
A Role for Mycobacterium tuberculosis Sigma Factor C in Copper Nutritional Immunity

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Absence of Significant Growth or Global Gene Expression Differences after Deletion of sigC from M. tuberculosis Strain Erdman Cultured in 7H9 Medium
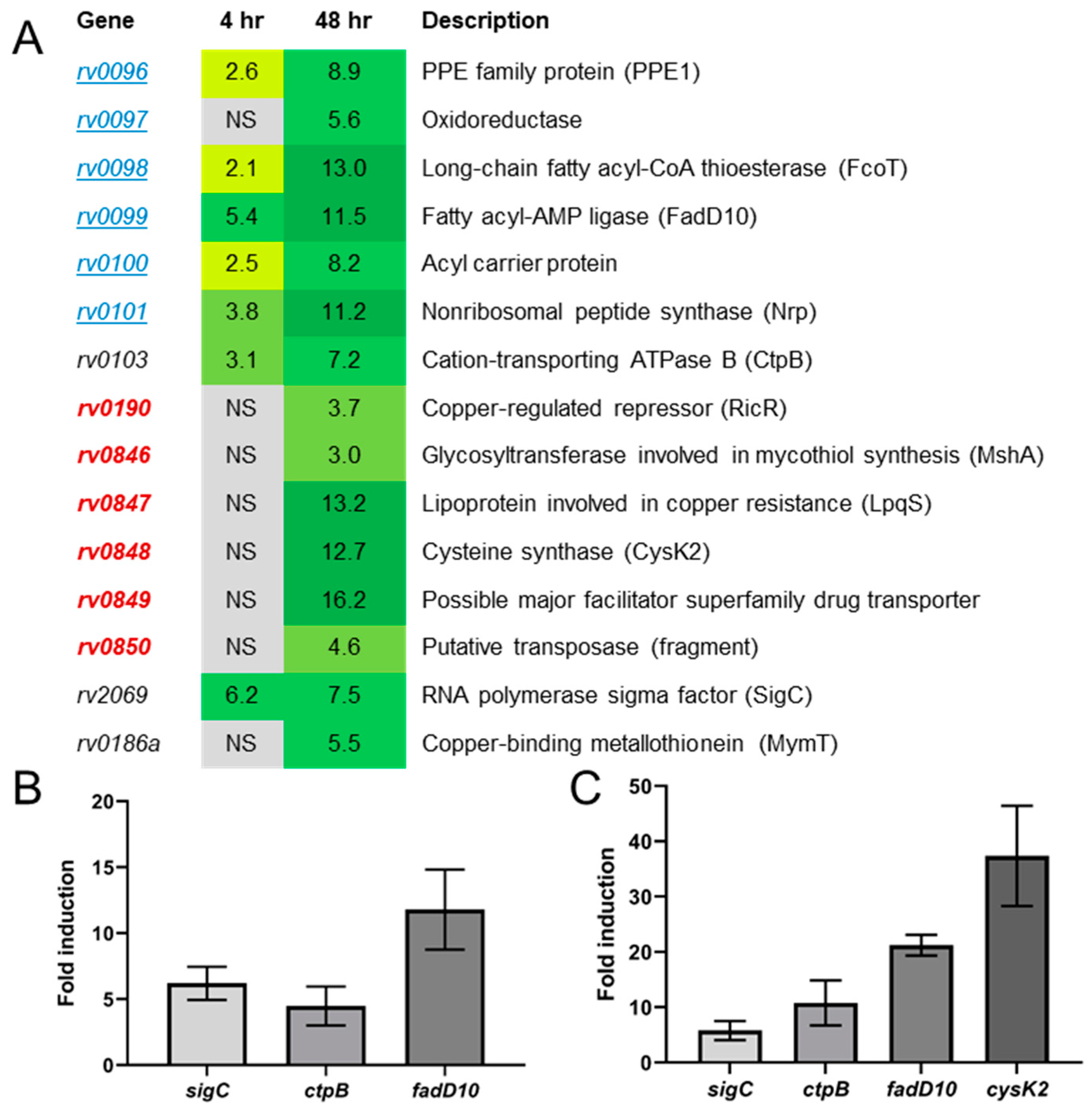
2.2. Artificial sigC Induction Points to a Role for SigC in Metal Transport
2.3. Identification of an In Vitro Phenotype for ΔsigC
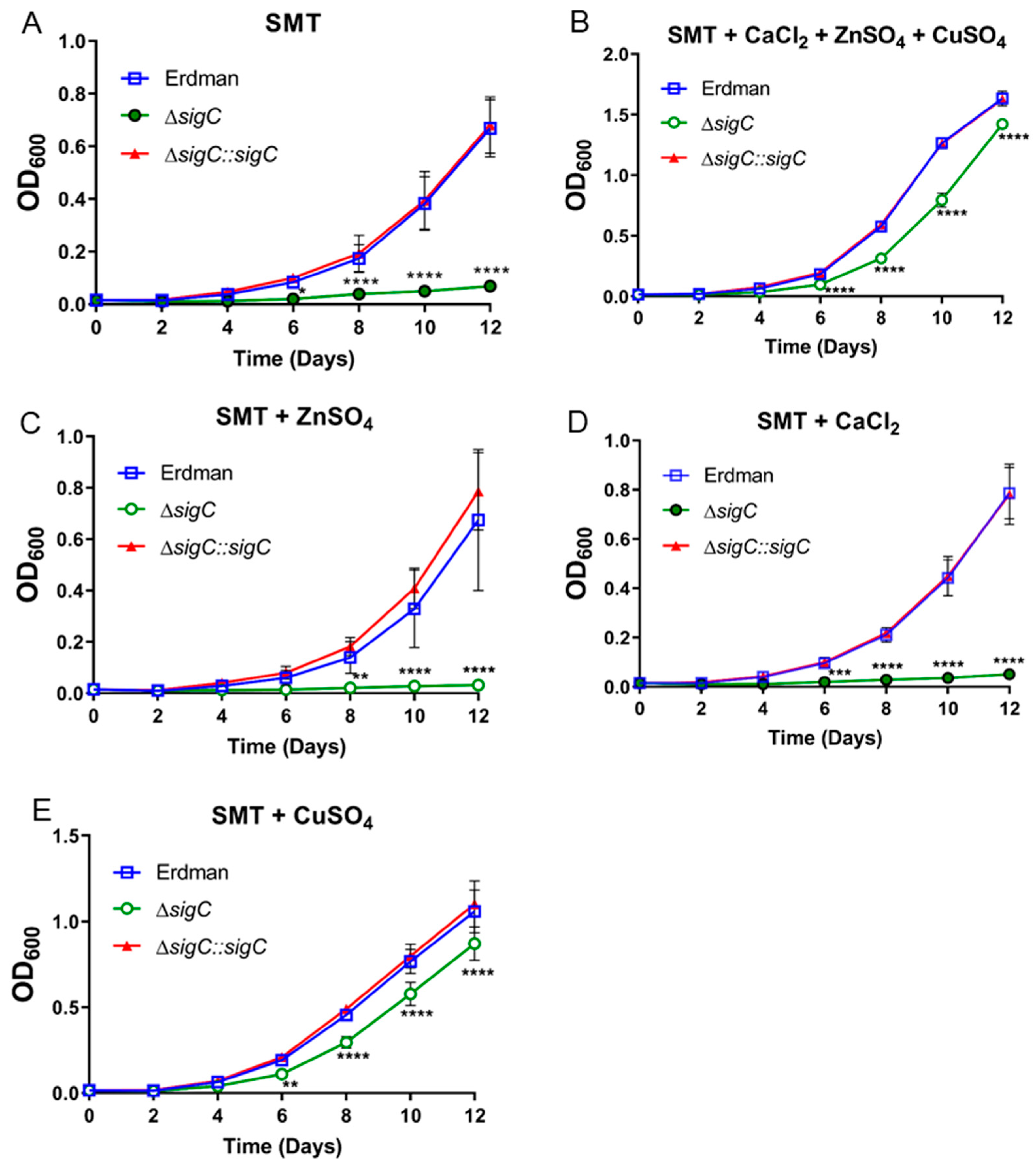
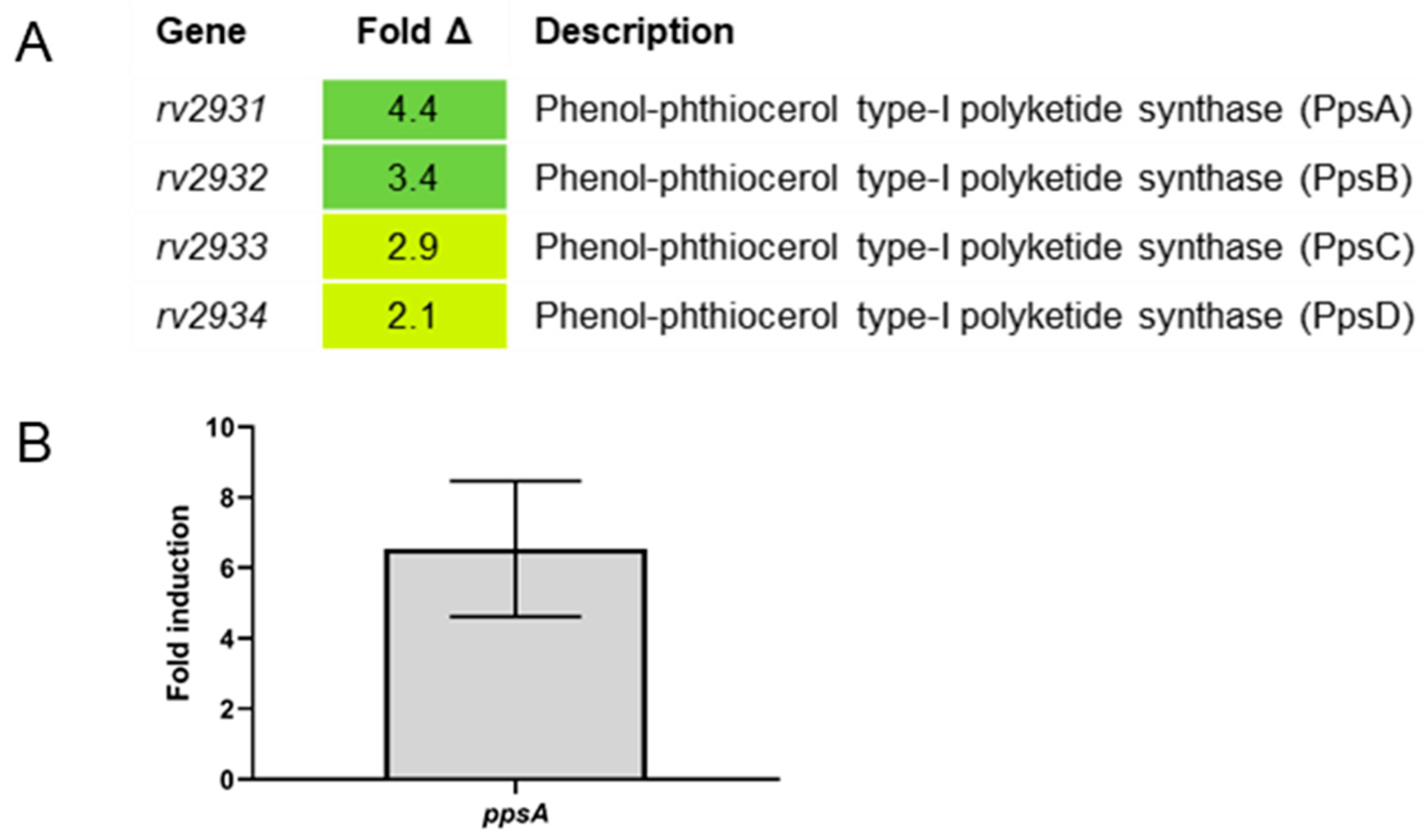
2.4. Differential Gene Expression between Erdman and ΔsigC in SMT Supports SigC Function in Copper Uptake
2.5. Copper Supplementation Eliminates Most Transcriptional Differences between ΔsigC and Erdman
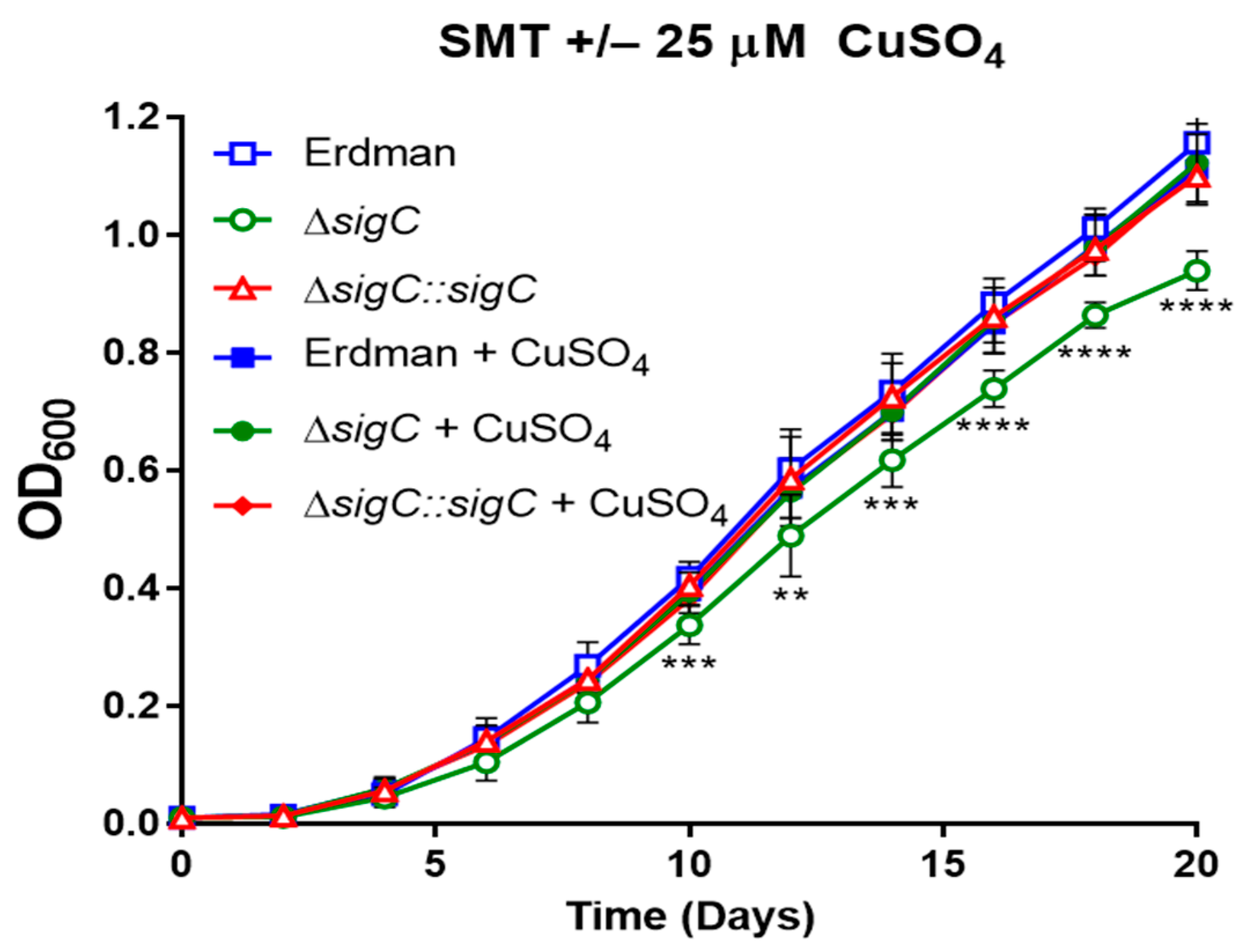
2.6. Complete Reversal of the ΔsigC Growth Defect by 25 µM Copper Sulfate Addition
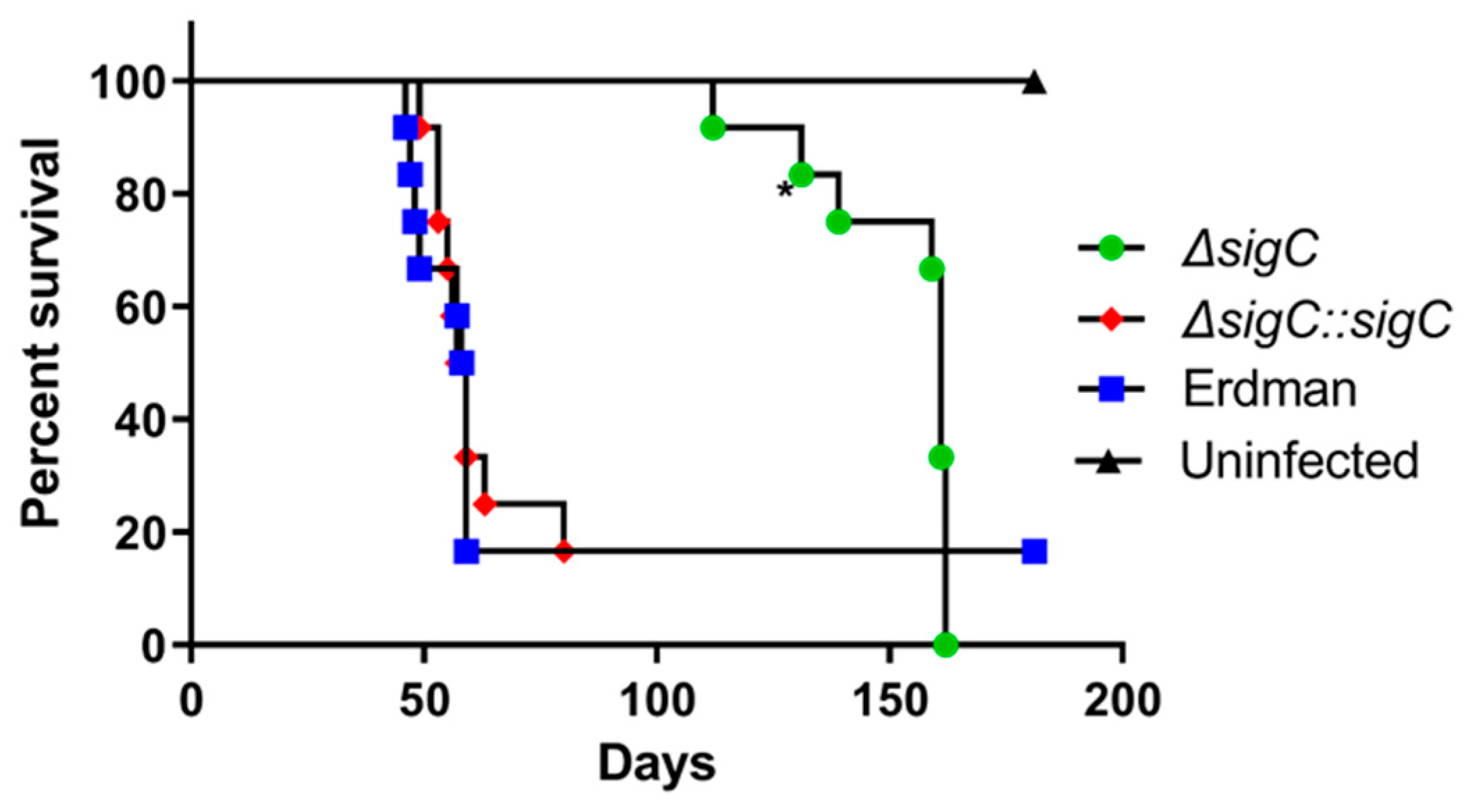
2.7. Attenuation of M. tuberculosis Erdman ΔsigC Virulence in SCID Mice
2.8. Future Directions
3. Materials and Methods
3.1. Bacterial Strains and Culture Conditions
3.2. RNA Isolation
3.3. Microarray Analyses
3.4. Quantitative RT-PCR Assays
3.5. Animal Infection Studies
3.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. Global Tuberculosis Report; World Health Organization: Geneva, Switzerland, 2020. [Google Scholar]
- Houben, R.M.; Dodd, P.J. The Global Burden of Latent Tuberculosis Infection: A Re-estimation Using Mathematical Modelling. PLoS Med. 2016, 13, e1002152. [Google Scholar] [CrossRef] [Green Version]
- Cole, S.T.; Brosch, R.; Parkhill, J.; Garnier, T.; Churcher, C.; Harris, D.; Gordon, S.V.; Eiglmeier, K.; Gas, S.; Barry, C.E., 3rd; et al. Deciphering the biology of Mycobacterium tuberculosis from the complete genome sequence. Nature 1998, 393, 537–544. [Google Scholar] [CrossRef] [PubMed]
- Manganelli, R. Sigma Factors: Key Molecules in Mycobacterium tuberculosis Physiology and Virulence. Microbiol. Spectr. 2014, 2, MGM2-0007-2013. [Google Scholar] [CrossRef] [Green Version]
- Karls, R.K.; Guarner, J.; McMurray, D.N.; Birkness, K.A.; Quinn, F.D. Examination of Mycobacterium tuberculosis sigma factor mutants using low-dose aerosol infection of guinea pigs suggests a role for SigC in pathogenesis. Microbiology 2006, 152, 1591–1600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdul-Majid, K.B.; Ly, L.H.; Converse, P.J.; Geiman, D.E.; McMurray, D.N.; Bishai, W.R. Altered cellular infiltration and cytokine levels during early Mycobacterium tuberculosis sigC mutant infection are associated with late-stage disease attenuation and milder immunopathology in mice. BMC Microbiol. 2008, 8, 151. [Google Scholar] [CrossRef] [Green Version]
- Sun, R.; Converse, P.J.; Ko, C.; Tyagi, S.; Morrison, N.E.; Bishai, W.R. Mycobacterium tuberculosis ECF sigma factor sigC is required for lethality in mice and for the conditional expression of a defined gene set. Mol. Microbiol. 2004, 52, 25–38. [Google Scholar] [CrossRef] [PubMed]
- Chang, A.; Smollett, K.L.; Gopaul, K.K.; Chan, B.H.; Davis, E.O. Mycobacterium tuberculosis H37Rv sigC is expressed from two promoters but is not auto-regulatory. Tuberculosis 2012, 92, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Manganelli, R.; Dubnau, E.; Tyagi, S.; Kramer, F.R.; Smith, I. Differential expression of 10 sigma factor genes in Mycobacterium tuberculosis. Mol. Microbiol. 1999, 31, 715–724. [Google Scholar] [CrossRef]
- Rustad, T.R.; Minch, K.J.; Ma, S.; Winkler, J.K.; Hobbs, S.; Hickey, M.; Brabant, W.; Turkarslan, S.; Price, N.D.; Baliga, N.S.; et al. Mapping and manipulating the Mycobacterium tuberculosis transcriptome using a transcription factor overexpression-derived regulatory network. Genome Biol. 2014, 15, 502. [Google Scholar] [CrossRef]
- Rodrigue, S.; Brodeur, J.; Jacques, P.E.; Gervais, A.L.; Brzezinski, R.; Gaudreau, L. Identification of mycobacterial sigma factor binding sites by chromatin immunoprecipitation assays. J. Bacteriol. 2007, 189, 1505–1513. [Google Scholar] [CrossRef] [Green Version]
- Andreini, C.; Bertini, I.; Cavallaro, G.; Holliday, G.L.; Thornton, J.M. Metal ions in biological catalysis: From enzyme databases to general principles. J. Biol. Inorg. Chem. 2008, 13, 1205–1218. [Google Scholar] [CrossRef]
- Hood, M.I.; Skaar, E.P. Nutritional immunity: Transition metals at the pathogen-host interface. Nat. Rev. Microbiol. 2012, 10, 525–537. [Google Scholar] [CrossRef]
- Weinberg, E.D. Nutritional immunity. Host’s attempt to withold iron from microbial invaders. JAMA 1975, 231, 39–41. [Google Scholar] [CrossRef] [PubMed]
- Hennigar, S.R.; McClung, J.P. Nutritional Immunity: Starving Pathogens of Trace Minerals. Am. J. Lifestyle Med. 2016, 10, 170–173. [Google Scholar] [CrossRef] [PubMed]
- Darwin, K.H. Mycobacterium tuberculosis and Copper: A Newly Appreciated Defense against an Old Foe? J. Biol. Chem. 2015, 290, 18962–18966. [Google Scholar] [CrossRef] [Green Version]
- Fu, Y.; Chang, F.M.; Giedroc, D.P. Copper transport and trafficking at the host-bacterial pathogen interface. Acc. Chem. Res. 2014, 47, 3605–3613. [Google Scholar] [CrossRef]
- Sassetti, C.M.; Boyd, D.H.; Rubin, E.J. Genes required for mycobacterial growth defined by high density mutagenesis. Mol. Microbiol. 2003, 48, 77–84. [Google Scholar] [CrossRef]
- Griffin, J.E.; Gawronski, J.D.; Dejesus, M.A.; Ioerger, T.R.; Akerley, B.J.; Sassetti, C.M. High-resolution phenotypic profiling defines genes essential for mycobacterial growth and cholesterol catabolism. PLoS Pathog. 2011, 7, e1002251. [Google Scholar] [CrossRef] [Green Version]
- DeJesus, M.A.; Gerrick, E.R.; Xu, W.; Park, S.W.; Long, J.E.; Boutte, C.C.; Rubin, E.J.; Schnappinger, D.; Ehrt, S.; Fortune, S.M.; et al. Comprehensive Essentiality Analysis of the Mycobacterium tuberculosis Genome via Saturating Transposon Mutagenesis. mBio 2017, 8, e02133-16. [Google Scholar] [CrossRef] [Green Version]
- Koch, A.; Mizrahi, V. Mycobacterium tuberculosis. Trends Microbiol. 2018, 26, 555–556. [Google Scholar] [CrossRef] [PubMed]
- Rodrigue, S.; Provvedi, R.; Jacques, P.E.; Gaudreau, L.; Manganelli, R. The sigma factors of Mycobacterium tuberculosis. FEMS Microbiol. Rev. 2006, 30, 926–941. [Google Scholar] [CrossRef] [Green Version]
- Leon-Torres, A.; Arango, E.; Castillo, E.; Soto, C.Y. CtpB is a plasma membrane copper (I) transporting P-type ATPase of Mycobacterium tuberculosis. Biol. Res. 2020, 53, 6. [Google Scholar] [CrossRef]
- Harris, N.C.; Sato, M.; Herman, N.A.; Twigg, F.; Cai, W.; Liu, J.; Zhu, X.; Downey, J.; Khalaf, R.; Martin, J.; et al. Biosynthesis of isonitrile lipopeptides by conserved nonribosomal peptide synthetase gene clusters in Actinobacteria. Proc. Natl. Acad. Sci. USA 2017, 114, 7025–7030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Zhu, M.; Zhang, Q.; Zhang, X.; Yang, P.; Liu, Z.; Deng, Y.; Zhu, Y.; Huang, X.; Han, L.; et al. Diisonitrile Natural Product SF2768 Functions As a Chalkophore That Mediates Copper Acquisition in Streptomyces thioluteus. ACS Chem. Biol. 2017, 12, 3067–3075. [Google Scholar] [CrossRef]
- Festa, R.A.; Jones, M.B.; Butler-Wu, S.; Sinsimer, D.; Gerads, R.; Bishai, W.R.; Peterson, S.N.; Darwin, K.H. A novel copper-responsive regulon in Mycobacterium tuberculosis. Mol. Microbiol. 2011, 79, 133–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gold, B.; Deng, H.; Bryk, R.; Vargas, D.; Eliezer, D.; Roberts, J.; Jiang, X.; Nathan, C. Identification of a copper-binding metallothionein in pathogenic mycobacteria. Nat. Chem. Biol. 2008, 4, 609–616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimbro, M.J. Difco & BBL Manual: Manual of Microbiological Culture Media, 2nd ed.; Becton, Dickinson and Company: Sparks, MD, USA, 2009. [Google Scholar]
- Parish, T.; Smith, D.A.; Roberts, G.; Betts, J.; Stoker, N.G. The senX3-regX3 two-component regulatory system of Mycobacterium tuberculosis is required for virulence. Microbiology 2003, 149, 1423–1435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rifat, D.; Belchis, D.A.; Karakousis, P.C. senX3-independent contribution of regX3 to Mycobacterium tuberculosis virulence. BMC Microbiol. 2014, 14, 265. [Google Scholar] [CrossRef] [Green Version]
- Marcus, S.A.; Sidiropoulos, S.W.; Steinberg, H.; Talaat, A.M. CsoR Is Essential for Maintaining Copper Homeostasis in Mycobacterium tuberculosis. PLoS ONE 2016, 11, e0151816. [Google Scholar] [CrossRef]
- He, W.; Soll, C.E.; Chavadi, S.S.; Zhang, G.; Warren, J.D.; Quadri, L.E. Cooperation between a coenzyme A-independent stand-alone initiation module and an iterative type I polyketide synthase during synthesis of mycobacterial phenolic glycolipids. J. Am. Chem. Soc. 2009, 131, 16744–16750. [Google Scholar] [CrossRef] [Green Version]
- Park, H.D.; Guinn, K.M.; Harrell, M.I.; Liao, R.; Voskuil, M.I.; Tompa, M.; Schoolnik, G.K.; Sherman, D.R. Rv3133c/dosR is a transcription factor that mediates the hypoxic response of Mycobacterium tuberculosis. Mol. Microbiol. 2003, 48, 833–843. [Google Scholar] [CrossRef] [Green Version]
- Rustad, T.R.; Harrell, M.I.; Liao, R.; Sherman, D.R. The enduring hypoxic response of Mycobacterium tuberculosis. PLoS ONE 2008, 3, e1502. [Google Scholar] [CrossRef] [PubMed]
- Fenhalls, G.; Stevens, L.; Moses, L.; Bezuidenhout, J.; Betts, J.C.; Helden Pv, P.; Lukey, P.T.; Duncan, K. In situ detection of Mycobacterium tuberculosis transcripts in human lung granulomas reveals differential gene expression in necrotic lesions. Infect. Immun. 2002, 70, 6330–6338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dahl, J.L.; Kraus, C.N.; Boshoff, H.I.; Doan, B.; Foley, K.; Avarbock, D.; Kaplan, G.; Mizrahi, V.; Rubin, H.; Barry, C.E., 3rd. The role of RelMtb-mediated adaptation to stationary phase in long-term persistence of Mycobacterium tuberculosis in mice. Proc. Natl. Acad. Sci. USA 2003, 100, 10026–10031. [Google Scholar] [CrossRef] [Green Version]
- Sherman, D.R.; Mdluli, K.; Hickey, M.J.; Barry, C.E., 3rd; Stover, C.K. AhpC, oxidative stress and drug resistance in Mycobacterium tuberculosis. Biofactors 1999, 10, 211–217. [Google Scholar] [CrossRef]
- Saxena, A.; Srivastava, V.; Srivastava, R.; Srivastava, B.S. Identification of genes of Mycobacterium tuberculosis upregulated during anaerobic persistence by fluorescence and kanamycin resistance selection. Tuberculosis 2008, 88, 518–525. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Lee, B.S.; Shin, S.J.; Kim, H.J.; Park, J.K. Differentially expressed genes in Mycobacterium tuberculosis H37Rv under mild acidic and hypoxic conditions. J. Med. Microbiol. 2008, 57, 1473–1480. [Google Scholar] [CrossRef]
- Betts, J.C.; Lukey, P.T.; Robb, L.C.; McAdam, R.A.; Duncan, K. Evaluation of a nutrient starvation model of Mycobacterium tuberculosis persistence by gene and protein expression profiling. Mol. Microbiol. 2002, 43, 717–731. [Google Scholar] [CrossRef]
- Farhana, A.; Guidry, L.; Srivastava, A.; Singh, A.; Hondalus, M.K.; Steyn, A.J.C. Reductive Stress in Microbes: Implications for Understanding Mycobacterium tuberculosis Diseases and Persistence. In Advances in Microbial Physiology; Poole, R.K., Ed.; Elsevier Academic Press: Oxford, UK, 2010; Volume 57, pp. 43–117. [Google Scholar]
- Rhee, K.Y.; de Carvalho, L.P.; Bryk, R.; Ehrt, S.; Marrero, J.; Park, S.W.; Schnappinger, D.; Venugopal, A.; Nathan, C. Central carbon metabolism in Mycobacterium tuberculosis: An unexpected frontier. Trends Microbiol. 2011, 19, 307–314. [Google Scholar] [CrossRef] [Green Version]
- Ehrt, S.; Rhee, K. Mycobacterium tuberculosis metabolism and host interaction: Mysteries and paradoxes. Curr. Top. Microbiol. Immunol. 2013, 374, 163–188. [Google Scholar] [CrossRef]
- Baughn, A.D.; Rhee, K.Y. Metabolomics of Central Carbon Metabolism in Mycobacterium tuberculosis. Microbiol. Spectr. 2014, 2, 323–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehrt, S.; Schnappinger, D.; Rhee, K.Y. Metabolic principles of persistence and pathogenicity in Mycobacterium tuberculosis. Nat. Rev. Microbiol. 2018, 16, 496–507. [Google Scholar] [CrossRef] [PubMed]
- Cook, G.M.; Hards, K.; Vilcheze, C.; Hartman, T.; Berney, M. Energetics of Respiration and Oxidative Phosphorylation in Mycobacteria. Microbiol. Spectr. 2014, 2, 389–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marrero, J.; Rhee, K.Y.; Schnappinger, D.; Pethe, K.; Ehrt, S. Gluconeogenic carbon flow of tricarboxylic acid cycle intermediates is critical for Mycobacterium tuberculosis to establish and maintain infection. Proc. Natl. Acad. Sci. USA 2010, 107, 9819–9824. [Google Scholar] [CrossRef] [Green Version]
- Nienaber, L.; Cave-Freeman, E.; Cross, M.; Mason, L.; Bailey, U.M.; Amani, P.; Davis, R.A.; Taylor, P.; Hofmann, A. Chemical probing suggests redox-regulation of the carbonic anhydrase activity of mycobacterial Rv1284. FEBS J. 2015, 282, 2708–2721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beste, D.J.; Bonde, B.; Hawkins, N.; Ward, J.L.; Beale, M.H.; Noack, S.; Noh, K.; Kruger, N.J.; Ratcliffe, R.G.; McFadden, J. (1)(3)C metabolic flux analysis identifies an unusual route for pyruvate dissimilation in mycobacteria which requires isocitrate lyase and carbon dioxide fixation. PLoS Pathog. 2011, 7, e1002091. [Google Scholar] [CrossRef] [Green Version]
- Giuffre, A.; Borisov, V.B.; Arese, M.; Sarti, P.; Forte, E. Cytochrome bd oxidase and bacterial tolerance to oxidative and nitrosative stress. Biochim. Biophys. Acta 2014, 1837, 1178–1187. [Google Scholar] [CrossRef] [Green Version]
- Voskuil, M.I.; Visconti, K.C.; Schoolnik, G.K. Mycobacterium tuberculosis gene expression during adaptation to stationary phase and low-oxygen dormancy. Tuberculosis 2004, 84, 218–227. [Google Scholar] [CrossRef]
- Milano, A.; Pasca, M.R.; Provvedi, R.; Lucarelli, A.P.; Manina, G.; Ribeiro, A.L.; Manganelli, R.; Riccardi, G. Azole resistance in Mycobacterium tuberculosis is mediated by the MmpS5-MmpL5 efflux system. Tuberculosis 2009, 89, 84–90. [Google Scholar] [CrossRef]
- Gold, B.; Rodriguez, G.M.; Marras, S.A.; Pentecost, M.; Smith, I. The Mycobacterium tuberculosis IdeR is a dual functional regulator that controls transcription of genes involved in iron acquisition, iron storage and survival in macrophages. Mol. Microbiol. 2001, 42, 851–865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sala, C.; Haouz, A.; Saul, F.A.; Miras, I.; Rosenkrands, I.; Alzari, P.M.; Cole, S.T. Genome-wide regulon and crystal structure of BlaI (Rv1846c) from Mycobacterium tuberculosis. Mol. Microbiol. 2009, 71, 1102–1116. [Google Scholar] [CrossRef]
- Roberts, G.; Vadrevu, I.S.; Madiraju, M.V.; Parish, T. Control of CydB and GltA1 expression by the SenX3 RegX3 two component regulatory system of Mycobacterium tuberculosis. PLoS ONE 2011, 6, e21090. [Google Scholar] [CrossRef]
- Singh, A.; Crossman, D.K.; Mai, D.; Guidry, L.; Voskuil, M.I.; Renfrow, M.B.; Steyn, A.J. Mycobacterium tuberculosis WhiB3 maintains redox homeostasis by regulating virulence lipid anabolism to modulate macrophage response. PLoS Pathog. 2009, 5, e1000545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simeone, R.; Constant, P.; Malaga, W.; Guilhot, C.; Daffe, M.; Chalut, C. Molecular dissection of the biosynthetic relationship between phthiocerol and phthiodiolone dimycocerosates and their critical role in the virulence and permeability of Mycobacterium tuberculosis. FEBS J. 2007, 274, 1957–1969. [Google Scholar] [CrossRef]
- Raman, S.; Puyang, X.; Cheng, T.Y.; Young, D.C.; Moody, D.B.; Husson, R.N. Mycobacterium tuberculosis SigM positively regulates Esx secreted protein and nonribosomal peptide synthetase genes and down regulates virulence-associated surface lipid synthesis. J. Bacteriol. 2006, 188, 8460–8468. [Google Scholar] [CrossRef] [Green Version]
- Li, D.D.; Zhang, W.; Wang, Z.Y.; Zhao, P. Serum Copper, Zinc, and Iron Levels in Patients with Alzheimer’s Disease: A Meta-Analysis of Case-Control Studies. Front. Aging. Neurosci. 2017, 9, 300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhatt, K.; Machado, H.; Osorio, N.S.; Sousa, J.; Cardoso, F.; Magalhaes, C.; Chen, B.; Chen, M.; Kim, J.; Singh, A.; et al. A Nonribosomal Peptide Synthase Gene Driving Virulence in Mycobacterium tuberculosis. mSphere 2018, 3, e00352-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hotter, G.S.; Wards, B.J.; Mouat, P.; Besra, G.S.; Gomes, J.; Singh, M.; Bassett, S.; Kawakami, P.; Wheeler, P.R.; de Lisle, G.W.; et al. Transposon mutagenesis of Mb0100 at the ppe1-nrp locus in Mycobacterium bovis disrupts phthiocerol dimycocerosate (PDIM) and glycosylphenol-PDIM biosynthesis, producing an avirulent strain with vaccine properties at least equal to those of M. bovis BCG. J. Bacteriol. 2005, 187, 2267–2277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhar, N.; McKinney, J.D. Mycobacterium tuberculosis persistence mutants identified by screening in isoniazid-treated mice. Proc. Natl. Acad. Sci. USA 2010, 107, 12275–12280. [Google Scholar] [CrossRef] [Green Version]
- Camacho, L.R.; Constant, P.; Raynaud, C.; Laneelle, M.A.; Triccas, J.A.; Gicquel, B.; Daffe, M.; Guilhot, C. Analysis of the phthiocerol dimycocerosate locus of Mycobacterium tuberculosis. Evidence that this lipid is involved in the cell wall permeability barrier. J. Biol. Chem. 2001, 276, 19845–19854. [Google Scholar] [CrossRef] [Green Version]
- Astarie-Dequeker, C.; Le Guyader, L.; Malaga, W.; Seaphanh, F.K.; Chalut, C.; Lopez, A.; Guilhot, C. Phthiocerol dimycocerosates of M. tuberculosis participate in macrophage invasion by inducing changes in the organization of plasma membrane lipids. PLoS Pathog. 2009, 5, e1000289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, J.S.; Chen, B.; McNeil, M.; Jacobs, W.R., Jr. Complex lipid determines tissue-specific replication of Mycobacterium tuberculosis in mice. Nature 1999, 402, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Kirksey, M.A.; Tischler, A.D.; Simeone, R.; Hisert, K.B.; Uplekar, S.; Guilhot, C.; McKinney, J.D. Spontaneous phthiocerol dimycocerosate-deficient variants of Mycobacterium tuberculosis are susceptible to gamma interferon-mediated immunity. Infect. Immun. 2011, 79, 2829–2838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braunstein, M.; Bardarov, S.S.; Jacobs, W.R., Jr. Genetic methods for deciphering virulence determinants of Mycobacterium tuberculosis. Methods Enzymol. 2002, 358, 67–99. [Google Scholar] [CrossRef] [PubMed]
- Kong, D.; Kunimoto, D.Y. Secretion of human interleukin 2 by recombinant Mycobacterium bovis BCG. Infect. Immun. 1995, 63, 799–803. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.H.; Dudoit, S.; Luu, P.; Lin, D.M.; Peng, V.; Ngai, J.; Speed, T.P. Normalization for cDNA microarray data: A robust composite method addressing single and multiple slide systematic variation. Nucleic Acids Res. 2002, 30, e15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Grosse-Siestrup, B.T. Examination of the SigC Regulon and Cobalamin Biosynthesis in Mycobacterium tuberculosis. Ph.D. Thesis, University of Georgia, Athens, Georgia, 2012. [Google Scholar]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grosse-Siestrup, B.T.; Gupta, T.; Helms, S.; Tucker, S.L.; Voskuil, M.I.; Quinn, F.D.; Karls, R.K. A Role for Mycobacterium tuberculosis Sigma Factor C in Copper Nutritional Immunity. Int. J. Mol. Sci. 2021, 22, 2118. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22042118
Grosse-Siestrup BT, Gupta T, Helms S, Tucker SL, Voskuil MI, Quinn FD, Karls RK. A Role for Mycobacterium tuberculosis Sigma Factor C in Copper Nutritional Immunity. International Journal of Molecular Sciences. 2021; 22(4):2118. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22042118
Chicago/Turabian StyleGrosse-Siestrup, Benjamin T., Tuhina Gupta, Shelly Helms, Samantha L. Tucker, Martin I. Voskuil, Frederick D. Quinn, and Russell K. Karls. 2021. "A Role for Mycobacterium tuberculosis Sigma Factor C in Copper Nutritional Immunity" International Journal of Molecular Sciences 22, no. 4: 2118. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22042118