1. Introduction
More than two billion obese people are registered worldwide [
1]. According to the latest data, by 2040, the number of patients with diabetes is projected to increase to 642 million people—10% of the world’s population [
1]. Almost 95% of them are patients with (T2DM) [
2]. Complications from carbohydrate metabolism are caused by the chronic subclinical inflammation, an imbalance in redox processes, overexpression of reactive oxygen species (ROS), and oxidative stress.
Interleukin (IL)-6 is one of the main cytokines of chronic subclinical inflammation and is a cytokine with multidirectional effects. IL-6 promotes the insulin resistance (IR) and impairs glucose homeostasis [
3,
4]. Strong experimental data support the ability of IL-6 to potentiate IR in hepatocytes, while the results for adipocytes and skeletal muscle are not always consistent [
5,
6].
The IL-6 family cytokines and IL-11 belong to the same family, because each cytokine’s receptor complex contains two subunits of the signaling receptor gp130 [
6].
The IL-6 family regulates the processes of liver damage/regeneration and maintains the balance between regulatory and effector T cells [
6]. Chronic inflammatory processes lead to fatty infiltration of the liver and the spread of foci of inflammation (cellular infiltration) in the parenchyma. In this regard, the role of IL-6 in the liver is of interest in obesity.
A decrease in local and systemic inflammation can be due to the regulation of mitochondrial dynamics and associated processes, such as the activation of cell differentiation and regulation of glucose oxidation processes. Under the influence of oxidative stress, mitochondria adapt by regulating the processes of division/fusion and optimizing mtDNA matrix processes and their biogenesis, thereby exerting a wide influence on metabolic status [
7]. Mitochondrial biogenesis is achieved through the organized induction of several transcriptional regulators, the activity of which is critically determined by the prevailing energy requirements [
8]. Peroxisome proliferator-activated gamma coactivator (PGC)-1α, (TFAM) [
9], and nuclear respiratory factor 1 (NRF-1) are important regulators of mitochondrial biogenesis, and their activity is regulated by post-translational modifications that fine-tune mitochondrial metabolism.
Mitochondrial mechanisms of control of cellular bioenergetic homeostasis require a close connection with the nucleus (interaction between the mitochondrial and nuclear genome) and specifically include mechanisms related to antioxidant defense, protein homeostasis (import, folding, and degradation of proteins through the mitochondrial unfolded protein response), and autophagy for mitochondrial metabolism through the process of mitophagy [
7].
Mitophagy is a mitochondria-specific analogue of the autophagy process. The destruction of damaged and dysfunctional mitochondria occurs through autophagy. Mitophagy is an important process for maintaining mitochondrial quality and insulin sensitivity [
10]. The fusion or elongation of mitochondria allows the exchange of their contents, mtDNA, and metabolic intermediates and the restoration of the activity of damaged or depolarized membranes [
10].
Fusion is predominantly controlled by mitofusins 1 and 2 (Mfn1 and Mfn2) located on the outer mitochondrial membrane, and optic atrophy protein 1 (Opa1) located on the inner mitochondrial membrane [
11]. Division or fragmentation increases the number of mitochondria and prepares the cell for division. Division is regulated by fission protein 1 (FIS1), which is transported to the outer mitochondrial membrane dynamin-related protein (Drp) 1 during the process of division initiation [
11]. The dynamic transition of mitochondria between fused and fragmented states allows them to reorganize their structure and eliminate damaged or dysfunctional components using mitophagy mechanisms. Cells exposed to a nutrient-rich environment tend to keep their mitochondria in a fragmented state (fission). However, under conditions of starvation and nutrient depletion, mitochondria tend to remain bound or elongated for longer periods of time (fusion).
The fusion and fission processes continuously balance each other: inactivation of one activates the other, and vice versa [
7]. In general, these dynamic events regulate mitochondrial function. They carry out mitochondrial distribution, shape, and quality control.
Collectively, liver mitochondria temporarily adapt to increased lipid availability by increasing their oxidative capacity [
11]. Loss of mitochondrial adaptation contributes to lipid deposition, ROS accumulation, and, as a consequence, IR. Finally, excessive lipid overload disrupts the cell’s antioxidant capacity and energy balance. This leads to damage to various cell systems, in particular, mitochondrial biogenesis. As a result, IR is aggravated, and steatohepatitis develops.
In this regard, the study aimed to investigate the role of the IL-6 cytokine family (sIL-6Ra, gp130/sIL-6Rb, and IL-11) in the regulation of mitochondrial dynamics in the liver of obese patients and to assess the contribution of these cytokines to the pathogenesis of T2DM.
2. Materials and Methods
This study included 134 obese patients. Of these, 70 obese patients had T2DM (48.9 ± 7.28 years; 50.62 ± 8.83 kg/m2; 31 men and 39 women), and 115 obese patients did not have carbohydrate metabolism disorders (43.7 ± 9.4 years; 43.8 ± 7.2 kg/m2; 52 men and 63 women). The presence of arterial hypertension was noted in 41% of patients. The control group included 41 apparently healthy donors with normal anthropometric and biochemical parameters (39.1 ± 10.1 years; 22.6 ± 3.1 kg/m2; 23 men and 18 women). Liver samples were taken during routine laparoscopic operations from conditionally healthy donors: inguinal hernia on the right or left, femoral, diaphragmatic and ventral hernias, and nephroptosis.
All the study participants provided informed consent to participate in a research study. The study was carried out according to the World Medical Association (WMA) Declaration of Helsinki (2000) and the Protocol to the Convention on Human Rights and Biomedicine (1999). The Local Ethics Committee approved the study protocol of the Innovation Park of the Immanuel Kant Baltic Federal University (Protocol No. 4 from 23 October 2013).
The analysis of biochemical parameters in the blood serum was carried out on a Furuno CA-180 biochemical analyzer (Furuno Electric Company, Hyogo, Japan) using DiaSys test systems (DiaSys Diagnostic Systems, Holzheim, Germany). Plasma hormone levels were assessed by flow fluorimetry (Bio-Plex Protein Assay System, Bio-Rad, Heracles, CA, USA) using Bio-Plex Pro™ Human Inflammation Panel 1 test systems, 37-Plex (Bio-Rad, Heracles, CA, USA).
Total RNA from liver biopsies was isolated with ExtractRNA reagent (Evrogen, Moscow, Russia). Reverse transcription of total RNA samples was conducted with the MMLV RT kit (Evrogen, Moscow, Russia). Real-time PCR was conducted on a CFX96 Touch Real-Time PCR Detection System (Bio-Rad, Heracles, CA, USA). PCR data processing was performed using REST 2009 (QIAGEN, Valencia, CA, USA).
Semi-quantitative determination of proteins by immunoblotting was conducted using specific monoclonal antibodies (Thermo Fisher, Waltham, MA, USA) and the Mini-PROTEAN® Tetra Cell Systems and Mini Trans-Blot®Turbo blotting systems (Bio-Rad, Heracles, CA, USA). Total protein was isolated from liver biopsies by RIPA buffer (Thermo Fisher, Waltham, MA, USA). The Bradford method was used to measure the total protein concentration (Pierce BCA Protein Assay Kit, Thermo Fisher, Waltham, MA, USA). The specific proteins were detected on a ChemiDoc MP Imaging System (Bio-Rad, Heracles, CA, USA). Densitometry was performed using ImageLab software (Bio-Rad, Heracles, CA, USA).
Paraffin slices of liver biopsies were stained with haematoxylin–eosin. Traditional histological examination was conducted using a Leica DM3000 microscope (Leica Microsystems, Weitzlar, Germany). Additionally, the degree of steatosis and lymphocyte infiltration were assessed using a Pannoramic 250 FLASH (3DHISTECH, Hungary, Budapest) scanning microscope and ImageJ software.
Verification of the normality of quantitative indicator distribution was carried out using the Shapiro-Wilk test. Because the investigated samples fitted a normal distribution, the hypothesis of the equality of the mean sample values was verified using Student’s t-tests. To assess the significance of differences between independent quantitative samples that did not follow a normal distribution law, the non-parametric Kruskal-Wallis test was used. To detect statistically significant differences between groups, a pairwise analysis was performed using the non-parametric Mann-Whitney U test for independent groups.
Correlations between the studied indices were determined using Pearson correlation analysis. Differences were considered significant at the level of p < 0.05. All statistical analyses were performed in R language (version 3.3.1).
4. Discussion
The liver is exposed to higher levels of nutrients from food than peripheral tissues [
12]. Nutrients penetrate the liver the fastest through the hepatic portal vein [
13]. Hepatocytes can store and release glucose to minimize pre/post-meal imbalances [
14].
In addition to carbohydrate metabolism, the liver plays an important role in lipid, protein, and pigment metabolism [
13]. Lipid metabolism disorders were typical for patients with T2DM and contributed to liver diseases in our study. The liver’s excretory function is bile secretion, which includes direct bilirubin, creatinine, urea, and cholesterol [
13]. The total bilirubin level was increased in obese patients with and without T2DM, indicating abnormalities in the hepatic parenchyma.
Obesity, inflammation in adipose tissue, and IR are closely related to each other, forming a vicious circle that leads to steatosis [
15,
16]. Various serum factors in the blood of subjects with obesity induce inflammation and oxidative stress through various mechanisms and can negatively affect insulin signaling [
16]. We measured the levels of pro-inflammatory factors involved in the development of chronic subclinical inflammation. CRP and IL-6 levels in obese patients with T2DM were elevated in plasma and indicated systemic inflammation in this patient population. In obese patients with T2DM, CRP levels were lower than those in obese patients without T2DM. This is unusual; other authors have shown no difference in CRP levels between patients with steatosis and steatohepatitis [
17]. CRP is synthesized in the liver, and its decrease in patients with T2DM may result from liver dysfunction 130.
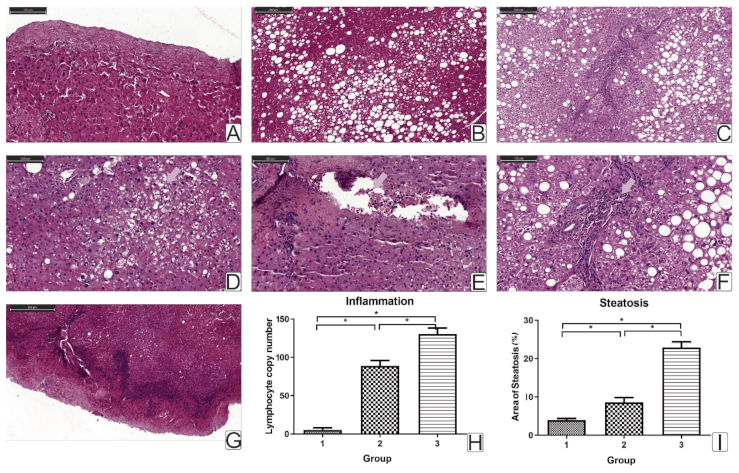
Most obese patients have been diagnosed with steatosis. In obese patients with T2DM, steatosis was higher than that in obese patients without T2DM. Steatosis leads to increased NFkB signaling through the activation of an inhibitor of nuclear factor-κB kinase β (IKKβ) [
18]. NFkB activation induces pro-inflammatory mediator production. Studies in mice have shown that activation of hepatocyte inflammation may be a link between initial metabolic stress and the death of hepatocytes and stimulation of fibrogenesis in steatohepatitis [
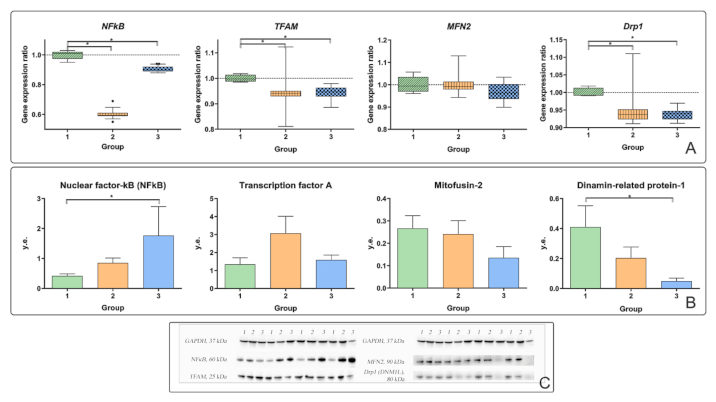
19]. In patients with T2DM, the production of NFkB protein in the liver was higher than that in control patients (
Figure 3B).
Hepatic inflammation in obesity leads to the progression of steatosis and steatohepatitis. We have shown that chronic inflammatory processes in obese patients lead to steatosis and liver lymphocyte infiltration. A study of liver biopsy histology showed an increase in the number of lymphocytes in obese patients with T2DM.
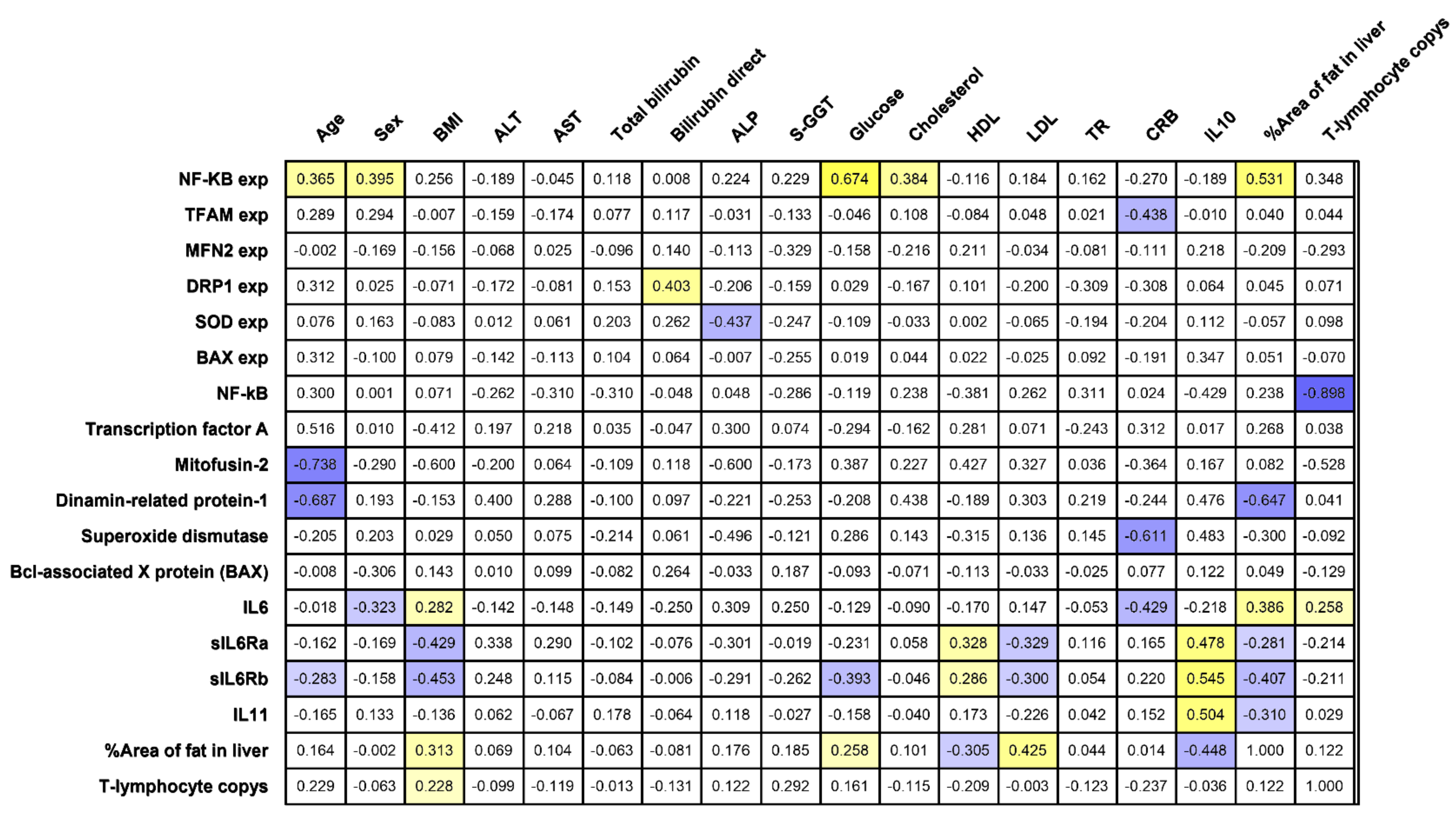
The percent steatosis and liver lymphocyte infiltration were associated with BMI (
r = 0.31,
r = 0.29,
p < 0.05) and with the level of plasma IL-6 (
r = 0.39,
r = 0.26,
p < 0.05) (
Figure 1). The area of fat inclusions was positively correlated with the levels of glucose (r = 0.25) and LDL (
r = 0.425) and negatively correlated with HDL (
r = −0.30) (
p < 0.05) in all obese patients, which formed a vicious circle with a positive inverse link. These facts characterize a pro-inflammatory component at the local (liver biopsy) and systemic (peripheral blood) levels in T2DM.
IL-6 binds to IL-6R or sIL-6R and initiates subsequent signaling through interaction with gp130 [
5,
6]. The IL-6/sIL-6R complex acts as an agonist of gp130-mediated IL-6 signaling and is called trans-signaling. This type of signaling broadens the spectrum of potential targets for IL-6 to virtually any cell type due to the ubiquitous expression of gp130. However, gp130 can also be found in a soluble form, which results in the inhibition of trans-IL-6 signaling while not affecting the classic type of signaling [
6]. Soluble gp130 (sgp130) is found in healthy people’s bloodstream, inhibiting the circulating IL-6 complex’s systemic response with sIL-6Ra [
6,
20,
21,
22].
Plasma sIL-6Ra and gp130/sIL-6Rb increased in obese patients without T2DM compared with obese patients with T2DM and controls. Both sIL-6Ra and gp130/sIL-6Rb receptors negatively correlated with BMI (r = −0.43, p < 0.05) (r = −0.45, p < 0.05) and LDL levels (r = −0.33, p < 0.05) (r = −0.30, p < 0.05) and positively correlated with HDL levels (r = 0.33, p < 0.05) (r = 0.29, p < 0.05). This emphasizes the protective role of compensatory increased sIL-6Ra and gp130/sIL-6Rb in maintaining normal lipid balance in obese patients. In patients without T2DM, fat infiltration hepatocytes were found in a much smaller number of cases than in patients with T2DM.
Classic IL-6 signaling in T cells plays a key role in the early stages of obesity, while trans-signals are more important later. Mice with hepatocyte-specific IL-6Rα deletion exhibit decreased insulin sensitivity and glucose tolerance, indicating a potential protective role for hepatic IL-6 signaling [
23]. Sgp130Fc can block trans-signaling IL-6 without affecting classical signaling. Therefore, future IL-6-based therapies may use a specific blockade of trans-IL-6 signaling rather than a global blockade of all IL-6 activities [
22]. The blockade of trans-IL-6 signals was sufficient to suppress the inflammatory state [
24]. Moreover, these data demonstrate the importance of sIL-6R levels for the pathophysiology of inflammatory diseases.
The anti-inflammatory and regenerative properties of IL-6 are mediated by a classical signaling mode, while pro-inflammatory responses in many pathological conditions involve trans-signaling.
Classic IL-6 signaling occurs via IL-6R and stimulates liver regeneration, protecting the liver from damage. An increase in plasma IL-6 levels leads to lipolysis in skeletal muscle and systemic fatty acid oxidation [
6]. Infusion of IL-6 into human AT enhances fatty acid and glycerol uptake, which promotes lipolysis. IL-6 suppressed CRP production. Plasma IL-6 levels were negatively correlated with CRP (
r = −0.43,
p < 0.05).
Activation of classic IL-6 signaling in steatosis leads to increased proliferation of liver cells and the replication stress due to the accumulation of replication errors, contributing to the progression of pathogenesis and degeneration of hepatocytes. In the group of obese patients with T2DM, almost all patients had dystrophy of hepatocytes and karyolysis manifestations; manifestations of intracellular cholestasis and lymphocytic infiltration of patients were noted. Oxidative stress in patients with NAFLD leads to the death of hepatocytes [
18,
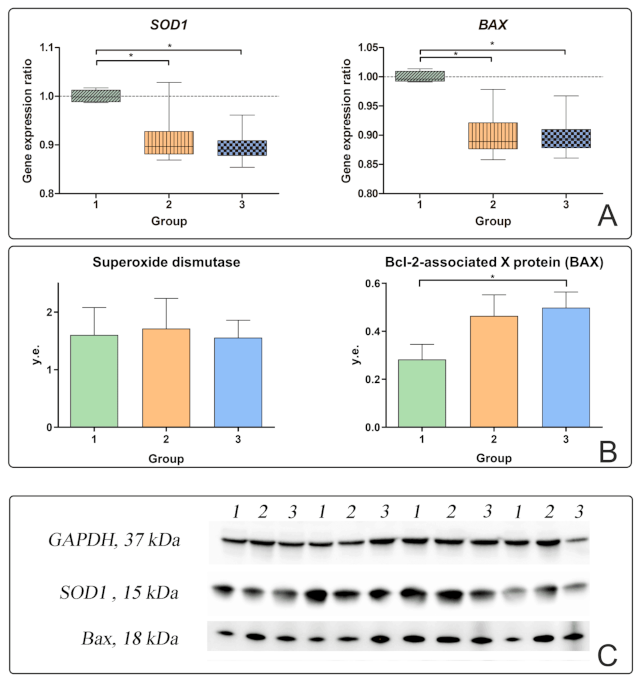
20]. In patients with T2DM, the production of pro-apoptotic BAX protein increased together with the production of NFkB. This proves that hepatic inflammation stimulates liver cell apoptosis. Liver cells infiltrated with fatty inclusions die through various mechanisms, including apoptosis, necrosis, and necroptosis. In an attempt to regenerate new cells, NASH progresses to fibrosis and cirrhosis with hepatocytes replaced by scar tissue collagen type 1 produced by stellate cells [
18].
The IL-6 begins to be synthesized in response to liver damage. The feedback mechanism requires more detailed study. In our study, no correlations of IL-6 in blood plasma with any characteristics of apoptosis were found. However, a negative correlation of IL-6 in blood plasma was found with the level of SOD1 gene expression in the liver in all obese patients (r = −0.815). The level of SOD1 gene expression in the liver decreased relative to the controls in obese patients with and without T2DM, which indicates a low antioxidant activity of the liver tissue. SOD protein production was not associated with any of the investigated factors. Against the background of an increase in plasma IL-6 levels and the development of liver cell inflammation and death, superoxide dismutase ceases to inhibit steatohepatitis in patients with T2DM.
IL-11 inhibits the induction of apoptosis in the liver. The highest levels of IL-11 were found in patients without T2DM and were negatively correlated with the production of BAX protein in liver biopsies of obese patients. IL-11 has been implicated in fibrosis of the heart [
25], liver [
12], and lung [
26]. Blocking IL-11 is considered a promising target for the fight against liver disease, with an established mechanism of action in both acute and chronic liver diseases [
27].
IL-11 is increased in obese patients without T2DM compared with patients with T2DM and controls. The IL-11 level is also positively correlated with IL-10, which may indicate its protective role in T2DM. However, IL-11 was not associated with lipid metabolism. Plasma IL-11 levels were negatively correlated with BAX protein production in liver biopsies. IL-11 is also soluble [
22,
28] and is blocked by sgp130 binding. How sgp130 affects the pathways triggered by these cytokines and how these pathways influence the effects of sgp130 deserve further study.
Strengthening mitochondrial dynamics is associated with increased cell proliferation during apoptosis. We did not find any significant changes in mitochondrial dynamics in obese patients without T2DM; therefore, we tested the hypothesis about the effect of the microenvironment of chronic subclinical inflammation on the liver in patients with T2DM. In obese patients, mitochondrial dysfunction, mediated by an increase in TFAM in the presence of an inflammatory process in the liver, may be a factor contributing to the formation of T2DM.
In this case, oxidative stress can cause changes in mtDNA. mtDNA is especially sensitive to oxidative damage because it is directly adjacent to ROS production and DNA repair systems. In addition, oxidative damage to nuclear DNA can lead to mitochondrial dysfunction by disrupting the transcription of nuclear-encoded mitochondrial genes. For example, the expression levels of genes involved in mitochondrial metabolism and biogenesis, such as
TFAM,
NRF-2, and
PGC-1α, are reduced in NAFLD [
29].
Plasma IL-6 suppresses the production of TFAM protein in liver biopsies (
r = −0.615,
p < 0.05). TFAM is a mitochondrial transcription factor responsible for the regulation of division and replication. TFAM protein production was independent of other parameters and was associated only with IL-6 levels. IL-6 can regulate the processes of replication and transcription of mitochondrial DNA. Considering the important role of IL-6 and its receptors in regulating lipid metabolism, its effect on fatty liver infiltration deserves special attention. TFAM is a multifunctional DNA-binding protein required for the activation of transcription and organization of mtDNA [
29]. TFAM is one of the key regulators of mtDNA transcription associated with altered mitochondrial division/fusion processes and mtDNA replication [
29]. Thus, in patients with T2DM, an increase in TFAM transcription and a decrease in its protein level were observed in in vitro models [
29]. The TFAM protein is synthesized in the cell and transported into mitochondria, where it activates transcription and participates in mtDNA replication [
29]. In obese patients with T2DM, the level of
TFAM gene expression and TFAM protein production increased in some patients, but these changes were not significant. The
TFAM gene plays an important role in cell physiology, participates in mtDNA maintenance, and regulates the number of mtDNA copies [
29]. Therefore, an increase in the expression level of the
TFAM gene may indicate an increase in mtDNA transcription under conditions of inflammation of the liver tissue. The expression level of the
TFAM gene was positively correlated with the expression level of the
DRP1 (
r = 0.62) and
SOD1 (
r = 0.53) genes.
Proteins that regulate the fusion and division of mitochondria are mainly responsible for changing the dynamics of mitochondria: MFN1, MFN2, DRP1, and FIS1. MFN2 is required for mitochondrial fusion, while DRP1 is required for mitochondrial division [
29,
30]. Mitofusin is located on the outer mitochondrial membrane and promotes fusion of the outer mitochondrial membranes. In addition to its role in mitochondrial fusion, mitofusin may play a role in developing oxidative stress during metabolic syndrome. For the division processes, one of the main proteins is DRP1. DRP1 is found mainly in the cytosol and is recruited to the outer mitochondrial membrane by the FIS1 protein located on the outer membrane.
In obese patients with T2DM, the level of gene expression and production of MFN2 and DRP1 proteins decreased. In obese patients with T2DM, the expression of the
DRP1 gene and the production of the DRP-1 protein were reduced relative to the controls (
p < 0.05) (
Figure 3A,B).
It is assumed that mitochondrial fusion processes are induced under conditions that require the optimization of mitochondrial bioenergetics. At the same time, the processes of division are associated with the degradation of mitochondria. Consequently, the processes of fusion and division are induced under conditions of mitochondrial damage. Repression of MFN2 is associated with a decrease in cellular metabolism. However, no changes in MFN2 were found in our study.
Drp1 is also involved in the regulation of apoptosis [
31]. Mitophagy, a specialized form of macroautophagy, mediates selective degradation of damaged mitochondria [
7]. In response to reduced membrane potential, ubiquitin is activated in mitochondria, removing Mfn and several other outer mitochondrial membrane proteins. Ultimately, this leads to the capture of mitochondria by the autophagosome and subsequent catabolism in the lysosomes [
7]. The expression level of the
DRP1 gene was positively correlated with the level of BAX gene expression in the liver (
r = 0.40,
p < 0.05). With an increase in inflammation in the liver, the processes of mitophagy and mitochondrial division are activated.
Dysregulation of energy metabolism in insulin-dependent tissues can be a crucial cause of T2DM. With the development of T2DM, the processes of mitochondrial and peroxisomal oxidation are triggered; the latter’s activation is associated with excessive accumulation of ROS and the development of oxidative stress [
32]. Under the influence of oxidative stress and increased ROS production, obesity disrupts the functioning of mitochondria, represented by a decrease in the synthesis of ATP [
32] by mitochondria, increased mitophagy, and processes of its division. Thus, in liver biopsies of patients with T2DM, replication processes, transcription, and division were reduced.

.JPG)
 ,
, 






{kind=link}
{kind=link}
{kind=link}
{kind=link}