Combined Effects of Methylated Cytosine and Molecular Crowding on the Thermodynamic Stability of DNA Duplexes


Abstract
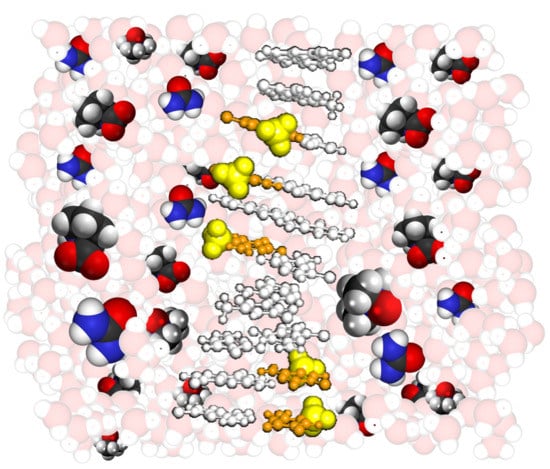
:
1. Introduction
2. Results
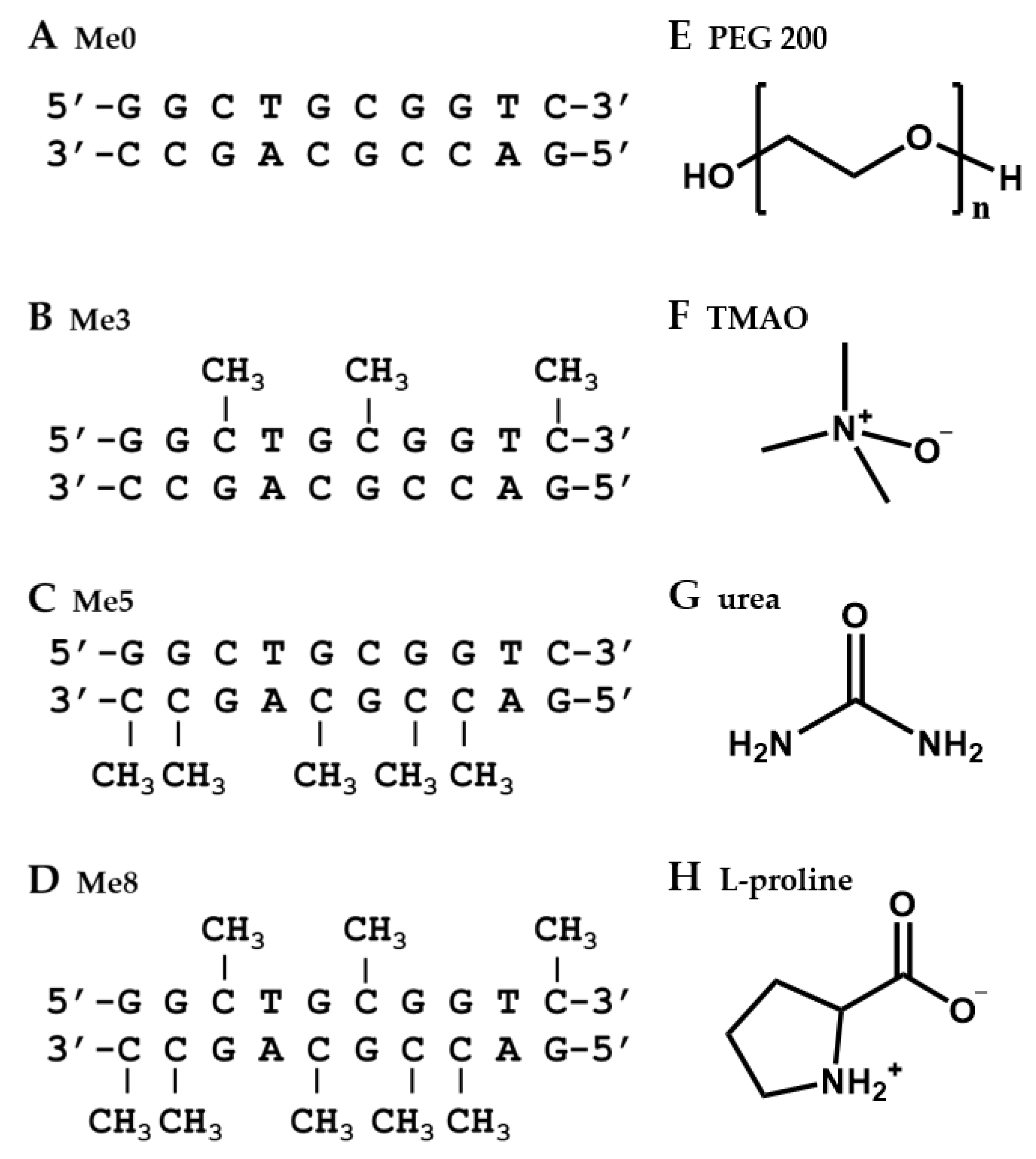
2.1. Design of DNA Sequences and Crowding Reagents
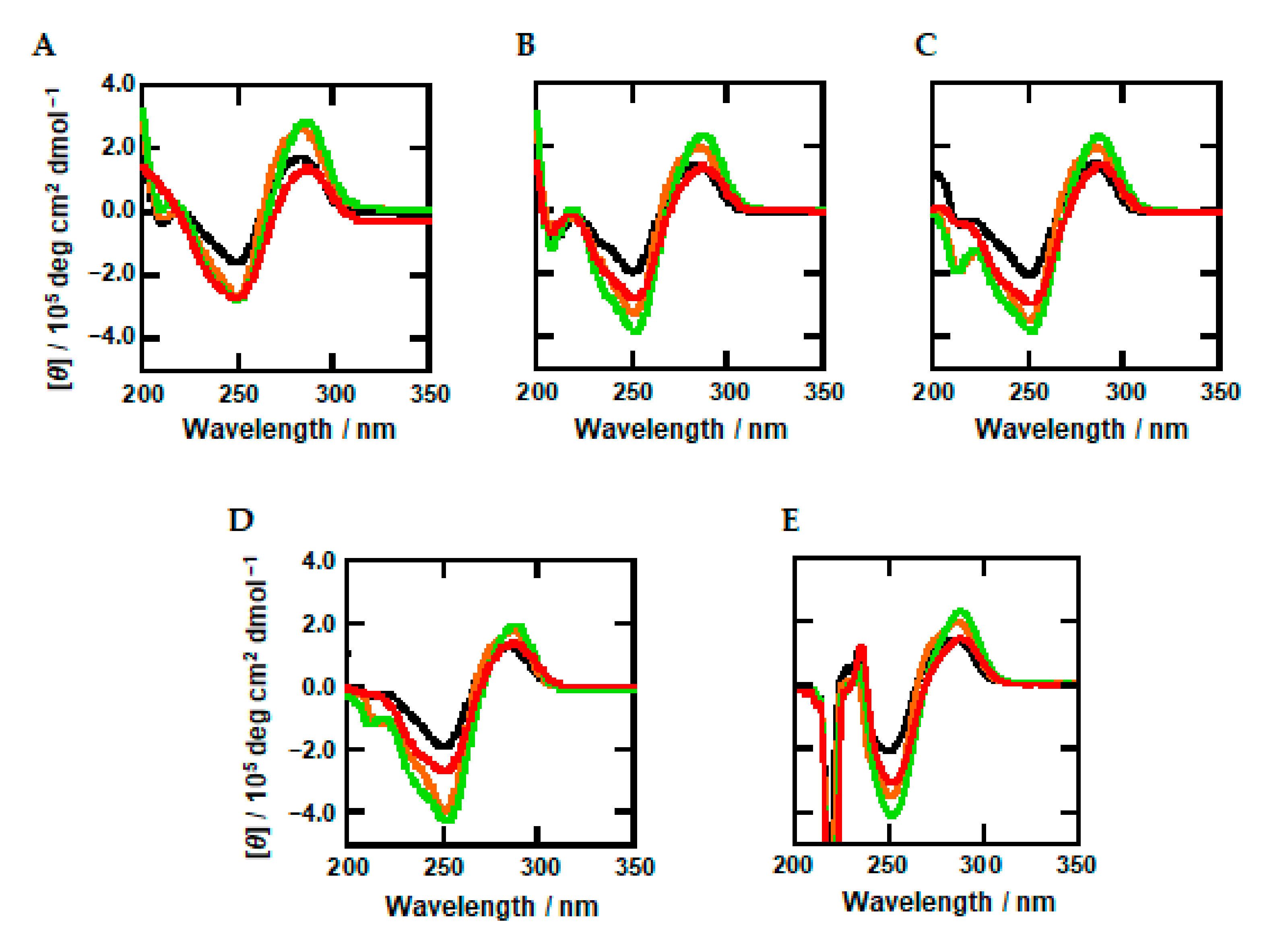
2.2. Effects of Methylation and Molecular Crowding on DNA Structure
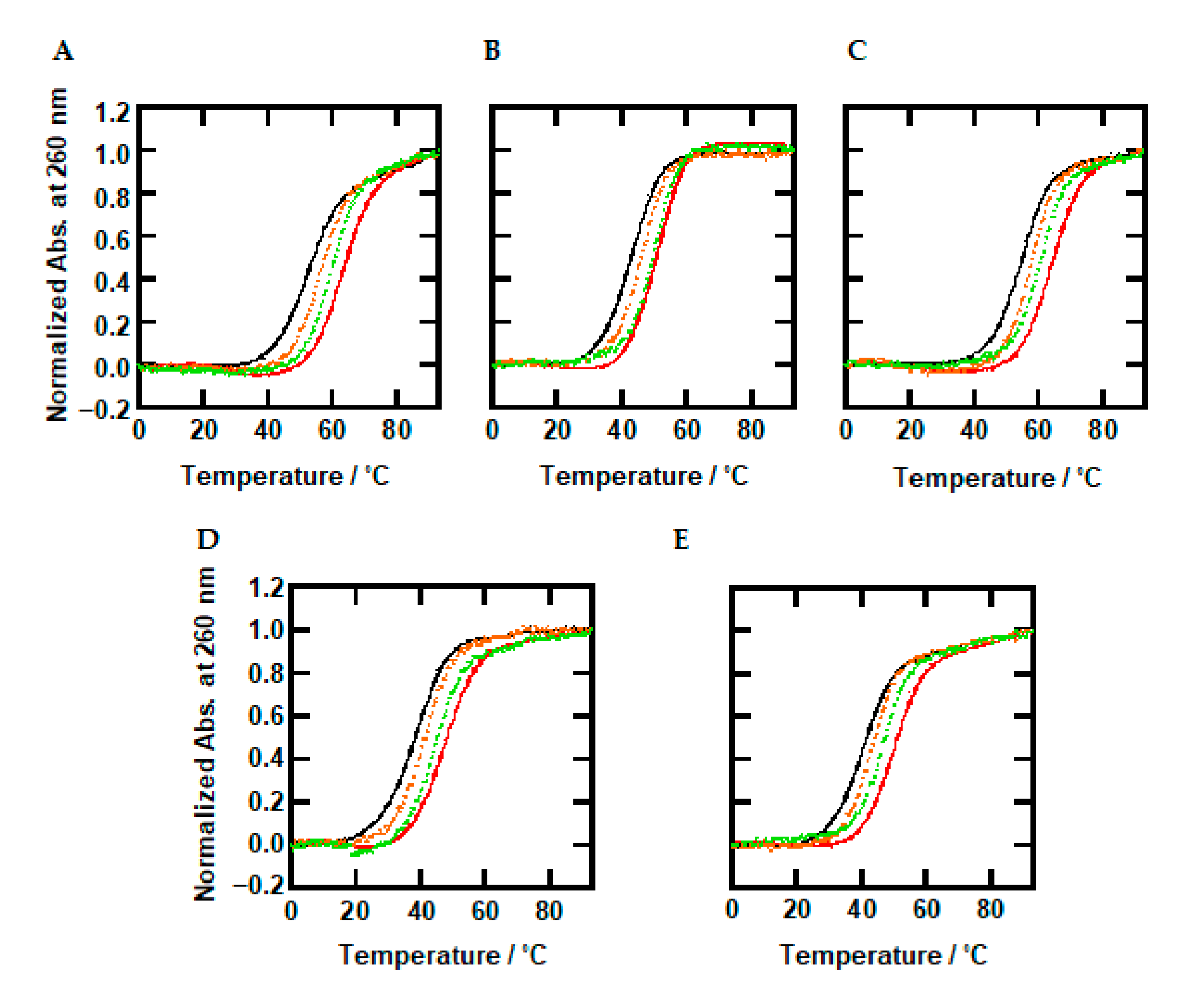
2.3. Thermal Stability of DNA Duplexes with Methylations under Dilute and Molecular Crowding Conditions
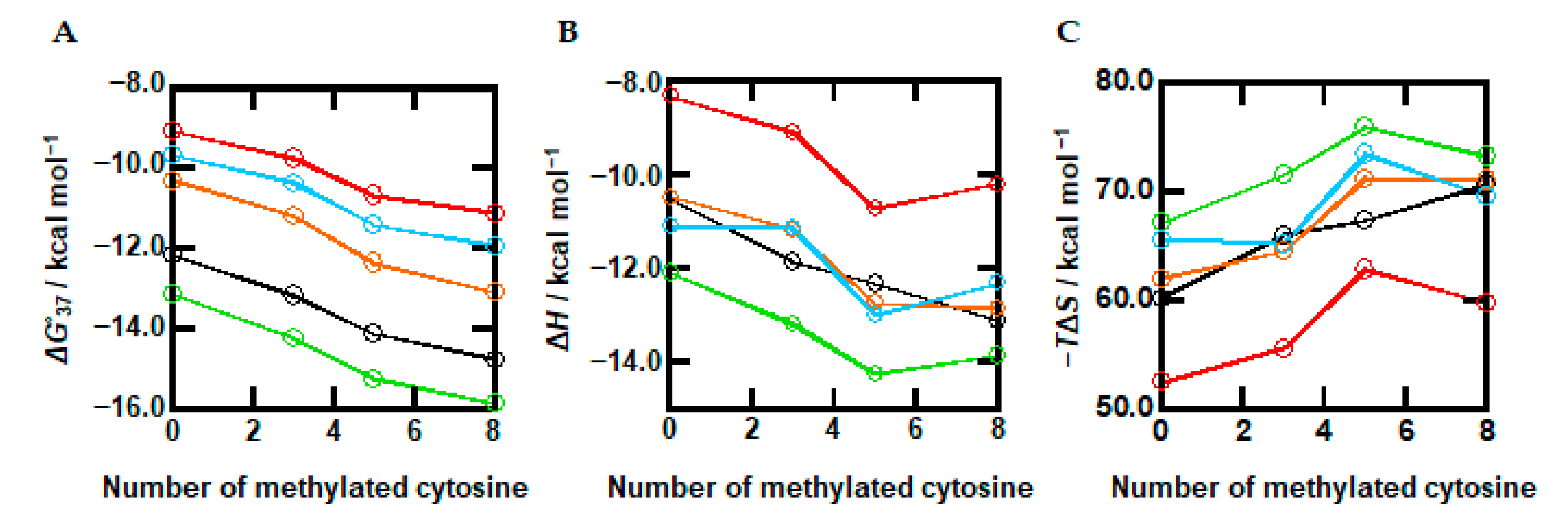
2.4. The Combined Effects of Methylation and Cosolutes on the Thermodynamics of DNA Duplexes
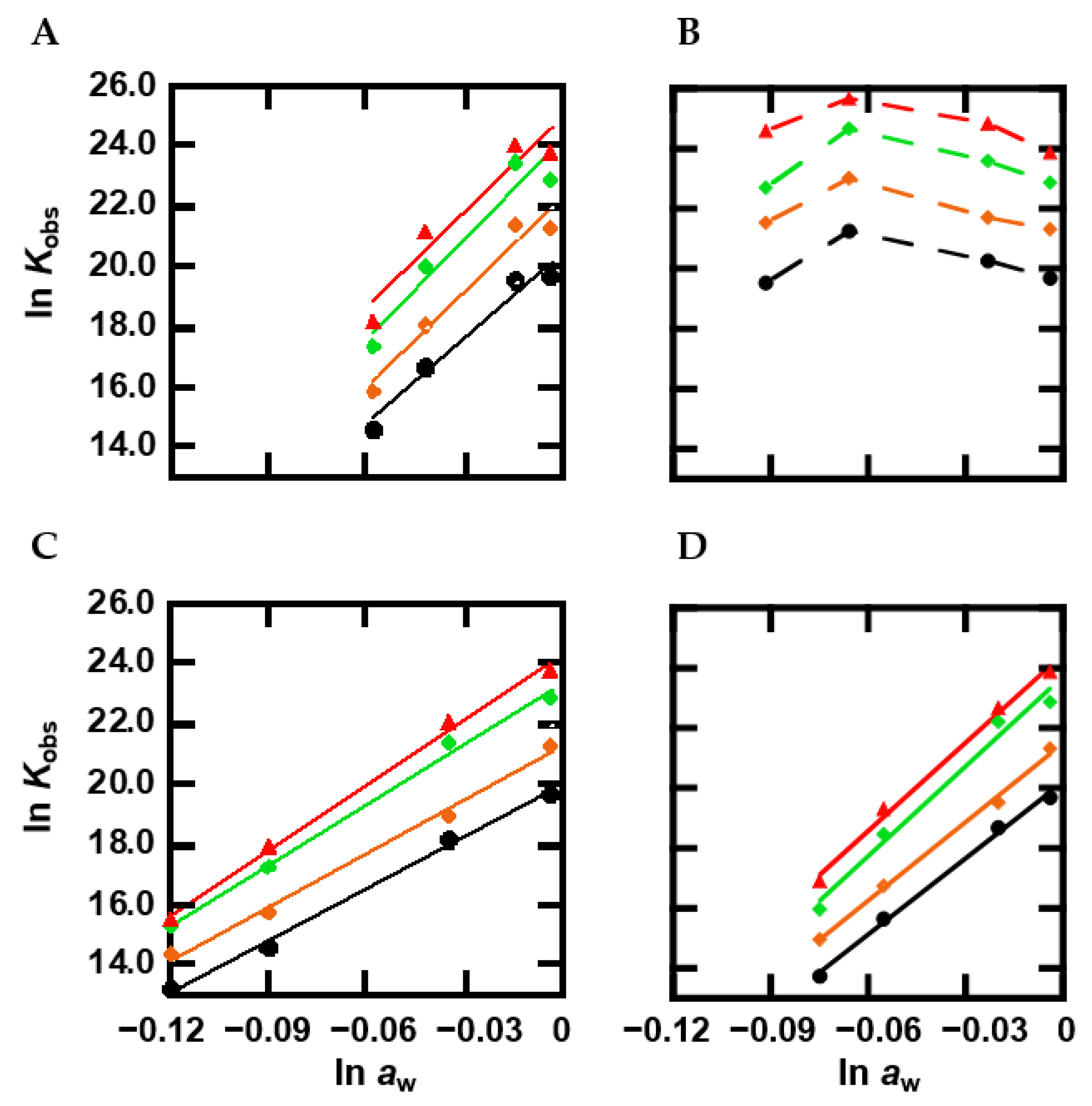
2.5. The Effect of Methylation on Hydration
3. Discussion
3.1. Structural Flexibility of Methylated DNA Duplexes under Molecular Crowding Conditions
3.2. Thermodynamics of DNA Duplexes with Methylations under Molecular Crowding Conditions
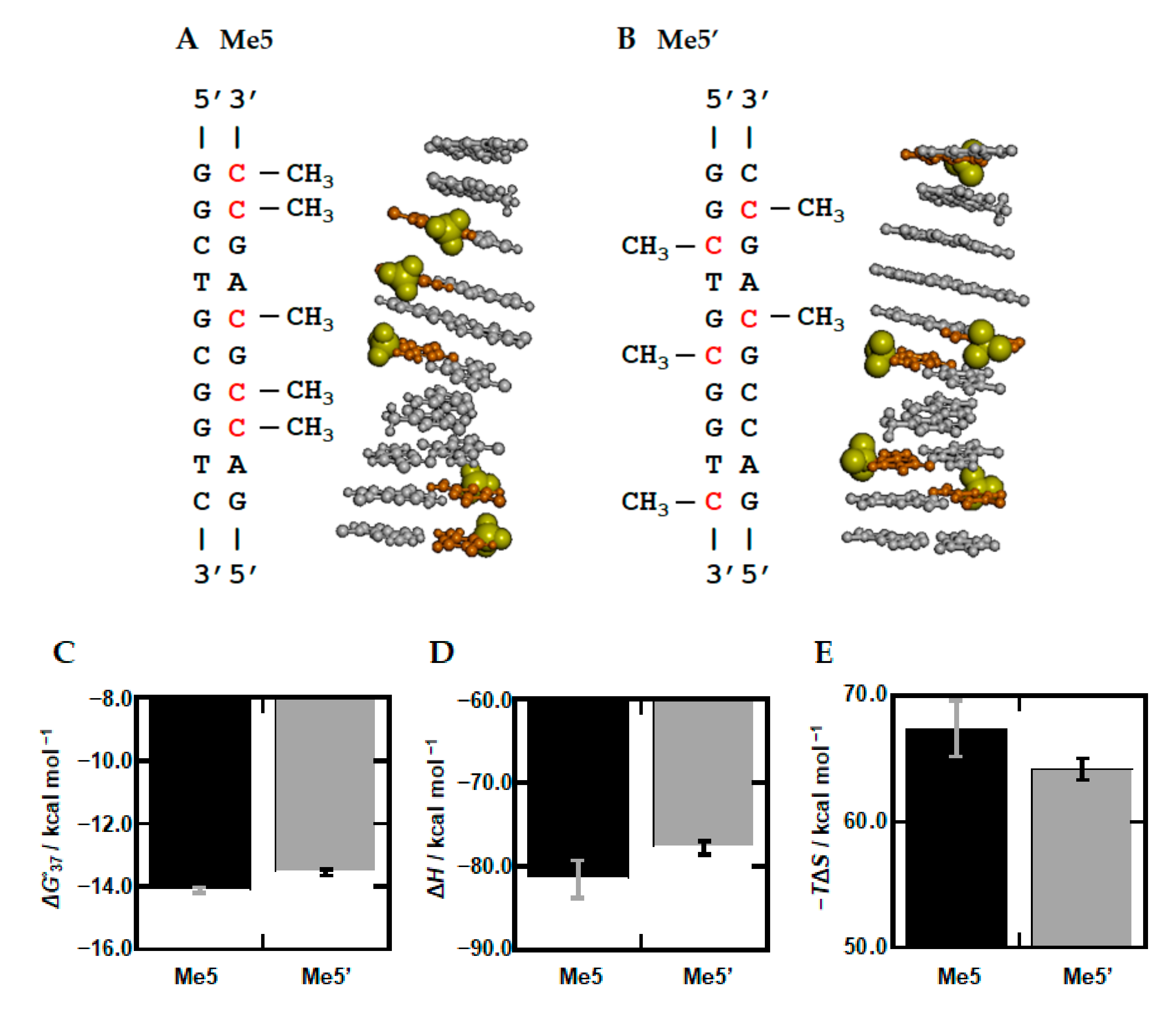
3.3. Stabilization Mechanism of DNA Duplexes by Methylation
3.4. Sequence Dependency of Methylation
4. Materials and Methods
4.1. Materials
4.2. CD Spectroscopy
4.3. Thermal and Thermodynamic Analysis
4.4. Water Activity Measurements
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Holliday, R.; Pugh, J.E. DNA modification mechanisms and gene activity during development. Science 1975, 187, 226–232. [Google Scholar] [CrossRef] [PubMed]
- Smith, Z.D.; Meissner, A. DNA methylation: Roles in mammalian development. Nat. Rev. 2013, 14, 204–220. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Rechkoblit, O.; Bestor, T.H.; Patel, D.J. Structure of DNMT1-DNA complex reveals a role for autoinhibition in maintenance DNA methylation. Science 2011, 331, 1036–1040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehrich, M.; Gama-Sosa, M.A.; Huang, L.-H.; Midgett, R.M.; Kuo, K.C.; Mccune, R.A.; Gehrke, C. Amount and distribution of 5-methylcytosine in human DNA from different types of tissues or cells. Nucleic Acid Res. 1982, 10, 2709–2721. [Google Scholar] [CrossRef] [PubMed]
- Bird, A.P.; Taggart, M.H.; Nicholls, R.D.; Higgs, D.R. Non-methylated CpG-rich islands at the human α-globin locus: Implications for evolution of the α-globin pseudogene. EMBO J. 1987, 6, 999–1004. [Google Scholar] [CrossRef] [PubMed]
- Bird, A.P. DNA methylation patterns and epigenetic memory. Genes Dev. 2002, 16, 6–21. [Google Scholar] [CrossRef] [Green Version]
- Cokus, S.J.; Feng, S.; Zhang, X.; Chen, Z.; Merriman, B.; Haudenschild, C.D.; Pradhan, S.; Nelson, S.F.; Pellegrini, M.; Jacobsen, S.E. Shotgun bisulphite sequencing of the Arabidopsis genome reveals DNA methylation patterning. Nature 2008, 452, 215–219. [Google Scholar] [CrossRef] [Green Version]
- Lister, R.; O’Malley, R.C.; Tonti-Filippini, J.; Gregory, B.D.; Berry, C.C.; Millar, A.H.; Ecker, J.R. Highly integrated single-base resolution maps of the epigenome in Arabidopsis. Cell 2008, 133, 523–536. [Google Scholar] [CrossRef] [Green Version]
- Lister, R.; Pelizzola, M.; Dowen, R.H.; Hawkins, R.D.; Hon, G.; Tonti-Filippini, J.; Joseph, R.N.; Lee, L.; Ye, Z.; Ngo, Q.-M.; et al. Human DNA methylomes at base resolution show widespread epigenomic differences. Nature 2009, 462, 315–322. [Google Scholar] [CrossRef] [Green Version]
- Shirane, K.; Toh, H.; Kobayashi, H.; Kono, T.; Sasaki, H. Mouse oocyte methylomes at base resolution reveal genome-wide accumulation of non-CpG methylation and role of DNA methyltransferases. PLoS Genet. 2013, 9, e1003439. [Google Scholar] [CrossRef] [Green Version]
- Laurent, L.; Wong, E.; Li, G.; Huynh, T.; Tsirigos, A. Dynamic changes in the human methylome during differentiation. Genome Res. 2010, 20, 320–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramsahoye, B.H.; Biniszkiewicz, D.; Lyko, F.; Clark, V.; Bird, A.P.; Jaenisch, R. Non-CpG methylation is prevalent in embryonic stem cells and may be mediated by DNA methyltransferase 3a. Proc. Natl. Acad. Sci. USA 2000, 97, 5237–5242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haines, T.R.; Rodenhiser, D.I.; Ainsworth, P.J. Allele-specific non-CpG methylation of the Nf1 gene during early mouse development. Dev. Biol. 2001, 240, 585–598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lister, R.; Pelizzola, M.; Kida, Y.S.; Hawkins, R.D.; Nery, J.R.; Hon, G.; Antosiewicz-Bourget, J.; O’Mally, R.; Castanon, R.; Klugman, S.; et al. Hotspots of aberrant epigenomic reprogramming in human induced pluripotent stem cells. Nature 2011, 47, 68–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaenisch, R.; Bird, A. Epigenetic regulation of gene expression: How the genome integrates intrinsic and environmental signals. Nat. Genet. 2003, 33, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Riggs, A.D. X inactivation, differentiation, and DNA methylation. Cytogenet. Cell Genet. 1975, 14, 9–25. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.A. Functions of DNA methylation: Islands, start sites, gene bodies and beyond. Nat. Rev. 2012, 13, 484–492. [Google Scholar] [CrossRef]
- Robertson, K.D.; Jones, P.A. DNA methylation: Past, present and future directions. Carcinogenesis 2000, 21, 461–467. [Google Scholar] [CrossRef] [Green Version]
- Dawson, M.A.; Kouzarides, T. Cancer Epigenetics: From Mechanism to Therapy. Cell 2012, 150, 12–27. [Google Scholar] [CrossRef] [Green Version]
- Hwang, J.-Y.; Aromolaran, K.A.; Zukin, R.S. The Emerging Field of Epigenetics in Neurodegeneration and Neuroprotection. Nat. Rev. Neurosci. 2017, 18, 347–361. [Google Scholar] [CrossRef]
- Buck-Koehntop, B.A.; Defossez, P.-A. On how mammalian transcription factors recognize methylated DNA. Epigenetics 2013, 8, 131–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schübeler, D. Function and information content of DNA methylation. Nature 2015, 517, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Luo, C.; Ecker, J.R. Exceptional epigenetics in the brain. Science 2015, 348, 1094–1095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geahigan, K.B.; Meints, G.A.; Hatcher, M.E.; Orban, J.; Drobny, G.P. The Dynamic impact of CpG methylation in DNA. Biochemistry 2000, 39, 4939–4946. [Google Scholar] [CrossRef] [PubMed]
- Lefebvre, A.; Mauffret, O.; Antri, S.E.; Monnot, M.; Lescot, E.; Fermandlian, S. Sequence dependent effects of CpG methylated cytosine A joint 1H-NMR and 31P-NMR study. Eur. J. Biochem. 1995, 229, 445–454. [Google Scholar] [CrossRef]
- Renciuk, D.; Blacpue, O.; Vorlickova, M.; Spingler, B. Crystal structures of B-DNA dodecamer containing the epigenetic modifications 5-hydroxymethylcytosine or 5-methylcytosine. Nucleic Acids Res. 2013, 41, 9891–9900. [Google Scholar] [CrossRef] [Green Version]
- Mayer-Jung, C.; Moras, D.; Timsit, Y. Effect of methylation on DNA-DNA recognition at CpG steps. J. Mol. Biol. 1997, 270, 328–335. [Google Scholar] [CrossRef]
- Hodges-Garcia, Y.; Hagerman, P.J. Methylated cytosine can induce local distortions in the structure of duplex DNA. Biochemistry 1992, 31, 7595–7599. [Google Scholar] [CrossRef]
- Nardo, L.; Lamperti, M.; Salerno, D.; Cassina, V.; Missana, N.; Bondani, M.; Tempestini, A.; Mantegazza, F. Effects of non-CpG site methylation on DNA thermal stability: A fluorescence study. Nucleic Acids Res. 2015, 43, 10722–10733. [Google Scholar] [CrossRef] [Green Version]
- Mayer-Jung, C.; Moras, D.; Timsit, Y. Hydration and recognition of methylated CpG steps in DNA. EMBO J. 1998, 17, 2709–2718. [Google Scholar] [CrossRef] [Green Version]
- Derreumaux, S.; Chaoui, M.; Tevanian, G.; Fermandjian, S. Impact of CpG methylation on structure, dynamics and solvation of cAMP DNA responsive element. Nucleic Acids Res. 2001, 29, 2314–2326. [Google Scholar] [CrossRef] [PubMed]
- Carr, C.E.; Ganugula, R.; Shikiya, R.; Soto, A.M.; Marky, L.A. Effects of dC-d(m5C) substitution on the folding of intramolecular triplexes with mixed TAT and C+GC base triplets. Biochemie 2018, 146, 156–165. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Kool, E.T. Origins of the large differences in stability of DNA and RNA helices: C-5 methyl and 2′-hydroxyl effects. Biochemistry 1995, 34, 4125–4132. [Google Scholar] [CrossRef] [PubMed]
- Zendlova, L.; Hobza, P.; Kabelac, M. Potential energy surfaces of the microhydrated guanine-cytosine base pair and its methylated analogue. ChemPhysChem 2006, 7, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Nakano, S.; Miyoshi, D.; Sugimoto, N. Effects of molecular crowding on the structures, interactions, and functions of nucleic acids. Chem. Rev. 2014, 114, 2733–2758. [Google Scholar] [CrossRef]
- Miyoshi, D.; Karimata, H.; Sugimoto, N. Hydration regulates thermodynamics of G-quadruplex formation under molecular crowding conditions. J. Am. Chem. Soc. 2006, 128, 7957–7963. [Google Scholar] [CrossRef]
- Miyoshi, D.; Sugimoto, N. Molecular crowding effects on structure and stability of DNA. Biochimie 2008, 90, 1040–1051. [Google Scholar] [CrossRef]
- Ueda, Y.; Zouzumi, Y.; Maruyama, A.; Nakano, S.; Sugimoto, N.; Miyoshi, D. Effects of trimethylamine N-oxide and urea on DNA duplex and G-quadruplex. Sci. Technol. Adv. Mater. 2016, 17, 753–759. [Google Scholar] [CrossRef] [Green Version]
- Xu, B.; Devi, G.; Shao, F. Regulation of telomeric i-motif stability by 5-methylcytosine and 5-hydroxymethylcytosine modification. Org. Biomol. Chem. 2015, 13, 5646–5651. [Google Scholar] [CrossRef]
- Spink, C.H.; Chairs, J.B. Effects of Hydration, Ion Release, and Excluded Volume on the Melting of Triplex and Duplex DNA. Biochemistry 1999, 38, 496–508. [Google Scholar] [CrossRef]
- Goobes, R.; Kahana, N.; Cohen, O.; Minsky, A. Metabolic buffering exerted by macromolecular crowding on DNA-DNA interactions: Origin and physiological significance. Biochemistry 2003, 42, 2431–2440. [Google Scholar] [CrossRef] [PubMed]
- Ilia, B.; Bolen, D.W. Forcing Thermodynamically Unfolded Proteins to Fold. J. Biol. Chem. 1998, 273, 4831–4834. [Google Scholar] [CrossRef] [Green Version]
- Miyoshi, D.; Nakao, A.; Sugimoto, N. Molecular Crowding Regulates the Structural Switch of the DNA G-Quadruplex. Biochemistry 2002, 41, 15017–15024. [Google Scholar] [CrossRef]
- Takahashi, S.; Yamamoto, J.; Kitamura, A.; Kinjo, M.; Sugimoto, N. Characterization of intracellular crowding environment with topology-based DNA quadruplex sensors. Anal. Chem. 2019, 91, 2586–2590. [Google Scholar] [CrossRef] [Green Version]
- Qu, Y.; Bolen, C.L.; Bolen, D.W. Osmolyte- driven contraction of a random coil protein. Proc. Natl. Acad. Sci. USA 1998, 16, 9268–9273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, T.Y.; Timasheff, S.N. Why Do Some Organisms Use a Urea-Methylamine Mixture as Osmolyte? Thermodynamic Compensation of Urea and Trimethylamine N-Oxide Interactions with Protein. Biochemistry 1994, 33, 12695–12701. [Google Scholar] [CrossRef] [PubMed]
- Bennion, B.J.; Daggett, V. The molecular basis for the chemical denaturation of proteins by urea. Proc. Natl. Acad. Sci. USA 2003, 9, 5142–5147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canchi, D.R.; Paschek, D.; Garcia, A.E. Equilibrium Study of Protein Denaturation by Urea. J. Am. Chem. Soc. 2010, 132, 2338–2344. [Google Scholar] [CrossRef]
- Guinn, E.J.; Schwinefus, J.J.; Cha, H.K.; McDevitt, J.L.; Merker, W.E.; Ritzer, R.; Muth, S.W.; Engelsgjerd, S.W.; Mangold, K.E.; Thompson, P.J.; et al. Quantifying Functional Group Interactions That Determine Urea Effects on Nucleic Acid Helix Formation. J. Am. Chem. Soc. 2013, 135, 5823–5838. [Google Scholar] [CrossRef] [Green Version]
- Diehl, R.C.; Guinn, E.J.; Capp, M.W.; Tsodikov, O.V.; Record, M.T., Jr. Quantifying additive interactions of the osmolyte proline with individual functional groups of proteins: Comparisons with urea and glycine betaine, interpretation of m-values. Biochemistry 2013, 52, 5997–6010. [Google Scholar] [CrossRef] [Green Version]
- Rajendrakumar, C.S.; Suryanarayana, T.; Reddy, A.R. DNA helix destabilization by proline and betaine: Possible role in the salinity tolerance process. FEBS Lett. 1997, 410, 201–205. [Google Scholar] [CrossRef] [Green Version]
- Auton, M.; Bolen, D.W.; Rösgen, J. Structural thermodynamics of protein preferential solvation: Osmolyte solvation of proteins, aminoacids, and peptides. Proteins 2008, 73, 802–813. [Google Scholar] [CrossRef] [PubMed]
- Riazance, J.H.; Baase, W.A.; Johnson, W.C., Jr.; Hall, K.; Cruz, P.; Tinoco, I., Jr. Evidence for Z-form RNA by vacuum UV circular dichroism. Nucleic. Acids Res. 1985, 13, 4983–4989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thalhammer, A.; Hansen, A.S.; El-Sagheer, A.H.; Brownb, T.; Schofield, C.J. Hydroxylation of methylated CpG dinucleotides reverses stabilisation of DNA duplexes by cytosine 5-methylation. Chem. Commun. 2011, 47, 5325–5327. [Google Scholar] [CrossRef] [PubMed]
- Nakano, S.-I.; Karimata, H.; Ohmichi, T.; Kawakami, J.; Sugimoto, N. The effect of molecular crowding with nucleotide length and cosolute structure on DNA duplex stability. J. Am. Chem. Soc. 2004, 126, 14330–14331. [Google Scholar] [CrossRef] [PubMed]
- Theruvathu, J.A.; Yin, Y.W.; Pettitt, B.M.; Sowers, L.C. Comparison of the structural and dynamic effects of 5-methylcytosine and 5-chlorocytosine in a CpG dinucleotide sequence. Biochemistry 2013, 52, 8590–8598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lercher, L.; McDonough, M.A.; El-Sagheer, A.H.; Thalhammer, A.; Kriaucionis, S.; Brown, T.; Schofield, C.J. Structural insights into how 5-hydroxymethylation influences transcription factor binding. Chem. Commun. 2014, 50, 1794–1796. [Google Scholar] [CrossRef] [Green Version]
- Szulik, M.W.; Pallan, P.S.; Nocek, B.; Voehler, M.; Banerjee, S.; Brooks, S.; Joachimiak, A.; Egli, M.; Eichman, B.F.; Stone, M.P. Differential Stabilities and Sequence-Dependent Base Pair Opening Dynamics of Watson−Crick Base Pairs with 5-Hydroxymethylcytosine, 5-Formylcytosine, or 5-Carboxylcytosine. Biochemistry 2015, 54, 1294–1305. [Google Scholar] [CrossRef] [Green Version]
- Severin, P.M.D.; Zou, X.; Gaub, H.E.; Schulten, K. Cytosine methylation alters DNA mechanical properties. Nucleic Acids Res. 2011, 39, 8740–8751. [Google Scholar] [CrossRef] [Green Version]
- Ngo, T.T.M.; Yoo, J.; Dai, Q.; Zhang, Q.; He, C.; Aksimentiev, A.; Ha, T. Effects of cytosine modifications on DNA flexibility and nucleosome mechanical stability. Nat. Commun. 2016, 7, 10813–10822. [Google Scholar] [CrossRef] [Green Version]
- Nathan, D.; Crothers, D.M. Bending and flexibility of methylated and unmethylated EcoRI DNA. J. Mol. Biol. 2002, 316, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Mirsaidov, U.; Timp, W.; Zou, X.; Dimitrov, V.; Schulten, K.; Feinberg, A.P.; Timp, G. Nanoelectromechanics of methylated DNA in a synthetic nanopore. Biophys. J. 2009, 96, L32–L34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luger, K.; Mader, A.W.; Richmond, R.K.; Sargent, D.F.; Richmond, T.J. Crystal structure of the nucleosome core particle at 2.8 angstrom resolution. Nature 1997, 389, 251–260. [Google Scholar] [CrossRef]
- Churchman, L.S.; Weissman, J.S. Nascent transcript sequencing visualizes transcription at nucleotide resolution. Nature 2011, 469, 368–373. [Google Scholar] [CrossRef] [Green Version]
- Acosta-Silva, C.; Branchadell, V.; Bertran, J.; Oliva, A. Mutual Relationship between Stacking and Hydrogen Bonding in DNA. Theoretical Study of Guanine-Cytosine, Guanine-5-methylcytosine, and Their Dimers. J. Phys. Chem. B 2010, 114, 10217–10227. [Google Scholar] [CrossRef] [PubMed]
- Hognon, C.; Besancenot, V.; Gruez, A.; Grandemange, S.; Monari, A. Cooperative Effects of Cytosine Methylation on DNA Structure and Dynamics. J. Phys. Chem. B 2019, 123, 7365–7371. [Google Scholar] [CrossRef] [PubMed]
- Madugundu, G.S.; Cadet, J.; Wagner, J.R. Hydroxyl-radical-induced oxidation of 5-methylcytosine in isolated and cellular DNA. Nucleic Acids Res. 2014, 42, 7450–7460. [Google Scholar] [CrossRef]
- Plum, G.E.; Park, Y.-W.; Singleton, S.F.; Dervan, P.B.; Breslauer, K.J. Thermodynamic characterization of the stability and the melting behavior of a DNA triplex: A spectroscopic and calorimetric study. Proc. Natl. Acad. Sci. USA 1990, 87, 9436–9440. [Google Scholar] [CrossRef] [Green Version]
- Roberts, R.W.; Crothers, D.M. Stability and properties of double and triple helices: Dramatic effects of RNA or DNA backbone composition. Science 1992, 258, 1463–1466. [Google Scholar] [CrossRef]
- Wu, Q.; Wong, J.R.; Yeo, P.L.Q.; Zhang, D.; Shao, F. Methylation on CpG repeats modulates hydroxymethylcytosine induced duplex destabilization. RSC Adv. 2016, 6, 48858–48862. [Google Scholar] [CrossRef]
- Povsic, T.J.; Dervan, P.B. Triple helix formation by oligonucleotides on DNA extended to the physiological pH range. J. Am. Chem. Soc. 1989, 111, 3059–3061. [Google Scholar] [CrossRef]
- Tretyakova, N.; Guza, R.; Matter, B. Endogenous cytosine methylation and the formation of carcinogen carcinogen–DNA adducts. Nucleic Acids Symp. Ser. 2008, 52, 49–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, V.R.; Thonhauser, T.; Puzder, A.; Schroder, E.; Lundqvist, B.I.; Langreth, D.C. Stacking Interactions and the Twist of DNA. J. Am. Chem. Soc. 2008, 130, 1304–1308. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Gu, X.; Nakano, S.; Miyoshi, D.; Sugimoto, N. Beads-on-a-String Structure of Long Telomeric DNAs under Molecular Crowding Conditions. J. Am. Chem. Soc. 2012, 134, 20060–20069. [Google Scholar] [CrossRef] [PubMed]
- Muhuri, S.; Mimura, K.; Miyoshi, D.; Sugimoto, N. Stabilization of Three-Way Junctions of DNA under Molecular Crowding Conditions. J. Am. Chem. Soc. 2009, 131, 9268–9280. [Google Scholar] [CrossRef]
- Miyoshi, D.; Nakamura, K.; Tateishi-Karimata, H.; Ohmichi, T.; Sugimoto, N. Hydration of Watson-Crick Base Pairs and Dehydration of Hoogsteen Base Pairs Inducing Structural Polymorphism under Molecular Crowding Conditions. J. Am. Chem. Soc. 2009, 131, 3522–3531. [Google Scholar] [CrossRef]
- Miyoshi, D.; Shizuka, M.; Nakano, S.; Sugimoto, N. Duplex Dissociation of Telomere DNAs Induced by Molecular Crowding. J. Am. Chem. Soc. 2004, 126, 165–169. [Google Scholar] [CrossRef]
- Sugimoto, N.; Nakano, S.; Katoh, M. Thermodynamic parameters to predict stability of RNA/DNA hybrid duplexes. Biochemistry 1995, 34, 11211–11216. [Google Scholar] [CrossRef]
- Nakano, S.; Fujimoto, M.; Hara, H. Nucleic acid duplex stability: Influence of base composition on cation effects. Nucleic Acids Res. 1999, 27, 2957–2965. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abbreviation | Cosolutes | ||||
|---|---|---|---|---|---|
| Without Cosolute | PEG200 | TMAO | Urea | L-Proline | |
| Me0 | 51.4 ± 0.52 | 42.8 ± 0.0 | 54.0 ± 0.2 | 37.5 ± 0.0 | 40.0 ± 0.1 |
| Me3 | 54.4 ± 0.5 | 46.4 ± 0.2 | 57.2 ± 0.1 | 40.7 ± 0.1 | 42.8 ± 0.2 |
| Me5 | 58.0 ± 0.5 | 50.1 ± 0.2 | 59.9 ± 0.3 | 44.4 ± 0.2 | 46.2 ± 0.4 |
| Me8 | 59.5 ± 1.8 | 52.9 ± 0.2 | 63.0 ± 0.3 | 46.7 ± 0.2 | 48.7 ± 0.1 |
| Abbreviation | −Δnw | |||
|---|---|---|---|---|
| PEG200 | TMAO 2 | Urea | L-Proline | |
| Me0 | 96 ± 4 | - | 57 ± 1 | 85 ± 3 |
| Me3 | 105 ± 2 | - | 60 ± 2 | 87 ± 1 |
| Me5 | 108 ± 2 | - | 67 ± 1 | 100 ± 3 |
| Me8 | 106 ± 11 | - | 72 ± 3 | 98 ± 8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsuruta, M.; Sugitani, Y.; Sugimoto, N.; Miyoshi, D. Combined Effects of Methylated Cytosine and Molecular Crowding on the Thermodynamic Stability of DNA Duplexes. Int. J. Mol. Sci. 2021, 22, 947. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020947
Tsuruta M, Sugitani Y, Sugimoto N, Miyoshi D. Combined Effects of Methylated Cytosine and Molecular Crowding on the Thermodynamic Stability of DNA Duplexes. International Journal of Molecular Sciences. 2021; 22(2):947. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020947
Chicago/Turabian StyleTsuruta, Mitsuki, Yui Sugitani, Naoki Sugimoto, and Daisuke Miyoshi. 2021. "Combined Effects of Methylated Cytosine and Molecular Crowding on the Thermodynamic Stability of DNA Duplexes" International Journal of Molecular Sciences 22, no. 2: 947. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020947