
Immunopathogenicity of Acanthamoeba spp. in the Brain and Lungs


1
Department of Biology and Medical Parasitology, Faculty of Pharmacy, Medical Biotechnology and Laboratory Medicine, Pomeranian Medical University in Szczecin, Powstańców Wielkopolskich 72, 70-111 Szczecin, Poland
2
Independent Laboratory of Pharmaceutical Botany, Faculty of Pharmacy, Medical Biotechnology and Laboratory Medicine, Pomeranian Medical University in Szczecin, Powstańców Wielkopolskich 72, 70-111 Szczecin, Poland
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2021, 22(3), 1261; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031261
Submission received: 12 January 2021
/
Revised: 24 January 2021
/
Accepted: 25 January 2021
/
Published: 27 January 2021
(This article belongs to the Section Biochemistry)
Abstract
:Free-living amoebas, including Acanthamoeba spp., are widely distributed in soil, water, and air. They are capable of causing granulomatous amebic encephalitis, Acanthamoeba pneumonia, Acanthamoeba keratitis, and disseminated acanthamoebiasis. Despite low occurrence worldwide, the mortality rate of Acanthamoeba spp. infections is very high, especially in immunosuppressed hosts. Acanthamoeba infections are a medical problem, owing to limited improvement in diagnostics and treatment, which is associated with incomplete knowledge of pathophysiology, pathogenesis, and the host immune response against Acanthamoeba spp. infection. The aim of this review is to present the biochemical and molecular mechanisms of Acanthamoeba spp.–host interactions, including the expression of Toll-like receptors, mechanisms of an immune response, the activity of metalloproteinases, the secretion of antioxidant enzymes, and the expression and activity of cyclooxygenases. We show the relationship between Acanthamoeba spp. and the host at the cellular level and host defense reactions that lead to changes in the selected host’s organs.
1. Introduction
Acanthamoeba spp. are free-living amoebas (FLA) widely distributed in the environment, including soil, water, and air [1]. They are capable of causing cerebral (granulomatous amebic encephalitis, GAE) and extracerebral (Acanthamoeba keratitis, AK; Acanthamoeba pneumonia, AP; cutaneous acanthamoebiasis and disseminated acanthamoebiasis) life-threatening infections among humans [2]. Acanthamoeba spp. exist as an active (trophozoite) and dormant (cyst) forms. The cysts can abide and survive in various environments for years and transform into trophozoites under favorable environmental conditions [3].
Cerebral and extracerebral infections caused by Acanthamoeba spp. occur mostly among immunocompromised individuals, including HIV positive, organ transplant recipients, patients with chronic diseases, and those who undergo immunosuppressive therapy. Acanthamoebiasis has also been observed in immunocompetent individuals [4,5]. Despite low occurrence worldwide, the mortality rate of Acanthamoeba spp. infections is very high. The pathogenesis of acanthamoebiasis is not fully understood. Acanthamoeba spp. may invade through different ways, such as the respiratory tract or breaks in the skin, resulting in hematogenous dissemination to the brain [6]. Acanthamoeba spp. infections cases are often underdiagnosed and hence strong clinical suspicion along with laboratory technical expertise is required for early diagnosis and therapeutic intervention [7]. The review aims to present the biochemical and molecular mechanisms of Acanthamoeba spp.–host interactions based on the available data.
1.1. Biology of Acanthamoeba spp.
Acanthamoeba spp. exist in two distinct forms: an actively feeding and dividing trophozoite (15–45 μm) and a dormant cyst stage (12–25 μm) [8]. Both stages are infective to humans. Trophozoites of Acanthamoeba spp. actively feed on bacteria, yeast, algae and small organic particles. Cysts are formed after exposure to unfavorable environmental conditions, including changes in humidity, temperature, or environmental pH [4]. Cysts are resistant to many chemical and physical factors, including UV radiation, osmotic pressure, disinfectants, and antiseptics [9,10]. They can survive in the environment for more than 20 years [11].
1.2. Genome of Acanthamoeba spp.
Acanthamoeba species are identified based on the analysis of the diagnostic fragment 3 (DF3) region of the ribosomal DNA gene, designated Acanthamoeba-specific amplimer S1 (ASA.S1) [12]. DF3 encodes the highly variable stem 29-1 of the nuclear small subunit 18S rRNA gene [13]. Acanthamoeba genotypes are distinguished by a 5% or greater sequence difference between isolates [14,15]. To date, 22 sequence types have been identified, designated T1 through T22 [16]. However, Booton et al. [17], examining the DF3 subregion of ASA.S1 in the samples of T3 and T4 isolates, observed several unique sequences within T3 and T4 isolates. These sequences were designated T3/1–T3/5 and T4/1–T4/10 [17]. Due to the fact that different laboratories applied redundant numerical labels to identify different sequences, Fuerst and Booton [16] suggested new subtypes for the T4 genotype, which have been labeled T4A–T4F and T4Neff. The authors also described different sequences within T3 (labeled T3/01–T3/13), T5 (labeled T5/01–T5/15), T11 (labeled T11/01–T11/14), and T15 (labeled T15/01–T15/11). Types T2 and T6 are the most closely related pair of sequence types within Acanthamoeba, and these two types are considered as a supergroup in which five subtypes are formed: T2, T2/6A, T2/6B, T2/6C and T6 [16].
Among all genotypes, T4 is the most frequently isolated from nature, and it includes many pathogenic strains that have been associated with neurological and pulmonary acanthamoebiasis. It is assumed that T4 is characterized by increased virulence and increased resistance to chemotherapeutic agents [18]. The genotypes T1, T2, T4, T5, T10, T11, and T12 are the factors of granulomatous amebic encephalitis [13,19], whereas T2, T4, T5, T16, and T18 may be the factors of Acanthamoeba pneumonia [20,21,22,23].
1.3. Occurrence of Acanthamoeba spp. in the Environment
Acanthamoeba spp. are free-living protozoa that are widely distributed in the environment. Trophozoites and cysts of Acanthamoeba spp. with varying degrees of pathogenicity were found in the water, soil, and air samples. They occur in the rivers, seas, ocean sediments, lakes, ponds, hot springs, water sewage, swimming pools, rainwater, and even in mineral and bottled water [5]. In recent years, Acanthamoeba spp. have been isolated from tap water in Lithuania and city water in Iran [24,25]. In Poland, Derda et al. [26] isolated Acanthamoeba spp. from fountains, and Górnik and Kuźna-Grygiel [27] found the presence of pathogenic Acanthamoeba strains in natural recreational tanks and indoor as well as outdoor swimming pools. Acanthamoeba spp. strains were also isolated from potting soil, dust, air conditioning, dental units and dialysis stations [28]. Besides, amoebas were isolated from vegetables, fruits, mushrooms, but also from biological materials, such as swabs from the nasal mucosa, throat, and purulent secretions from the ear [26]. In Nigeria, the colonization of nasal mucosa by Acanthamoeba spp. was found in 24% of the studied participants [29]. The ubiquity of Acanthamoeba spp. is confirmed by the fact that 80% of the human population has natural IgG antibodies against Acanthamoeba spp. [30].
2. Granulomatous Amebic Encephalitis (GAE)
2.1. Epidemiology
Acanthamoeba spp. causes rare fatal cerebral infection, particularly among immunocompromised patients [31]. The mortality rate of GAE is around 97–98% [4]. In Pubmed, since 1990–2020, 75 cases of patients with GAE caused by Acanthamoeba spp. have been published. Most patients came from the USA, but there were also cases from India, Austria, China, Turkey, Italy, Thailand, Japan, Spain, Germany, Peru, Taiwan, Venezuela, Sweden, Canada, Saudi Arabia, Mexico, Iran and United Kingdom [5]. However, due to non-specific symptoms and diagnostic difficulties, it is presumed that many cases were not recognized or incorrectly diagnosed as bacterial, viral, or fungal infections [32,33,34]. GAE occurs mostly among immunocompromised patients, but immunocompetent cases with GAE have also been reported. Nineteen patients survived cerebral infection of Acanthamoeba spp., among which twelve patients (63%) were immunocompetent (Table 1). All patients received combination therapy, apart from one patient; there is no information about treatment. Risk factors associated with GAE include immunosuppressive states such as acquired immunodeficiency syndrome (AIDS), hematological malignancies, organ transplantation, intake of steroids or other immunosuppressive therapy, systemic lupus erythematosus, diabetes mellitus, and other factors such as prolonged and excessive use of antibiotics, chronic alcoholism, liver cirrhosis, malnutrition, pregnancy, surgical trauma, burns, wounds, and radiation therapy [6,35].
2.2. Symptoms
The nasal cavity is usually the gateway to infestation. The Acanthamoeba spp. infection mostly occurs through inhalation of air or aspiration of water contaminated with an infective stages of these parasites [54]. Trophozoites migrate to the central nervous system (CNS) through the nasal mucosa and the endothelium of the brain’s capillaries [55]. Oral mucosa, damaged or ulcerated skin, and intestinal mucosa can also be sites of infection [56]. It is assumed that penetration into the CNS of the hosts may occur through the bloodstream, mainly from primary lesions in the skin, lungs, or kidneys. The mechanisms involved in the pathogenesis of GAE are not fully understood. Infection is chronic or subacute, with focal necrotic lesions of the brain leading to death 8 days to several months after the onset of symptoms [56]. Clinical symptoms associated with the presence of parasites in the brainstem are as follows: diencephalon or disorientation, lethargy, changes in behavior with hallucinations and irritability, as well as disturbances in taste and smell. The patients also had a headache, especially in the frontal area, neck stiffness, changes in body temperature, seizures, epilepsy, and nausea [9,11].
2.3. Mechanisms Involved in GAE
2.3.1. Immune Responses
The activation of a specific immune response takes time, so in the first days of the infection, non-specific response mechanisms are important as the first line of defense. Toll-like receptors (TLRs) identify highly conserved structural motifs, known as pathogen-associated microbial patterns (PAMPs), which are expressed by microorganisms, or damage-associated molecular patterns (DAMPs) that are endogenous particles released from necrotic or dying cells [57,58]. The main task of TLRs is to activate the cells of the immune system. They are found on the surface of many cells, such as dendritic cells, mast cells, monocytes, neutrophils, and macrophages. In the brain, TLR2 and TLR4 are expressed on microglia, oligodendrocytes, astrocytes, and in neurons [59,60]. The best known TLRs are TLR2 and TLR4. On the basis of experimental studies, it was noted that the ligands for TLR2 are the products of decay of dead cells [61], and the ligands for TLR4 are heat shock proteins, which Acanthamoeba spp. have on their surface [62].
There are far fewer papers concerning the role of TLR in response to parasitic infections. There are also few data on the role of TLR in the innate and acquired responses of the host immune system during acanthamoebiasis. Wojtkowiak-Giera et al. [63] studied the expression of TLR2 and TLR4 in the brain of uninfected and Acanthamoeba spp. infected mice using quantitative polymerase chain reaction (qPCR) and immunohistochemistry (IHC) methods. Using both methods, the uninfected mice showed very similar mRNA expression of TLR2 and TLR4. In the brains of Acanthamoeba spp. infected mice, the TLR2 expression was significantly higher than the TLR4 expression. Wojtkowiak-Giera et al. [63] reported that the TLR2 expression statistically increased at the beginning of infection (at 2 and 4 days post Acanthamoeba spp. infection, dpi) and then decreased (16 and 30 dpi) to the level similar to the uninfected mice. The TLR4 expression in the brains of Acanthamoeba spp. infected mice increased only at 2 dpi, whereas at 4, 16, and 30 dpi, it was at a similar level compared with uninfected mice. Observed changes in the TLR2 and TLR4 expressions in the brain of Acanthamoeba spp. suggest the role of these receptors in the recognition of Acanthamoeba spp. molecules and in the pathomechanism of GAE [63].
The role of TLR receptors in the activation of Th1 (cellular) and Th2 (humoral) responses, acquired immune responses, has been well described in bacterial infections [64,65]. The activation of TLR induces the expression of antigen-presenting cell (APC)-derived cytokines, such as interleukin (IL)-12, IL-23, and IL-27, which promote the differentiation of Th1 cells and inhibit the differentiation of Th2 cells, which preferentially secrete IL-4, IL-5, and IL-13 [66,67]. Relatively few studies have looked at TLR response induction in parasite infections. Parasites can both activate and inhibit the signal across TLRs [68], but the role of these mechanisms in the parasite–host interaction requires further research. Massilamany et al. [69] characterized the inflammatory cells in the brains and spinal cords of Acanthamoeba castellanii infected mice. The infiltrates were comprised of both T cells (~51%) and non-T cells (~49%). Non-T cells were represented by macrophages, dendritic cells, neutrophils, B cells, and natural killer cells. The proportions of CD4 T cells and CD8 T cells were similar [69]. The authors determined the frequencies of Th1, Th2, and Th17 cytokine-producing cells based on intracellular staining by flow cytometry. The analyses included a panel of cytokines: interferon gamma, IFN-γ (Th1 cells), IL-4 and IL-10 (Th2 cells), and IL-17A, IL-17F, and IL-22 (Th17 cells). It was reported that antigen-sensitized lymphocytes from A. castellanii-infected mice contain mostly Th1 cytokine-producing cells; the levels of Th2 and Th17 were similar. The cytokines can be arranged in the following order: IFN-γ> IL-10 > IL-17F > IL-4 or IL-17A or IL-22. The level of IFN-γ-producing cells were three-to-four times higher than Th2 or Th17 cytokine-producing cells. Massilamany et al. [69] showed that Acanthamoeba spp. infection can generate encephalitogenic cross-reactive T cells by antigenic mimicry, and Acanthamoeba antigen-specific T cells are capable of secreting IFN-γ.
2.3.2. Matrix Metalloproteinases (MMPs) and Tissue Inhibitors of MMPs (TIMPs)
MMPs are proteins that play an important role in regulating processes, such as cell differentiation and migration, the regulation of growth factor activity, angiogenesis, and the development of inflammation. MMPs also influence cell survival or its entry into apoptosis, and cell-to-cell communication [70,71]. To date, more than 20 subtypes of MMPs have been identified. The activity and expression of MMPs are controlled mainly (i) by the action of tissue inhibitors of metalloproteinases—TIMPs, (ii) by proenzyme activation, because MMPs are initially synthesized in the form of inactive zymogens, (iii) and by the regulation of gene transcription [72]. Metalloproteinases can be viewed as mediators of nervous system disorders [73]. Increased levels of MMPs are observed in CNS lesions and diseases, such as stroke, multiple sclerosis, and neuropathy; the increased concentration of these enzymes is caused not only by the synthesis from incoming leukocytes, but also by increased synthesis in brain neurons and endothelial cells. In the course of CNS damage, the activity of MMPs is increased under the influence of inflammatory cytokines and chemokines, which causes the interactions between MMPs and nitric oxide (NO), reactive oxygen species (ROS), extracellular matrix (ECM), growth factors and their receptors, integrins and other proteins. The degradation of cytokines by MMPs modulates inflammation, either inducing a worsening and neurotoxic effect or extinguishing it, which in turn promotes cell survival [73,74,75].
Metalloproteinases are found in some parasitic infections of CNS, including Plasmodium falciparum, Trypanosoma brucei spp., and Toxoplasma gondii [76]. In Naegleria fowleri, which is also free-living amoeba with high neurotropism, MMP-2 was found as an integral membrane protein in trophozoites, whereas MMP-9 was either a molecule in the cytosol or a molecule in peripheral membrane. However, the presence of MMPs did not correspond with the virulence of the strain [77]. The secretion of metalloproteinases plays a role in the parasite invasion of the CNS, which elicits the thought that the inhibition of MMPs may hinder the early invasion process [76,77]. To date, it has been reported that Acanthamoeba spp. express and activate several virulence proteins, which allow them to penetrate the biological barriers of the host. Lorenzo-Morales et al. [8] reported that Acanthamoeba spp. produce contact-dependent metalloproteinases during the adhesion to host cells. These proteinases induce the degradation of membranes and host cell death [8].
Imbalances in MMPs and their TIMPs in the selected structures of immunocompetent and immunosuppressed mice were reported by Łanocha-Arendarczyk et al. [78]. The activity of MMPs (−2 and −9) and TIMPs (−1 and −3) in the hippocampus and cerebral cortex were measured by enzyme-linked immunosorbent assay (ELISA) and IHC methods. Immunocompetent Acanthamoeba spp.-infected mice at the beginning of infection showed higher activity of MMP-9 in the cerebral cortex compared to the uninfected group of mice, whereas immunosuppressed Acanthamoeba spp.-infected mice at 24 dpi showed lower MMP-9 levels compared to immunosuppressed uninfected animals. Moreover, MMP-9 level in Acanthamoeba spp.-infected mice differed between the hippocampus and cerebral cortex at 16 dpi. The authors also determined the activity of MMP2 in the cerebral cortex and hippocampus of immunocompetent and immunocompromised hosts, but none of the differences were reported. Łanocha-Arendarczyk et al. [78] found a higher level of TIMP-1 protein in the cerebral cortex of immunocompetent Acanthamoeba spp.-infected mice than in the uninfected mice at 8 dpi, and lower TIMP-3 level in the hippocampus of immunosuppressed Acanthamoeba spp.-infected mice compared to the immunosuppressed uninfected mice at 16 dpi. The MMP-9/TIMP-1 ratio was greater in the hippocampus of the immunosuppressed Acanthamoeba spp.-infected mice than of the uninfected animals at the beginning of infection. A similar trend was reported in Acanthamoeba spp.-infected immunosuppressed mice in an MMP-2/TIMP-3 ratio at 8 dpi, but the difference was not significant. Łanocha-Arendarczyk et al. [78] indicate that the increase in the activity of matrix metalloproteinases during acanthamoebiasis may be primarily the result of the inflammation process, probably an increased activity of proteolytic processes, but also a defense mechanism, preventing the processes of neurodegeneration. Moreover, the authors suggest that MMPs might represent suitable therapeutic targets to prevent the unsealing of the brain–blood barrier in inducted amoebic brain infection [78].
2.3.3. Neurotrophins: Brain-Derived Neurotrophic Factor (BDNF) and Neutrotrophin-4 (NT-4)
Brain-derived neurotrophic factor (BDNF) and neurotrophin-4 (NT-4) are members of the neurotrophin family. BDNF supports the differentiation, maturation, and survival of neurons in the central and peripheral nervous system and shows a neuroprotective effect under adverse conditions, such as hypoglycaemia, cerebral ischemia, neurotoxicity [79]. BDNF stimulates and controls the growth of new neurons from neural stem cells (neurogenesis) [80,81]. BDNF protein and mRNA have been reported in most brain structures, including the hippocampus, cortex, olfactory bulb, and spinal cord [81]. Decreased BDNF levels were observed in neurodegenerative diseases, such as multiple sclerosis [82], Parkinson’s disease [83], and Hungtington’s disease [84]. By contrast, increased BDNF expression was reported in patients with severe cerebral malaria [85] and ocular toxoplasmosis [86]. Łanocha-Arendarczyk et al. [87] determined BDNF levels in the hippocampus and cerebral cortex of immunocompetent and immunocompromised mice infected with Acanthamoeba spp. using the ELISA method. The highest level of BDNF was reported in the hippocampus and cerebral cortex in the mice at the beginning of Acanthamoeba spp. infection (8 dpi). Increased BDNF level in the immunocompetent and immunosuppressed Acanthamoeba spp.-infected mice correlated with neurological symptoms of mice. Along with the duration of infection, the level of BDNF was decreasing in both immunocompetent and immunosuppressed Acanthamoeba spp.-infected mice. Łanocha-Arendarczyk et al. [87] reported no differences in the BDNF level between the hippocampus and cerebral cortex of Acanthamoeba spp.-infected mice and no differences between immunocompetent and immunosuppressed animals. On the basis of the study, it was reported that increased BDNF level in the brain may have neuroprotective effects, and the reduction in the activity of this factor may be related to the progressive process of atrophy and/or neuronal death, usually observed in the cerebral form of acanthamoebiasis [87].
Neurotrophin-4 plays an important role in the development of the nervous system. NT-4 has a similar role to BDNF; it controls the survival and differentiation of vertebrate neurons [88]. NT-4 might also play a role in long-term potentiation and plasticity [89,90]. Chan et al. [91] showed that treatment with NT-4 reduced the infarction volume in a permanent focal cerebral ischemic rat model, demonstrating that NT-4 is involved in ischemic brain injury. Chung et al. [92] reported that NT-4 might participate in the recovery process in brain damage. Łanocha-Arendarczyk et al. [93] determined the activity of NT-4 in the cerebral cortex and hippocampus of mice infected with Acanthamoeba spp. with regard to the immunological status of animals using ELISA method. The highest activity of NT-4 was noticed in the cerebral cortex and hippocampus of immunocompetent and immunosuppressed mice on the 8th day post Acanthamoeba spp. infection. In immunocompetent infected mice, NT-4 activity decreased with the duration of the infection. Łanocha-Arendarczyk et al. [93] reported the differing activity of NT-4 between the cerebral cortex and hippocampus of immunocompetent mice at 16 dpi and immunosuppressed mice at 8 dpi.
2.3.4. Histopathological Changes
Based on the results of the research and the analysis of clinical cases, it was found that Acanthamoeba spp. exhibited the properties of a parasite with noticeable neurotropism. The first experimental studies on the pathogenicity of Acanthamoeba spp. were carried out by Culbertson et al. [94]. The authors found that the intranasal administration of amoebas resulted in the death of mice within 4 days of Acanthamoeba spp. infection. Ulceration of the nasal mucosa was observed in histological preparations, and the presence of amoebas was observed in the frontal lobes of the brain [94]. In another study, Culbertson et al. [95] observed that Acanthamoeba spp., by destroying the nasal mucosa and olfactory bulbs, caused fatal changes in the brain, often accompanied by changes in the lungs. In 1966, Culbertson et al. [96] isolated new strains of Acanthamoeba spp. that induced granulomas forming in the forebrain, mainly in the olfactory bulbs. Histological analysis showed that the occurrence of amoebas in the brain was accompanied by an infiltration of neutrophilic granulocytes, gradually changing into infiltration, consisting of mononuclear cells [96]. Extensive inflammatory lesions of the brain, including the olfactory and frontal lobes, were found in mice infected intranasally with strains of free-living amoebas isolated from lakes in Poznań, central Poland [97]. Diffuse infarctions of nerve and glial cells in the nervous tissue were found [97]. In the study of histological changes in the brain of mice infected with amoebas isolated from water reservoirs in the Lublin region (east part of Poland), Gieryng and Gieryng [98] observed general cerebral congestion with large ecchymoses in the area of the frontal and parietal lobes and the cerebellum. There were small pus spots in the superficial substance of the cerebral cortex and in the olfactory bulbs, which were usually clearly red and showed necrotic lesions. Histological and morphological analysis of the brain of mice infected intranasally with environmental Acanthamoeba strains from northwestern Poland revealed swelling, shallowing of the furrows, and severe congestion of the meninges [27]. Moreover, the detachment of the meninges from the cortical part of the cerebral hemispheres was observed, mainly in the area of the frontal lobes [27]. It was found that there is a close relationship between the severity of inflammation in the brain and the survival time of the mice. In animals that died in the first days post Acanthamoeba spp. infection, slight brain lesions were observed, while in mice that survived 10–17 days, the greatest level of brain damage was observed [98]. Similarly, Rucka [97] found that the degree of intensification of inflammatory processes in the brain depended on the survival time of mice experimentally infected with amoebic strains. It was also noted that only trophozoites of Acanthamoeba spp. appeared in the brain of animals infected with amoebas [98], while Martinez [99] observed both trophozoites and cysts of Acanthamoeba spp. in the brain of a patient with granulomatous encephalitis.
Histopathological examinations of the brains of patients with Acanthamoeba spp. infections showed cerebral edema and numerous necrotic as well as hemorrhagic areas. The brainstem, hemispheres, and cerebellum showed effusions and inflammatory infiltrates consisting of neutrophils, macrophages, mononuclear cells, and multinuclear giant cells. Necrotizing arteritis, selective accumulation of trophozoites and cysts of Acanthamoeba spp. in the perivascular spaces were also observed. The cerebrospinal fluid (CSF) examination revealed lymphocyte-predominant pleocytosis, elevated protein levels, and normal or low glucose levels [2,100,101].
All described mechanisms in the Acanthamoeba spp. infected brain are presented in Figure 1.
3. Acanthamoeba pneumonia (AP)
3.1. Epidemiology
So far, 19 case reports of Acanthamoeba pneumonia (AP) or disseminated acanthamoebiasis with lung infection have been published. Most patients came from the USA, but there were also cases from Poland, Austria, France, Korea, Japan, and India (Table 2). None of the patients survived.
3.2. Symptoms
Acanthamoeba spp. infection to the lungs occurs mostly in patients with a low immune response [102,104,112]. The pathomechanism of AP is not clear. Acanthamoeba pneumonia was found in patients not only as a site of Acanthamoeba spp. infection [104,112] but also as a disseminated infection [102]. In patients with AP, a decrease in body weight and respiratory efficiency was observed, and in the radiological examination, interstitial changes with visible pulmonary edema were observed [112]. Martinez [105] found multiple nodular lesions in the patient’s lungs infected with A. castellanii. Additionally, it has been noticed that the infection of amoebas into the lungs may be bilateral with patchy infiltrates [102]. Im and Kim [112] noticed changes in peripheral blood parameters in a 7-month-old AP patient. During the infection, there was a decreasing trend in the level of hemoglobin (15.0–11.1 mg/dl), hematocrit (48.0–37.6%), and a decrease in the number of white blood cells (10,900–5460/mm3) and platelets (253–65/103). However, no changes were found in the sodium and chloride levels [112]. Infection is most often diagnosed post mortem by isolating amoebas from bronchoaspirate fluid, bronchoalveolar lavage (BAL), bronchial washing, and a fragment of lung tissue obtained during a biopsy [20,102,103,104,114].
3.3. Mechanisms Involved in AP
3.3.1. Immune Responses
TLRs are components of host defense activation in the case of infectious and non-infectious pulmonary disorders, such as acute lung injury, interstitial lung diseases, asthma, and chronic obstructive pulmonary disease. Receptors also play an important role in lung cancer [115,116]. Derda et al. [117] determined the TLR2 and TLR4 expressions in the lungs of Acanthamoeba spp.-infected mice using qPCR and IHC methods. The authors found that the TLR2 expression was higher in mice at 2, 4, 16, and 30 days post Acanthamoeba spp. infection than the TLR4 expression. The authors reported an increased expression of both receptors from 2 to 30 dpi, compared with the uninfected mice. Statistically significant differences between the uninfected and Acanthamoeba spp. infected mice were observed in TLR2 and TLR4 expressions at 16 dpi [117]. Derda et al. [117] showed that TLR2 and TLR4 were upregulated in the lungs of Acanthamoeba spp. infected mice and TLRs were located in the interstitial cells, pneumocytes, and the apical portion of epithelial cells of the bronchial tree. The authors suggested that TLRs are involved in the recognition of Acanthamoeba spp. in the lungs.
PAMPs are recognized by a diverse set of pattern recognition receptors (PRRs), such as Toll-like receptors, C-type lectin receptors, and protease-activated receptor (PAR) 2. The immune response may be polarized towards Th1, Th2, Th17, or Treg lymphocytes. Mechanisms of inducing Th-1 mediated by TLRs are well understood [118], whereas the mechanisms triggering other types of responses are not clear. In the case of a Th2-mediated response, there are two hypotheses concerning the mechanisms that trigger this response: (i) it develops in the absence or incomplete induction of Th1 inflammation, or (ii) there are distinct receptors and signaling pathways responsible for recognizing PAMP pathogens, including the Th2 response [119].
An airway allergic reaction and the presence of parasites in the host organism often causes an increased synthesis of immunoglobulin E (IgE) and Th2 interleukins, such as IL-4, IL-5, and IL-13. These cytokines lead to the recruitment of basophils, eosinophils, and mast cells. As a consequence, this mechanism causes damage to the parasite and its expulsion from the host organism [120,121]. Park et al. [122] determined the activity of several chemokines in the bronchoalveolar lavage fluid (BALF) of mice infected with low (100 trophozoites) and high dose (4 × 105 trophozoites) of A. lugdunensis. In mice infected with a high dose of trophozoites—the levels of IL-4, IL-5, and IL-13 in BALF increased compared to uninfected animals. The IL-17 levels in BALF of A. lugdunensis-infected mice did not change. It was found that A. lugdunensis excretory–secretory (ES) proteins induced the activation of dendric cells (DCs), which accelerated the differentiation of naive CD4+ T cells into Th2 cells [122]. The authors also determine whether PAR2 mediates the airway inflammation induced by proteins that are expressed on Acanthamoeba spp. trophozoites. PAR2 knockout and wild-type mice were inoculated by Acanthamoeba ES proteins. The number of eosinophils and neutrophils was higher in the ES protein administered hosts than in uninfected mice. The lungs of PAR2 knockout and wild-type mice-treated ES proteins showed the hyperplasia of lung epithelial cells, enhanced mucin production, and immune cell infiltration around the bronchial tracts. The cytokine levels, such as IL-4, IL-5, and IL-13, increased in BALF of mice; nonetheless, the levels of studied cytokines were lower in the PAR2 knockout mice. Park et al. [122] suggest that Acanthamoeba trophozoites promoted Th2 responses by activating DCs via PAR2 signaling in the lungs of hosts.
3.3.2. Cyclooxygenases (COXs)
The inflammatory process in the lungs increases the expression of many inflammatory proteins, including cytokines, chemokines, and adhesion molecules. It can also change the activity of enzymes related to the synthesis of eicosanoids, which play the role of the most peripherally located secondary informants in the cell [123,124]. These compounds locally in the cell affect the activity of hormones and neurotransmitters, and the profile of eicosanoids formed depends on the enzymatic composition of individual cells. Cyclooxygenases (COX) are responsible for the formation of prostaglandins, including strongly pro-inflammatory PGE2 and thromboxane (TXB2), which is an activator of platelet aggregation, a factor promoting vasoconstriction, and a strong mitogen of smooth muscles [125,126]. Cyclooxygenase, also known as prostaglandin H synthase, is an enzyme that catalyzes the conversion of arachidonic acid to prostaglandins (PGs), prostacyclins (PGIs), and thromboxanes (TXBs). In the human body, it exists in the form of two isoforms—cyclooxygenase-1 (COX-1) and cyclooxygenase-2 (COX-2) [124]. COX-1 is constantly expressed in almost all tissues and is responsible for maintaining the homeostasis of the organism, e.g., by the cytoprotection of mucous membranes, platelet aggregation, and the maintenance of normal renal blood flow. The presence of COX-1 has been demonstrated in both the upper and lower respiratory tracts, as well as in the pleura [127,128]. COX-2 is an inducible enzyme that is active in cells involved in pathological processes, such as the onset and maintenance of acute and chronic inflammation, pain, fever, and cancer. COX-2 expression has been reported to be associated with inflammation or proliferative changes in the airway epithelium, including pneumonia and lung cancer [129]. COX-2 is induced by parasitic infections, but the mechanism is not clear [130].
An experimental study shows that COX-1 and COX-2 are important mediators of the AP [131]. Łanocha-Arendarczyk et al. [131] aimed to check the impact of Acanthamoeba spp. infection on the transformation of arachidonic acid and the production of eicosanoids (PGs and TXs) in the lungs of immunocompetent and immunosuppressed mice using IHC and Western blot methods. Molecular analysis showed that the strong expression of COX-1 and COX-2 in the lungs of the immunocompetent host induced by Acanthamoeba spp. did not correspond to the significant differences in the levels of PGE2 and TXB2. It is likely that the inflammatory process caused by the parasite initiated the immune response, consisting of the inhibition of the enzyme activity by its product. The highest expressions of COX-1 and COX-2 proteins in the lungs were observed in 24 days post Acanthamoeba spp.-infected immunocompetent mice, and they were, respectively, 44% and 80% higher than in the uninfected animals. In contrast, steroid-induced immunosuppression in Acanthamoeba spp.-infected mice reduced the COX-1 and COX-2 expressions in lung, but not at the beginning of the infection. Moreover, the analysis of IHC showed a decrease in the detection of COX-1 in the Acanthamoeba spp.-infected immunosuppressed mice, compared to the Acanthamoeba spp.-infected immunocompetent mice. Łanocha-Arendarczyk et al. [131] concluded that Acanthamoeba spp. increase the expression and activity of COX-1 and COX-2 in the lungs of the hosts. Moreover, the authors [131] suggested that increased expression may be a result of a direct regulatory function of pulmonary epithelial cells and the release of pro-inflammatory cytokines from alveolar macrophages, or it may be related to increased levels of free radicals and decreased antioxidant enzyme activity in pulmonary acanthamoebiasis.
3.3.3. Antioxidant Defense
An intense or long-lasting state of oxidative stress may induce pathological reactions, leading to cell and tissue damage. The biological consequences of the pro and antioxidant imbalance include protein denaturation, lipid peroxidation, carbohydrate damage, changes in DNA structure leading to mutations, or cytotoxic effects [132]. Increasing oxidative damage in cells is the molecular basis of the development of many diseases, also in the inflammatory processes of the lungs [133]. The violation of the oxidation-reduction potential balance also plays a role in the pathogenesis of parasitic diseases.
The parasite–host interactions are influenced by the defense mechanisms of the parasite and the host. Parasites have developed mechanisms to avoid the effects of ROS. Protozoa and helminths have at least one of the three main antioxidant enzymes that play a key role in defense response against the host’s reaction [134]. Two forms of superoxide dismutases (Fe-SOD and Cu, Zn-SOD) have been detected in Acanthamoeba spp. [8]. Dismutases in Acanthamoeba spp. may not only protect them from oxidative stress but also protect against ROS released by the cells of the immune system upon contact with amoebas [135,136]. Moreover, a correlation between the activity of glutathione peroxidase (GPx) in Acanthamoeba spp. and the invasiveness of these amoebas was established. The role of catalase (CAT) in the antioxidant defense system of Acanthamoeba spp. was not found [137]. Despite the presence of two antioxidant enzymes (SOD and GPx) in Acanthamoeba spp., it has been found that the increased production of ROS inhibits development and, consequently, degrades the amoeba cells [138].
Łanocha-Arendarczyk et al. [139] assessed the severity of oxidative stress by measuring the concentration of lipid peroxidation products in the lungs of mice infected with the Acanthamoeba AM 22 strain and assessed the effectiveness of protection against oxidative stress by measuring the activity of antioxidant enzymes (SOD, CAT, glutathione reductase (GR), GPx) regarding the immune status of the host. The study showed a significant increase in lipid peroxidation products (LPO) in immunocompetent and immunocompromised mice at 8- and 16-days post Acanthamoeba spp. infection, compared to uninfected mice. It was also reported that, at the beginning of the infection, the LPO levels were significantly different between immunocompetent and immunocompromised mice infected with Acanthamoeba spp. A significant increase in LPO was demonstrated in Acanthamoeba spp. infected immunocompetent mice compared to Acanthamoeba spp. infected immunocompromised mice. The study has shown that the Acanthamoeba spp. infection of mice is accompanied by changes in the activity of antioxidant enzymes in the lungs during the course of the infection. In immunocompromised Acanthamoeba spp.-infected mice, significantly lower activity of SOD in the lungs was found compared to the uninfected immunocompromised mice at 8 dpi. In the lungs of immunocompetent hosts, a significant increase in CAT activity was noted at 16 days post Acanthmoeba spp. infection. It was shown that the activity of this enzyme was significantly higher in immunocompetent Acanthamoeba spp.-infected mice than in immunocompetent hosts at 16 dpi. The study also reported a decrease in GR activity in the lungs of immunosuppressed Acanthamoeba spp.-infected vs. uninfected mice at 24 dpi. At 8 dpi, in the lungs of immunocompetent and immunocompromised mice, a decrease in GPx activity was found compared to the control groups, as well as a significantly lower activity of this enzyme was noted in the immunocompromised mice compared to the control group in 16 dpi, which could result from the depletion of the body’s antioxidant abilities during an intensified inflammatory process taking place in the lung tissue [139].
The increased activity of the immune system during Acanthamoeba spp. infection may lead to a shift of the antioxidant balance in the body towards oxidation, which may initiate or aggravate inflammation. An effect of the inflammatory response in immunocompetent and immunocompromised hosts in the course of the experimental Acanthamoeba spp. infection is a reduction in the antioxidant potential of the lungs, resulting from changes in the activity of antioxidant enzymes. In pulmonary acanthamoebiasis, the changes in the activity of antioxidant enzymes may be a consequence of oxidative stress, which is manifested by an increase in the concentration of lipid peroxidation products [139].
3.3.4. Histopathological Changes
In animal studies, Culbertson et al. [94] observed Acanthamoeba spp. accompanied by a strong fibrin-purulent exudate reaction in the lungs of mice after the intranasal administration of amoebas. Hemorrhage was also commonly observed. The changes also included pulmonary veins with numerous clots and embolisms. In other studies, Culbertson et al. [95] observed that low doses of amoebas caused animals death due to lung lesions. Rucka [97] found the infiltration of plasma and mononuclear cells, with the destruction of the vessel wall and bronchus, in the perivascular and peribronchial tissue in the histological examination of the lungs of mice inoculated with free-living amoebas. Large exudative cells and phlegmon-containing amoebas were present in the lung tissue. Moreover, numerous foci of necrosis and purulent inflammation were observed in the lung parenchyma [97]. Gieryng et al. [140] found the destruction of the bronchial epithelium, bronchioles, and endothelium in blood vessels, to a large extent in the histological examination of the lungs of experimentally Acanthamoeba spp.-infected mice. The presence of Acanthamoeba spp. and small hemorrhages were observed at the sites of vascular and bronchial damage. The walls of the alveoli became thickened, and in some places, the destruction of the walls by Acanthamoeba spp. was present. As the disease progressed, pathological changes in the lungs worsened [140]. Górnik and Kuźna-Grygiel [27] noticed the hyperplasia of the bronchiolar epithelium, thickening, and congestion of the alveolar walls in the lung of Acanthamoeba spp.-infected mice. Park et al. [122] reported massive inflammatory cell infiltration, bronchial epithelial cell hyperplasia, and goblet cell hyperplasia in the lungs of mice infected with a high dose of Acanthamoeba spp. (4 × 105 trophozoites). In the lungs infected with a low dose of Acanthamoeba spp. (100 trophozoites), Park et al. [122] observed mild infiltration of inflammation in the lungs and the hyperplasia of some bronchial epithelial cells. Some forms of Acanthamoeba spp. were observed in alveoli, and several eosinophils were noticed around amoebas.
All described mechanisms in the Acanthamoeba spp.-infected lungs are presented in Figure 2.
4. Treatment
Currently, there is no reliably effective drug therapy for Acanthamoeba spp. infections of the brain and lungs. Most published cases have used multiple drug combinations to provide synergistic effects and improved treatment results. Clinical cases reporting successful treatment outcomes remain limited because of the inability of most drugs to overcome the biological barriers and to penetrate into the parenchyma of the brain or lungs with sufficient concentrations to kill the amoeba. The FDA-approved drugs used in Acanthamoeba spp. infections are amphotericin B, azithromycin, paromomycin, itraconazole, fluconazole, ketoconazole, pentamidine, sulfadiazine, cotrimoxazole, 5-flucytosine, rifampicin, and pyrimethamine [141].
The Infectious Diseases Society of America (IDSA) guidelines for Acanthamoeba CNS infection have a recommendation for either fluconazole+ sulfadiazine+ pyrimethamine or trimethoprim-sulfamethoxazole+ rifampicin+ ketoconazole [142]. The Centers for Disease Control and Prevention (CDC) announced the availability of anticancer and anti-leishmaniasis drug miltefosine as a drug to treat CNS infection [143]. Table 1 presents treatment that was given to survivors of GAE; miltefosine as a part of combination therapy was used in six cases. Webster et al. [52] found that miltefosine and oral voriconazole reduced serological titers and brain lesions in immunocompetent patients with GAE. In AP, amphotericin B in combination with trimethoprim, miltefosine, voriconazole, pentamidine, 5-fluorocytosine, and itraconazole were used [21,22,102]. It is important to point out that patients with pulmonary acanthamoebiasis often receive therapy that is targeted on viral, bacterial, and fungal infections instead of Acanthamoeba spp. infection.
Among the potential drugs that may have possible effects on Acanthamoeba spp. infection, Elsheikha et al. [141] presented and described amlodipine, loperamide, prochlorperazine, a combination of chlorpromazine and rokitamycin, procyclidine, digoxin, corifungin, tigecycline, heterocyclic alkylphosphocholines (structural analogue of miltefosine) and chloroquine.
5. Conclusions and Future Research Perspectives
The biochemical and molecular mechanisms known to date that are involved in the response to Acanthamoeba spp. infection in the hosts provide valuable information on the pathogenesis and factors that affect the course of cerebral and pulmonary acanthamoebiasis. The presented mechanisms, to a small extent, allow us to understand the interaction between Acanthamoeba spp. and the hosts. The parasite caused the inflammation in the organs through the host’s immunological reactions, changes in antioxidant defense, metalloproteinases and cyclooxygenases activity. Owing to the fact that Acanthamoeba spp. are involved in a variety of processes in the host’s organism, it can be concluded that the parasite may also affect other mechanisms than those presented in this review. In an attempt to understand all the mechanisms in acanthamoebiasis, future research is needed to describe the molecular pathways of the immune response. Understanding the mechanisms of acanthamoebiasis pathogenesis may contribute to developing an effective pharmacological therapy for these infections and to limit the degree of cell damage.
Author Contributions
K.K.: literature search and review, manuscript drafts preparation, writing manuscript, and preparation of manuscript revision. N.Ł.-A.: literature search and review, participation in writing the manuscript. D.K.-B.: participation in writing the manuscript, preparation of manuscript revision, final acceptance of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lass, A.; Guerrero, M.; Li, X.; Karanis, G.; Ma, L.; Karanis, P. Detection of Acanthamoeba spp. in water samples collected from natural water reservoirs, sewages, and pharmaceutical factory drains using LAMP and PCR in China. Sci. Total Environ. 2017, 584–585, 489–494. [Google Scholar] [CrossRef] [PubMed]
- Marciano-Cabral, F.; Cabral, G. Acanthamoeba spp. as agents of disease in humans. Clin. Microbiol. Rev. 2003, 16, 273–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juárez, M.M.; Tártara, L.I.; Cid, A.G.; Real, J.P.; Bermúdez, J.M.; Rajal, V.B.; Palma, S.D. Acanthamoeba in the eye, can the parasite hide even more? Latest developments on the disease. Cont. Lens. Anterior Eye 2018, 41, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Kot, K.; Łanocha-Arendarczyk, N.A.; Kosik-Bogacka, D.I. Amoebas from the genus Acanthamoeba and their pathogenic properties. Ann. Parasitol. 2018, 64, 299–308. [Google Scholar] [CrossRef]
- Kalra, S.K.; Sharma, P.; Shyam, K.; Tejan, N.; Ghoshal, U. Acanthamoeba and its pathogenic role in granulomatous amebic encephalitis. Exp. Parasitol. 2020, 208, 107788. [Google Scholar] [CrossRef]
- Duggal, S.D.; Rongpharpi, S.R.; Duggal, A.K.; Kumar, A.; Biswal, I. Role of Acanthamoeba in granulomatous encephalitis: A review. J. Infect. Dis. Immune Ther. 2017, 1, 2. [Google Scholar]
- Parija, S.C.; Dinoop, K.; Venugopal, H. Management of granulomatous amebic encephalitis: Laboratory diagnosis and treatment. Trop. Parasitol. 2015, 5, 23–28. [Google Scholar] [CrossRef] [Green Version]
- Lorenzo-Morales, J.; Khan, N.A.; Walochnik, J. An update on Acanthamoeba keratitis: Diagnosis, pathogenesis and treatment. Parasite 2015, 22, 10. [Google Scholar] [CrossRef] [Green Version]
- Khan, N.A. Acanthamoeba: Biology and increasing importance in human health. FEMS Microbiol. Rev. 2006, 30, 564–595. [Google Scholar] [CrossRef] [Green Version]
- Lorenzo-Morales, J.; Martín-Navarro, C.M.; López-Arencibia, A.; Arnalich-Montiel, F.; Piñero, J.E.; Valladares, B. Acanthamoeba keratitis: An emerging disease gathering importance worldwide? Trends Parasitol. 2013, 29, 181–187. [Google Scholar] [CrossRef]
- Visvesvara, G.S. Infections with free-living amebae. Handb. Clin. Neurol. 2013, 114, 153–168. [Google Scholar] [CrossRef] [PubMed]
- Taher, E.E.; Méabed, E.M.H.; Abdallah, I.; Abdel Wahed, W.Y. Acanthamoeba keratitis in noncompliant soft contact lenses users: Genotyping and risk factors, a study from Cairo, Egypt. J. Infect. Public Health 2018, 11, 377–383. [Google Scholar] [CrossRef] [PubMed]
- Megha, K.; Sehgal, R.; Khurana, S. Genotyping of Acanthamoeba spp. isolated from patients with granulomatous amoebic encephalitis. Indian J. Med. Res. 2018, 148, 456–459. [Google Scholar] [CrossRef] [PubMed]
- Gast, R.J.; Ledee, D.R.; Fuerst, P.A.; Byers, T.J. Subgenus systematics of Acanthamoeba: Four nuclear 18S rDNA sequence types. J. Eukaryot. Microbiol. 1996, 43, 498–504. [Google Scholar] [CrossRef]
- Fuerst, P.A.; Booton, G.C.; Crary, M. Phylogenetic analysis and the evolution of the 18S rRNA gene typing system of Acanthamoeba. J. Eukaryot. Microbiol 2015, 62, 69–84. [Google Scholar] [CrossRef] [PubMed]
- Fuerst, P.A.; Booton, G.C. Species, Sequence Types and Alleles: Dissecting Genetic Variation in Acanthamoeba. Pathogens 2020, 9, 534. [Google Scholar] [CrossRef]
- Booton, G.C.; Kelly, D.J.; Chu, Y.W.; Seal, D.V.; Houang, E.; Lam, D.S.; Byers, T.J.; Fuerst, P.A. 18S ribosomal DNA typing and tracking of Acanthamoeba species isolates from corneal scrape specimens, contact lenses, lens cases, and home water supplies of Acanthamoeba keratitis patients in Hong Kong. J. Clin. Microbiol. 2002, 40, 1621–1625. [Google Scholar] [CrossRef] [Green Version]
- Siddiqui, R.; Khan, N.A. Biology and pathogenesis of Acanthamoeba. Parasit. Vectors 2012, 10, 6. [Google Scholar] [CrossRef] [Green Version]
- Behera, H.S.; Panda, A.; Satpathy, G.; Bandivadekar, P.; Vanathi, M.; Agarwal, T.; Nayak, N.; Tandon, R. Genotyping of Acanthamoeba spp. and characterization of the prevalent T4 type along with T10 and unassigned genotypes from amoebic keratitis patients in India. J. Med. Microbiol. 2016, 65, 370–376. [Google Scholar] [CrossRef]
- Łanocha, N.; Kosik-Bogacka, D.; Maciejewska, A.; Sawczuk, M.; Wilk, A.; Kuźna-Grygiel, W. The occurence Acanthamoeba (Free Living Amoeba) in environmental and respiratory samples in Poland. Acta Protozool. 2009, 48, 271–279. [Google Scholar]
- Aichelburg, A.C.; Walochnik, J.; Assadian, O.; Prosch, H.; Steuer, A.; Perneczky, G.; Visvesvara, G.S.; Aspöck, H.; Vetter, N. Successful treatment of disseminated Acanthamoeba sp. infection with miltefosine. Emerg. Infect. Dis. 2008, 14, 1743–1746. [Google Scholar] [CrossRef] [PubMed]
- Barete, S.; Combes, A.; de Jonckheere, J.F.; Datry, A.; Varnous, S.; Martinez, V.; Ptacek, S.G.; Caumes, E.; Capron, F.; Francès, C.; et al. Fatal disseminated Acanthamoeba lenticulata infection in a heart transplant patient. Emerg. Infect. Dis. 2007, 13, 736–738. [Google Scholar] [CrossRef] [PubMed]
- Matsui, T.; Maeda, T.; Kusakabe, S.; Arita, H.; Yagita, K.; Morii, E.; Kanakura, Y. A case report of granulomatous amoebic encephalitis by Group 1 Acanthamoeba genotype T18 diagnosed by the combination of morphological examination and genetic analysis. Diagn. Pathol. 2018, 13, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gavarāne, I.; Trofimova, J.; Mališevs, A.; Valciņa, O.; Kirjušina, M.; Rubeniņa, I.; Bērziņš, A. DNA extraction from amoebal isolates and genotype determination of Acanthamoeba from tap water in Latvia. Parasitol. Res. 2018, 117, 3299–3303. [Google Scholar] [CrossRef] [PubMed]
- Javanmard, E.; Niyyati, M.; Lorenzo-Morales, J.; Lasjerdi, Z.; Behniafar, H.; Mirjalali, H. Molecular identification of waterborne free living amoebae (Acanthamoeba, Naegleria and Vermamoeba) isolated from municipal drinking water and environmental sources, Semnan province, north half of Iran. Exp. Parasitol. 2017, 183, 240–244. [Google Scholar] [CrossRef] [PubMed]
- Derda, M.; Hadaś, E.; Wojtkowiak-Giera, A.; Wojt, W.J.; Cholewiński, M.; Skrzypczak, Ł. The occurrence of free-living amoebae in fountains. Probl. Hig. Epidemiol. 2013, 94, 147–150. [Google Scholar]
- Górnik, K.; Kuźna-Grygiel, W. Histological studies of selected organs of mice experimentally infected with Acanthamoeba spp. Folia Morphol. 2005, 64, 161–167. [Google Scholar]
- Łanocha-Arendarczyk, N.; Kosik-Bogacka, D.; Galant, K.; Zaorski, W.; Kot, K.; Łanocha, A. Pathogenic free-living amoeba. Post Mikrobiol. 2017, 1, 106–112. [Google Scholar]
- Clarke, B.; Sinha, A.; Parmar, D.N.; Sykakis, E. Advances in the diagnosis and treatment of Acanthamoeba keratitis. J. Ophthal Mol. 2012, 2012, 484–892. [Google Scholar] [CrossRef] [Green Version]
- Brindley, N.; Matin, A.; Khan, N.A. Acanthamoeba castellanii: High antibody prevalence in racially and ethnically diverse populations. Exp. Parasitol. 2009, 121, 254–256. [Google Scholar] [CrossRef]
- Zamora, A.; Henderson, H.; Swiatlo, E. Acanthamoeba encephalitis: A Case Report and Review of Therapy. Surg. Neurol. Int. 2014, 5, 68. [Google Scholar] [CrossRef] [PubMed]
- Yagi, S.; Schuster, F.L.; Visvesvara, G.S. Demonstration of Balamuthia and Acanthamoeba mitochondrial DNA in sectioned archival brain and other tissues by the polymerase chain reaction. Parasitol. Res. 2008, 102, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Reddy, R.; Vijayasaradhi, M.; Uppin, M.S.; Challa, S.; Jabeen, A.; Borghain, R. Acanthamoeba meningoencephalitis in an immunocompetent patient: An autopsy case report. Neuropathology 2011, 31, 183–187. [Google Scholar] [CrossRef] [PubMed]
- Trabelsi, H.; Dendana, F.; Sellami, A.; Cheikhrouhou, F.; Neji, S.; Makni, F.; Ayadi, A. Pathogenic free-living amoebe: Epidemiology and clinical review. Pathol. Biol. 2012, 60, 399–405. [Google Scholar] [CrossRef]
- Martinez, A.J. Infection of the central nervous system due to Acanthamoeba. Rev. Infect. Dis. 1991, 13, 399–402. [Google Scholar] [CrossRef]
- Das, S.; Gunasekaran, K.; Ajjampur, S.S.R.; Abraham, D.; George, T.; Janeela, M.A.; Iyadurai, R. Acanthamoeba encephalitis in immunocompetent hosts: A report of two cases. J. Fam. Med. Prim. Care 2020, 9, 1240–1243. [Google Scholar] [CrossRef]
- Khanna, V.; Shastri, B.; Anusha, G.; Mukhopadhayay, C.; Khanna, R. Acanthamoeba meningoencephalitis in immunocompetent: A case report and review of literature. Trop. Parasitol. 2014, 4, 115–118. [Google Scholar] [CrossRef] [Green Version]
- Khurana, S.; Mewara, A.; Verma, S.; Totadri, S.K. Central nervous system infection with Acanthamoeba in a malnourished child. Case Rep. 2012, 2012, bcr2012007449. [Google Scholar] [CrossRef]
- Ranjan, R.; Handa, A.; Choudhary, A.; Kumar, S. Acanthamoeba infection in an interhemispheric ependymal cyst: A case report. Surg. Neurol. 2009, 72, 185–189. [Google Scholar] [CrossRef]
- Ghadage, D.P.; Choure, A.C.; Wankhade, A.B.; Bhore, A.V. Opportunistic free: Living amoeba now becoming a usual pathogen? Indian J. Pathol. Microbiol. 2017, 60, 601–603. [Google Scholar] [CrossRef]
- Hamide, A.; Sarkar, E.; Kumar, N.; Das, A.K.; Narayan, S.K.; Parija, S.C. Acanthameba meningoencephalitis: A case report. Neurol. India 2002, 50, 484–486. [Google Scholar] [PubMed]
- Saxena, A.; Mittal, S.; Burman, P.; Garg, P. Acanthameba meningitis with successful outcome. Indian J. Pediatr. 2009, 76, 1063–1064. [Google Scholar] [CrossRef] [PubMed]
- Hanafiah, M.; Shahizon, A.M.M.; Hamdan, M.F.; Wong, S.W.; Kanaheswari, Y. Acanthamoeba encephalitis in an immunocompeteny child and review of the imaging features of intracranical acanthamoebic infections in immunocompetent patients. Neurol. Asia 2018, 23, 179–184. [Google Scholar]
- Seijo Martinez, M.; Gonzalez-Mediero, G.; Santiago, P.; Rodriguez De Lope, A.; Diz, J.; Conde, C.; Visvesvara, G.S. Granulomatous amebic encephalitis in a patient with AIDS: Isolation of acanthamoeba sp. Group II from brain tissue and successful treatment with sulfadiazine and fluconazole. J. Clin. Microbiol. 2000, 38, 3892–3895. [Google Scholar] [CrossRef] [Green Version]
- Petry, F.; Torzewski, M.; Bohl, J.; Wilhelm-Schwenkmezger, T.; Scheid, P.; Walochnik, J.; Michel, R.; Zöller, L.; Werhahn, K.J.; Bhakdi, S.; et al. Early diagnosis of Acanthamoeba infection during routine cytological examination of cerebrospinal fluid. J. Clin. Microbiol. 2006, 44, 1903–1904. [Google Scholar] [CrossRef] [Green Version]
- Fung, K.T.; Dhillon, A.P.; McLaughlin, J.E.; Lucas, S.B.; Davidson, B.; Rolles, K.; Patch, D.; Burroughs, A.K. Cure of Acanthamoeba cerebral abscess in a liver transplant patient. Liver Transpl. 2008, 14, 308–312. [Google Scholar] [CrossRef]
- Maritschnegg, P.; Sovinz, P.; Lackner, H.; Benesch, M.; Nebl, A.; Schwinger, W.; Walochnik, J.; Urban, C. Granulomatous amebic encephalitis in a child with acute lymphoblastic leukemia successfully treated with multimodal antimicrobial therapy and hyperbaric oxygen. J. Clin. Microbiol. 2011, 49, 446–448. [Google Scholar] [CrossRef] [Green Version]
- Lackner, P.; Beer, R.; Broessner, G.; Helbok, R.; Pfausler, B.; Brenneis, C.; Auer, H.; Walochnik, J.; Schmutzhard, E. Acute granulomatous acanthamoeba encephalitis in an immunocompetent patient. Neurocrit. Care 2010, 12, 91–94. [Google Scholar] [CrossRef]
- Walochnik, J.; Aichelburg, A.; Assadian, O.; Steuer, A.; Visvesvara, G.; Vetter, N.; Aspöck, H. Granulomatous amoebic encephalitis caused by Acanthamoeba amoebae of genotype T2 in a human immunodeficiency virus-negative patient. J. Clin. Microbiol. 2008, 46, 338–340. [Google Scholar] [CrossRef] [Green Version]
- Modica, S.; Miracco, C.; Cusi, M.G.; Tordini, G.; Muzii, V.F.; Iacoangeli, F.; Nocentini, C.; Ali, I.K.M.; Roy, S.; Cerase, A.; et al. Non-granulomatous cerebellar infection by Acanthamoeba spp. in an immunocompetent host. Infection 2018, 46, 885–889. [Google Scholar] [CrossRef]
- El Sahly, H.; Udayamurthy, M.; Parkerson, G.; Hasbun, R. Survival of an AIDS patient after infection with Acanthamoeba sp. of the central nervous system. Infection 2017, 45, 715–718. [Google Scholar] [CrossRef] [PubMed]
- Webster, D.; Umar, I.; Kolyvas, G.; Bilbao, J.; Guiot, M.C.; Duplisea, K.; Qvarnstrom, Y.; Visvesvara, G.S. Treatment of granulomatous amoebic encephalitis with voriconazole and miltefosine in an immunocompetent soldier. Am. J. Trop. Med. Hyg. 2012, 87, 715–718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheng, W.H.; Hung, C.C.; Huang, H.H.; Liang, S.Y.; Cheng, Y.J.; Ji, D.D.; Chang, S.C. First case of granulomatous amebic encephalitis caused by Acanthamoeba castellanii in Taiwan. Am. J. Trop. Med. Hyg. 2009, 81, 277–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasprzak, W. Free-Living Amoeba with Pathogenic Properties to Human and Animals, 1st ed.; Państwowe Wydawnictwo Naukowe: Warsaw, Poland, 1985; pp. 8–15. [Google Scholar]
- Khan, N.A.; Siddiqui, R. The neuropathogenesis of Acanthamoeba encephalitis: Barriers to overcome. J. Cell Sci. Ther. 2011, S3. [Google Scholar] [CrossRef] [Green Version]
- Visvesvara, G.S.; Stehr-Green, J.K. Epidemiology of free-living ameba infections. J. Protozool. 1990, 37, 25S–33S. [Google Scholar] [CrossRef]
- Kawasaki, T.; Kawai, T. Toll-like receptor signaling pathways. Front. Immunol. 2014, 5, 461. [Google Scholar] [CrossRef] [Green Version]
- El-Zayat, S.R.; Sibaii, H.; Mannaa, F.A. Toll-like receptors activation, signaling, and targeting: An overview. Bull. Natl. Res. Cent. 2019, 43, 187. [Google Scholar] [CrossRef] [Green Version]
- Olson, J.K.; Miller, S.D. Microglia initiate central nervous system innate and adaptive immune responses through multiple TLRs. J. Immunol. 2004, 173, 3916–3924. [Google Scholar] [CrossRef] [Green Version]
- Tang, S.C.; Arumugam, T.V.; Xu, X.; Cheng, A.; Mughal, M.R.; Jo, D.G.; Lathia, J.D.; Siler, D.A.; Chigurupati, S.; Ouyang, X.; et al. Pivotal role for neuronal Toll-like receptors in ischemic brain injury and functional deficits. Proc. Natl. Acad. Sci. USA 2007, 104, 13798–13803. [Google Scholar] [CrossRef] [Green Version]
- Sauter, B.; Albert, M.L.; Francisco, L.; Larson, M.; Somersan, S.; Bhardwaj, N. Consequences of cell death: Exposure to necrotic tumor cells, but not primary tissue cells or apoptotic cells, induces the maturation of immunostimulatory dendritic cells. J. Exp. Med 2000, 191, 423–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vabulas, R.M.; Ahmad-Nejad, P.; Ghose, S.; Kirschning, C.J.; Issels, R.D.; Wagner, H. HSP70 as endogenous stimulus of the Toll/interleukin-1 receptor signal pathway. J. Biol. Chem. 2002, 277, 15107–15112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wojtkowiak-Giera, A.; Derda, M.; Kolasa-Wołosiuk, A.; Hadaś, E.; Kosik-Bogacka, D.; Solarczyk, P.; Jagodziński, P.P.; Wandurska-Nowak, E. Toll-like receptors in the brain of mice following infection with Acanthamoeba spp. Parasitol. Res. 2016, 115, 4335–4344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Elios, M.M.; Benagiano, M.; Della Bella, C.; Amedei, A. T-cell response to bacterial agents. J. Infect. Dev. Ctries. 2011, 5, 640–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shepherd, F.R.; McLaren, J.E. T Cell Immunity to Bacterial Pathogens: Mechanisms of Immune Control and Bacterial Evasion. Int. J. Mol. Sci. 2020, 21, 6144. [Google Scholar] [CrossRef] [PubMed]
- Robinson, D.S.; O’Garra, A. Further checkpoints in Th1 development. Immunity 2002, 16, 755–758. [Google Scholar] [CrossRef] [Green Version]
- Dabbagh, K.; Lewis, D.B. Toll-like receptors and T-helper-1/T-helper-2 responses. Curr. Opin. Infect. Dis. 2003, 16, 199–204. [Google Scholar] [CrossRef]
- Fouzder, C.; Mukhuty, A.; Das, S.; Chattopadhyay, D. TLR signaling on protozoan and helminthic parasite. In Toll-Like Receptors; InTechOpen: London, UK, 2019. [Google Scholar] [CrossRef]
- Massilamany, C.; Marciano-Cabral, F.; Rocha-Azevedo, B.d.; Jamerson, M.; Gangaplara, A.; Steffen, D.; Zabad, R.; Illes, Z.; Sobel, R.A.; Reddy, J. SJL mice infected with Acanthamoeba castellanii develop central nervous system autoimmunity through the generation of cross-reactive T cells for myelin antigens. PLoS ONE 2014, 9, e98506. [Google Scholar] [CrossRef] [Green Version]
- Parks, W.C.; Wilson, C.L.; López-Boado, Y.S. Matrix metalloproteinases as modulators of inflammation and innate immunity. Nat. Rev. Immunol. 2004, 4, 617–629. [Google Scholar] [CrossRef]
- Sternlicht, M.D.; Werb, Z. How matrix metalloproteinases regulate cell behavior. Annu. Rev. Cell Dev. Biol. 2001, 17, 463–516. [Google Scholar] [CrossRef] [Green Version]
- Raeeszadeh-Sarmazdeh, M.; Do, L.D.; Hritz, B.G. Metalloproteinases and Their Inhibitors: Potential for the Development of New Therapeutics. Cells 2020, 9, 1313. [Google Scholar] [CrossRef]
- Yong, V.W. Metalloproteinases: Mediators of pathology and regeneration in the CNS. Nat. Rev. Neurosci. 2005, 6, 931–944. [Google Scholar] [CrossRef] [PubMed]
- Yong, V.W.; Krekoski, C.A.; Forsyth, P.A.; Bell, R.; Edwards, D.R. Matrix metalloproteinases and diseases of the CNS. Trends Neurosci. 1998, 21, 75–80. [Google Scholar] [CrossRef]
- Agrawal, S.M.; Lau, L.; Yong, V.W. MMPs in the central nervous system: Where the good guys go bad. Semin. Cell Dev. Biol. 2008, 19, 42–51. [Google Scholar] [CrossRef]
- Bruschi, F.; Pinto, B. The significance of matrix metalloproteinases in parasitic infections involving the central nervous system. Pathogens 2013, 2, 105–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lam, C.; Jamerson, M.; Cabral, G.; Carlesso, A.M.; Marciano-Cabral, F. Expression of matrix metalloproteinases in Naegleria fowleri and their role in invasion of the central nervous system. Microbiology 2017, 163, 1436–1444. [Google Scholar] [CrossRef]
- Łanocha-Arendarczyk, N.; Baranowska-Bosiacka, I.; Gutowska, I.; Kolasa-Wołosiuk, A.; Kot, K.; Łanocha, A.; Metryka, E.; Wiszniewska, B.; Chlubek, D.; Kosik-Bogacka, D. The Activity of Matrix Metalloproteinases (MMP-2, MMP-9) and Their Tissue Inhibitors (TIMP-1, TIMP-3) in the Cerebral Cortex and Hippocampus in Experimental Acanthamoebiasis. Int. J. Mol. Sci. 2018, 19, 4128. [Google Scholar] [CrossRef] [Green Version]
- Bathina, S.; Das, U.N. Brain-derived neurotrophic factor and its clinical implications. Arch. Med. Sci. 2015, 11, 1164–1178. [Google Scholar] [CrossRef]
- Zigova, T.; Pencea, V.; Wiegand, S.J.; Luskin, M.B. Intraventricular administration of BDNF increases the number of newly generated neurons in the adult olfactory bulb. Mol. Cell Neurosci. 1998, 11, 234–245. [Google Scholar] [CrossRef]
- Benraiss, A.; Chmielnicki, E.; Lerner, K.; Roh, D.; Goldman, S.A. Adenoviral brain-derived neurotrophic factor induces both neostriatal and olfactory neuronal recruitment from endogenous progenitor cells in the adult forebrain. J. Neurosci. 2001, 21, 6718–6731. [Google Scholar] [CrossRef] [Green Version]
- Sohrabji, F.; Lewis, D.K. Estrogen-BDNF interactions: Implications for neurodegenerative diseases. Front. Neuroendocrinol. 2006, 27, 404–414. [Google Scholar] [CrossRef] [Green Version]
- Palasz, E.; Wysocka, A.; Gasiorowska, A.; Chalimoniuk, M.; Niewiadomski, W.; Niewiadomska, G. BDNF as a Promising Therapeutic Agent in Parkinson’s Disease. Int. J. Mol. Sci. 2020, 21, 1170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuccato, C.; Liber, D.; Ramos, C.; Tarditi, A.; Rigamonti, D.; Tartari, M.; Valenza, M.; Cattaneo, E. Progressive loss of BDNF in a mouse model of Huntington’s disease and rescue by BDNF delivery. Pharmacol. Res. 2005, 52, 133–139. [Google Scholar] [CrossRef] [PubMed]
- McDonald, C.R.; Conroy, A.L.; Hawkes, M.; Elphinstone, R.E.; Gamble, J.L.; Hayford, K.; Namasopo, S.; Opoka, R.O.; Liles, W.C.; Kain, K.C. Brain-derived Neurotrophic Factor Is Associated with Disease Severity and Clinical Outcome in Ugandan Children Admitted to Hospital With Severe Malaria. Pediatr. Infect. Dis. J. 2017, 36, 146–150. [Google Scholar] [CrossRef] [PubMed]
- Cordeiro, C.A.; Vieira, E.L.; Rocha, N.P.; Castro, V.M.; Oréfice, J.L.; Barichello, T.; Costa, R.A.; Oréfice, F.; Young, L.; Teixeira, A.L. Serum levels of neurotrophic factors in active toxoplasmic retinochoroiditis. Braz. J. Infect. Dis. 2017, 21, 176–179. [Google Scholar] [CrossRef] [PubMed]
- Łanocha-Arendarczyk, N.; Kosik-Bogacka, D.; Kot, K.; Baranowska-Bosiacka, I.; Metryka, E. Brain-derived neurotrophic factor (BDNF) in the brain cortex and hippocampus of mice infected with Acanthamoeba spp. In Proceedings of the Conference: Biotechnologia-Problemy i Wyzwania, Szczecin, Poland, 8 June 2018; pp. 86–87. Abstract book. (In Polish). [Google Scholar]
- Ibáñez, C.F. Neurotrophin-4: The odd one out in the neurotrophin family. Neurochem. Res. 1996, 21, 787–793. [Google Scholar] [CrossRef]
- Ferrer, I.; Krupinski, J.; Goutan, E.; Martí, E.; Ambrosio, S.; Arenas, E. Brain-derived neurotrophic factor reduces cortical cell death by ischemia after middle cerebral artery occlusion in the rat. Acta Neuropathol. 2001, 101, 229–238. [Google Scholar] [CrossRef]
- Klintsova, A.Y.; Greenough, W.T. Synaptic plasticity in cortical systems. Curr. Opin. Neurobiol. 1999, 9, 203–208. [Google Scholar] [CrossRef]
- Chan, K.M.; Lam, D.T.; Pong, K.; Widmer, H.R.; Hefti, F. Neurotrophin-4/5 treatment reduces infarct size in rats with middle cerebral artery occlusion. Neurochem. Res. 1996, 21, 763–767. [Google Scholar] [CrossRef]
- Chung, J.Y.; Kim, M.W.; Bang, M.S.; Kim, M. Increased expression of neurotrophin 4 following focal cerebral ischemia in adult rat brain with treadmill exercise. PLoS ONE 2013, 8, e52461. [Google Scholar] [CrossRef] [Green Version]
- Łanocha-Arendarczyk, N.; Kot, K.; Baranowska-Bosiacka, I.; Metryka, E.; Kapczuk, P.; Kosik-Bogacka, D. Activity of neurotrophin (NT-4) in the brain cortex and hippocampus in experimental acanthamoebiasis with regard to the immunological status of the host. Ann. Parasitol. 2019, 65, 117. [Google Scholar]
- Culbertson, C.G.; Smith, J.W.; Minner, J.R. Acanthamoeba: Observations on animal pathogenicity. Science 1958, 127, 1506. [Google Scholar] [CrossRef] [PubMed]
- Culbertson, C.G.; Smith, J.W.; Cohen, H.K.; Minner, J.R. Experimental infection of mice and monkeys by Acanthamoeba. Am. J. Pathol. 1959, 35, 185–197. [Google Scholar] [PubMed]
- Culberston, C.G.; Ensminger, P.W.; Overton, W.M. Hartmennella (Acanthamoeba) experimental chronic, granulomatous brain infections produced by new isolated of low virulence. Am. J. Clin. Pathol. 1966, 46, 305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rucka, A. Histopathological changes in the brain and lungs of mice infected with pathogenic amoebas. Wiad. Parazytol. 1974, 2–3, 247–250. (In Polish) [Google Scholar]
- Gieryng, H.; Gieryng, R. Histopathological changes in the brain of mice infected with the amoebas from the limax group. Ann. Univ. Mariae Curie Sklodowska 1987, 16, 103–109. (In Polish) [Google Scholar]
- Martinez, A.J. Is Acanthamoeba encephalitis an opportunistic infection? Neurology 1980, 30, 567–574. [Google Scholar] [CrossRef]
- Visvesvara, G.S.; Moura, H.; Schuster, F.L. Pathogenic and opportunistic free-living amoebae: Acanthamoeba spp., Balamuthia mandrillaris, Naegleria fowleri, and Sappinia diploidea. FEMS Immunol. Med. Microbiol. 2007, 50, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Schuster, F.L.; Yagi, S.; Gavali, S.; Michelson, D.; Raghavan, R.; Blomquist, I.; Glastonbury, C.; Bollen, A.W.; Scharnhorst, D.; Reed, S.L.; et al. Under the radar: Balamuthia amebic encephalitis. Clin. Infect. Dis. 2009, 48, 879–887. [Google Scholar] [CrossRef]
- Kaul, D.R.; Lowe, L.; Visvesvara, G.S.; Farmen, S.; Khaled, Y.A.; Yanik, G.A. Acanthamoeba infection in a patient with chronic graft-versus-host disease occurring during treatment with voriconazole. Transpl. Infect. Dis. 2008, 10, 437–441. [Google Scholar] [CrossRef] [Green Version]
- Young, A.L.; Leboeuf, N.R.; Tsiouris, S.J.; Husain, S.; Grossman, M.E. Fatal disseminated Acanthamoeba infection in a liver transplant recipient immunocompromised by combination therapies for graft-versus-host disease. Transpl. Infect. Dis. 2010, 12, 529–537. [Google Scholar] [CrossRef]
- Visvesvara, G.S.; Mirra, S.S.; Brandt, F.H.; Moss, D.M.; Mathews, H.M.; Martinez, A.J. Isolation of two strains of Acanthamoeba castellanii from human tissue and their pathogenicity and isoenzyme profiles. J. Clin. Microbiol. 1983, 18, 1405–1412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez, A.J. Acanthamoebiasis and immunosuppression. Case report. J. Neuropathol. Exp. Neurol. 1982, 41, 548–557. [Google Scholar] [CrossRef] [PubMed]
- Anderlini, P.; Przepiorka, D.; Luna, M.; Langford, L.; Andreeff, M.; Claxton, D.; Deisseroth, A.B. Acanthamoeba meningoencephalitis after bone marrow transplantation. Bone Marrow Transplant. 1994, 14, 459–461. [Google Scholar] [PubMed]
- Castellano-Sanchez, A.; Popp, A.C.; Nolte, F.S.; Visvesvara, G.S.; Thigpen, M.; Redei, I.; Somani, J. Acanthamoeba castellani encephalitis following partially mismatched related donor peripheral stem cell transplantation. Transpl. Infect. Dis. 2003, 5, 191–194. [Google Scholar] [CrossRef] [PubMed]
- Murakawa, G.J.; McCalmont, T.; Altman, J.; Telang, G.H.; Hoffman, M.D.; Kantor, G.R.; Berger, T.G. Disseminated acanthamebiasis in patients with AIDS. A report of five cases and a review of the literature. Arch. Dermatol. 1995, 131, 1291–1296. [Google Scholar] [CrossRef] [PubMed]
- Duarte, A.G.; Sattar, F.; Granwehr, B.; Aronson, J.F.; Wang, Z.; Lick, S. Disseminated acanthamoebiasis after lung transplantation. J. Heart Lung Transplant. 2006, 25, 237–240. [Google Scholar] [CrossRef]
- Afshar, K.; Boydking, A.; Ganesh, S.; Herrington, C.; McFadden, P.M. Rapidly fatal disseminated acanthamoebiasis in a single lung transplant recipient. Ann. Transplant. 2013, 18, 108–111. [Google Scholar] [CrossRef]
- Readinger, A.; Blumberg, M.; Colome-Grimmer, M.I.; Kelly, E. Disseminated Acanthamoeba infection with sporotrichoid nodules. Int. J. Dermatol. 2006, 45, 942–943. [Google Scholar] [CrossRef]
- Im, K.; Kim, D.S. Acanthamoebiasis in Korea: Two new cases with clinical cases review. Yonsei Med. J. 1998, 39, 478–484. [Google Scholar] [CrossRef]
- Mutreja, D.; Jalpota, Y.; Madan, R.; Tewari, V. Disseminated acanthamoeba infection in a renal transplant recipient: A case report. Indian J. Pathol. Microbiol. 2007, 50, 346–348. [Google Scholar]
- Gordeeva, L.M. Amoeba of limax group from the respiratory tract of man interactions with cell cultures. In Proceedings of the 4th International Congress of Parasitology, Warszawa, Poland, 19–26 August 1978; pp. 34–35. [Google Scholar]
- Gu, J.; Liu, Y.; Xie, B.; Ye, P.; Huang, J.; Lu, Z. Roles of toll-like receptors: From inflammation to lung cancer progression. Biomed. Rep. 2018, 8, 126–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arora, S.; Ahmad, S.; Irshad, R.; Goyal, Y.; Rafat, S.; Siddiqui, N.; Dev, K.; Husain, M.; Ali, S.; Mohan, A.; et al. TLRs in pulmonary diseases. Life Sci. 2019, 233, 116671. [Google Scholar] [CrossRef] [PubMed]
- Derda, M.; Wojtkowiak-Giera, A.; Kolasa-Wołosiuk, A.; Kosik-Bogacka, D.; Hadaś, E.; Jagodziński, P.P.; Wandurska-Nowak, E. Acanthamoeba infection in lungs of mice expressed by toll-like receptors (TLR2 and TLR4). Exp. Parasitol. 2016, 165, 30–34. [Google Scholar] [CrossRef] [PubMed]
- Kay, E.; Scotland, R.S.; Whiteford, J.R. Toll-like receptors: Role in inflammation and therapeutic potential. Biofactors 2014, 40, 284–294. [Google Scholar] [CrossRef] [PubMed]
- Phythian-Adams, A.T.; Cook, P.C.; Lundie, R.J.; Jones, L.H.; Smith, K.A.; Barr, T.A.; Hochweller, K.; Anderton, S.M.; Hämmerling, G.J.; Maizels, R.M.; et al. CD11c depletion severely disrupts Th2 induction and development in vivo. J. Exp. Med. 2010, 207, 2089–2096. [Google Scholar] [CrossRef] [PubMed]
- Paul, W.E.; Zhu, J. How are T(H)2-type immune responses initiated and amplified? Nat. Rev. Immunol. 2010, 10, 225–235. [Google Scholar] [CrossRef]
- Maizels, R.M.; Balic, A.; Gomez-Escobar, N.; Nair, M.; Taylor, M.D.; Allen, J.E. Helminth parasites: Masters of regulation. Immunol. Rev. 2004, 201, 89–116. [Google Scholar] [CrossRef]
- Park, M.K.; Cho, M.K.; Kang, S.A.; Park, H.K.; Kim, D.H.; Yu, H.S. Acanthamoeba protease activity promotes allergic airway inflammation via protease-activated receptor 2. PLoS ONE 2014, 9, e92726. [Google Scholar] [CrossRef] [Green Version]
- Lee, I.T.; Yang, C.M. Inflammatory signalings involved in airway and pulmonary diseases. Mediat. Inflamm. 2013, 2013, 791231. [Google Scholar] [CrossRef] [Green Version]
- Korbecki, J.; Baranowska-Bosiacka, I.; Gutowska, I.; Chlubek, D. Cyclooxygenase pathways. Acta Biochim. Pol. 2014, 61, 639–649. [Google Scholar] [CrossRef] [Green Version]
- Dannhardt, G.; Kiefer, W. Cyclooxygenase inhibitors—Current status and future prospects. Eur. J. Med. Chem. 2001, 36, 109–126. [Google Scholar] [CrossRef]
- Claar, D.; Hartert, T.V.; Peebles, R.S., Jr. The role of prostaglandins in allergic lung inflammation and asthma. Expert Rev. Respir. Med. 2015, 9, 55–72. [Google Scholar] [CrossRef] [Green Version]
- Hasturk, S.; Kemp, B.; Kalapurakal, S.K.; Kurie, J.M.; Hong, W.K.; Lee, J.S. Expression of cyclooxygenase-1 and cyclooxygenase-2 in bronchial epithelium and nonsmall cell lung carcinoma. Cancer 2002, 94, 1023–1031. [Google Scholar] [CrossRef]
- Radi, Z.A. Pathophysiology of cyclooxygenase inhibition in animal models. Toxicol. Pathol. 2009, 37, 34–46. [Google Scholar] [CrossRef] [Green Version]
- Szymanski, K.V.; Toennies, M.; Becher, A.; Fatykhova, D.; N’Guessan, P.D.; Gutbier, B.; Klauschen, F.; Neuschaefer-Rube, F.; Schneider, P.; Rueckert, J.; et al. Streptococcus pneumoniae-induced regulation of cyclooxygenase-2 in human lung tissue. Eur. Respir. J. 2012, 40, 1458–1467. [Google Scholar] [CrossRef] [Green Version]
- Kosik-Bogacka, D.I.; Baranowska-Bosiacka, I.; Kolasa-Wołosiuk, A.; Lanocha-Arendarczyk, N.; Gutowska, I.; Korbecki, J.; Namięta, H.; Rotter, I. The inflammatory effect of infection with Hymenolepis diminuta via the increased expression and activity of COX-1 and COX-2 in the rat jejunum and colon. Exp. Parasitol. 2016, 169, 69–76. [Google Scholar] [CrossRef]
- Łanocha-Arendarczyk, N.; Baranowska-Bosiacka, I.; Kot, K.; Gutowska, I.; Kolasa-Wołosiuk, A.; Chlubek, D.; Kosik-Bogacka, D. Expression and Activity of COX-1 and COX-2 in Acanthamoeba sp.-Infected Lungs According to the Host Immunological Status. Int. J. Mol. Sci. 2018, 19, 121. [Google Scholar] [CrossRef] [Green Version]
- Birben, E.; Sahiner, U.M.; Sackesen, C.; Erzurum, S.; Kalayci, O. Oxidative stress and antioxidant defense. World Allergy Organ. J. 2012, 5, 9–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hosakote, Y.M.; Liu, T.; Castro, S.M.; Garofalo, R.P.; Casola, A. Respiratory syncytial virus induces oxidative stress by modulating antioxidant enzymes. Am. J. Respir. Cell Mol. Biol. 2009, 41, 348–357. [Google Scholar] [CrossRef] [PubMed]
- Kolodziejczyk, L.; Siemieniuk, E.; Skrzydlewska, E. Antioxidant potential of rat liver in experimental infection with Fasciola hepatica. Parasitol. Res. 2005, 96, 367–372. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Na, B.K.; Song, K.J.; Park, M.H.; Park, Y.K.; Kim, T.S. Functional expression and characterization of an iron-containing superoxide dismutase of Acanthamoeba castellanii. Parasitol. Res. 2012, 111, 1673–1682. [Google Scholar] [CrossRef] [PubMed]
- Motavalli, M.; Khodadadi, I.; Fallah, M.; Maghsood, A.H. Effect of oxidative stress on vital indicators of Acanthamoeba castellanii (T4 genotype). Parasitol. Res. 2018, 117, 2957–2962. [Google Scholar] [CrossRef] [PubMed]
- Hadas, E.; Mazur, T. Proteolytic enzymes of pathogenic and non-pathogenic strains of Acanthamoeba spp. Trop. Med. Parasitol. 1993, 44, 197–200. [Google Scholar]
- Jha, B.K.; Jung, H.J.; Seo, I.; Suh, S.I.; Suh, M.H.; Baek, W.K. Juglone induces cell death of Acanthamoeba through increased production of reactive oxygen species. Exp. Parasitol. 2015, 159, 100–106. [Google Scholar] [CrossRef]
- Łanocha-Arendarczyk, N.; Baranowska-Bosiacka, I.; Gutowska, I.; Kot, K.; Metryka, E.; Kosik-Bogacka, D.I. Relationship between antioxidant defense in Acanthamoeba spp. infected lungs and host immunological status. Exp. Parasitol. 2018, 193, 58–65. [Google Scholar] [CrossRef]
- Gieryng, H.; Gieryng, R.; Piróg, Z. Histological changes in the lungs of mice experimentally infected with amoebas from the limax group. Histopathological changes in the lungs. Wiad. Parazytol. 1993, 39, 367–372. (In Polish) [Google Scholar]
- Elsheikha, H.M.; Siddiqui, R.; Khan, N.A. Drug Discovery against Acanthamoeba Infections: Present Knowledge and Unmet Needs. Pathogens 2020, 9, 405. [Google Scholar] [CrossRef]
- Tunkel, A.R.; Glaser, C.A.; Bloch, K.C.; Sejvar, J.J.; Marra, C.M.; Roos, K.L.; Hartman, B.J.; Kaplan, S.L.; Scheld, W.M.; Whitley, R.J. Infectious Diseases Society of America. The management of encephalitis: Clinical practice guidelines by the Infectious Diseases Society of America. Clin. Infect. Dis. 2008, 47, 303–327. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC). Investigational drug available directly from CDC for the treatment of infections with free-living amebae. MMWR Morb. Mortal. Wkly. Rep. 2013, 62, 666. [Google Scholar]
Figure 1.
Host defense mechanisms against Acanthamoeba spp. infection in the brain: 1. Acanthamoeba spp. infection affects the growth of Toll-like receptors (TLR) expression, mainly TLR2 and TLR4. Moreover, it was found that antigen-sensitized lymphocytes from the brain of Acanthamoeba spp. infected mice contain mostly Th1 cytokine-producing cells; 2. Acanthamoebiasis causes an increase in the activity of metalloproteinases-9 (MMP-9) and the tissue inhibitor of metalloproteinases-1 (TIMP-1) in immunocompetent hosts (IC), and a decrease in MMP-9 and TIMP-3 in immunosuppressed hosts (IS). In immunocompromised hosts, an increase in the ratio of MMP-9/ TIMP-1 was also observed; 3. Acanthamoeba spp. infection affects the growth of brain derived neurotrophic factor (BDNF) and neurotrophin-4 (NT-4) activity in both immunocompetent (IC) and immunosuppressed (IS) hosts; 4. The amoebas induced inflammatory and necrotic lesions in the brain structures.
Figure 1.
Host defense mechanisms against Acanthamoeba spp. infection in the brain: 1. Acanthamoeba spp. infection affects the growth of Toll-like receptors (TLR) expression, mainly TLR2 and TLR4. Moreover, it was found that antigen-sensitized lymphocytes from the brain of Acanthamoeba spp. infected mice contain mostly Th1 cytokine-producing cells; 2. Acanthamoebiasis causes an increase in the activity of metalloproteinases-9 (MMP-9) and the tissue inhibitor of metalloproteinases-1 (TIMP-1) in immunocompetent hosts (IC), and a decrease in MMP-9 and TIMP-3 in immunosuppressed hosts (IS). In immunocompromised hosts, an increase in the ratio of MMP-9/ TIMP-1 was also observed; 3. Acanthamoeba spp. infection affects the growth of brain derived neurotrophic factor (BDNF) and neurotrophin-4 (NT-4) activity in both immunocompetent (IC) and immunosuppressed (IS) hosts; 4. The amoebas induced inflammatory and necrotic lesions in the brain structures.

Figure 2.
Host defense mechanisms against Acanthamoeba spp. infection in the lungs: 1. Acanthamoeba spp. infection affects the growth of Toll-like receptor (TLR) expression, mainly TLR2 and TLR4. Moreover, it was found that Acanthamoeba spp. increased the synthesis of Th2 cytokines, such as interleukin (IL)-4, IL-5, and IL-13; 2. Acanthamoebiasis causes an increase in expression of cyclooxygenase-1 (COX-1) and -2 (COX-2) in immunocompetent (IC) and immunosuppressed (IS) hosts; 3. Acanthamoeba spp. infection causes oxidative stress manifested by an increase in lipid peroxidation (LPO) and changes in antioxidant enzyme activity in the immunocompetent hosts; an increase in catalase (CAT) and a decrease in glutathione peroxidase (GPx). In immunosuppressed hosts, Acanthamoeba spp. causes a decrease in LPO and a decrease in superoxide dismutase (SOD), glutathione reductase (GR) and GPx; 4. The amoebas induced hemorrhage, inflammatory and necrotic lesions in the lungs.
Figure 2.
Host defense mechanisms against Acanthamoeba spp. infection in the lungs: 1. Acanthamoeba spp. infection affects the growth of Toll-like receptor (TLR) expression, mainly TLR2 and TLR4. Moreover, it was found that Acanthamoeba spp. increased the synthesis of Th2 cytokines, such as interleukin (IL)-4, IL-5, and IL-13; 2. Acanthamoebiasis causes an increase in expression of cyclooxygenase-1 (COX-1) and -2 (COX-2) in immunocompetent (IC) and immunosuppressed (IS) hosts; 3. Acanthamoeba spp. infection causes oxidative stress manifested by an increase in lipid peroxidation (LPO) and changes in antioxidant enzyme activity in the immunocompetent hosts; an increase in catalase (CAT) and a decrease in glutathione peroxidase (GPx). In immunosuppressed hosts, Acanthamoeba spp. causes a decrease in LPO and a decrease in superoxide dismutase (SOD), glutathione reductase (GR) and GPx; 4. The amoebas induced hemorrhage, inflammatory and necrotic lesions in the lungs.


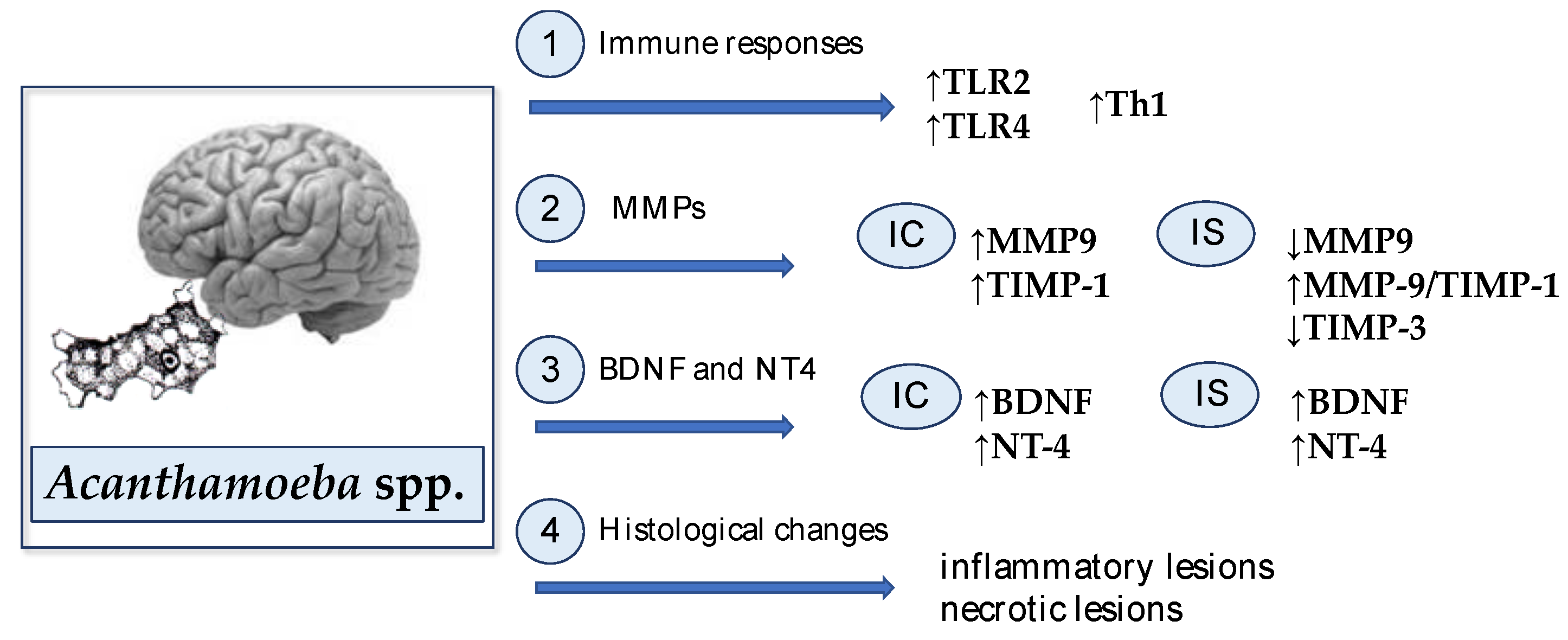{kind=link}
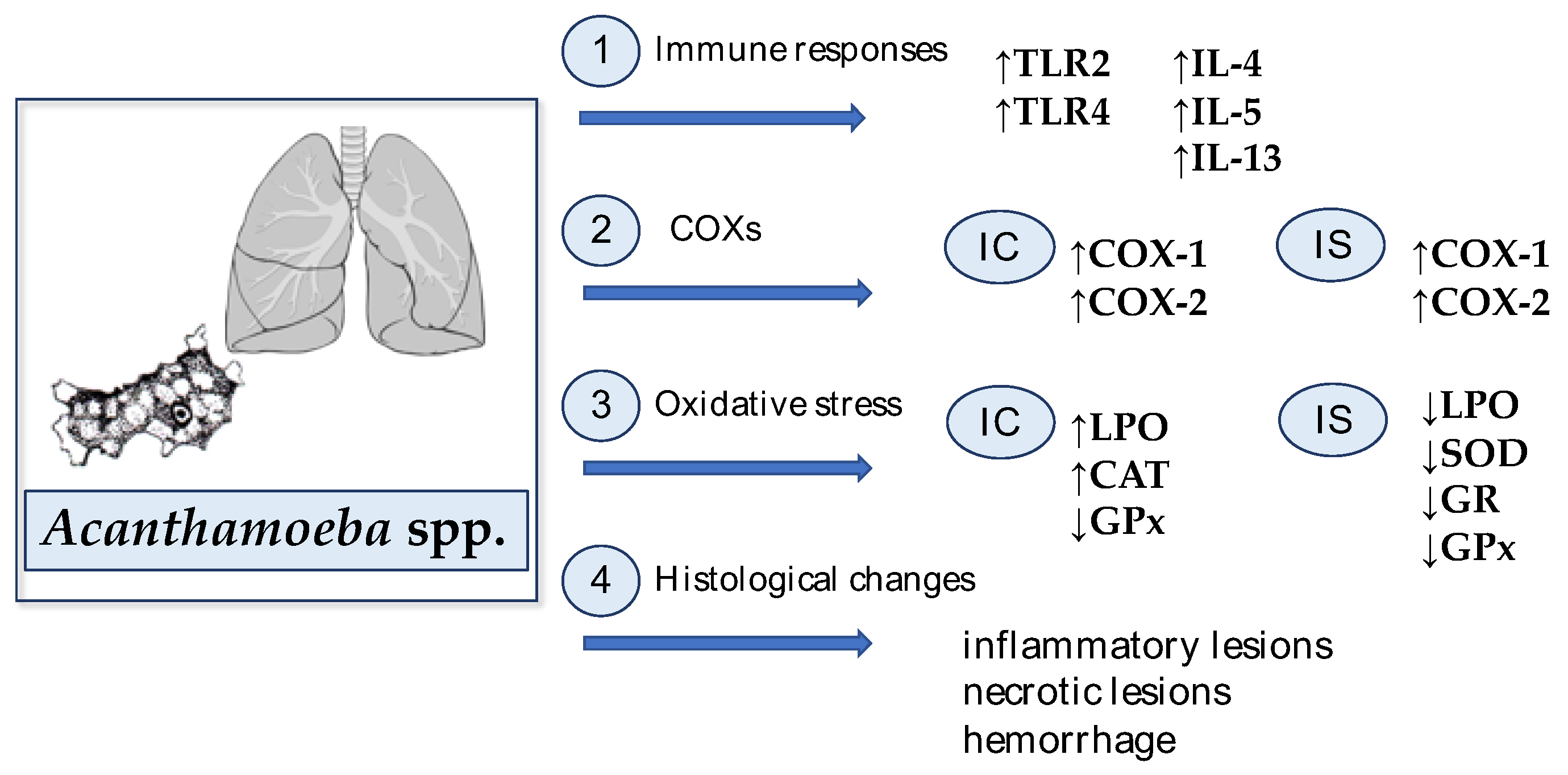{kind=link}
Table 1.
Cases of survivors of granulomatous amebic encephalitis (GAE) caused by Acanthamoeba spp. reported during 1990–2020.
Table 1.
Cases of survivors of granulomatous amebic encephalitis (GAE) caused by Acanthamoeba spp. reported during 1990–2020.
| No. | Place of Study | Patient Details | Biological Sample | Treatment | References |
|---|---|---|---|---|---|
| 1 | India | 22 years; IC | CSF | fluconazole, trimethoprim-sulfamethoxazole, metronidazole, rifampicin, miltefosin; for 3 months | [36] |
| 2 | India | 3 years, male; child on chronic malnutrition | CSF | - | [13] |
| 3 | India | 30 years, male; IC | CSF | rifampicin, trimethoprim-sulfamethoxazole, fluconazole; for 14 days | [37] |
| 4 | India | 3 years, male; malnourished child | CSF | cotrimoxazole, rifampicin, ketoconazole | [38] |
| 5 | India | 25 years, male; IC | frontotemporal craniotomy | rifampicin, trimethoprim-sulfamethoxazole, fluconazole | [39] |
| 6 | India | 2 years, male; IC | CSF | vancomycin, ceftriaxone, dexamethasone | [40] |
| 7 | India | 45 years, female; IC | CSF | rifampicin, cotrimoxazole, fluconazole, albendazole, carbamazepine | [41] |
| 8 | India | 15 years, female; IC | CSF | ketoconazole, rifampicin, cotrimoxazole; for 9 months | [42] |
| 9 | Malaysia | 1 year, female; IC | brain biopsy | fluconazole, trimethoprim, rifampicin; for 106 days | [43] |
| 10 | Spain | 33 years, male; patient with AIDS | brain biopsy | fluconazole, sulfadiazine and surgical excision of lesions | [44] |
| 11 | Germany | 64 years, female; IC | CSF | fluconazole, rifampicin, metronidazole, sulfadiazine; for 14 days | [45] |
| 12 | UK | 41 years, male; liver transplant recipient | brain tissue | rifampicin, co-trimoxazole; for 3 months | [46] |
| 13 | Austria | 25 months, male; patients with ALL | CSF | trimethoprim-sulfamethoxazole, fluconazole, pentamidine, miltefosine | [47] |
| 14 | Austria | 17 years, male; IC | CSF | fluconazole, rifampicin, cotrimoxazole; for 2 months | [48] |
| 15 | Austria | 25 years, male; patients with tuberculous meningitis | CSF, biopsy | fluconazole, trimethoprim-sulfamethoxazole, amphotericin B, flucytosine, sulfadiazine, miltefosine | [49] |
| 16 | Italy | 35 years, male; IC | brain tissue biopsy | fluconazole, trimethoprim-sulfamethoxazole, miltefosine | [50] |
| 17 | USA | 34 years, male; patient with HIV | CSF | trimethoprim-sulfamethoxazole, flucytosine, fluconazole, miltefosine | [51] |
| 18 | Canada | 38 years, male; IC | serum sample, brain biopsy | voriconazole, miltefosine | [52] |
| 19 | Taiwan | 63 years, male; IC | CSF | amphotericin B, rifampicin, dexamethasone | [53] |
ALL, acute lymphoblastic leukemia; CSF, cerebrospinal fluid; HIV, human immunodeficiency virus; IC, immunocompetent.
Table 2.
Cases of Acanthamoeba pneumonia (AP) or disseminated acanthamoebiasis with lung infection reported during 1990–2020.
Table 2.
Cases of Acanthamoeba pneumonia (AP) or disseminated acanthamoebiasis with lung infection reported during 1990–2020.
| No. | Place of Study | Patient Details | Biological Sample | References |
|---|---|---|---|---|
| 1 | Poland | 15 years, male; patient with AML | bronchoaspirate fluid | [20] |
| 2 | Poland | 53 years, male; patient with ACL | bronchoaspirate fluid | [20] |
| 3 | Poland | newborn with atypical pneumonia | bronchoalveolar lavage (BAL) | [20] |
| 4 | Poland | newborn with atypical pneumonia | bronchoalveolar lavage (BAL) | [20] |
| 5 | Austria | 25 years, male from India; IS | lung biopsy | [21] |
| 6 | France | 39 years, male; heart transplant recipient | postmortem, biopsy of lungs | [22] |
| 7 | USA | 53 years, male; patient with ALL | postmortem, biopsy of lungs | [102] |
| 8 | USA | 63 years, male; liver transplant recipient | postmortem, biopsy of lungs | [103] |
| 9 | USA | 38 years, male; kidney transplant recipient | postmortem, biopsy of lungs | [104] |
| 10 | USA | 28 years, male; kidney transplant recipient | postmortem, biopsy of lungs | [105] |
| 11 | USA | 39 years, female; patient with CML | postmortem, biopsy of lungs | [106] |
| 12 | USA | 45 years, male; patient with AML | postmortem, biopsy of lungs | [107] |
| 13 | USA | 35 years, male; patient with AIDS | bronchial washing | [108] |
| 14 | USA | 60 years, male; bilateral lung transplant recipient | postmortem, biopsy of lungs | [109] |
| 15 | USA | 62 years, male; single lung transplant recipient | postmortem, biopsy of lungs | [110] |
| 16 | USA | 60 years, male; lung transplant recipient | postmortem, biopsy of lungs | [111] |
| 17 | Korea | 7 months, female; congenital immunodeficiency patient | bronchial washing | [112] |
| 18 | Japan | 52 years, male; allogenic bone marrow transplantation | postmortem, biopsy of lungs | [23] |
| 19 | India | 36 years, female; kidney transplant recipient | postmortem, biopsy of lungs | [113] |
ACL, acute chronic marrow leukemia; AML, acute myeloblastic leukemia; ALL, acute lymphoblastic leukemia; CML, chronic myelogenous leukemia; IC, immunocompetent; IS, immunosuppressed.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kot, K.; Łanocha-Arendarczyk, N.; Kosik-Bogacka, D. Immunopathogenicity of Acanthamoeba spp. in the Brain and Lungs. Int. J. Mol. Sci. 2021, 22, 1261. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031261
AMA Style
Kot K, Łanocha-Arendarczyk N, Kosik-Bogacka D. Immunopathogenicity of Acanthamoeba spp. in the Brain and Lungs. International Journal of Molecular Sciences. 2021; 22(3):1261. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031261
Chicago/Turabian StyleKot, Karolina, Natalia Łanocha-Arendarczyk, and Danuta Kosik-Bogacka. 2021. "Immunopathogenicity of Acanthamoeba spp. in the Brain and Lungs" International Journal of Molecular Sciences 22, no. 3: 1261. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031261
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.