev. 2020, 17, 709–716. [Google Scholar] [CrossRef] [PubMed]
- Sosne, G.; Szliter, E.A.; Barrett, R.; Kernacki, K.A.; Kleinman, H.; Hazlett, L.D. Thymosin beta 4 promotes corneal wound healing and decreases inflammation in vivo following alkali injury. Exp. Eye Res. 2002, 74, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Meduri, A.; Scalinci, S.Z.; Morara, M.; Ceruti, P.; Grenga, P.L.; Zigiotti, G.L.; Scorolli, L. Effect of basic fibroblast growth factor in transgenic mice: Corneal epithelial healing process after excimer laser photoablation. Ophthalmologica 2009, 223, 139–144. [Google Scholar] [CrossRef]
- Scalinci, S.Z.; Scorolli, L.; Meduri, A.; Grenga, P.L.; Corradetti, G.; Metrangolo, C. Effect of basic fibroblast growth factor and cytochrome c peroxidase combination in transgenic mice corneal epithelial healing process after excimer laser photoablation. Clin. Ophthalmol. 2011, 5, 215–221. [Google Scholar] [CrossRef] [Green Version]
- Meduri, A.; Grenga, P.L.; Scorolli, L.; Ceruti, P.; Ferreri, G. Role of cysteine in corneal wound healing after photorefractive keratectomy. Ophthalmic Res. 2009, 41, 76–82. [Google Scholar] [CrossRef]
- Wang, G. Human antimicrobial peptides and proteins. Pharmaceuticals 2014, 7, 545–594. [Google Scholar] [CrossRef] [Green Version]
- Otvos, L. Antibacterial peptides and proteins with multiple cellular targets. J. Pept. Sci. 2005, 11, 697–706. [Google Scholar] [CrossRef]
- Hsu, C.H.; Chen, C.; Jou, M.L.; Lee, A.Y.L.; Lin, Y.C.; Yu, Y.P.; Huang, W.T.; Wu, S.H. Structural and DNA-binding studies on the bovine antimicrobial peptide, indolicidin: Evidence for multiple conformations involved in binding to membranes and DNA. Nucleic Acids Res. 2005, 33, 4053–4064. [Google Scholar] [CrossRef] [Green Version]
- Boman, H.G.; Agerberth, B.; Boman, A. Mechanisms of action on Escherichia coli of cecropin P1 and PR-39, two antibacterial peptides from pig intestine. Infect. Immun. 1993, 61, 2978–2984. [Google Scholar] [CrossRef] [Green Version]
- Funderburg, N.; Lederman, M.M.; Feng, Z.; Drage, M.G.; Jadlowsky, J.; Harding, C.V.; Weinberg, A.; Sieg, S.F. Human β-defensin-3 activates professional antigen-presenting cells via Toll-like receptors 1 and 2. Proc. Natl. Acad. Sci. USA 2007, 104, 18631–18635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van den Berg, R.H.; Faber-Krol, M.C.; Van Wetering, S.; Hiemstra, P.S.; Daha, M.R. Inhibition of activation of the classical pathway of complement by human neutrophil defensins. Blood 1998, 92, 3898–3903. [Google Scholar] [CrossRef] [PubMed]
- Befus, A.D.; Mowat, C.; Gilchrist, M.; Hu, J.; Solomon, S.; Bateman, A. Neutrophil defensins induce histamine secretion from mast cells: Mechanisms of action. J. Immunol. 1999, 163, 947–953. [Google Scholar] [PubMed]
- Zhou, L.; Beuerman, R.W.; Huang, L.; Barathi, A.; Yong, H.F.; Li, S.F.Y.; Fook, T.C.; Tan, D. Proteomic analysis of rabbit tear fluid: Defensin levels after an experimental corneal wound are correlated to wound closure. Proteomics 2007, 7, 3194–3206. [Google Scholar] [CrossRef] [PubMed]
- Vallespi, M.G.; Glaria, L.A.; Reyes, O.; Garay, H.E.; Ferrero, J.; Araña, M.J. A Limulus antilipopolysaccharide factor-derived peptide exhibits a new immunological activity with potential applicability in infectious diseases. Clin. Diagn. Lab. Immunol. 2000, 7, 669–675. [Google Scholar] [CrossRef] [Green Version]
- Territo, M.C.; Ganz, T.; Selsted, M.E.; Lehrer, R. Monocyte-chemotactic activity of defensins from human neutrophils. J. Clin. Investig. 1989, 84, 2017–2020. [Google Scholar] [CrossRef]
- Yang, H.; Young, D.W.; Gusovsky, F.; Chow, J.C. Cellular events mediated by lipopolysaccharide-stimulated toll-like receptor 4. MD-2 is required for activation of mitogen-activated protein kinases and Elk-1. J. Biol. Chem. 2000, 275, 20861–20866. [Google Scholar] [CrossRef] [Green Version]
- Van Watering, S.; Sterk, P.J.; Rabe, K.F.; Hiemstra, P.S. Defensins: Key players or bystanders in infection, injury, and repair in the lung? J. Allergy Clin. Immunol. 1999, 104, 1131–1138. [Google Scholar] [CrossRef]
- McDermott, A.M. The role of antimicrobial peptides at the ocular surface. Ophthalmic Res. 2009, 41, 60–75. [Google Scholar] [CrossRef] [Green Version]
- Haynes, R.J.; Tighe, P.J.; Dua, H.S. Antimicrobial defensin peptides of the human ocular surface. Br. J. Ophthalmol. 1999, 83, 737–741. [Google Scholar] [CrossRef]
- Zhou, L.; Huang, L.Q.; Beuerman, R.W.; Grigg, M.E.; Li, S.F.Y.; Chew, F.T.; Ang, L.; Stern, M.E.; Tan, D. Proteomic analysis of human tears: Defensin expression after ocular surface surgery. J. Proteome Res. 2004, 3, 410–416. [Google Scholar] [CrossRef] [PubMed]
- McDermott, A.M.; Redfern, R.L.; Zhang, B.; Pei, Y.; Huang, L.; Proske, R.J. Defensin expression by the cornea: Multiple signalling pathways mediate IL-1β stimulation of hBD-2 expression by human corneal epithelial cells. Investig. Ophthalmol. Vis. Sci. 2003, 44, 1859–1865. [Google Scholar] [CrossRef] [PubMed]
- Hattenbach, L.O.; Gumbel, H.; Kippenberger, S. Identification of beta-defensins in human conjunctiva. Antimicrob. Agents Chemother. 1998, 42, 3332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gökçınar, N.B.; Karabulut, A.A.; Onaran, Z.; Yumuşak, E.; Budak Yıldıran, F.A. Elevated Tear Human Neutrophil Peptides 1-3, Human Beta Defensin-2 Levels and Conjunctival Cathelicidin LL-37 Gene Expression in Ocular Rosacea. Ocul. Immunol. Inflamm. 2019, 27, 1174–1183. [Google Scholar] [CrossRef]
- McIntosh, R.S.; Cade, J.E.; Al-Abed, M.; Shanmuganathan, V.; Gupta, R.; Bhan, A.; Tighe, P.J.; Dua, H.S. The spectrum of antimicrobial peptide expression at the ocular surface. Investig. Ophthalmol. Vis. Sci. 2005, 46, 1379–1385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, L.C.; Jean, D.; Proske, R.J.; Reins, R.Y.; McDermott, A.M. Ocular surface expression and in vitro activity of antimicrobial peptides. Curr. Eye Res. 2007, 32, 595–609. [Google Scholar] [CrossRef] [Green Version]
- Abedin, A.; Mohammed, I.; Hopkinson, A.; Dua, H.S. A novel antimicrobial peptide on the ocular surface shows decreased expression in inflammation and infection. Investig. Ophthalmol. Vis. Sci. 2008, 49, 28–33. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.C.; Petkova, T.D.; Reins, R.Y.; Proske, R.J.; McDermott, A.M. Multifunctional roles of human cathelicidin (LL-37) at the ocular surface. Investig. Ophthalmol. Vis. Sci. 2006, 47, 2369–2380. [Google Scholar] [CrossRef]
- Gregory, M.S. Innate immune system and the eye. Encycl. Eye 2010, 439–445. [Google Scholar] [CrossRef]
- Kalmodia, S.; Son, K.N.; Cao, D.; Lee, B.S.; Surenkhuu, B.; Shah, D.; Ali, M.; Balasubramaniam, A.; Jain, S.; Aakalu, V.K. Presence of Histatin-1 in Human Tears and Association with Aqueous Deficient Dry Eye Diagnosis: A Preliminary Study. Sci. Rep. 2019, 9, 10304. [Google Scholar] [CrossRef] [Green Version]
- Harder, J.; Schröder, J.M. RNase 7, a novel innate immune defense antimicrobial protein of healthy human skin. J. Biol. Chem. 2002, 277, 46779–46784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohammed, I.; Yeung, A.; Abedin, A.; Hopkinson, A.; Dua, H.S. Signalling pathways involved in ribonuclease-7 expression. Cell. Mol. Life Sci. 2011, 68, 1941–1952. [Google Scholar] [CrossRef] [PubMed]
- Ganz, T. Defensins: Antimicrobial peptides of innate immunity. Nat. Rev. Immunol. 2003, 3, 710–720. [Google Scholar] [CrossRef]
- Faurschou, M.; Sørensen, O.E.; Johnsen, A.H.; Askaa, J.; Borregaard, N. Defensin-rich granules of human neutrophils: Characterization of secretory properties. Biochim. Biophys. Acta Mol. Cell Res. 2002, 1591, 29–35. [Google Scholar] [CrossRef] [Green Version]
- Zanetti, M.; Gennaro, R.; Romeo, D. Cathelicidins: A novel protein family with a common proregion and a variable C-terminal antimicrobial domain. FEBS Lett. 1995, 374, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.C.; Reins, R.Y.; Gallo, R.L.; McDermott, A.M. Cathelicidin-deficient (Cnlp-/-) mice show increased susceptibility to Pseudomonas aeruginosa keratitis. Investig. Ophthalmol. Vis. Sci. 2007, 48, 4498–4508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, M.; McClellan, S.A.; Barrett, R.P.; Zhang, Y.; Hazlett, L.D. β-Defensins 2 and 3 Together Promote Resistance to Pseudomonas aeruginosa Keratitis. J. Immunol. 2009, 183, 8054–8060. [Google Scholar] [CrossRef] [Green Version]
- Dua, H.S.; Otri, A.M.; Hopkinson, A.; Mohammed, I. In vitro studies on the antimicrobial peptide human beta-defensin 9 (HBD9): Signalling pathways and pathogen-related response (an American ophthalmological society thesis). Trans. Am. Ophthalmol. Soc. 2014, 112, 50–73. [Google Scholar]
- Mohammed, I.; Said, D.G.; Dua, H.S. Human antimicrobial peptides in ocular surface defense. Prog. Retin. Eye Res. 2017, 61, 1–22. [Google Scholar] [CrossRef]
- Mohammed, I.; Abedin, A.; Tsintzas, K.; Abedin, S.A.; Otri, A.M.; Hopkinson, A.; Mathew, M.; Dua, H.S. Increased expression of hepcidin and toll-like receptors 8 and 10 in viral keratitis. Cornea 2011, 30, 899–904. [Google Scholar] [CrossRef]
- Mohammed, I.; Mohanty, D.; Said, D.G.; Barik, M.R.; Reddy, M.M.; Alsaadi, A.; Das, S.; Dua, H.S.; Mittal, R. Antimicrobial peptides in human corneal tissue of patients with fungal keratitis. Br. J. Ophthalmol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Takeda, K.; Kaisho, T.; Akira, S. Toll-like receptors. Annu. Rev. Immunol. 2003, 21, 335–376. [Google Scholar] [CrossRef] [PubMed]
- Redfern, R.L.; Reins, R.Y.; McDermott, A.M. Toll-like receptor activation modulates antimicrobial peptide expression by ocular surface cells. Exp. Eye Res. 2011, 92, 209–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Shen, J.B.; Beuerman, R.W. Expression of toll-like receptors in human limbal and conjunctival epithelial cells. Mol. Vis. 2007, 13, 813–822. [Google Scholar] [PubMed]
- McNamara, N.A.; Van, R.; Tuchin, O.S.; Fleiszig, S.M.J. Ocular surface epithelia express mRNA for human beta defensin-2. Exp. Eye Res. 1999, 69, 483–490. [Google Scholar] [CrossRef]
- Kumar, A.; Tassopoulos, A.M.; Li, Q.; Yu, F.S.X. Staphylococcus aureus protein A induced inflammatory response in human corneal epithelial cells. Biochem. Biophys. Res. Commun. 2007, 354, 955–961. [Google Scholar] [CrossRef] [Green Version]
- Kolar, S.S.; Baidouri, H.; McDermott, A.M. Role of pattern recognition receptors in the modulation of antimicrobial peptide expression in the corneal epithelial innate response to F. solani. Investig. Ophthalmol. Vis. Sci. 2017, 58, 2463–2472. [Google Scholar] [CrossRef] [Green Version]
- Hou, A.; Tin, M.Q.; Tong, L. Toll-like receptor 2-mediated NF-kappa B pathway activation in ocular surface epithelial cells. Eye Vis. 2017, 4, 17. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Xu, K.; Ambati, B.; Yu, F.S.X. Toll-like receptor 5-mediated corneal epithelial inflammatory responses to Pseudomonas aeruginosa flagellin. Investig. Ophthalmol. Vis. Sci. 2003, 44, 4247–4254. [Google Scholar] [CrossRef] [Green Version]
- Roy, S.; Marla, S.; Praneetha, D.C. Recognition of Corynebacterium pseudodiphtheriticum by toll-like receptors and up-regulation of antimicrobial peptides in human corneal epithelial cells. Virulence 2015, 6, 716–721. [Google Scholar] [CrossRef] [Green Version]
- Talreja, D.; Singh, P.K.; Kumar, A. In vivo role of TLR2 and MyD88 signaling in eliciting innate immune responses in staphylococcal endophthalmitis. Investig. Ophthalmol. Vis. Sci. 2015, 56, 1719–1732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, A.; Zhang, J.; Yu, F.S.X. Innate immune response of corneal epithelial cells to Staphylococcus aureus infection: Role of peptidoglycan in stimulating proinflammatory cytokine secretion. Investig. Ophthalmol. Vis. Sci. 2004, 45, 3513–3522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, P.I.; Abraham, T.A.; Park, Y.; Zivony, A.S.; Harten, B.; Edelhauser, H.F.; Ward, S.L.; Armstrong, C.A.; Ansel, J.C. The expression of functional LPS receptor proteins CD14 and toll-like receptor 4 in human corneal cells. Investig. Ophthalmol. Vis. Sci. 2001, 42, 2867–2877. [Google Scholar]
- Park, B.S.; Lee, J.O. Recognition of lipopolysaccharide pattern by TLR4 complexes. Exp. Mol. Med. 2013, 45, e66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lang, L.L.; Wang, L.; Liu, L. Exogenous MD-2 confers lipopolysaccharide responsiveness to human corneal epithelial cells with intracellular expression of TLR4 and CD14. Inflammation 2011, 34, 371–378. [Google Scholar] [CrossRef]
- Roy, S.; Sun, Y.; Pearlman, E. Interferon-γ-induced MD-2 protein expression and lipopolysaccharide (LPS) responsiveness in corneal epithelial cells is mediated by Janus tyrosine kinase-2 activation and direct binding of STAT1 protein to the MD-2 promoter. J. Biol. Chem. 2011, 286, 23753–23762. [Google Scholar] [CrossRef] [Green Version]
- Blais, D.R.; Vascotto, S.G.; Griffith, M.; Altosaar, I. LBP and CD14 secreted in tears by the lacrimal glands modulate the LPS response of corneal epithelial cells. Investig. Ophthalmol. Vis. Sci. 2005, 46, 4235–4244. [Google Scholar] [CrossRef]
- Huff, T.; Müller, C.S.G.; Otto, A.M.; Netzker, R.; Hannappel, E. Β-Thymosins, Small Acidic Peptides with Multiple Functions. Int. J. Biochem. Cell Biol. 2001, 33, 205–220. [Google Scholar] [CrossRef]
- Hannapel, E.; van Kampen, M. Determination of thymosin β4 in human blood cells and serum. J. Chromatogr. A 1987, 397, 279–285. [Google Scholar] [CrossRef]
- Goldstein, A.L.; Hannappel, E.; Sosne, G.; Kleinman, H.K. Thymosin β4: A multi-functional regenerative peptide. Basic properties and clinical applications. Expert Opin. Biol. Ther. 2012, 12, 37–51. [Google Scholar] [CrossRef]
- Sosne, G.; Hafeez, S.; Greenberry, A.L.; Kurpakus-Wheater, M. Thymosin β4 promotes human conjunctival epithelial cell migration. Curr. Eye Res. 2002, 24, 268–273. [Google Scholar] [CrossRef] [PubMed]
- Malinda, K.M.; Sidhu, G.S.; Mani, H.; Banaudha, K.; Maheshwari, R.K.; Goldstein, A.L.; Kleinman, H.K. Thymosin beta4 Accelerates Wound Healing. J. Investig. Dermatol. 1999, 113, 364–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sosne, G.; Chan, C.C.; Thai, K.; Kennedy, M.; Szliter, E.A.; Hazlett, L.D.; Kleinman, H.K.; Sosne, G.; Chan, C.C.; Thai, K.; et al. Thymosin beta 4 promotes corneal wound healing and modulates inflammatory mediators in vivo. Exp. Eye Res. 2001, 72, 605–608. [Google Scholar] [CrossRef] [PubMed]
- Sosne, G.; Xu, L.; Prach, L.; Mrock, L.K.; Kleinman, H.K.; Letterio, J.J.; Hazlett, L.D.; Kurpakus-Wheater, M. Thymosin beta 4 stimulates laminin-5 production independent of TGF-beta. Exp. Cell Res. 2004, 293, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.M.; Kang, S.W.; Sung, J.; Kim, K.; Kleinman, H.K. Purinergic Signaling Involvement in Thymosin β4-mediated Corneal Epithelial Cell Migration. Curr. Eye Res. 2020, 45, 1352–1358. [Google Scholar] [CrossRef]
- Sosne, G.; Siddiqi, A.; Kurpakus-Wheater, M. Thymosin-β4 inhibits corneal epithelial cell apoptosis after ethanol exposure in vitro. Investig. Ophthalmol. Vis. Sci. 2004, 45, 1095–1100. [Google Scholar] [CrossRef] [Green Version]
- Sosne, G.; Albeiruti, A.R.; Hollis, B.; Siddiqi, A.; Ellenberg, D.; Kurpakus-Wheater, M. Thymosin β4 inhibits benzalkonium chloride-mediated apoptosis in corneal and conjunctival epithelial cells in vitro. Exp. Eye Res. 2006, 83, 502–507. [Google Scholar] [CrossRef]
- Pahl, H.L. Activators and target genes of Rel/NF-κB transcription factors. Oncogene 1999, 18, 6853–6866. [Google Scholar] [CrossRef] [Green Version]
- Perkins, N.D. Integrating cell-signalling pathways with NF-κB and IKK function. Nat. Rev. Mol. Cell Biol. 2007, 8, 49–62. [Google Scholar] [CrossRef]
- Qiu, P.; Wheater, M.K.; Qiu, Y.; Sosne, G. Thymosin β 4 inhibits TNF-α-induced NF-κB activation, IL-8 expression, and the sensitizing effects by its partners PINCH-1 and ILK. FASEB J. 2011, 25, 1815–1826. [Google Scholar] [CrossRef] [Green Version]
- Sosne, G.; Dunn, S.P.; Kim, C. Thymosin β4 significantly improves signs and symptoms of severe dry eye in a phase 2 randomized trial. Cornea 2015, 34, 491–496. [Google Scholar] [CrossRef] [PubMed]
- Sosne, G.; Ousler, G.W. Thymosin beta 4 ophthalmic solution for dry eye: A randomized, placebo-controlled, phase II clinical trial conducted using the controlled adverse environment (CAETM) model. Clin. Ophthalmol. 2015, 9, 877–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- RegeneRx Joint Venture Expects to Complete Phase 3 Dry Eye Trial in November 2020. 2020. Available online: http://www.regenerx.com/2020-08-31-RegeneRx-Joint-Venture-Expects-to-Complete-Phase-3-Dry-Eye-Trial-in-November-2020 (accessed on 15 September 2020).
- Jin, R.; Li, Y.; Li, L.; Kim, D.; Yang, C.D.; Son, H.S.; Choi, J.H.; Yoon, H.J.; Yoon, K.C. Anti-inflammatory effects of glycine thymosin β4 eye drops in experimental dry eye. Biomed. Rep. 2020, 12, 319–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres, P.; Castro, M.; Reyes, M.; Torres, V.A. Histatins, wound healing, and cell migration. Oral Dis. 2018, 24, 1150–1160. [Google Scholar] [CrossRef] [Green Version]
- Oudhoff, M.J.; Bolscher, J.G.M.; Nazmi, K.; Kalay, H.; Hof, W.; Amerongen, A.V.N.; Veerman, E.C.I. Histatins are the major wound-closure stimulating factors in human saliva as identified in a cell culture assay. FASEB J. 2008, 22, 3805–3812. [Google Scholar] [CrossRef] [Green Version]
- Shah, D.; Ali, M.; Pasha, Z.; Jaboori, A.J.; Jassim, S.H.; Jain, S.; Aakalu, V.K. Histatin-1 expression in human lacrimal epithelium. PLoS ONE 2016, 11, e0148018. [Google Scholar] [CrossRef] [Green Version]
- Oydanich, M.; Epstein, S.P.; Gadaria-Rathod, N.; Guers, J.J.; Fernandez, K.B.; Asbell, P.A. In Vivo Efficacy of Histatin-1 in a Rabbit Animal Model. Curr. Eye Res. 2018, 43, 1215–1220. [Google Scholar] [CrossRef]
- Srinivasan, S.; Thangavelu, M.; Zhang, L.; Green, K.B.; Nichols, K.K. iTRAQ quantitative proteomics in the analysis of tears in dry eye patients. Investig. Ophthalmol. Vis. Sci. 2012, 53, 5052–5059. [Google Scholar] [CrossRef] [Green Version]
- Ananthi, S.; Venkatesh Prajna, N.; Lalitha, P.; Valarnila, M.; Dharmalingam, K. Pathogen Induced Changes in the Protein Profile of Human Tears from Fusarium Keratitis Patients. PLoS ONE 2013, 8, e53018. [Google Scholar] [CrossRef] [Green Version]
- Sanghi, S.; Kumar, R.; Lumsden, A.; Dickinson, D.; Klepeis, V.; Trinkaus-Randall, V.; Frierson, H.F.; Laurie, G.W. cDNA and genomic cloning of lacritin, a novel secretion enhancing factor from the human lacrimal gland. J. Mol. Biol. 2001, 310, 127–139. [Google Scholar] [CrossRef] [Green Version]
- Vantaku, V.R.; Gupta, G.; Rapalli, K.C.; Karnati, R. Lacritin Salvages Human Corneal Epithelial Cells from Lipopolysaccharide Induced Cell Death. Sci. Rep. 2015, 5, 18362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, N.; Zimmerman, K.; Raab, R.W.; Mckown, R.L.; Hutnik, C.M.L.; Talla, V.; Tyler, M.F.; Lee, J.K.; Laurie, G.W. Lacritin rescues stressed epithelia via rapid forkhead box O3 (FOXO3)-associated autophagy that restores metabolism. J. Biol. Chem. 2013, 288, 18146–18161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Wang, N.; Xie, J.; Walton, S.C.; McKown, R.L.; Raab, R.W.; Ma, P.; Beck, S.L.; Coffman, G.L.; Hussaini, I.M.; et al. Restricted epithelial proliferation by lacritin via PKCα-dependent NFAT and mTOR pathways. J. Cell Biol. 2006, 174, 689–700. [Google Scholar] [CrossRef]
- Wang, W.; Despanie, J.; Shi, P.; Edman, M.C.; Lin, Y.A.; Cui, H.; Heur, M.; Fini, M.E.; Hamm-Alvarez, S.F.; Mackay, J.A. Lacritin-mediated regeneration of the corneal epithelia by protein polymer nanoparticles. J. Mater. Chem. B 2014, 2, 8131–8141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, P.; Beck, S.L.; Raab, R.W.; McKown, R.L.; Coffman, G.L.; Utani, A.; Chirico, W.J.; Rapraeger, A.C.; Laurie, G.W. Heparanase deglycanation of syndecan-1 is required for binding of the epithelial-restricted prosecretory mitogen lacritin. J. Cell Biol. 2006, 174, 1097–1106. [Google Scholar] [CrossRef]
- McKown, R.L.; Coleman Frazier, E.V.; Zadrozny, K.K.; Deleault, A.M.; Raab, R.W.; Ryan, D.S.; Sia, R.K.; Lee, J.K.; Laurie, G.W. A cleavage-potentiated fragment of tear lacritin is bactericidal. J. Biol. Chem. 2014, 289, 22172–22182. [Google Scholar] [CrossRef] [Green Version]
- Francisco, V.V.; Romano, J.A.; McKown, R.L.; Green, K.; Zhang, L.; Raab, R.W.; Ryan, D.S.; Hutnik, C.M.L.; Frierson, H.F.; Laurie, G.W. Tissue transglutaminase is a negative regulator of monomeric lacritin bioactivity. Investig. Ophthalmol. Vis. Sci. 2013, 54, 2123–2132. [Google Scholar] [CrossRef]
- Aragona, P.; Aguennouz, M.; Rania, L.; Postorino, E.; Sommario, M.S.; Roszkowska, A.M.; De Pasquale, M.G.; Pisani, A.; Puzzolo, D. Matrix metalloproteinase 9 and transglutaminase 2 expression at the ocular surface in patients with different forms of dry eye disease. Ophthalmology 2015, 122, 62–71. [Google Scholar] [CrossRef]
- Dias-Teixeira, K.; Horton, X.; McKown, R.; Romano, J.; Laurie, G.W. The Lacritin-Syndecan-1-Heparanase Axis in Dry Eye Disease. In Heparanase: From Basic Research to Clinical Applications; Vlodavsky, I., Sanderson, R.D., Ilan, N., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 747–757. ISBN 978-3-030-34521-1. [Google Scholar]
- Li, B.; Sheng, M.; Li, J.; Yan, G.; Lin, A.; Li, M.; Wang, W.; Chen, Y. Tear proteomic analysis of Sjögren syndrome patients with dry eye syndrome by two-dimensional-nano-liquid chromatography coupled with tandem mass spectrometry. Sci. Rep. 2014, 4, 5772. [Google Scholar] [CrossRef] [Green Version]
- McNamara, N.A.; Ge, S.; Lee, S.M.; Enghauser, A.M.; Kuehl, L.; Chen, F.Y.T.; Gallup, M.; McKown, R.L. Reduced levels of tear lacritin are associated with corneal neuropathy in patients with the ocular component of Sjögren’s syndrome. Investig. Ophthalmol. Vis. Sci. 2016, 57, 5237–5243. [Google Scholar] [CrossRef]
- Vijmasi, T.; Chen, F.Y.T.; Balasubbu, S.; Gallup, M.; McKown, R.L.; Laurie, G.W.; McNamara, N.A. Topical administration of lacritin is a novel therapy for aqueous-deficient dry eye disease. Investig. Ophthalmol. Vis. Sci. 2014, 55, 5401–5409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, F.D.; Hao, Z.Q.; Gao, W.; Xing, Y.Q. Effect of topical 0.05% cyclosporine a on the tear protein lacritin in a rat model of dry eye. Int. J. Ophthalmol. 2019, 12, 189–193. [Google Scholar] [CrossRef] [PubMed]
- Georgiev, G.A.; Gh, M.S.; Romano, J.; Teixeira, K.L.D.; Ryan, D.S.; Sia, R.K.; Kitt, J.P.; Harris, J.M.; Hsu, K.; Libby, A.; et al. Lacritin proteoforms prevent tear film collapse and maintain epithelial homeostasis. J. Biol. Chem. 2020. [Google Scholar] [CrossRef] [PubMed]
- Lacripep™ in Subjects with Dry Eye Associated with Primary Sjögren’s Syndrome. Available online: https://clinicaltrials.gov/ct2/show/study/NCT03226444 (accessed on 15 September 2020).
- Inatomi, T.; Spurr-Michaud, S.; Tisdale, A.S.; Gipson, I.K. Human corneal and conjunctival epithelia express MUC1 mucin. Investig. Ophthalmol. Vis. Sci. 1995, 145, 230–234. [Google Scholar]
- Pflugfelder, S.C.; Liu, Z.; Monroy, D.; Li, D.Q.; Carvajal, M.E.; Price-Schiavi, S.A.; Idris, N.; Solomon, A.; Perez, A.; Carraway, K.L. Detection of sialomucin complex (MUC4) in human ocular surface epithelium and tear fluid. Investig. Ophthalmol. Vis. Sci. 2000, 41, 1316–1326. [Google Scholar]
- Argüeso, P.; Spurr-Michaud, S.; Russo, C.L.; Tisdale, A.; Gipson, I.K. MUC16 mucin is expressed by the human ocular surface epithelia and carries the H185 carbohydrate epitope. Investig. Ophthalmol. Vis. Sci. 2003, 44, 2487–2495. [Google Scholar] [CrossRef] [Green Version]
- Yu, D.F.; Chen, Y.; Han, J.M.; Zhang, H.; Chen, X.P.; Zou, W.J.; Liang, L.Y.; Xu, C.C.; Liu, Z.G. MUC19 expression in human ocular surface and lacrimal gland and its alteration in Sjögren syndrome patients. Exp. Eye Res. 2008, 86, 403–411. [Google Scholar] [CrossRef]
- Menon, B.B.; Kaiser-Marko, C.; Spurr-Michaud, S.; Tisdale, A.S.; Gipson, I.K. Suppression of Toll-like receptor-mediated innate immune responses at the ocular surface by the membrane-associated mucins MUC1 and MUC16. Mucosal Immunol. 2015, 8, 1000–1008. [Google Scholar] [CrossRef] [Green Version]
- Uchino, Y.; Uchino, M.; Yokoi, N.; Dogru, M.; Kawashima, M.; Okada, N.; Inaba, T.; Tamaki, S.; Komuro, A.; Sonomura, Y.; et al. Alteration of tear mucin 5AC in office workers using visual display terminals: The Osaka study. JAMA Ophthalmol. 2014, 132, 985–992. [Google Scholar] [CrossRef]
- Corrales, R.M.; Narayanan, S.; Fernández, I.; Mayo, A.; Galarreta, D.J.; Fuentes-Páez, G.; Chaves, F.J.; Herreras, J.M.; Calonge, M. Ocular mucin gene expression levels as biomarkers for the diagnosis of dry eye syndrome. Investig. Ophthalmol. Vis. Sci. 2011, 52, 8363–8369. [Google Scholar] [CrossRef]
- Shimazaki-Den, S.; Dogru, M.; Higa, K.; Shimazaki, J. Symptoms, visual function, and mucin expression of eyes with tear film instability. Cornea 2013, 32, 1211–1218. [Google Scholar] [CrossRef]
- Argüeso, P.; Balaram, M.; Spurr-Michaud, S.; Keutmann, H.T.; Dana, M.R.; Gipson, I.K. Decreased levels of the goblet cell mucin MUC5AC in tears of patients with Sjögren syndrome. Investig. Ophthalmol. Vis. Sci. 2002, 43, 1004–1011. [Google Scholar]
- Caffery, B.; Heynen, M.L.; Joyce, E.; Jones, L.; Robert, R.; Senchyna, M. MUC1 expression in Sjogren’s syndrome, KCS, and control subjects. Mol. Vis. 2010, 16, 1720–1727. [Google Scholar]
- Caffery, B.; Joyce, E.; Heynen, M.L.; Jones, L.; Ritter, R.; Garnache, D.A.; Senchyna, M. MUC16 expression in Sjogren’s syndrome, KCS, and control subjects. Mol. Vis. 2008, 14, 2547–2555. [Google Scholar] [PubMed]
- Argüeso, P.; Guzman-Aranguez, A.; Mantelli, F.; Cao, Z.; Ricciuto, J.; Panjwani, N. Association of cell surface mucins with galectin-3 contributes to the ocular surface epithelial barrier. J. Biol. Chem. 2009, 284, 23037–23045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uchino, Y.; Mauris, J.; Woodward, A.M.; Dieckow, J.; Amparo, F.; Dana, R.; Mantelli, F.; Argüeso, P. Alteration of galectin-3 in tears of patients with dry eye disease. Am. J. Ophthalmol. 2015, 159, 1027–1035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Díaz-Alvarez, L.; Ortega, E. The Many Roles of Galectin-3, a Multifaceted Molecule, in Innate Immune Responses against Pathogens. Mediat. Inflamm. 2017, 2017, 9247574. [Google Scholar] [CrossRef] [Green Version]
- Burguillos, M.A.; Svensson, M.; Schulte, T.; Boza-Serrano, A.; Garcia-Quintanilla, A.; Kavanagh, E.; Santiago, M.; Viceconte, N.; Oliva-Martin, M.J.; Osman, A.M.; et al. Microglia-secreted galectin-3 acts as a toll-like receptor 4 ligand and contributes to microglial activation. Cell Rep. 2015, 10, 1626–1638. [Google Scholar] [CrossRef] [Green Version]
- Narayanan, S.; Miller, W.L.; McDermott, A.M. Expression of human β-defensins in conjunctival epithelium: Relevance to dry eye disease. Investig. Ophthalmol. Vis. Sci. 2003, 44, 3795–3801. [Google Scholar] [CrossRef] [Green Version]
- Kawasaki, S.; Kawamoto, S.; Yokoi, N.; Connon, C.; Minesaki, Y.; Kinoshita, S.; Okubo, K. Up-regulated gene expression in the conjunctival epithelium of patients with Sjögren’s syndrome. Exp. Eye Res. 2003, 77, 17–26. [Google Scholar] [CrossRef]
- Zhou, L.; Beuerman, R.W.; Choi, M.C.; Shao, Z.Z.; Xiao, R.L.; Yang, H.; Tong, L.; Liu, S.; Stern, M.E.; Tan, D. Identification of tear fluid biomarkers in dry eye syndrome using iTRAQ quantitative proteomics. J. Proteome Res. 2009, 8, 4889–4905. [Google Scholar] [CrossRef] [PubMed]
- Dunn, S.P.; Heidemann, D.G.; Chow, C.Y.C.; Crockford, D.; Turjman, N.; Angel, J.; Allan, C.B.; Sosne, G. Treatment of chronic nonhealing neurotrophic corneal epithelial defects with thymosin β4. Ann. N. Y. Acad. Sci. 2010, 1194, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Terreni, E.; Burgalassi, S.; Chetoni, P.; Tampucci, S.; Zucchetti, E.; Fais, R.; Ghelardi, E.; Lupetti, A.; Monti, D. Development and characterization of a novel peptide-loaded antimicrobial ocular insert. Biomolecules 2020, 10, 664. [Google Scholar] [CrossRef] [PubMed]
- Dutta, D.; Vijay, A.K.; Kumar, N.; Willcox, M.D.P. Melimine-coated antimicrobial contact lenses reduce microbial keratitis in an animal model. Investig. Ophthalmol. Vis. Sci. 2016, 57, 5616–5624. [Google Scholar] [CrossRef] [Green Version]
- Dutta, D.; Kumar, N.; Willcox, M.D.P. Antimicrobial activity of four cationic peptides immobilised to poly-hydroxyethylmethacrylate. Biofouling 2016, 32, 429–438. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Molecule | Non-Sjögren DED | Sjögren DED | References |
|---|---|---|---|
| Gel-forming mucins | |||
| MUC5AC | Decrease | Decrease | [113,115] |
| MUC19 | - | Decrease | [110] |
| Membrane associated mucins | |||
| MUC1 | Decrease | Increase | [113,116] |
| MUC4 | Decrease | - | [113] |
| MUC16 | Decrease | Increase | [114,117] |
| Galectin-3 | Increase | Increase | [119] |
| Antimicrobial Peptides and Proteins | |||
| hBD-2 | Increase | Increase | [122,123] |
| hBD-9 | Decrease | - | [37] |
| Histatin-1 | Decrease | Decrease | [40] |
| Lacritin | Decrease | Decrease | [89,101] |
| S100 A8 | Increase | [124] | |
| S100 A9 | Increase | [124] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eshac, Y.; Redfern, R.L.; Aakalu, V.K. The Role of Endogenous Antimicrobial Peptides in Modulating Innate Immunity of the Ocular Surface in Dry Eye Diseases. Int. J. Mol. Sci. 2021, 22, 721. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020721
Eshac Y, Redfern RL, Aakalu VK. The Role of Endogenous Antimicrobial Peptides in Modulating Innate Immunity of the Ocular Surface in Dry Eye Diseases. International Journal of Molecular Sciences. 2021; 22(2):721. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020721
Chicago/Turabian StyleEshac, Youssof, Rachel L. Redfern, and Vinay Kumar Aakalu. 2021. "The Role of Endogenous Antimicrobial Peptides in Modulating Innate Immunity of the Ocular Surface in Dry Eye Diseases" International Journal of Molecular Sciences 22, no. 2: 721. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020721