Plant Mitophagy in Comparison to Mammals: What Is Still Missing?

Centre for Cell and Developmental Biology, State Key Laboratory of Agrobiotechnology, School of Life Sciences, The Chinese University of Hong Kong, Hong Kong, China
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2021, 22(3), 1236; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031236
Submission received: 22 December 2020
/
Revised: 13 January 2021
/
Accepted: 20 January 2021
/
Published: 27 January 2021
(This article belongs to the Special Issue New Insight into Signaling and Autophagy in Plants)
Abstract
:Mitochondrial homeostasis refers to the balance of mitochondrial number and quality in a cell. It is maintained by mitochondrial biogenesis, mitochondrial fusion/fission, and the clearance of unwanted/damaged mitochondria. Mitophagy represents a selective form of autophagy by sequestration of the potentially harmful mitochondrial materials into a double-membrane autophagosome, thus preventing the release of death inducers, which can trigger programmed cell death (PCD). Recent advances have also unveiled a close interconnection between mitophagy and mitochondrial dynamics, as well as PCD in both mammalian and plant cells. In this review, we will summarize and discuss recent findings on the interplay between mitophagy and mitochondrial dynamics, with a focus on the molecular evidence for mitophagy crosstalk with mitochondrial dynamics and PCD.
1. Introduction
The mitochondrion is an essential organelle conserved in eukaryotic organisms, which conducts several key biological functions. The primary function of mitochondria is to manufacture ATP, the major energy resource of the cells, through oxidative respiration reactions [1]. In addition, mitochondria are versatile participants in many catabolic and signaling pathways. For example, studies in animals have established an essential role of mitochondria in fatty acid synthesis for lipoic acid production, mitochondrial translation and mitochondrial oxidative reaction [2]. Another conserved function of mitochondria is to store Ca2+ ions to maintain Ca2+ homeostasis [3]. In plants, recent findings have also identified a crucial role of mitochondria in mediating salicylic acid signaling [4], multiple anti-stress [5] and anti-pathogen responses [6]. In addition, mitochondria are indispensable for programmed cell death (PCD) regulation in both animals [7] and plants [8]. As a highly active factory where redox reactions occur, mitochondria also produce a lot of reactive oxygen species (ROS), such as H2O2, which can damage themselves as well as other organelles [8]. Accumulating ROS in the mitochondrion will ultimately lead to a loss of mitochondrial membrane potential difference (also known as ΔφΜ) and increase its mitochondrial outer membrane permeability (MOMP) [9]. As a result, mitochondrial materials (e.g., cytochrome C), which will activate downstream PCD machinery, are released from the damaged mitochondria [9]. Thus, it is crucial for the cells to surveil the conditions of the mitochondria and to eliminate those damaged/unwanted ones efficiently by regulating mitochondrial dynamics [10].
The overall balance in the quantity and the quality of mitochondria in a cell is referred to as mitochondrial homeostasis maintenance. Mitochondrial fission/fusion and autophagy-selective degradation (also known as mitophagy) represent two major approaches to balance the number and size of mitochondria, or to eliminate the unwanted ones [11,12]. Through the mitochondrial fission/fusion, mitochondria are organized into a huge dynamic network, which increases their anti-stress ability as the mildly damaged mitochondria can be repaired by fusing with healthy ones, while the severely damaged parts of mitochondria can be segregated from the network by fission [13]. On the other hand, the segregated damaged parts of mitochondria can be selectively degraded via mitophagy [14]. During mitophagy, unwanted mitochondrial materials are recognized and enclosed by a double-membrane structure named an autophagosome, which is finally delivered into the lysosome (in animal) or vacuole (in plant and yeast) for degradation [15].
Five core autophagy-related (ATG) complexes are required to aid the formation of an autophagosome, which are largely conserved in plant and animal genomes: (1) ATG1 (ortholog of ULK1 [unc-51 like autophagy activating kinase 1] in animals) kinase complex; (2) ATG9 complex; (3) ATG6 (ortholog of Beclin1 in animals)-VPS15-VPS34 PI3K (phosphatidylinositol 3-kinase) complex; (4) ATG5-ATG12 conjugate; (5) ATG8 (ortholog of LC3 in animals) conjugate [16]. Among them, only ATG8 covalently binds to phosphatidylethanolamine (PE) on the autophagosome membrane, and therefore widely serves as a reporter of autophagosome structures and autophagy activity. The function of the core ATG complexes in autophagosome formation have been extensively summarized in a number of recent reviews, and thus will not be covered here [16,17,18,19].
In recent decades, exciting new findings have unveiled a critical role of mitophagy in plant development. In germinating Arabidopsis seedlings, it has been observed that inactive mitochondria are engulfed by autophagosomes, suggesting that mitophagy facilitates the degradation of mitochondria that fail to recover from dormancy [20]. Recently, it has been also demonstrated that the de-etiolation process is suppressed in atg5 mutant plants during the transition from darkness to light [21]. In addition, intact mitochondria were observed in the vacuole of Arabidopsis under oxidative stress, implying the occurrence of mitophagy in response to stress [15]. Furthermore, mitophagy has also been implicated to participate in PCD regulation and senescence under nutrient deficiency conditions [12,22]. Importantly, recent studies in animal have unveiled a close interconnection between mitophagy and mitochondrial dynamics, as well as PCD, for a balance in mitochondrial quality control mechanisms. Here we will highlight these recent findings to compare with the plant system, with a focus on the interplay among mitophagy regulators, mitochondrial fusion/fission machineries and PCD regulators.
2. Mitophagy: More than One Way to Get Rid of a Mitochondrion
During mitophagy, two key events are involved: (1) the formation of the phagophore, which finally expands and encloses the cargo into the autophagosome; (2) the recognition of the mitochondrial cargo, which is mainly mediated by interaction between the mitochondria cargo receptor/adaptor and ATG8/LC3, which are covalently linked to phosphatidylethanolamine (PE) on the autophagosome membrane [23] (Figure 1). According to the nature of the targeting mitochondrial proteins, mitophagy can be classified into the following categories: (1) ubiquitin-dependent, (2) receptor-dependent and (3) lipid-dependent (Figure 1).
2.1. Ubiquitin-Dependent Mitophagy
In humans, the first pathway identified to mediate mitophagy was the PINK1–Parkin system, which is involved in Parkinson’s disease [47]. When mitochondria lose their ΔφΜ induced by ROS, PTEN-induced kinase 1 (PINK1) kinase proteins accumulate on the outer mitochondrial membrane (OMM) to recruit and activate Parkin [48]. Parkin, which is featured by its RING1-in-between-ring (IBR)-RING2 domains, is a ring-in-between-ring (RBR) E3 ubiquitin ligase to ubiquitinate multiple OMM proteins [49]. Afterwards, some adaptor proteins, such as p62, recognizes the poly-ubiquitinated mitochondrial OMMs and targets them to the autophagosome via binding to LC3 [24]. In addition, emerging evidence has indicated that other E3-ubiquitin ligases may participate in mitophagy. For example, mitochondrial ubiquitin ligase activator of NFKB 1 (MUL1/MAPL/MULAN) functions as an alternative OMM-localized E3 ligase in PARKIN-independent mitophagy [25]. UKL1, as well as dynamin-related protein 1 (DRP1), have both been identified as the substrates of MUL1/MAPL/MULAN via ubiquitination and SUMOylating, respectively [26,27]. Another E3 ligase, SIAH1, forms a complex with Synphilin1, which interacts with PINK1 and is recruited to the OMM, to ubiquitinate mitochondrial proteins in a PINK1-Parkin-independent manner [28]. Moreover, a recent study identified another alternative PRB E3 ligase called ARIH1/HHARI, to mediate the mitophagy pathway together with PINK1 in a Parkin-independent manner [29].
In plants, counterparts of these E3-ligases have not yet been identified to date. Instead, the Arabidopsis ARI family, a putative RBR E3-ligase family, has been suggested to play an essential role under stress conditions, and actively participates in several stress-related hormone signaling pathways via the ubiquitin proteasome system [50]. Recently, an RBR E3-ligase ubiquitin conjugating enzyme 26 (UBC26) has been reported to participate in the phytohormone abscisic acid (ABA) response in Arabidopsis, likely by promoting the degradation of ABA receptors in response to abiotic stress [51]. However, direct evidence for linking RBR E3-ligases in plant mitophagy is still missing. Recently, it was reported that another plant E3 ligase, SP1, a counterpart of mammalian MUL1/MAPL/MULAN, which participates in chloroplast and peroxisome biogenesis, also targets to mitochondria [52]. It might be worthwhile to identify the mitochondrial substrates for these E3 ligases during mitophagy in future to further unveil their possible roles in mitochondrial homeostasis.
2.2. Receptor-Dependent Mitophagy
Despite the ubiquitin-dependent pathway, ATG8/LC3 also directly binds to several OMM receptors which contain the conserved ATG8-interacting motif/LC3-interacting region (AIM/LIR). OMM receptors such as FUN14 domain-containing protein 1 (FUNDC1) [30], BCL2 Interacting Protein 1 (BNIP1) [32] and NIX [31], are phosphorylated and activated upon mitochondrial damage to recruit LC3 or gamma-aminobutyric acid receptor-associated protein, another LC3 isoform (GABARAP) for binding to different mitochondrial OMM proteins respectively. Recently, a novel receptor named FKBP Prolyl Isomerase 8 (FKBP8) has been identified, but it is recycled from the mitochondria once mitophagy is activated [33]. Interestingly, it has been reported that Bcl2-L-13, functions as another mitophagy receptor to recruit both ULK1 and LC3B to regulate mitophagy [39]. Additionally, an OMM receptor activating molecule in BECN1-regulated autophagy protein 1 (AMBRA1), by recruiting Beclin1, induces PINK1/Parkin-dependent mitophagy pathway [34] and the receptor-dependent pathway [35]. On the other hand, some inner mitochondrial membrane (IMM) mitophagy receptors have also been reported. For example, prohibitin 2 (PHB2) was found to be exported to the OMM for directly binding to LC3 to stimulate mitophagy [36]. Differently, another IMM protein, choline dehydrogenase (CHDH), is initially recognized by p62 adaptor and then recruits LC3 [37]. Moreover, it has also been reported that E3 ligase MUL1/MAPL/MULAN contains an LIR-like motif to interact with GABARAP, another LC3 isoform in animals, implying that E3 ligases may alternatively participate in mitochondrial turnover in a receptor-dependent manner as well [38].
In budding yeast, the only identified receptor is ATG32, an OMM-localized protein interacting with both ATG8 and ATG11 [42]. Upon mitophagy induction, casein kinase 2 (CK2) will activate ATG32 via phosphorylation. Subsequently, the phosphorylated ATG32 is recognized by ATG11 [43]. Meanwhile, ATG32 also requires an IMM protease FtsH/Yme1 for its C-terminus processing in order to interact with ATG11 [44]. Although ATG32 contains an AIM motif for binding to ATG8, mutation of ATG32–ATG8 interaction dose not completely block mitophagy [45]. On the other hand, ATG11 binds to Dynamin 1 (DNM1), which functions in mitochondrial fission [53]. However, it remains unknown whether the ATG32–ATG11 complex promotes mitochondrial fission by interacting with DNM1. Differently, in fission yeast, it has been found that another OMM protein, named ATG43, might function as a mitophagy receptor by coordinating with the mitochondrial import factors instead [46].
In plants, ATG11 has also been suggested to function as an adaptor in autophagy-dependent mitochondrial degradation [22]. Arabidopsis ATG11 contains AIM for interacting with ATG8, and its deficiency leads to the accumulation of mitochondria [22]. Recently, by searching for the AIM motif in mitochondrial proteins, 12 OMM proteins and 36 IMM proteins containing the AIM-like motif were predicted [12]. Of note, another recent screening study suggested that the Arabidopsis mitochondrial OMM Ca2+ channel, voltage-dependent anion channel 2 (VDAC2), is also predicted to harbor an AIM [54]. In Arabidopsis, defects in number and size of mitochondria were observed in most vdac mutants, but vdac2 and vdac4 mutant plants displayed a more severe growth retardation phenotype when compared with other vdac mutants, suggesting their possible distinct roles in plant growth [55,56]. In animals, it has been shown that VDAC2 is poly-ubiquitinated by Parkin and involved in mitochondrial quality control [24]. Plant PHBs, which are homologs of the mammalian IMM mitophagy receptor, may function as a candidate receptor in plant mitophagy. Loss of plant PHB3 leads to altered mitochondrial morphology and numbers [57]. A recent study has also reported that the deficiency in PHB3 reduces the levels of Salicylic acid (SA) but increases hypersensitive cell death in response to stress [58]. Nevertheless, the molecular link for plant PHBs in mitochondria or autophagy is still unclear. On the other hand, mutation of FtsH4 protease, which also locates to the IMM, causes mitochondria swelling and ROS accumulation [59]. Another study has further demonstrated that the autophagy-related defects, including leaf senescence caused by FtSH4, are dependent on the core ATG proteins [59]. It would be interesting to identify the mitochondrial substrates that are targeted by FtsH4 and to elucidate their functions in plant mitophagy. Taken together, future studies to explore the function of these OMM/IMM proteins, as well as their relationships with the ATG machinery during mitophagy should provide novel insights into the underlying mechanism of plant mitophagy.
2.3. Lipid-Dependent Mitophagy
Recently, a highly conserved IMM lipid cardiolipin (CL) was reported to recruit LC3 upon mitochondrial damage, suggesting that the lipid might serve as a targeting signal for mitophagy as well [40]. Under normal conditions, cardiolipin mediates IMM fusion together with OPA1 [60], but is exported to the OMM by the phospholipid scramblase-3 (PLS3) upon mitochondrial damage [40]. Another lipid named ceramide (CER), is synthesized into different forms by ceramide synthetase families (CerS) and released to the OMM to induce mitophagy as well [41]. It was observed that these CERs may promote the lipidation of LC3-PE to the OMM, and thus activate autophagy [41]. It was shown that the loss of cardiolipin synthetase in Arabidopsis leads to abnormal mitochondrial morphology, defective growth and reduced anti-stress response [5]. However, whether the lipid products are also exported to regulate mitophagy awaits further investigation in plants.
3. Interplay between Mitophagy and Mitochondrial Dynamics
3.1. Mitochondrial Fusion and Fission Machineries in Mitophagy
Mitochondrial fusion and fission is essential for maintaining the morphology and functions of healthy mitochondria. The following section will discuss different and conserved regulators in mammal and plant mitochondrial fusion/fission, as well as their interconnections with mitophagy (Table 1).
3.1.1. Mitochondrial Fusion Machinery
In mammalian cells, OMM-located GTPases, MFN1/2 (mitofusin1/2), which will form homodimers or heterodimers to facilitate membrane tethering, play an essential role in mediating OMM fusion with adjacent mitochondria [71]. In addition, with other tethering factors, MFN1/2 forms a protein complex named ER-mitochondria encounter structure (ERMES) to mediate the fusion between ER membrane and mitochondria, also known as the ER-mitochondria contact (EMC). It has been observed that autophagosomal structures containing mitochondria were colocalized with ERMES, whereas disturbance of this complex affects the mitophagy level [72]. Thus, it has been suggested that the ER-mitochondria contact might provide membrane source to aid phagophore initiation and mitochondria sequestration. Another tethering complex, comprising mitochondria-localized VDAC1, inositol 1,4,5-triphosphate receptor 3 (IP3R) and ER-localized glucose-regulated protein 75 kDa (GRP75), mainly functions in the regulation of the intracellular Ca2+ homeostasis between ER and mitochondria [73]. VDAC1 has been previously identified as a Parkin substrate and ubiquitination of VDAC1 will further induce mitophagy, but it has also been suggested that other substrates rather than VDAC1 is responsible for the activation [24,74]. Interestingly, a recent study showed that Parkin promotes MFN2 phospho-ubiquitination, followed by MFN2 disassembly from the mitochondrial membrane to disrupt MFN2-mediated EMCs [75]. As a result, Parkin further enhanced ubiquitination VDAC1 and/or other mitochondrial substrates to facilitate the degradation of mitochondria, thus unveiling a novel molecular mechanism by interfering with EMCs via the Parkin-dependent ubiquitination of MFN2. However, FZO-like (FZL) protein, a homolog of MFN in plant, has been reported to function in plant chloroplast biogenesis, and no obvious defects in mitochondrial morphology was observed in fzl mutants [61].
On the other hand, the IMM fusion machinery also plays an essential role to maintain the architecture of mitochondria. Invagination of the IMM produces cristae, which requires optic atrophy 1 (OPA1) and the mitochondrial contact site (MICOS) complex [76]. OPA1 is a mitochondrial dynamin-like GTPase, which is processed by several proteases to produce the soluble short form (OPA1S) or the IMM-anchored form (OPA1L) [77]. Under severe stress conditions, only OPA1L participates in IMM fusion. Furthermore, when OPA1L is processed into OPA1S, IMM fusion is inhibited and mitochondrial fission is promoted [78]. Interestingly, OPA1 has also been shown to maintain the shape of mitochondrial cristae in a fusion-independent manner by antagonizing the Bcl2 associated protein X (BAX)-mediated apoptosis [79].
Recently, sorting assembly machinery 50 kDa subunit (SAM50), a β-barrel channel on the mitochondrial outer membrane, has been suggested to participate in mitophagy [80]. SAM50 forms a protein complex with MICOS to sustain the structural integrity of cristae [81]. Of note, the depletion of SAM50 induces fragmented mitochondria, which subsequently fused to form large abnormal spherical mitochondria [80]. By further interaction analysis, SAM50 was found to interact with PINK1 to regulate PINK1 stability. Moreover, it was also observed that the level of PINK1 was enhanced and Parkin-dependent mitophagy was accelerated in the SAM50 deficiency cells.
The majority of the mitochondria-related tethering machineries have not been identified in plants. By mitochondrial membrane proteome analysis, a list of novel plant proteins without any predicted mitochondria distribution were verified to distribute on the mitochondrial outer membrane, further suggesting that the mitochondrial tethering machinery might be diversified and specialized in plant species [82]. Recently, a plant-unique MICOS complex subunit, called DGD1 SUPPRESSOR1 (DGS1), has been found to regulate mitochondria and chloroplast morphology, as well as lipid homeostasis [83]. Particularly, the OMM-localized DGS1 serves as a bridge to link OMM-localized TOM40/20 and IMM-localized MIC60 [84]. However, orthologs of DGS1 are only conserved in yeast, but not in metazoan, implying a complexity in plant mitochondrial homeostasis during evolution. Nevertheless, a conserved protein in eukaryotic organisms, named FRIENDLY, which belongs to the CLUSTERED MITOCHONDRIA (CLUH) superfamily, has been reported to function in mitochondrial fusion in Arabidopsis [62]. It has been shown that mitochondria in friendly mutant cells formed large clusters. Interestingly, a recent study also uncovered a unique role of FRIENDLY in plant mitophagy [21]. In friendly mutants, abnormal mitophagosomes accumulated, and the level of mitophagy was significantly reduced upon mitochondrial uncoupler treatment, implying a defect in mitophagy due to the loss of FRIENDLY. Importantly, it was observed that Friendly overlapped with ATG8 upon mitophagy induction. This study has thus demonstrated an essential role of the mitochondrial fusion regulator in plant mitophagy for recycling damaged mitochondria during stress and development.
3.1.2. Mitochondrial Fission Machinery
The mitochondrial fission machinery is conserved in most eukaryotes. Small GTPase including dynamin-related protein 1 (DRP1) in mammals and DRP3 in plants both have been shown to function as a driving force to segregate mitochondria [85]. When recruited to the OMM, DRP1 oligomerizes into a ring-like complex to strap the mitochondrial fission site, and ultimately segregates the mitochondrion [85]. However, DRP1 itself does not bind to the OMM directly, and is recruited by several mitochondrial receptors, respectively, including mitochondrial fission factor (MFF), mitochondrial elongation factor 1/2, also known as MiD51 and MiD49 (MIEF1/2) as well as mitochondrial fission 1 protein (FIS1), respectively [86]. Therefore, by interacting with a number of fission regulators, a high-order DRP1 complex is assembled on the mitochondrial surface to induce mitochondrial fission via its GTPase activity [87]. Mitochondrial fission, by isolating the damaged mitochondrial parts, has been generally regarded to make “bite-size” mitochondria prior to their engulfment by the autophagosome. However, a recent study suggested that the fission step might protect healthy mitochondria from degradation, likely by reducing the Parkin activity. It was also observed that loss of DRP1 triggered the recruitment of Parkin to mitochondria, and thus enhanced mitophagy activity [88]. Furthermore, comparing with drp1 or parkin single mutation, the loss of both DRP1 and Parkin significantly compromised mitochondrial degradation, suggesting that both DRP1-dependent and DRP1-independent pathways are involved in mitophagy [89].
Indeed, it has been reported that mitochondrial division for mitophagy might be achieved by the core ATG machinery in coordination with autophagosome formation [90]. In particular, it was found that FIS1 interacts with syntaxin 17 (STX17), an EMC-related SNARE protein [91]. The FIS1-STX17 interaction relocates STX17 from ER onto mitochondria, subsequently recruiting other core ATG proteins, including ATG14. Loss of Fis1 induces the accumulation of STX17 on the mitochondria, which further enhances the formation of autophagosomes for mitophagy. It would be interesting to further investigate whether DRP1 and FIS1 participate in this process coordinately.
In Arabidopsis, there are two functional redundant orthologs of DRP1, named DRP3A and DRP3B, and both drp3a and drp3b mutants contain an abnormal elongated mitochondrial network [63]. The mitochondrial anchor for DRP1 is also conserved in Arabidopsis, including FIS1A (also known as BIGYIN) and FIS1B. Similar to the phenotype in the DRP3 mutant, deficiency of either FIS1A and FIS1B both caused arrested plant growth and abnormal clustered mitochondria [64,66]. Of note, elongated mitochondria 1 (ELM1) and peroxisomal and mitochondrial division factors (PMDs), were found to function as plant-unique fission factors in Arabidopsis. ELM1 is required for the relocation of DRP3 to the mitochondria, while PMD might regulate mitochondrial fission in a DRP3-independent manner, as a deficiency of PMD does not affect the DRP3 targeting the mitochondria [67,68]. Nevertheless, physical interaction for ELM1 with DRP3 or FIS1 has not been experimentally verified. It has been shown that the mutation of elm1 reduced DRP3A/B recruitment to mitochondria when compared with the bigyin mutant in liverworts, suggesting ELM1 might function upstream of mitochondrial fission in plants [65].
In addition, the tethering machinery may coordinate with other trafficking regulators to reinforce membrane fission physically. It has been shown that mammalian DRP1 may interact with Bcl2-interacting factor 1 (BIF1), a BAR domain containing pro-apoptosis protein to mediate mitochondrial fission [92]. The BAR domain is able to induce membrane curvature by bending, thus providing a physical scaffold to facilitate the fission process [93]. Another recent publication has revealed that BIF1 interacts with both PHB2 and OPA1 in human cells to regulate mitochondrial dynamics and mitophagy [94]. Moreover, two Rho GTPases, mitochondrial Rho GTPase 1 and 2 (MIRO1 and MIRO2), were also suggested to participate in mitochondrial fission to mediate mitochondrial trafficking by bridging microtubules [95]. Loss of MIRO1 produced aggregated and thread-like mitochondria, suggesting impaired mitochondrial trafficking that hampered their fission. Recently, it was found that MIRO1 is degraded in Parkin-dependent mitophagy, which prevents the movement of damaged mitochondria to escape from clearance by autophagosomes [96]. Another recent study also reported that MIRO1 may interact with a low level of Parkin under normal conditions, but suppression of MIRO impaired Parkin translocation to mitochondria and suppresses mitophagy [97]. Counterparts of MIRO also exist in plants, with three isoforms in Arabidopsis. Although dysfunction of Arabidopsis MIRO1 caused embryo lethality, elongated mitochondria were detected in pollen tubes [69]. Interestingly, Arabidopsis MIRO2 knockout plants showed no obvious growth defects, but when there was overexpression of an active MIRO2 variant, larger and fewer mitochondria were observed in stable contact with the ER [70]. Contrary to the dominant function of MIRO in mitochondrial movement, MIRO might function in a plant-specific manner for both mitochondrial fusion and motility.
Taken together, multiple fusion/fission machineries participate in mitochondria dynamic and degradation (Table 1). Regarding the conservation and divergence of these fission/fusion machineries in plants, future investigation is required to further dissect their interaction relationships in plant mitochondria dynamic and mitophagy.
3.2. AMPK/SnRK1: A Master Energy Sensor in Balancing Mitophagy and Mitochondrial Dynamics
In addition to a tight coordination between mitochondria fission/fusion machineries and mitophagy regulators, upstream signal molecules also play important roles in coordinating mitochondrial dynamics and mitophagy activity. As one conserved key energy sensor, AMP-activated kinase (AMPK)/sucrose nonfermenting 1 (SNF1)/Snf1-related kinase1 (SnRK1), represents one of the upstream sensors in response to the cellular energy status, and subsequently phosphorylating various substrates to reprogram the cellular metabolism activities to balance mitochondrial biogenesis, mitochondrial fission/fusion and mitophagy [98] (Figure 2).
Firstly, it has been reported that several transcription factors essential for mitochondrial biogenesis are phosphorylated by AMPK in mammals. For example, AMPK phosphorylates peroxisome proliferator-activated receptor-γ (PPARγ) co-activator 1α (PGC1α), which activates mitochondrial biogenesis genes through interaction with PPARγ or oestrogen-related receptors (ERRs) [99,100,101]. On the other hand, AMPK also activates the transcription factor EB (TFEB), which further induces the expression of PGC1α upon binding to their promoter regions [102]. In addition, Acetyl-CoA carboxylase 2 (ACC2), a mitochondrial outer membrane protein involved in lipid metabolism, has also been found to be a substrate of AMPK. Of note, it has been suggested that ACC2 might enhance TFEB target gene expression upon phosphorylation by AMPK, suggesting positive feedback upon AMPK phosphorylation [103,104]. Moreover, it has been reported that AMPK might phosphorylate the mitochondrial outer-membrane receptor MFF, to control the number of mitochondria [105]. The activated MFF then recruits DRP1 to regulate mitochondrial fission process [105].
Particularly, AMPK may regulate mitophagy activity to control the degradation of mitochondria via direct phosphorylation of the core ATG proteins, including ULK1 Kinase. Upon mitochondrial oxidative stress, it has been shown that phosphorylation of AMPK and ULK1 were both increased [106,107]. Then, AMPK-dependent activation of ULK1 kinase facilitates its recruitment to the mitochondria, and further phosphorylates other downstream regulators, such as ATG9, which is suggested to transport the lipids for autophagosome formation, and small GTPase Rab9 for the transport of damaged mitochondria into the lysosome [108,109]. In addition, it was also shown that ULK1 phosphorylates mitophagy receptors to promote mitophagy [110,111]. Another key ATG protein that has been implicated in mitophagy is Beclin1 (ATG6 homolog), which forms different class III PI3K complexes for either intracellular vesicle trafficking or autophagy [112]. Interestingly, it was found that AMPK facilitates the phosphorylation of Beclin1 to form the autophagy PI3K complex to induce autophagy [113,114]. Beclin1 directly interacts with the protein kinase PTEN-induced kinase 1 (PINK1) to aid in the translocation of the E3 ligase Parkin to mitochondrial membrane to induce mitophagy [115,116]. However, anti-apoptotic protein B-cell lymphoma 2 (Bcl-2) binds to Beclin1 to negatively regulate the pro-survival role of autophagy [117]. In addition, a recent study showed that energy stress enhanced Beclin1 interaction with a Toll-like receptor 9, which further activated AMPK activity but inhibited Beclin1-Bcl-2 interaction [118], thus providing another positive feedback loop to enhance autophagy for cell survival.
In plants, the Sucrose Nonfermenting-Related Kinase 1 (SnRK1) family serves as the counterpart of AMPK as an energy sensor and actively participates in plant stress response [119]. Of note, it has recently been shown that SnRK1 subunit KINβγ regulates the biogenesis of mitochondria to monitor ROS levels in pollen [120]. A mutation of kinβγ leads to abnormal mitochondrial biogenesis, as well as reduced ROS production. However, the plant SnRK1 regulation mechanism might be different from that in mammals, as downregulation of KIN10/11 and KINβγ reduced the number of mitochondria, while more mitochondria are observed in AMPK defective mouse liver cells. Nevertheless, ATG1 proteins have also been shown to be phosphorylated by the catalytic α-subunit of SnRK1, KIN10 in Arabidopsis [121,122]. Recently, it was also reported that KIN10 can phosphorylate ATG6 to promote autophagy [123]. In addition, an NAC transcription factor, named suppressor of gamma response 1 (SOG1), was identified to interact with SnRK1 in response to low energy [124]. Moreover, both sog1 and kin10/kin11 mutant can partially restore a mitochondrial mutant sd3, which contains low abundant ATP due to a defect in a mitochondrial inner membrane protein. Another example is the regulation of the alternative oxidase (AOX), a key enzyme involved in the electron transport chain in mitochondria. KIN10 may coordinate with cyclin-dependent kinase E1 (CDKE1) to induce AOX1a, likely via NAC17 transcription factor [125,126]. On the other hand, it was reported SnRK1 might target one subfamily of basic leucine zipper transcription factors (bZIPs), S1-bZIPs, which also participates in regulating mitochondrial respiratory genes [127]. However, whether other mitochondrial fission/fusion regulators are also targeted by SnRK1, such as those in mammals, remains unclear (Figure 2). Nevertheless, it seems that multiple layers of regulation in mitochondrial dynamics and mitophagy by AMPK/SnRK1 are conserved in both mammals and plants in mitochondrial quality control.
4. Interplay between Mitophagy and PCD: A New Role of the BAG Protein Family
Although mitophagy and mitochondrial fission may aid in the clearance of harmful/unwanted mitochondrial material, exactly how these quality-control pathways sense different levels of mitochondrial damage to ensure proper execution remains poorly understood. Upon stress induction and mitochondrial damage, the MOMP is increased, accompanied by the loss of ΔφM, which will further activate PCD [7,128]. This raises another important fundamental question as to how mitophagy and PCD are balanced for cell fate decisions, particularly at the molecular level. In animals, the versatile Bcl-2 protein family functions as a key regulator for PCD [129,130]. There are both pro-survival and pro-death Bcl-2 members reported in animal PCD. In response to stress or developmental signals, pro-death Bcl2-associated protein x (BAX) forms a Cytochrome c channel to facilitate the release of the latter, while pro-survival proteins Bcl-2 and Bcl-XL can either suppress the downstream Caspase or the release of Cytochrome c, respectively, for downstream execution of PCD [130,131]. However, no counterparts of Bcl-2 protein have been identified in the plant genome.
Although no Bcl-2 counterparts have been identified in plants, the Bcl-2-associated athanogene (BAG) family proteins, which were initially discovered as Bcl-2-interacting partners to participate in PCD regulation, are remarkedly conserved in plant genomes [132]. Of note, recent studies in animals and plants implied a crucial role of the BAG proteins in mitochondrial homeostasis, PCD and stress response (Figure 3). Importantly, several BAG proteins have been implicated to negatively or positively regulate Parkin-mediated mitophagy [133]. As cochaperones binding to heat shock proteins heat shock protein 70 (Hsp70), BAGs probably serve as an “off-on” switch mechanism between PCD and mitophagy by bridging various molecular chaperones and target proteins, thus balancing the level of PCD and mitophagy.
There are six BAG proteins identified in animals, and all of them contain at least one BAG domain for binding to the ATPase domain of Hsc70 [134,151]. As the first identified Bcl-2 associated BAG protein, BAG1 has been extensively studied. BAG1 contains an N terminus ubiquitin-like domain (UBL) to form a complex with Hsc70, and an E3 ligase named C terminus of Hsc70-interacting protein (CHIP) for proteasome-mediated degradation [134,151]. Differently, BAG3 comprises a tryptophan–tryptophan (WW) domain and PxxP motif, both of which bind to proline-rich proteins. In particular, BAG3 harbors two additional IPV (Ile-Pro-Val) motifs for recruiting specific HSP70, and links the BAG3-containing protein aggregates via the autophagic adaptor p62 [136]. Interestingly, it has been suggested that a shift in binding with Hsc70 from BAG1 to BAG3 may serve as a switch-off mechanism to balance proteasome-dependent proteolysis and selective autophagy [137]. Another piece of evidence showed that BAG3 interacts with an actin-binding protein Synaptopodin, named SYNPO, to facilitate the clearance of microtubule-associated protein tau (MAPT), further supporting the specific role of BAG3 in selective autophagy [138]. Recently, another study also demonstrated that BAG3 may promote mitophagy by recruiting Parkin to the depolarized mitochondria [135]. Conversely, BAG4 and BAG5 have been implicated in disturbing the translocation of PARKIN to the mitochondria, thus suppressing mitophagy [139,140]. Of note, BAG5 directly binds to Parkin to suppress its E3 ligase activity for mitophagy, but promotes Parkin-mediated proteolysis degradation for PCD upon depolarization of mitochondria, indicating a role of BAG5 in switching the pro-survival and pro-death activity of Parkin [139]. Differently, the nucleus-localized BAG6 might trigger a cell death pathway via binding to the apoptosis inducer REAPER, and it also regulates the unfolded protein response for protein quality control in the cytosol [141].
In A. thaliana, seven BAG proteins have been predicted based on their consensus BAG domain, and are distinguished into two groups [132]. Arabidopsis BAG1-4 share similar structures as human BAG1, which comprise an N-terminal UBL domain and a C-terminal BAG domain [129], likely playing a redundant role in Arabidopsis. Differently, BAG5-7 contain a plant-unique CaM (calmodium) binding motif (IQ) as well as the BAG domain, but lack the UBL domain [129]. It is implicated that all Arabidopsis BAGs are associated with Hsp70, except BAG6 [132,142]. Structural analysis has also revealed that Arabidopsis BAG1 and human BAG1 both use conserved residues within their BAG domain for interacting with Hsp70 (Figure 4) [143]. Moreover, recent progress has unveiled important roles of the BAG protein family in the plant stress response. For example, BAG1 has been reported to participate in the degradation of unimported plastid proteins, together with Hsp70 and CHIP, and overexpression of BAG1 impairs plant development and response to high salt conditions [142]. In another study, it was shown that BAG2 deficiency promotes plant growth [144]. However, both bag4 and bag6 mutant plants displayed an early senescence and higher susceptibility to salt stress [132]. All these data support that the level of BAG proteins is critical in plant development and stress response. Recently, through yeast two hybrid screening, it was also reported that BAG4 interacts with KAT1 potassium channel in mediating stomatal opening [145]. Regarding the essential role of stomata opening in drought tolerance and pathogen susceptibility, the BAG4–KAT1 interaction might function as a posttranslational regulation mechanism to participate in plant stress response.
Among all the Arabidopsis BAG proteins, only BAG5 has been reported to localize on the mitochondria, and structural analysis suggests a role of BAG5 in the spatiotemporal regulation of plant senescence [146]. Under normal conditions, BAG5 binds to CaM and Hsp70 independently [146]. However, when the mitochondrion is damaged and the matrix Ca2+ is released, the high Ca2+ induces a change in the binding mode for CaM with BAG5, which inhibits BAG5 binding to Hsp70 to enhance ROS level, and finally leads to senescence. It has also been shown that no significant developmental defect was observed in the bag5 mutant. Conversely, overexpression of BAG5 or its IQ domain accelerated leaf senescence, and senescence-associated genes (SAGs) were also upregulated. The direct evidence that links BAG protein in autophagy comes from recent studies of BAG6. It was reported that BAG6 triggers autophagy upon fungal infection [147]. Further analysis showed that the invading fungi insert their chitin hypha into the infected plant cell, which subsequently leads to the vacuolar cleavage of BAG6 on a specific caspase-1 site via binding to a C2 GRAM domain protein (BAGP1) and an aspartyl protease (APCB1) [148]. The truncated BAG6 then activated autophagic response to the pathogen, although the substrate for degradation is unknown [148]. Significantly, mutations on the BAG6 cleavage site suppressed autophagy in plants and inhibited disease resistance, implying that proteolytic activity is also required for BAG6 to execute its pro-survival function [148].
Compared with other BAGs, BAG7 has a unique distribution on the ER, and has been shown to interact with BIP2 in the bZIP28-mediated unfolded protein response (UPR) upon heat and cold stress [149]. An essential role of BAG7 in ER-induced cell death is supported by the observation that cell death is accelerated in bag7 mutants [149]. It was proposed that BAG7–BIP2 interaction might facilitate the retention of bZIP28 on the ER, whereby BAG7 undergoes proteolytic cleavage and is translocated into the nucleus under stress conditions [149]. After further SUMOylation modification, BAG7 interacts with the transcription factor WRKY DNA binding protein 29 (WRKY29) to mediate chaperone expression, while bZIP028 interacts with bZIP60 to activate UPR gene expression [150]. Therefore, BAG7 also functions as a “switch-on” signal for cell death. Surprisingly, an AIM-like motif is predicted in BAG7 [54]. Thus, it will be interesting to determine, in the future, whether BAG7 is a target of ATG8 to balance between cell death and cell survival. Furthermore, another study also unveiled an unexpected anti-survival role of BAX inhibitor 1 (BI1) in bZIP28-mediated pro-survival signaling in ER stress recovery. It has been shown that sensitivity to ER stress is partially recovered in the bzip28 bi1-2 double mutant when compared with bzip28 in roots, suggesting that BI1 may antagonize the effect of bZIP28 during ER stress recovery [152]. In addition, Arabidopsis BI1 also binds to ATG6, and the suppression of BI1 expression impaired the formation of autophagosomes upon viral infection, while its overexpression caused autophagy-dependent cell death [153]. Taken together, although plant genomes lack BCL-2 members, tremendous progress has been made to unveil a diversification of BAG proteins for the switching of cell fate in a plant-specific manner. Future efforts are required to further identify the BAG network to reveal their specific roles in balancing plant organelle quality control, particularly the mitochondria, and PCD.
5. Conclusions
Both mitochondrial biogenesis and mitochondrial degradation are essential for maintaining mitochondrial homeostasis. Mitophagy, as a key degradation process for the clearance of unwanted mitochondria, has been intensively studied in animal models for its close involvement with several diseases. Compared with other systems, it is obvious that there are lots of questions unsolved in plant mitophagy: What are the receptors linking the autophagosome and the mitochondrion under different conditions in plants? How do the mitochondrial fission/fusion machineries coordinate with the ATG machinery to facilitate mitophagy in plants? How is plant PCD executed to antagonize mitophagy under stress conditions? Future exploration into the molecular network for linking mitochondrial fission/fusion machinery, mitophagy and PCD should provide us with a better understanding of how mitochondrial homeostasis is maintained for plant development and stress response.
Author Contributions
X.Z. conceived of the concept and the organization of the manuscript; K.R., L.F., S.S. and X.Z. wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Research Grants Council of Hong Kong, grant number 24108820, N_CUHK405/20, G-CUHK404/18, C4002-17G, R4005-18F, AoE/M-05/12, AoE/M-403/16, and the Chinese University of Hong Kong Research Committee.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We apologize to researchers whose work has not been included in this manuscript owing to space limitations.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Karnkowska, A.; Vacek, V.; Zubáčová, Z.; Treitli, S.C.; Petrželková, R.; Eme, L.; Novák, L.; Žárský, V.; Barlow, L.D.; Herman, E.K.; et al. A Eukaryote without a Mitochondrial Organelle. Curr. Biol. 2016, 26, 1274–1284. [Google Scholar] [CrossRef] [PubMed]
- Van Vranken, J.G.; Jeong, M.Y.; Wei, P.; Chen, Y.C.; Gygi, S.P.; Winge, D.R.; Rutter, J. The mitochondrial acyl carrier protein (ACP) coordinates mitochondrial fatty acid synthesis with iron sulfur cluster biogenesis. eLife 2016, 5, e17828. [Google Scholar] [CrossRef] [PubMed]
- Ashkavand, Z.; Sarasija, S.; Ryan, K.C.; Laboy, J.T.; Norman, K.R. Corrupted ER-mitochondrial calcium homeostasis promotes the collapse of proteostasis. Aging Cell 2020, 19, e13065. [Google Scholar] [CrossRef] [PubMed]
- Belt, K.; Huang, S.; Thatcher, L.F.; Casarotto, H.; Singh, K.B.; Van Aken, O.; Millar, A.H. Salicylic Acid—Dependent Plant Stress Signaling via Mitochondrial Succinate Dehydrogenase. Plant Physiol. 2017, 173, 2029–2040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, R.; Jones, A.D.; Hu, J. Cardiolipin-mediated mitochondrial dynamics and stress response in Arabidopsis. Plant Cell 2014, 26, 391–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuchs, R.; Kopischke, M.; Klapprodt, C.; Hause, G.; Meyer, A.J.; Schwarzlander, M.; Fricker, M.D.; Lipka, V. Immobilized Subpopulations of Leaf Epidermal Mitochondria Mediate PENETRATION2-Dependent Pathogen Entry Control in Arabidopsis. Plant Cell 2016, 28, 130–145. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Youle, R.J. The role of mitochondria in apoptosis. Annu. Rev. Genet. 2009, 43, 95–118. [Google Scholar] [CrossRef] [Green Version]
- Lam, E.; Kato, N.; Lawton, M. Programmed cell death, mitochondria and the plant hypersensitive response. Nature 2001, 411, 848–853. [Google Scholar] [CrossRef]
- Garrido, C.; Galluzzi, L.; Brunet, M.; Puig, P.E.; Didelot, C.; Kroemer, G. Mechanisms of cytochrome c release from mitochondria. Cell Death Differ. 2006, 13, 1423–1433. [Google Scholar] [CrossRef] [Green Version]
- Scorrano, L.; Ashiya, M.; Buttle, K.; Weiler, S.; Oakes, S.A.; Mannella, C.A.; Korsmeyer, S.J. A distinct pathway remodels mitochondrial cristae and mobilizes cytochrome c during apoptosis. Dev. Cell 2002, 2, 55–67. [Google Scholar] [CrossRef] [Green Version]
- Arimura, S.I. Fission and Fusion of Plant Mitochondria, and Genome Maintenance. Plant Physiol. 2018, 176, 152–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broda, M.; Millar, A.H.; Van Aken, O. Mitophagy: A Mechanism for Plant Growth and Survival. Trends Plant Sci. 2018, 23, 434–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, D.C. Fusion and fission: Interlinked processes critical for mitochondrial health. Annu. Rev. Genet. 2012, 46, 265–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Twig, G.; Elorza, A.; Molina, A.J.; Mohamed, H.; Wikstrom, J.D.; Walzer, G.; Stiles, L.; Haigh, S.E.; Katz, S.; Las, G.; et al. Fission and selective fusion govern mitochondrial segregation and elimination by autophagy. EMBO J. 2008, 27, 433–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minibayeva, F.; Dmitrieva, S.; Ponomareva, A.; Ryabovol, V. Oxidative stress-induced autophagy in plants: The role of mitochondria. Plant Physiol. Biochem. 2012, 59, 11–19. [Google Scholar] [CrossRef]
- Liu, Y.; Bassham, D.C. Autophagy: Pathways for self-eating in plant cells. Annu. Rev. Plant Biol. 2012, 63, 215–237. [Google Scholar] [CrossRef] [Green Version]
- Soto-Burgos, J.; Zhuang, X.; Jiang, L.; Bassham, D.C. Dynamics of Autophagosome Formation. Plant Physiol. 2018, 176, 219–229. [Google Scholar] [CrossRef] [Green Version]
- Marshall, R.S.; Vierstra, R.D. Autophagy: The Master of Bulk and Selective Recycling. Annu. Rev. Plant Biol. 2018, 69, 173–208. [Google Scholar] [CrossRef]
- Zhuang, X.; Chung, K.P.; Luo, M.; Jiang, L. Autophagosome Biogenesis and the Endoplasmic Reticulum: A Plant Perspective. Trends Plant Sci. 2018, 23, 677–692. [Google Scholar] [CrossRef]
- Paszkiewicz, G.; Gualberto, J.M.; Benamar, A.; Macherel, D.; Logan, D.C. Arabidopsis Seed Mitochondria Are Bioenergetically Active Immediately upon Imbibition and Specialize via Biogenesis in Preparation for Autotrophic Growth. Plant Cell 2017, 29, 109–128. [Google Scholar] [CrossRef]
- Ma, J.; Liang, Z.; Zhao, J.; Wang, P.; Ma, W.; Fernandez Andrade, J.A.; Zeng, Y.; Grujic, N.; Jiang, L.; Dagdas, Y.; et al. Friendly regulates membrane depolarization induced mitophagy in Arabidopsis. BioRxiv 2020. [Google Scholar] [CrossRef]
- Li, F.; Chung, T.; Vierstra, R.D. AUTOPHAGY-RELATED11 plays a critical role in general autophagy- and senescence-induced mitophagy in Arabidopsis. Plant Cell 2014, 26, 788–807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Z.P.; Klionsky, D.J. Autophagosome formation: Core machinery and adaptations. Nat. Cell Biol. 2007, 9, 1102–1109. [Google Scholar] [CrossRef] [PubMed]
- Geisler, S.; Holmstrom, K.M.; Skujat, D.; Fiesel, F.C.; Rothfuss, O.C.; Kahle, P.J.; Springer, W. PINK1/Parkin-mediated mitophagy is dependent on VDAC1 and p62/SQSTM1. Nat. Cell Biol. 2010, 12, 119–131. [Google Scholar] [CrossRef] [PubMed]
- Yun, J.N.; Puri, R.; Yang, H.; Lizzio, M.A.; Wu, C.L.; Sheng, Z.H.; Guo, M. MUL1 acts in parallel to the PINK1/parkin pathway in regulating mitofusin and compensates for loss of PINK1/parkin. eLife 2014, 3, e01958. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Qi, W.; Chen, G.; Feng, D.; Liu, J.; Ma, B.; Zhou, C.; Mu, C.; Zhang, W.; Chen, Q.; et al. Mitochondrial outer-membrane E3 ligase MUL1 ubiquitinates ULK1 and regulates selenite-induced mitophagy. Autophagy 2015, 11, 1216–1229. [Google Scholar] [CrossRef] [Green Version]
- Prudent, J.; Zunino, R.; Sugiura, A.; Mattie, S.; Shore, G.C.; McBride, H.M. MAPL SUMOylation of Drp1 Stabilizes an ER/Mitochondrial Platform Required for Cell Death. Mol. Cell 2015, 59, 941–955. [Google Scholar] [CrossRef]
- Szargel, R.; Shani, V.; Abd Elghani, F.; Mekies, L.N.; Liani, E.; Rott, R.; Engelender, S. The PINK1, synphilin-1 and SIAH-1 complex constitutes a novel mitophagy pathway. Hum. Mol. Genet. 2016, 25, 3476–3490. [Google Scholar] [CrossRef] [Green Version]
- Villa, E.; Proics, E.; Rubio-Patino, C.; Obba, S.; Zunino, B.; Bossowski, J.P.; Rozier, R.M.; Chiche, J.; Mondragon, L.; Riley, J.S.; et al. Parkin-Independent Mitophagy Controls Chemotherapeutic Response in Cancer Cells. Cell Rep. 2017, 20, 2846–2859. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Feng, D.; Chen, G.; Chen, M.; Zheng, Q.X.; Song, P.P.; Ma, Q.; Zhu, C.Z.; Wang, R.; Qi, W.J.; et al. Mitochondrial outer-membrane protein FUNDC1 mediates hypoxia-induced mitophagy in mammalian cells. Nat. Cell Biol. 2012, 14, 177–185. [Google Scholar] [CrossRef]
- Rogov, V.V.; Suzuki, H.; Marinkovic, M.; Lang, V.; Kato, R.; Kawasaki, M.; Buljubasic, M.; Sprung, M.; Rogova, N.; Wakatsuki, S.; et al. Phosphorylation of the mitochondrial autophagy receptor Nix enhances its interaction with LC3 proteins. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Hanna, R.A.; Quinsay, M.N.; Orogo, A.M.; Giang, K.; Rikka, S.; Gustafsson, A.B. Microtubule-associated protein 1 light chain 3 (LC3) interacts with Bnip3 protein to selectively remove endoplasmic reticulum and mitochondria via autophagy. J. Biol. Chem. 2012, 287, 19094–19104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhujabal, Z.; Birgisdottir, A.B.; Sjottem, E.; Brenne, H.B.; Overvatn, A.; Habisov, S.; Kirkin, V.; Lamark, T.; Johansen, T. FKBP8 recruits LC3A to mediate Parkin-independent mitophagy. EMBO Rep. 2017, 18, 947–961. [Google Scholar] [CrossRef] [PubMed]
- Van Humbeeck, C.; Cornelissen, T.; Hofkens, H.; Mandemakers, W.; Gevaert, K.; De Strooper, B.; Vandenberghe, W. Parkin Interacts with Ambra1 to Induce Mitophagy. J. Neurosci. 2011, 31, 10249–10261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strappazzon, F.; Nazio, F.; Corrado, M.; Cianfanelli, V.; Romagnoli, A.; Fimia, G.M.; Campello, S.; Nardacci, R.; Piacentini, M.; Campanella, M.; et al. AMBRA1 is able to induce mitophagy via LC3 binding, regardless of PARKIN and p62/SQSTM1. Cell Death Differ. 2015, 22, 419–432. [Google Scholar] [CrossRef] [Green Version]
- Wei, Y.; Chiang, W.C.; Sumpter, R., Jr.; Mishra, P.; Levine, B. Prohibitin 2 Is an Inner Mitochondrial Membrane Mitophagy Receptor. Cell 2017, 168, 224–238. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Choi, S.G.; Yoo, S.M.; Son, J.H.; Jung, Y.K. Choline dehydrogenase interacts with SQSTM1/p62 to recruit LC3 and stimulate mitophagy. Autophagy 2014, 10, 1906–1920. [Google Scholar] [CrossRef] [Green Version]
- Ambivero, C.T.; Cilenti, L.; Main, S.; Zervos, A.S. Mulan E3 ubiquitin ligase interacts with multiple E2 conjugating enzymes and participates in mitophagy by recruiting GABARAP. Cell. Signal. 2014, 26, 2921–2929. [Google Scholar] [CrossRef]
- Murakawa, T.; Okamoto, K.; Omiya, S.; Taneike, M.; Yamaguchi, O.; Otsu, K. A Mammalian Mitophagy Receptor, Bcl2-L-13, Recruits the ULK1 Complex to Induce Mitophagy. Cell Rep. 2019, 26, 338–345. [Google Scholar] [CrossRef]
- Chu, C.T.; Ji, J.; Dagda, R.K.; Jiang, J.F.; Tyurina, Y.Y.; Kapralov, A.A.; Tyurin, V.A.; Yanamala, N.; Shrivastava, I.H.; Mohammadyani, D.; et al. Cardiolipin externalization to the outer mitochondrial membrane acts as an elimination signal for mitophagy in neuronal cells. Nat. Cell Biol. 2013, 15, 1197–1205. [Google Scholar] [CrossRef] [Green Version]
- Sentelle, R.D.; Senkal, C.E.; Jiang, W.H.; Ponnusamy, S.; Gencer, S.; Selvam, S.P.; Ramshesh, V.K.; Peterson, Y.K.; Lemasters, J.J.; Szulc, Z.M.; et al. Ceramide targets autophagosomes to mitochondria and induces lethal mitophagy. Nat. Chem. Biol. 2012, 8, 831–838. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, K.; Kondo-Okamoto, N.; Ohsumi, Y. Mitochondria-anchored receptor Atg32 mediates degradation of mitochondria via selective autophagy. Dev. Cell 2009, 17, 87–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aoki, Y.; Kanki, T.; Hirota, Y.; Kurihara, Y.; Saigusa, T.; Uchiumi, T.; Kang, D.C. Phosphorylation of Serine 114 on Atg32 mediates mitophagy. Mol. Biol. Cell 2011, 22, 3206–3217. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Jin, M.; Liu, X.; Klionsky, D.J. Proteolytic processing of Atg32 by the mitochondrial i-AAA protease Yme1 regulates mitophagy. Autophagy 2013, 9, 1828–1836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kondo-Okamoto, N.; Noda, N.N.; Suzuki, S.W.; Nakatogawa, H.; Takahashi, I.; Matsunami, M.; Hashimoto, A.; Inagaki, F.; Ohsumi, Y.; Okamoto, K. Autophagy-related protein 32 acts as autophagic degron and directly initiates mitophagy. J. Biol. Chem. 2012, 287, 10631–10638. [Google Scholar] [CrossRef] [Green Version]
- Fukuda, T.; Ebi, Y.; Saigusa, T.; Furukawa, K.; Yamashita, S.I.; Inoue, K.; Kobayashi, D.; Yoshida, Y.; Kanki, T. Atg43 tethers isolation membranes to mitochondria to promote starvation-induced mitophagy in fission yeast. eLife 2020, 9, e61245. [Google Scholar] [CrossRef]
- Lazarou, M.; Sliter, D.A.; Kane, L.A.; Sarraf, S.A.; Wang, C.; Burman, J.L.; Sideris, D.P.; Fogel, A.I.; Youle, R.J. The ubiquitin kinase PINK1 recruits autophagy receptors to induce mitophagy. Nature 2015, 524, 309–314. [Google Scholar] [CrossRef] [Green Version]
- Sauve, V.; Sung, G.; Soya, N.; Kozlov, G.; Blaimschein, N.; Miotto, L.S.; Trempe, J.F.; Lukacs, G.L.; Gehring, K. Mechanism of parkin activation by phosphorylation. Nat. Struct. Mol. Biol. 2018, 25, 623–630. [Google Scholar] [CrossRef]
- Dove, K.K.; Klevit, R.E. RING-Between-RING E3 Ligases: Emerging Themes amid the Variations. J. Mol. Biol. 2017, 429, 3363–3375. [Google Scholar] [CrossRef]
- Mladek, C.; Guger, K.; Hauser, M.T. Identification and characterization of the ARIADNE gene family in Arabidopsis. A group of putative E3 ligases. Plant Physiol. 2003, 131, 27–40. [Google Scholar] [CrossRef] [Green Version]
- Fernandez, M.A.; Belda-Palazon, B.; Julian, J.; Coego, A.; Lozano-Juste, J.; Inigo, S.; Rodriguez, L.; Bueso, E.; Goossens, A.; Rodriguez, P.L. RBR-Type E3 Ligases and the Ubiquitin-Conjugating Enzyme UBC26 Regulate Abscisic Acid Receptor Levels and Signaling. Plant Physiol. 2020, 182, 1723–1742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, R.; Hu, J. The Arabidopsis E3 Ubiquitin Ligase SP1 Targets to Chloroplasts, Peroxisomes, and Mitochondria. Plant Physiol. 2018, 176, 480–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, K.; Wang, K.; Liu, X.; Klionsky, D.J. The Scaffold Protein Atg11 Recruits Fission Machinery to Drive Selective Mitochondria Degradation by Autophagy. Dev. Cell 2013, 26, 9–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marshall, R.S.; Hua, Z.; Mali, S.; McLoughlin, F.; Vierstra, R.D. ATG8-binding UIM proteins define a new class of autophagy adaptors and receptors. Cell 2019, 177, 766–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robert, N.; d’Erfurth, I.; Marmagne, A.; Erhardt, M.; Allot, M.; Boivin, K.; Gissot, L.; Monachello, D.; Michaud, M.; Duchene, A.M. Voltage-dependent-anion-channels (VDACs) in Arabidopsis have a dual localization in the cell but show a distinct role in mitochondria. Plant Mol. Biol. 2012, 78, 431–446. [Google Scholar] [CrossRef] [PubMed]
- Tateda, C.; Watanabe, K.; Kusano, T.; Takahashi, Y. Molecular and genetic characterization of the gene family encoding the voltage-dependent anion channel in Arabidopsis. J. Exp. Bot. 2011, 62, 4773–4785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Aken, O.; Pecenkova, T.; van de Cotte, B.; De Rycke, R.; Eeckhout, D.; Fromm, H.; De Jaeger, G.; Witters, E.; Beemster, G.T.; Inze, D.; et al. Mitochondrial type-I prohibitins of Arabidopsis thaliana are required for supporting proficient meristem development. Plant J. 2007, 52, 850–864. [Google Scholar] [CrossRef] [Green Version]
- Seguel, A.; Jelenska, J.; Herrera-Vasquez, A.; Marr, S.K.; Joyce, M.B.; Gagesch, K.R.; Shakoor, N.; Jiang, S.C.; Fonseca, A.; Wildermuth, M.C.; et al. PROHIBITIN3 Forms Complexes with ISOCHORISMATE SYNTHASE1 to Regulate Stress-Induced Salicylic Acid Biosynthesis in Arabidopsis. Plant Physiol. 2018, 176, 2515–2531. [Google Scholar] [CrossRef] [Green Version]
- Smakowska, E.; Skibior-Blaszczyk, R.; Czarna, M.; Kolodziejczak, M.; Kwasniak-Owczarek, M.; Parys, K.; Funk, C.; Janska, H. Lack of FTSH4 Protease Affects Protein Carbonylation, Mitochondrial Morphology, and Phospholipid Content in Mitochondria of Arabidopsis: New Insights into a Complex Interplay. Plant Physiol. 2016, 171, 2516–2535. [Google Scholar] [CrossRef] [Green Version]
- Ban, T.; Ishihara, T.; Kohno, H.; Saita, S.; Ichimura, A.; Maenaka, K.; Oka, T.; Mihara, K.; Ishihara, N. Molecular basis of selective mitochondrial fusion by heterotypic action between OPA1 and cardiolipin. Nat. Cell Biol. 2017, 19, 856–863. [Google Scholar] [CrossRef]
- Gao, H.; Sage, T.L.; Osteryoung, K.W. FZL, an FZO-like protein in plants, is a determinant of thylakoid and chloroplast morphology. Proc. Natl. Acad. Sci. USA 2006, 103, 6759–6764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Zawily, A.M.; Schwarzlander, M.; Finkemeier, I.; Johnston, I.G.; Benamar, A.; Cao, Y.; Gissot, C.; Meyer, A.J.; Wilson, K.; Datla, R.; et al. FRIENDLY regulates mitochondrial distribution, fusion, and quality control in Arabidopsis. Plant Physiol. 2014, 166, 808–828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujimoto, M.; Arimura, S.; Mano, S.; Kondo, M.; Saito, C.; Ueda, T.; Nakazono, M.; Nakano, A.; Nishimura, M.; Tsutsumi, N. Arabidopsis dynamin-related proteins DRP3A and DRP3B are functionally redundant in mitochondrial fission, but have distinct roles in peroxisomal fission. Plant J. 2009, 58, 388–400. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Hu, J. Two small protein families, DYNAMIN-RELATED PROTEIN3 and FISSION1, are required for peroxisome fission in Arabidopsis. Plant J. 2009, 57, 146–159. [Google Scholar] [CrossRef] [PubMed]
- Nagaoka, N.; Yamashita, A.; Kurisu, R.; Watari, Y.; Ishizuna, F.; Tsutsumi, N.; Ishizaki, K.; Kohchi, T.; Arimura, S. DRP3 and ELM1 are required for mitochondrial fission in the liverwort Marchantia polymorpha. Sci. Rep. 2017, 7, 4600. [Google Scholar] [CrossRef] [Green Version]
- Scott, I.; Tobin, A.K.; Logan, D.C. BIGYIN, an orthologue of human and yeast FIS1 genes functions in the control of mitochondrial size and number in Arabidopsis thaliana. J. Exp. Bot. 2006, 57, 1275–1280. [Google Scholar] [CrossRef] [Green Version]
- Arimura, S.; Fujimoto, M.; Doniwa, Y.; Kadoya, N.; Nakazono, M.; Sakamoto, W.; Tsutsumi, N. Arabidopsis ELONGATED MITOCHONDRIA1 is required for localization of DYNAMIN-RELATED PROTEIN3A to mitochondrial fission sites. Plant Cell 2008, 20, 1555–1566. [Google Scholar] [CrossRef] [Green Version]
- Aung, K.; Hu, J. The Arabidopsis tail-anchored protein PEROXISOMAL AND MITOCHONDRIAL DIVISION FACTOR1 is involved in the morphogenesis and proliferation of peroxisomes and mitochondria. Plant Cell 2011, 23, 4446–4461. [Google Scholar] [CrossRef] [Green Version]
- Yamaoka, S.; Leaver, C.J. EMB2473/MIRO1, an Arabidopsis Miro GTPase, is required for embryogenesis and influences mitochondrial morphology in pollen. Plant Cell 2008, 20, 589–601. [Google Scholar] [CrossRef] [Green Version]
- White, R.R.; Lin, C.; Leaves, I.; Castro, I.G.; Metz, J.; Bateman, B.C.; Botchway, S.W.; Ward, A.D.; Ashwin, P.; Sparkes, I. Miro2 tethers the ER to mitochondria to promote mitochondrial fusion in tobacco leaf epidermal cells. Commun. Biol. 2020, 3, 161. [Google Scholar] [CrossRef] [Green Version]
- Koshiba, T.; Detmer, S.A.; Kaiser, J.T.; Chen, H.C.; McCaffery, J.M.; Chan, D.C. Structural basis of mitochondrial tethering by mitofusin complexes. Science 2004, 305, 858–862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Böckler, S.; Westermann, B. Mitochondrial ER Contacts Are Crucial for Mitophagy in Yeast. Dev. Cell 2014, 28, 450–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szabadkai, G.; Bianchi, K.; Varnai, P.; De Stefani, D.; Wieckowski, M.R.; Cavagna, D.; Nagy, A.I.; Balla, T.; Rizzuto, R. Chaperone-mediated coupling of endoplasmic reticulum and mitochondrial Ca2+ channels. J. Cell Biol. 2006, 175, 901–911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narendra, D.; Kane, L.A.; Hauser, D.N.; Fearnley, I.M.; Youle, R.J. p62/SQSTM1 is required for Parkin-induced mitochondrial clustering but not mitophagy; VDAC1 is dispensable for both. Autophagy 2010, 6, 1090–1106. [Google Scholar] [CrossRef] [PubMed]
- McLelland, G.L.; Goiran, T.; Yi, W.; Dorval, G.; Chen, C.X.; Lauinger, N.D.; Krahn, A.I.; Valimehr, S.; Rakovic, A.; Rouiller, I.; et al. Mfn2 ubiquitination by PINK1/parkin gates the p97-dependent release of ER from mitochondria to drive mitophagy. eLife 2018, 7, e32866. [Google Scholar] [CrossRef] [PubMed]
- Glytsou, C.; Calvo, E.; Cogliati, S.; Mehrotra, A.; Anastasia, I.; Rigoni, G.; Raimondi, A.; Shintani, N.; Loureiro, M.; Vazquez, J.; et al. Optic Atrophy 1 Is Epistatic to the Core MICOS Component MIC60 in Mitochondrial Cristae Shape Control. Cell Rep. 2016, 17, 3024–3034. [Google Scholar] [CrossRef] [Green Version]
- Ge, Y.F.; Shi, X.J.; Boopathy, S.; McDonald, J.; Smith, A.W.; Chao, L.H. Two forms of Opa1 cooperate to complete fusion of the mitochondrial inner-membrane. eLife 2020, 9, e50973. [Google Scholar] [CrossRef] [PubMed]
- Anand, R.; Wai, T.; Baker, M.J.; Kladt, N.; Schauss, A.C.; Rugarli, E.; Langer, T. The i-AAA protease YME1L and OMA1 cleave OPA1 to balance mitochondrial fusion and fission. J. Cell Biol. 2014, 204, 919–929. [Google Scholar] [CrossRef]
- Frezza, C.; Cipolat, S.; de Brito, O.M.; Micaroni, M.; Beznoussenko, G.V.; Rudka, T.; Bartoli, D.; Polishuck, R.S.; Danial, N.N.; De Strooper, B.; et al. OPA1 controls apoptotic cristae remodeling independently from mitochondrial fusion. Cell 2006, 126, 177–189. [Google Scholar] [CrossRef] [Green Version]
- Jian, F.; Chen, D.; Chen, L.; Yan, C.; Lu, B.; Zhu, Y.; Chen, S.; Shi, A.; Chan, D.C.; Song, Z. Sam50 Regulates PINK1-Parkin-Mediated Mitophagy by Controlling PINK1 Stability and Mitochondrial Morphology. Cell Rep. 2018, 23, 2989–3005. [Google Scholar] [CrossRef]
- Tang, J.; Zhang, K.; Dong, J.; Yan, C.; Hu, C.; Ji, H.; Chen, L.; Chen, S.; Zhao, H.; Song, Z. Sam50-Mic19-Mic60 axis determines mitochondrial cristae architecture by mediating mitochondrial outer and inner membrane contact. Cell Death Differ. 2020, 27, 146–160. [Google Scholar] [CrossRef] [PubMed]
- Duncan, O.; Taylor, N.L.; Carrie, C.; Eubel, H.; Kubiszewski-Jakubiak, S.; Zhang, B.; Narsai, R.; Millar, A.H.; Whelan, J. Multiple lines of evidence localize signaling, morphology, and lipid biosynthesis machinery to the mitochondrial outer membrane of Arabidopsis. Plant Physiol. 2011, 157, 1093–1113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Lavell, A.; Meng, X.; Berkowitz, O.; Selinski, J.; van de Meene, A.; Carrie, C.; Benning, C.; Whelan, J.; De Clercq, I.; et al. Arabidopsis DGD1 SUPPRESSOR1 Is a Subunit of the Mitochondrial Contact Site and Cristae Organizing System and Affects Mitochondrial Biogenesis. Plant Cell 2019, 31, 1856–1878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michaud, M.; Gros, V.; Tardif, M.; Brugiere, S.; Ferro, M.; Prinz, W.A.; Toulmay, A.; Mathur, J.; Wozny, M.; Falconet, D.; et al. AtMic60 Is Involved in Plant Mitochondria Lipid Trafficking and Is Part of a Large Complex. Curr. Biol. 2016, 26, 627–639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smirnova, E.; Griparic, L.; Shurland, D.L.; van der Bliek, A.M. Dynamin-related protein Drp1 is required for mitochondrial division in mammalian cells. Mol. Biol. Cell 2001, 12, 2245–2256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kraus, F.; Ryan, M.T. The constriction and scission machineries involved in mitochondrial fission. J. Cell Sci. 2017, 130, 2953–2960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otera, H.; Wang, C.X.; Cleland, M.M.; Setoguchi, K.; Yokota, S.; Youle, R.J.; Mihara, K. Mff is an essential factor for mitochondrial recruitment of Drp1 during mitochondrial fission in mammalian cells. J. Cell Biol. 2010, 191, 1141–1158. [Google Scholar] [CrossRef] [Green Version]
- Burman, J.L.; Pickles, S.; Wang, C.; Sekine, S.; Vargas, J.N.S.; Zhang, Z.; Youle, A.M.; Nezich, C.L.; Wu, X.; Hammer, J.A.; et al. Mitochondrial fission facilitates the selective mitophagy of protein aggregates. J. Cell Biol. 2017, 216, 3231–3247. [Google Scholar] [CrossRef] [Green Version]
- Kageyama, Y.; Hoshijima, M.; Seo, K.; Bedja, D.; Sysa-Shah, P.; Andrabi, S.A.; Chen, W.; Hoke, A.; Dawson, V.L.; Dawson, T.M.; et al. Parkin-independent mitophagy requires Drp1 and maintains the integrity of mammalian heart and brain. EMBO J. 2014, 33, 2798–2813. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, S.I.; Jin, X.; Furukawa, K.; Hamasaki, M.; Nezu, A.; Otera, H.; Saigusa, T.; Yoshimori, T.; Sakai, Y.; Mihara, K.; et al. Mitochondrial division occurs concurrently with autophagosome formation but independently of Drp1 during mitophagy. J. Cell Biol. 2016, 215, 649–665. [Google Scholar] [CrossRef]
- Xian, H.; Yang, Q.; Xiao, L.; Shen, H.M.; Liou, Y.C. STX17 dynamically regulated by Fis1 induces mitophagy via hierarchical macroautophagic mechanism. Nat. Commun. 2019, 10, 2059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, Y.; Karbowski, M.; Yamaguchi, H.; Kazi, A.; Wu, J.; Sebti, S.M.; Youle, R.J.; Wang, H.G. Loss of Bif-1 suppresses Bax/Bak conformational change and mitochondrial apoptosis. Mol. Cell Biol. 2005, 25, 9369–9382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simunovic, M.; Voth, G.A.; Callan-Jones, A.; Bassereau, P. When Physics Takes Over: BAR Proteins and Membrane Curvature. Trends Cell Biol. 2015, 25, 780–792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, S.G.; Xiao, X.; Wang, S.; Gao, H.; Rafikov, R.; Black, S.; Huang, S.; Ding, H.F.; Yoon, Y.; Kirken, R.A.; et al. Bif-1 Interacts with Prohibitin-2 to Regulate Mitochondrial Inner Membrane during Cell Stress and Apoptosis. J. Am. Soc. Nephrol. 2019, 30, 1174–1191. [Google Scholar] [CrossRef] [PubMed]
- Fransson, Å.; Ruusala, A.; Aspenstom, P. The atypical Rho GTPases Miro-1 and Miro-2 have essential roles in mitochondrial trafficking. Biochem. Bioph. Res. Commun. 2006, 344, 500–510. [Google Scholar] [CrossRef]
- Wang, X.N.; Winter, D.; Ashrafi, G.; Schlehe, J.; Wong, Y.L.; Selkoe, D.; Rice, S.; Steen, J.; LaVoie, M.J.; Schwarz, T.L. PINK1 and Parkin Target Miro for Phosphorylation and Degradation to Arrest Mitochondrial Motility. Cell 2011, 147, 893–906. [Google Scholar] [CrossRef] [Green Version]
- Safiulina, D.; Kuum, M.; Choubey, V.; Gogichaishvili, N.; Liiv, J.; Hickey, M.A.; Cagalinec, M.; Mandel, M.; Zeb, A.; Liiv, M.; et al. Miro proteins prime mitochondria for Parkin translocation and mitophagy. EMBO J. 2019, 38, e99384. [Google Scholar] [CrossRef]
- Ghillebert, R.; Swinnen, E.; Wen, J.; Vandesteene, L.; Ramon, M.; Norga, K.; Rolland, F.; Winderickx, J. The AMPK/SNF1/SnRK1 fuel gauge and energy regulator: Structure, function and regulation. FEBS J. 2011, 278, 3978–3990. [Google Scholar] [CrossRef]
- Puigserver, P.; Wu, Z.D.; Park, C.W.; Graves, R.; Wright, M.; Spiegelman, B.M. A cold-inducible coactivator of nuclear receptors linked to adaptive thermogenesis. Cell 1998, 92, 829–839. [Google Scholar] [CrossRef] [Green Version]
- Eichner, L.J.; Giguere, V. Estrogen related receptors (ERRs): A new dawn in transcriptional control of mitochondrial gene networks. Mitochondrion 2011, 11, 544–552. [Google Scholar] [CrossRef]
- Wu, Z.D.; Rosen, E.D.; Brun, R.; Hauser, S.; Adelmant, G.; Troy, A.E.; McKeon, C.; Darlington, G.J.; Spiegelman, B.M. Cross-regulation of C/EBP alpha and PPAR gamma controls the transcriptional pathway of adipogenesis and insulin sensitivity. Mol. Cell 1999, 3, 151–158. [Google Scholar] [CrossRef]
- Settembre, C.; De Cegli, R.; Mansueto, G.; Saha, P.K.; Vetrini, F.; Visvikis, O.; Huynh, T.; Carissimo, A.; Palmer, D.; Klisch, T.J.; et al. TFEB controls cellular lipid metabolism through a starvation-induced autoregulatory loop. Nat. Cell Biol. 2013, 15, 647–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abu-Elheiga, L.; Brinkley, W.R.; Zhong, L.; Chirala, S.S.; Woldegiorgis, G.; Wakil, S.J. The subcellular localization of acetyl-CoA carboxylase 2. Proc. Natl. Acad. Sci. USA 2000, 97, 1444–1449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, J.Y.; Xu, Z.X.; Ding, Z.Y.; Lu, Y.L.; Yu, Q.H.; Werle, K.D.; Zhou, G.; Park, Y.Y.; Peng, G.; Gambello, M.J.; et al. Myristoylation confers noncanonical AMPK functions in autophagy selectivity and mitochondrial surveillance. Nat. Commun. 2015, 6, 7926. [Google Scholar] [CrossRef] [PubMed]
- Toyama, E.Q.; Herzig, S.; Courchet, J.; Lewis, T.L.; Loson, O.C.; Hellberg, K.; Young, N.P.; Chen, H.; Polleux, F.; Chan, D.C.; et al. AMP-activated protein kinase mediates mitochondrial fission in response to energy stress. Science 2016, 351, 275–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egan, D.F.; Shackelford, D.B.; Mihaylova, M.M.; Gelino, S.; Kohnz, R.A.; Mair, W.; Vasquez, D.S.; Joshi, A.; Gwinn, D.M.; Taylor, R.; et al. Phosphorylation of ULK1 (hATG1) by AMP-activated protein kinase connects energy sensing to mitophagy. Science 2011, 331, 456–461. [Google Scholar] [CrossRef] [Green Version]
- Laker, R.C.; Drake, J.C.; Wilson, R.J.; Lira, V.A.; Lewellen, B.M.; Ryall, K.A.; Fisher, C.C.; Zhang, M.; Saucerman, J.J.; Goodyear, L.J.; et al. Ampk phosphorylation of Ulk1 is required for targeting of mitochondria to lysosomes in exercise-induced mitophagy. Nat. Commun. 2017, 8, 548. [Google Scholar] [CrossRef]
- Tian, W.L.; Li, W.; Chen, Y.Q.; Yan, Z.M.; Huang, X.; Zhuang, H.X.; Zhong, W.T.; Chen, Y.S.; Wu, W.X.; Lin, C.X.; et al. Phosphorylation of ULK1 by AMPK regulates translocation of ULK1 to mitochondria and mitophagy. FEBS Lett. 2015, 589, 1847–1854. [Google Scholar] [CrossRef] [Green Version]
- Mack, H.I.; Zheng, B.; Asara, J.M.; Thomas, S.M. AMPK-dependent phosphorylation of ULK1 regulates ATG9 localization. Autophagy 2012, 8, 1197–1214. [Google Scholar] [CrossRef] [Green Version]
- Joo, J.H.; Dorsey, F.C.; Joshi, A.; Hennessy-Walters, K.M.; Rose, K.L.; McCastlain, K.; Zhang, J.; Iyengar, R.; Jung, C.H.; Suen, D.F.; et al. Hsp90-Cdc37 Chaperone Complex Regulates Ulk1-and Atg13-Mediated Mitophagy. Mol. Cell 2011, 43, 572–585. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.X.; Tian, W.L.; Hu, Z.; Chen, G.; Huang, L.; Li, W.; Zhang, X.L.; Xue, P.; Zhou, C.Q.; Liu, L.; et al. ULK1 translocates to mitochondria and phosphorylates FUNDC1 to regulate mitophagy. EMBO Rep. 2014, 15, 566–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Funderburk, S.F.; Wang, Q.J.; Yue, Z. The Beclin 1-VPS34 complex—At the crossroads of autophagy and beyond. Trends Cell Biol. 2010, 20, 355–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.; Wang, W.; Sun, X.; Xu, D.; Wang, C.; Zhang, Q.; Wang, H.; Luo, W.; Chen, Y.; Chen, H.; et al. AMPK regulates autophagy by phosphorylating BECN1 at threonine 388. Autophagy 2016, 12, 1447–1459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Kim, Y.C.; Fang, C.; Russell, R.C.; Kim, J.H.; Fan, W.; Liu, R.; Zhong, Q.; Guan, K.L. Differential regulation of distinct Vps34 complexes by AMPK in nutrient stress and autophagy. Cell 2013, 152, 290–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gelmetti, V.; De Rosa, P.; Torosantucci, L.; Marini, E.S.; Romagnoli, A.; Di Rienzo, M.; Arena, G.; Vignone, D.; Fimia, G.M.; Valente, E.M. PINK1 and BECN1 relocalize at mitochondria-associated membranes during mitophagy and promote ER-mitochondria tethering and autophagosome formation. Autophagy 2017, 13, 654–669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choubey, V.; Cagalinec, M.; Liiv, J.; Safiulina, D.; Hickey, M.A.; Kuum, M.; Liiv, M.; Anwar, T.; Eskelinen, E.L.; Kaasik, A. BECN1 is involved in the initiation of mitophagy: It facilitates PARK2 translocation to mitochondria. Autophagy 2014, 10, 1105–1119. [Google Scholar] [CrossRef] [Green Version]
- Pattingre, S.; Tassa, A.; Qu, X.; Garuti, R.; Liang, X.H.; Mizushima, N.; Packer, M.; Schneider, M.D.; Levine, B. Bcl-2 antiapoptotic proteins inhibit Beclin 1-dependent autophagy. Cell 2005, 122, 927–939. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Nguyen, P.T.; Wang, X.; Zhao, Y.; Meacham, C.E.; Zou, Z.; Bordieanu, B.; Johanns, M.; Vertommen, D.; Wijshake, T.; et al. TLR9 and beclin 1 crosstalk regulates muscle AMPK activation in exercise. Nature 2020, 578, 605–609. [Google Scholar] [CrossRef]
- Wurzinger, B.; Nukarinen, E.; Nagele, T.; Weckwerth, W.; Teige, M. The SnRK1 Kinase as Central Mediator of Energy Signaling between Different Organelles. Plant Physiol. 2018, 176, 1085–1094. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.Q.; Liu, C.Z.; Li, D.D.; Zhao, T.T.; Li, F.; Jia, X.N.; Zhao, X.Y.; Zhang, X.S. The Arabidopsis KIN beta gamma Subunit of the SnRK1 Complex Regulates Pollen Hydration on the Stigma by Mediating the Level of Reactive Oxygen Species in Pollen. PLoS Genet. 2016, 12, e1006228. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Su, Z.Z.; Huang, L.; Xia, F.N.; Qi, H.; Xie, L.J.; Xiao, S.; Chen, Q.F. The AMP-Activated Protein Kinase KIN10 Is Involved in the Regulation of Autophagy in Arabidopsis. Front. Plant Sci. 2017, 8, 1201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soto-Burgos, J.; Bassham, D.C. SnRK1 activates autophagy via the TOR signaling pathway in Arabidopsis thaliana. PLoS ONE 2017, 12, e0182591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.; Zheng, C.; Liu, F.; Yang, C.; Zheng, P.; Lu, X.; Tian, J.; Chung, T.; Otegui, M.S.; Xiao, S.; et al. Genetic Analyses of the Arabidopsis ATG1 Kinase Complex Reveal Both Kinase-Dependent and Independent Autophagic Routes during Fixed-Carbon Starvation. Plant Cell 2019, 31, 2973–2995. [Google Scholar] [CrossRef] [PubMed]
- Hamasaki, H.; Kurihara, Y.; Kuromori, T.; Kusano, H.; Nagata, N.; Yamamoto, Y.Y.; Shimada, H.; Matsui, M. SnRK1 Kinase and the NAC Transcription Factor SOG1 Are Components of a Novel Signaling Pathway Mediating the Low Energy Response Triggered by ATP Depletion. Front. Plant Sci. 2019, 10, 503. [Google Scholar] [CrossRef] [PubMed]
- Ng, S.; Giraud, E.; Duncan, O.; Law, S.R.; Wang, Y.; Xu, L.; Narsai, R.; Carrie, C.; Walker, H.; Day, D.A.; et al. Cyclin-dependent kinase E1 (CDKE1) provides a cellular switch in plants between growth and stress responses. J. Biol. Chem. 2013, 288, 3449–3459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, X.; Li, L.; De Clercq, I.; Narsai, R.; Xu, Y.; Hartmann, A.; Claros, D.L.; Custovic, E.; Lewsey, M.G.; Whelan, J.; et al. ANAC017 Coordinates Organellar Functions and Stress Responses by Reprogramming Retrograde Signaling. Plant Physiol. 2019, 180, 634–653. [Google Scholar] [CrossRef] [Green Version]
- Pedrotti, L.; Weiste, C.; Nagele, T.; Wolf, E.; Lorenzin, F.; Dietrich, K.; Mair, A.; Weckwerth, W.; Teige, M.; Baena-Gonzalez, E.; et al. Snf1-RELATED KINASE1-Controlled C/S1-bZIP Signaling Activates Alternative Mitochondrial Metabolic Pathways to Ensure Plant Survival in Extended Darkness. Plant Cell 2018, 30, 495–509. [Google Scholar] [CrossRef] [Green Version]
- Takayama, S.; Reed, J.C. Molecular chaperone targeting and regulation by BAG family proteins. Nat. Cell Biol. 2001, 3, E237–E241. [Google Scholar] [CrossRef]
- Kabbage, M.; Kessens, R.; Bartholomay, L.C.; Williams, B. The Life and Death of a Plant Cell. Annu. Rev. Plant Biol. 2017, 68, 375–404. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.; Letai, A.; Sarosiek, K. Regulation of apoptosis in health and disease: The balancing act of BCL-2 family proteins. Nat. Rev. Mol. Cell Biol. 2019, 20, 175–193. [Google Scholar] [CrossRef]
- Kale, J.; Osterlund, E.J.; Andrews, D.W. BCL-2 family proteins: Changing partners in the dance towards death. Cell Death Differ. 2018, 25, 65–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doukhanina, E.V.; Chen, S.; van der Zalm, E.; Godzik, A.; Reed, J.; Dickman, M.B. Identification and functional characterization of the BAG protein family in Arabidopsis thaliana. J. Biol. Chem. 2006, 281, 18793–18801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapadia, M.; De Snoo, M.L.; Kalia, L.V.; Kalia, S.K. Regulation of Parkin-dependent mitophagy by Bcl-2-associated athanogene (BAG) family members. Neural Regen. Res. 2021, 16, 684–685. [Google Scholar] [PubMed]
- Sondermann, H.; Scheufler, C.; Schneider, C.; Hohfeld, J.; Hartl, F.U.; Moarefi, I. Structure of a Bag/Hsc70 complex: Convergent functional evolution of Hsp70 nucleotide exchange factors. Science 2001, 291, 1553–1557. [Google Scholar] [CrossRef] [PubMed]
- Tahrir, F.G.; Knezevic, T.; Gupta, M.K.; Gordon, J.; Cheung, J.Y.; Feldman, A.M.; Khalili, K. Evidence for the Role of BAG3 in Mitochondrial Quality Control in Cardiomyocytes. J. Cell Physiol. 2017, 232, 797–805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gamerdinger, M.; Hajieva, P.; Kaya, A.M.; Wolfrum, U.; Hartl, F.U.; Behl, C. Protein quality control during aging involves recruitment of the macroautophagy pathway by BAG3. EMBO J. 2009, 28, 889–901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minoia, M.; Boncoraglio, A.; Vinet, J.; Morelli, F.F.; Brunsting, J.F.; Poletti, A.; Krom, S.; Reits, E.; Kampinga, H.H.; Carra, S. BAG3 induces the sequestration of proteasomal clients into cytoplasmic puncta: Implications for a proteasome-to-autophagy switch. Autophagy 2014, 10, 1603–1621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, C.; Tang, M.; Zeidler, C.; Hohfeld, J.; Johnson, G.V. BAG3 and SYNPO (synaptopodin) facilitate phospho-MAPT/Tau degradation via autophagy in neuronal processes. Autophagy 2019, 15, 1199–1213. [Google Scholar] [CrossRef]
- De Snoo, M.L.; Friesen, E.L.; Zhang, Y.T.; Earnshaw, R.; Dorval, G.; Kapadia, M.; O’Hara, D.M.; Agapova, V.; Chau, H.; Pellerito, O.; et al. Bcl-2-associated athanogene 5 (BAG5) regulates Parkin-dependent mitophagy and cell death. Cell Death Dis. 2019, 10, 907. [Google Scholar] [CrossRef] [Green Version]
- Hasson, S.A.; Kane, L.A.; Yamano, K.; Huang, C.H.; Sliter, D.A.; Buehler, E.; Wang, C.; Heman-Ackah, S.M.; Hessa, T.; Guha, R.; et al. High-content genome-wide RNAi screens identify regulators of parkin upstream of mitophagy. Nature 2013, 504, 291–295. [Google Scholar] [CrossRef]
- Krenciute, G.; Liu, S.; Yucer, N.; Shi, Y.; Ortiz, P.; Liu, Q.; Kim, B.J.; Odejimi, A.O.; Leng, M.; Qin, J.; et al. Nuclear BAG6-UBL4A-GET4 complex mediates DNA damage signaling and cell death. J. Biol. Chem. 2013, 288, 20547–20557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, D.W.; Kim, S.J.; Oh, Y.J.; Choi, B.; Lee, J.; Hwang, I. Arabidopsis BAG1 Functions as a Cofactor in Hsc70-Mediated Proteasomal Degradation of Unimported Plastid Proteins. Mol. Plant 2016, 9, 1428–1431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quintana-Gallardo, L.; Martin-Benito, J.; Marcilla, M.; Espadas, G.; Sabido, E.; Valpuesta, J.M. The cochaperone CHIP marks Hsp70- and Hsp90-bound substrates for degradation through a very flexible mechanism. Sci. Rep. 2019, 9, 5102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, S.; Li, L.; Cui, B.; Men, S.; Shen, Y.; Yang, X. Structural insight into plant programmed cell death mediated by BAG proteins in Arabidopsis thaliana. Acta Crystallogr. D Biol. Crystallogr. 2013, 69, 934–945. [Google Scholar] [CrossRef]
- Locascio, A.; Marques, M.C.; Garcia-Martinez, G.; Corratge-Faillie, C.; Andres-Colas, N.; Rubio, L.; Fernandez, J.A.; Very, A.A.; Mulet, J.M.; Yenush, L. BCL2-ASSOCIATED ATHANOGENE4 Regulates the KAT1 Potassium Channel and Controls Stomatal Movement. Plant Physiol. 2019, 181, 1277–1294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.H.; Xing, Y.F.; Chang, D.; Fang, S.S.; Cui, B.Y.; Li, Q.; Wang, X.J.; Guo, S.; Yang, X.; Men, S.Z.; et al. CaM/BAG5/Hsc70 signaling complex dynamically regulates leaf senescence. Sci. Rep. 2016, 6, 31889. [Google Scholar] [CrossRef]
- Kabbage, M.; Kessens, R.; Dickman, M.B. A plant Bcl-2-associated athanogene is proteolytically activated to confer fungal resistance. Microb. Cell 2016, 3, 224–226. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.R.; Kabbage, M.; Liu, W.D.; Dickman, M.B. Aspartyl Protease-Mediated Cleavage of BAG6 Is Necessary for Autophagy and Fungal Resistance in Plants. Plant Cell 2016, 28, 233–247. [Google Scholar] [CrossRef] [Green Version]
- Williams, B.; Kabbage, M.; Britt, R.; Dickman, M.B. AtBAG7, an Arabidopsis Bcl-2-associated athanogene, resides in the endoplasmic reticulum and is involved in the unfolded protein response. Proc. Natl. Acad. Sci. USA 2010, 107, 6088–6093. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.R.; Williams, B.; Dickman, M. Arabidopsis B-cell lymphoma2 (Bcl-2)-associated athanogene 7 (BAG7)-mediated heat tolerance requires translocation, sumoylation and binding to WRKY29. New Phytol. 2017, 214, 695–705. [Google Scholar] [CrossRef]
- Kabbage, M.; Dickman, M.B. The BAG proteins: A ubiquitous family of chaperone regulators. Cell. Mol. Life Sci. 2008, 65, 1390–1402. [Google Scholar] [CrossRef] [PubMed]
- Ruberti, C.; Lai, Y.; Brandizzi, F. Recovery from temporary endoplasmic reticulum stress in plants relies on the tissue-specific and largely independent roles of bZIP28 and bZIP60, as well as an antagonizing function of BAX-Inhibitor 1 upon the pro-adaptive signaling mediated by bZIP28. Plant J. 2018, 93, 155–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, G.; Wang, S.; Han, S.; Xie, K.; Wang, Y.; Li, J.; Liu, Y. Plant Bax Inhibitor-1 interacts with ATG6 to regulate autophagy and programmed cell death. Autophagy 2017, 13, 1161–1175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Mitophagy pathways in different organisms. Upon stress damage, the ΔφΜ of the mitochondrion is decreased and the mitochondrial outer-membrane permeability (MOMP) is increased. This will lead to mitophagy, which involves the recruitment of the core autophagy-related (ATG) complexes, initiation of phagophore formation, recognition and sequestration of the unwanted mitochondrial materials into the autophagosome. Three types of mitophagy have been described based on the nature of the mitochondrial proteins recognized by the ATG proteins: (A) In animals, mitophagy can be executed through the ubiquitin-dependent, the receptor-dependent and the lipid-dependent pathways. In the ubiquitin-dependent pathway, an E3-Ub ligase is activated and poly-ubiquitinate outer mitochondrial membrane (OMM) substrates, which further recruit adaptor proteins to link the poly-Ub chain and LC3 [24,25,26,27,28,29]; in the receptor-dependent pathway, the receptor proteins directly bind to LC3 to recruit the phagophore [30,31,32,33,34,35,36,37,38], or form a complex together with ULK1 and LC3 [39]; in the lipid-dependent pathway, the lipids interact with LC3, such as cardiolipin [40] and ceramide [41]. (B) In budding yeast, the only identified receptor is ATG32, which is phosphorylated and recognized by ATG11 or ATG8 respectively [42,43,44,45]. Another mitochondrial outer membrane protein, ATG43, serves as the mitophagy receptor in fission yeast [46]. However, the function of cardiolipin is not clear in yeast mitophagy. (C) In plants, only ATG11, which binds to ATG8 directly, has been reported to participate in mitophagy regulation [22]. E3-Ub ligase family members as well as cardiolipin synthase have also been identified in plants to regulate mitochondria biogenesis, but whether they also participate in plant mitophagy remains unknown (question marks).
Figure 1.
Mitophagy pathways in different organisms. Upon stress damage, the ΔφΜ of the mitochondrion is decreased and the mitochondrial outer-membrane permeability (MOMP) is increased. This will lead to mitophagy, which involves the recruitment of the core autophagy-related (ATG) complexes, initiation of phagophore formation, recognition and sequestration of the unwanted mitochondrial materials into the autophagosome. Three types of mitophagy have been described based on the nature of the mitochondrial proteins recognized by the ATG proteins: (A) In animals, mitophagy can be executed through the ubiquitin-dependent, the receptor-dependent and the lipid-dependent pathways. In the ubiquitin-dependent pathway, an E3-Ub ligase is activated and poly-ubiquitinate outer mitochondrial membrane (OMM) substrates, which further recruit adaptor proteins to link the poly-Ub chain and LC3 [24,25,26,27,28,29]; in the receptor-dependent pathway, the receptor proteins directly bind to LC3 to recruit the phagophore [30,31,32,33,34,35,36,37,38], or form a complex together with ULK1 and LC3 [39]; in the lipid-dependent pathway, the lipids interact with LC3, such as cardiolipin [40] and ceramide [41]. (B) In budding yeast, the only identified receptor is ATG32, which is phosphorylated and recognized by ATG11 or ATG8 respectively [42,43,44,45]. Another mitochondrial outer membrane protein, ATG43, serves as the mitophagy receptor in fission yeast [46]. However, the function of cardiolipin is not clear in yeast mitophagy. (C) In plants, only ATG11, which binds to ATG8 directly, has been reported to participate in mitophagy regulation [22]. E3-Ub ligase family members as well as cardiolipin synthase have also been identified in plants to regulate mitochondria biogenesis, but whether they also participate in plant mitophagy remains unknown (question marks).

Figure 2.
Regulation of mitochondrial homeostasis by AMPK/SnRK. (A) In mammals, upon energy stress, AMPK phosphorylates several transcription regulators essential for mitochondrial biogenesis gene expression, including peroxisome proliferator-activated receptor-γ (PPARγ) co-activator 1α (PGC1α), which activates mitochondrial biogenesis genes through interaction with PPARγ or oestrogen-related receptors (ERRs) [99,100,101]. Meanwhile, the transcription factor EB (TFEB), is also activated by AMPK to further induce PGC1α expression upon binding to their promoter region [102]. Mitochondrial outer membrane proteins, including Acetyl-CoA carboxylase 2 (ACC2) and mitochondrial fission factor (MFF), have also been identified as AMPK substrates [103,105]. In addition, AMPK also activates core ATG genes (e.g., ULK1, and Beclin1), which further phosphorylates downstream regulators in mitophagy [106,107,108,109,110,111,113,114,115,116,117,118]. (B) In plants, SnRK1 phosphorylates ATG1 and ATG6 [121,122,123]. In addition, SnRK1 also activates NAC transcription factor (NAC) and basic leucine zipper transcription factors (bZIPs), which are involved in regulating mitochondrial gene expression [124,125,126,127]. However, whether SnRK1 phosphorylates mitochondrial fusion/fission regulators (e.g., elongated mitochondria1, ELM1) is still unknown (dashed lines).
Figure 2.
Regulation of mitochondrial homeostasis by AMPK/SnRK. (A) In mammals, upon energy stress, AMPK phosphorylates several transcription regulators essential for mitochondrial biogenesis gene expression, including peroxisome proliferator-activated receptor-γ (PPARγ) co-activator 1α (PGC1α), which activates mitochondrial biogenesis genes through interaction with PPARγ or oestrogen-related receptors (ERRs) [99,100,101]. Meanwhile, the transcription factor EB (TFEB), is also activated by AMPK to further induce PGC1α expression upon binding to their promoter region [102]. Mitochondrial outer membrane proteins, including Acetyl-CoA carboxylase 2 (ACC2) and mitochondrial fission factor (MFF), have also been identified as AMPK substrates [103,105]. In addition, AMPK also activates core ATG genes (e.g., ULK1, and Beclin1), which further phosphorylates downstream regulators in mitophagy [106,107,108,109,110,111,113,114,115,116,117,118]. (B) In plants, SnRK1 phosphorylates ATG1 and ATG6 [121,122,123]. In addition, SnRK1 also activates NAC transcription factor (NAC) and basic leucine zipper transcription factors (bZIPs), which are involved in regulating mitochondrial gene expression [124,125,126,127]. However, whether SnRK1 phosphorylates mitochondrial fusion/fission regulators (e.g., elongated mitochondria1, ELM1) is still unknown (dashed lines).

Figure 3.
Functions of BAG families in mitochondrial homeostasis and PCD in mammals and plants. (A) Balance between anti-apoptotic and anti-survival Bcl-2 proteins determines the levels of the pro-death and pro-survival pathways. In addition, 6 isoforms of BAG proteins are coded in mammals, among which BAG1 mainly participates in unfolded protein response [133,134]; BAG3 induces autophagy by competing with BAG1 and binding with p62 as well microtubule regulator, or by directly binding with OMM protein to promote autophagy [135,136,137,138]; BAG4 and BAG5 both inhibit mitophagy by suppressing Parkin activity and pro-survival pathway [139,140]; BAG6 forms a ternary complex in the nucleus to promote pro-apoptotic activity via REAPER, and also participates in proteasome-mediated unfolded protein response [141]. (B) In plants, although no homologues of Bcl-2 proteins have been identified, seven isoforms of BAG proteins are found, which have diverse distributions. BAG1-3 has been implicated in binding to HSC70, mediating the unfolded protein response [132,142,143,144]; BAG4 has been shown to interact with the potassium channel KAT1 to participate in stomatal movement [145]; BAG5 is a mitochondrial-localized protein, whose activity is switched by Ca2+ levels via regulating its binding affinity towards CaM and Hsc70 [146]; BAG6 requires a proteolytic process in the vacuole for its activation, which participates in pathogen response and autophagy as well [147,148]; BAG7 is an ER-localized protein, which forms a complex with Hsc70 ortholog BIP protein and bZIP28 upon ER stress [149]. Additionally, after cleavage, BAG7 will undergo SUMOylating and tranlocation to interact with the WRKY29 in the nucleus to regulate the expression of several anti-stress genes [150].
Figure 3.
Functions of BAG families in mitochondrial homeostasis and PCD in mammals and plants. (A) Balance between anti-apoptotic and anti-survival Bcl-2 proteins determines the levels of the pro-death and pro-survival pathways. In addition, 6 isoforms of BAG proteins are coded in mammals, among which BAG1 mainly participates in unfolded protein response [133,134]; BAG3 induces autophagy by competing with BAG1 and binding with p62 as well microtubule regulator, or by directly binding with OMM protein to promote autophagy [135,136,137,138]; BAG4 and BAG5 both inhibit mitophagy by suppressing Parkin activity and pro-survival pathway [139,140]; BAG6 forms a ternary complex in the nucleus to promote pro-apoptotic activity via REAPER, and also participates in proteasome-mediated unfolded protein response [141]. (B) In plants, although no homologues of Bcl-2 proteins have been identified, seven isoforms of BAG proteins are found, which have diverse distributions. BAG1-3 has been implicated in binding to HSC70, mediating the unfolded protein response [132,142,143,144]; BAG4 has been shown to interact with the potassium channel KAT1 to participate in stomatal movement [145]; BAG5 is a mitochondrial-localized protein, whose activity is switched by Ca2+ levels via regulating its binding affinity towards CaM and Hsc70 [146]; BAG6 requires a proteolytic process in the vacuole for its activation, which participates in pathogen response and autophagy as well [147,148]; BAG7 is an ER-localized protein, which forms a complex with Hsc70 ortholog BIP protein and bZIP28 upon ER stress [149]. Additionally, after cleavage, BAG7 will undergo SUMOylating and tranlocation to interact with the WRKY29 in the nucleus to regulate the expression of several anti-stress genes [150].

Figure 4.
Conserved interactions with Hsp70 for Arabidopsis BAG1 or human BAG1. (A) Sequence alignment of BAG domain from Arabidopsis thaliana and Homo sapiens BAG1 protein. Secondary structure and residue numbers were numbered according to Arabidopsis BAG1. Conserved residues forming the interaction surface with the Hsc70 are highlighted in red. (B) Structure comparison of complex of Hsp70 with BAG domain from Arabidopsis (PDB code 4HWI) and humans (PDB code 1HX1). (C) Enlarged schematic diagram of the interactions between BAG domain of Arabidopsis BAG1 and Human Hsp70 from B. (D) Enlarged schematic diagram of the interactions between BAG domain of human BAG1 and Bos taurus Hsp70 from B.
Figure 4.
Conserved interactions with Hsp70 for Arabidopsis BAG1 or human BAG1. (A) Sequence alignment of BAG domain from Arabidopsis thaliana and Homo sapiens BAG1 protein. Secondary structure and residue numbers were numbered according to Arabidopsis BAG1. Conserved residues forming the interaction surface with the Hsc70 are highlighted in red. (B) Structure comparison of complex of Hsp70 with BAG domain from Arabidopsis (PDB code 4HWI) and humans (PDB code 1HX1). (C) Enlarged schematic diagram of the interactions between BAG domain of Arabidopsis BAG1 and Human Hsp70 from B. (D) Enlarged schematic diagram of the interactions between BAG domain of human BAG1 and Bos taurus Hsp70 from B.


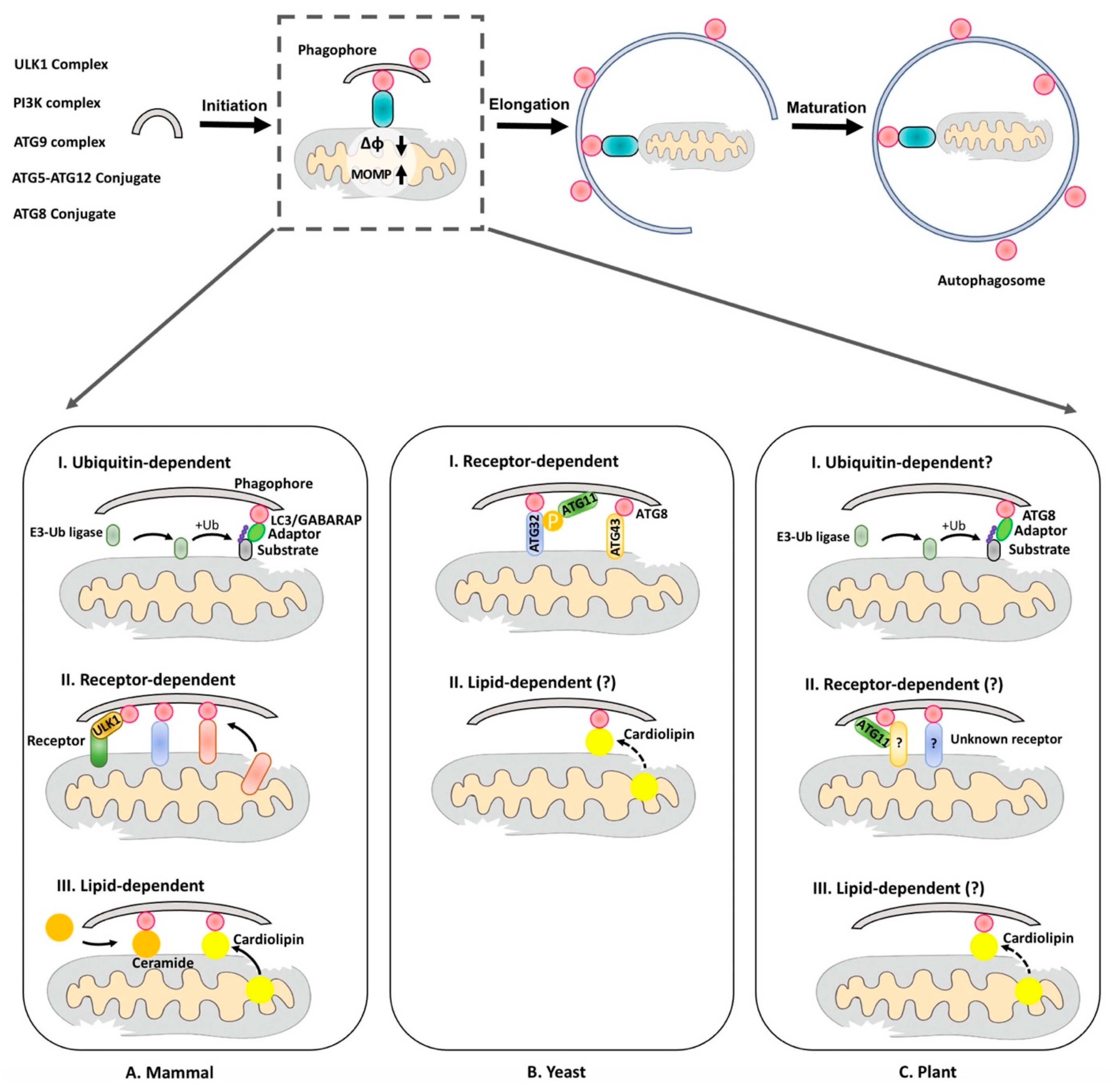{kind=link}
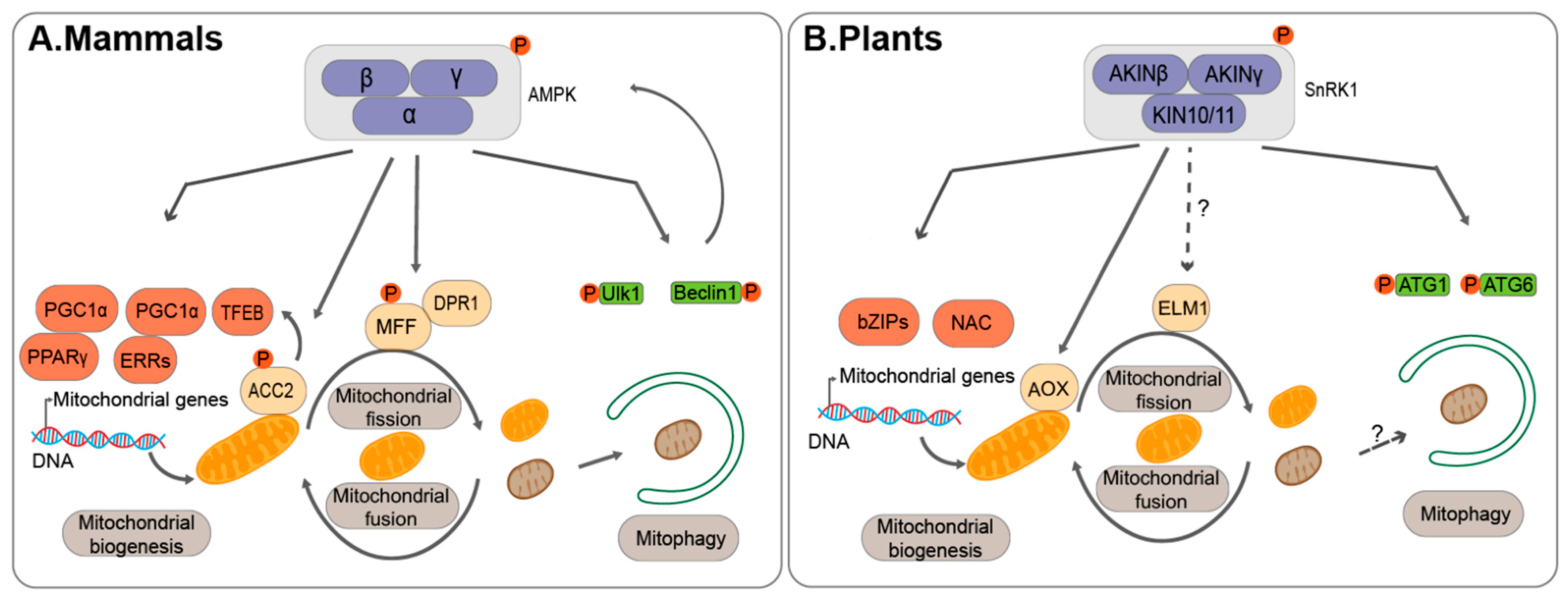{kind=link}
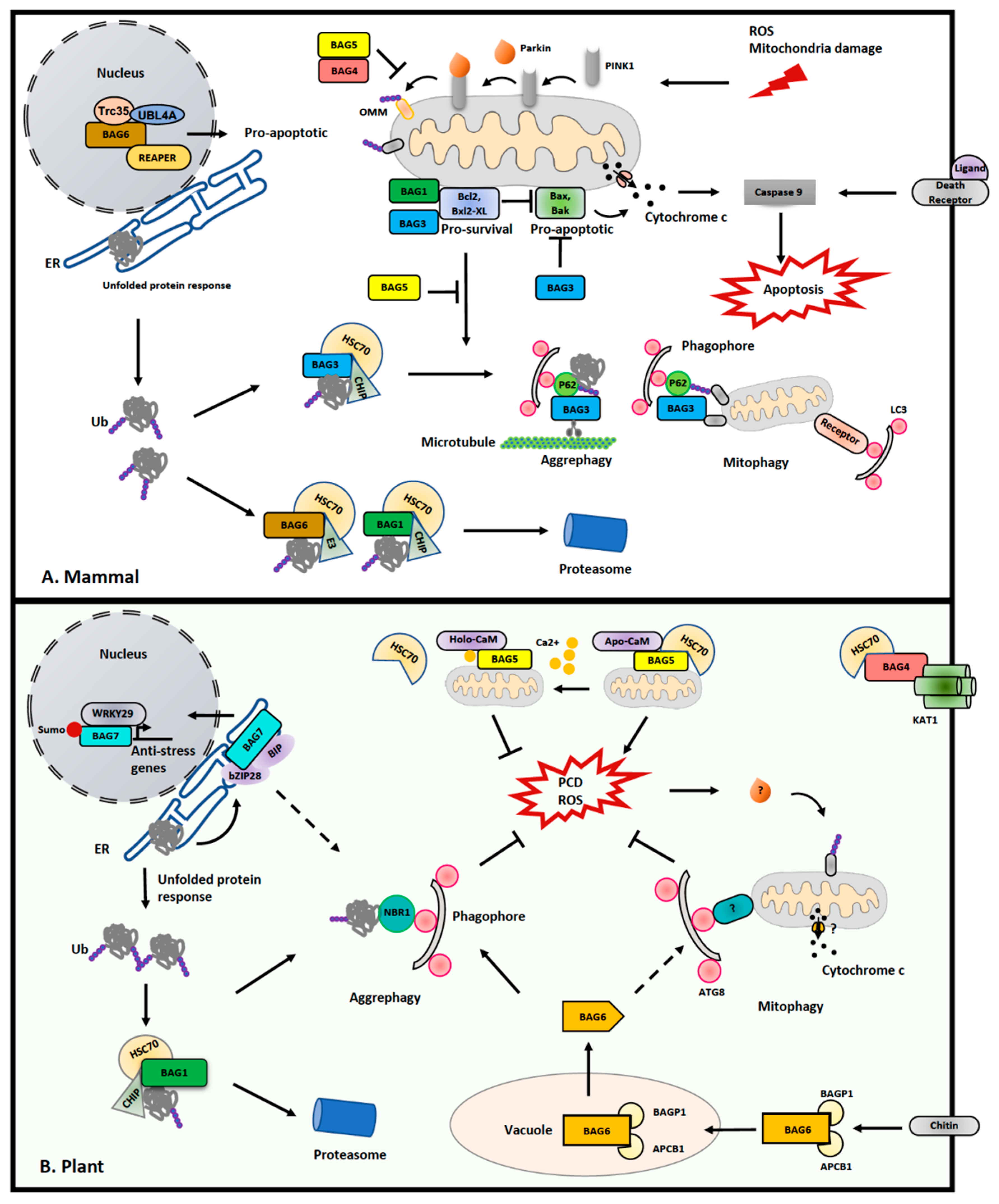{kind=link}
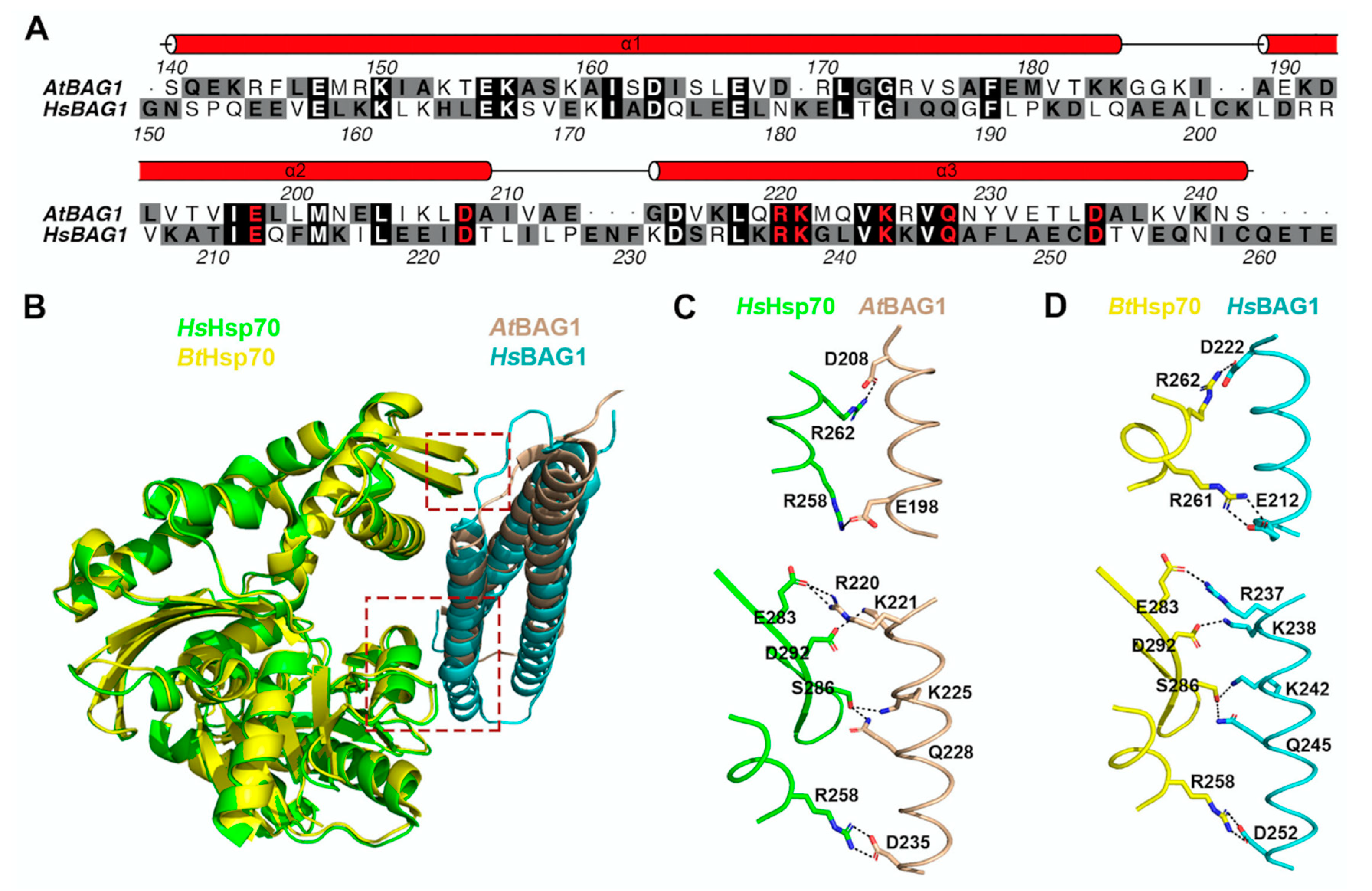{kind=link}
Table 1.
Key mitochondrial fusion and fission factors *.
| Hs | Sc | At | Function in Mitochondrial Dynamic | Phenotype in Plants | References in Plants | |
|---|---|---|---|---|---|---|
| Fusion | MFN1/2 | FZO1 | FZL | Mitochondria-organelle contacts | Abnormal chloroplast and thylakoid morphology | [61] |
| OPA1 | - | - | Inner mitochondrial membrane fusion; Cristae shape maintenance | - | - | |
| IP3R3 | - | - | ER mitochondria contacts | - | - | |
| VDAC1 | POR1 | VDAC1-6 | Swollen and fewer mitochondria | [55,56] | ||
| GRP75 | - | - | - | - | ||
| SAM50 | SAM50 | - | Cristae shape maintenance | - | - | |
| CLUH | CLU1 | FRIENDLY | Mitochondrial fusion | Clustered but not fused mitochondria; defective in mitophagy | [21,62] | |
| Fission | Drp1 | DRP1 | DRP3A; DRP3B | Mitochondrial segregation | Elongated, networked mitochondria | [63,64,65] |
| FIS1 | FIS1 | FIS1A (BIGYIN); FIS1B | Recruitment of dynamins to the outer mitochondrial membrane | Enlarged and fewer mitochondria | [64,66] | |
| MFF | - | - | - | - | ||
| MIEF1/2 (MiD49/51) | - | - | - | - | ||
| - | - | ELM1 | Elongated and fewer mitochondria | [65,67] | ||
| - | - | PMD1; PMD2 | DRP3-independent mitochondrial segregation | Elongated mitochondria | [68] | |
| BIF1 | - | - | Membrane curvature | - | - | |
| MIRO1/2 | - | MIRO1/2/3 | Mitochondrial movement; ER-mitochondria contacts | Elongated mitochondria in miro1 mutant; Few ER-mitochondria contacts in miro2 mutant. | [69,70] |
* Not all regulators are listed. Abbreviations: Hs, Homo sapiens; Sc, Saccharomyces cerevisiae; At, Arabidopsis thaliana.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ren, K.; Feng, L.; Sun, S.; Zhuang, X. Plant Mitophagy in Comparison to Mammals: What Is Still Missing? Int. J. Mol. Sci. 2021, 22, 1236. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031236
AMA Style
Ren K, Feng L, Sun S, Zhuang X. Plant Mitophagy in Comparison to Mammals: What Is Still Missing? International Journal of Molecular Sciences. 2021; 22(3):1236. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031236
Chicago/Turabian StyleRen, Kaike, Lanlan Feng, Shuangli Sun, and Xiaohong Zhuang. 2021. "Plant Mitophagy in Comparison to Mammals: What Is Still Missing?" International Journal of Molecular Sciences 22, no. 3: 1236. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031236
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.