
Fructose Induces Fluconazole Resistance in Candida albicans through Activation of Mdr1 and Cdr1 Transporters

Abstract
:1. Introduction
2. Results and Discussion
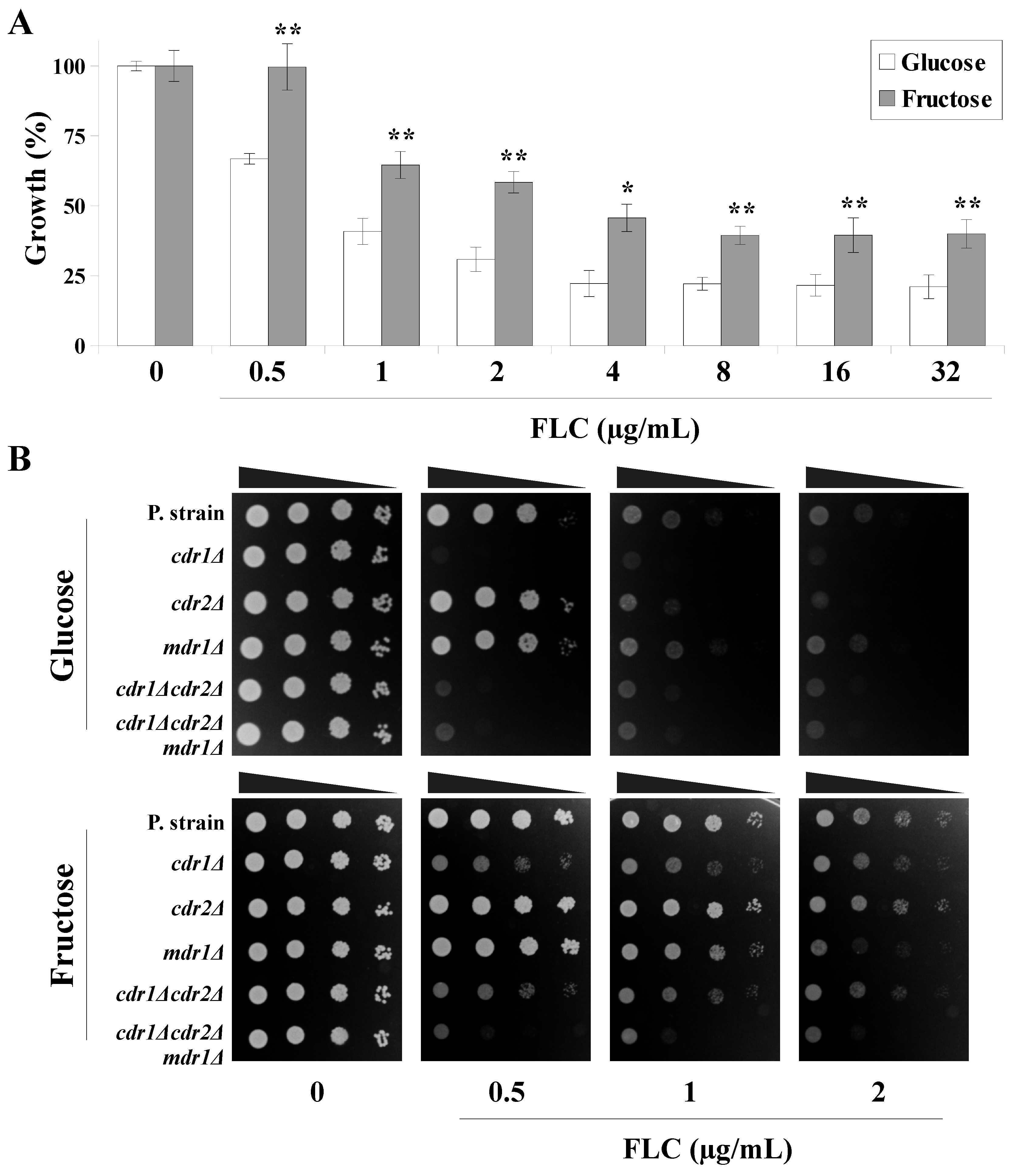
2.1. Fructose as a Carbon Source Promotes FLC Resistance and Increased Efflux Activities of Cdr1p and Mdr1p
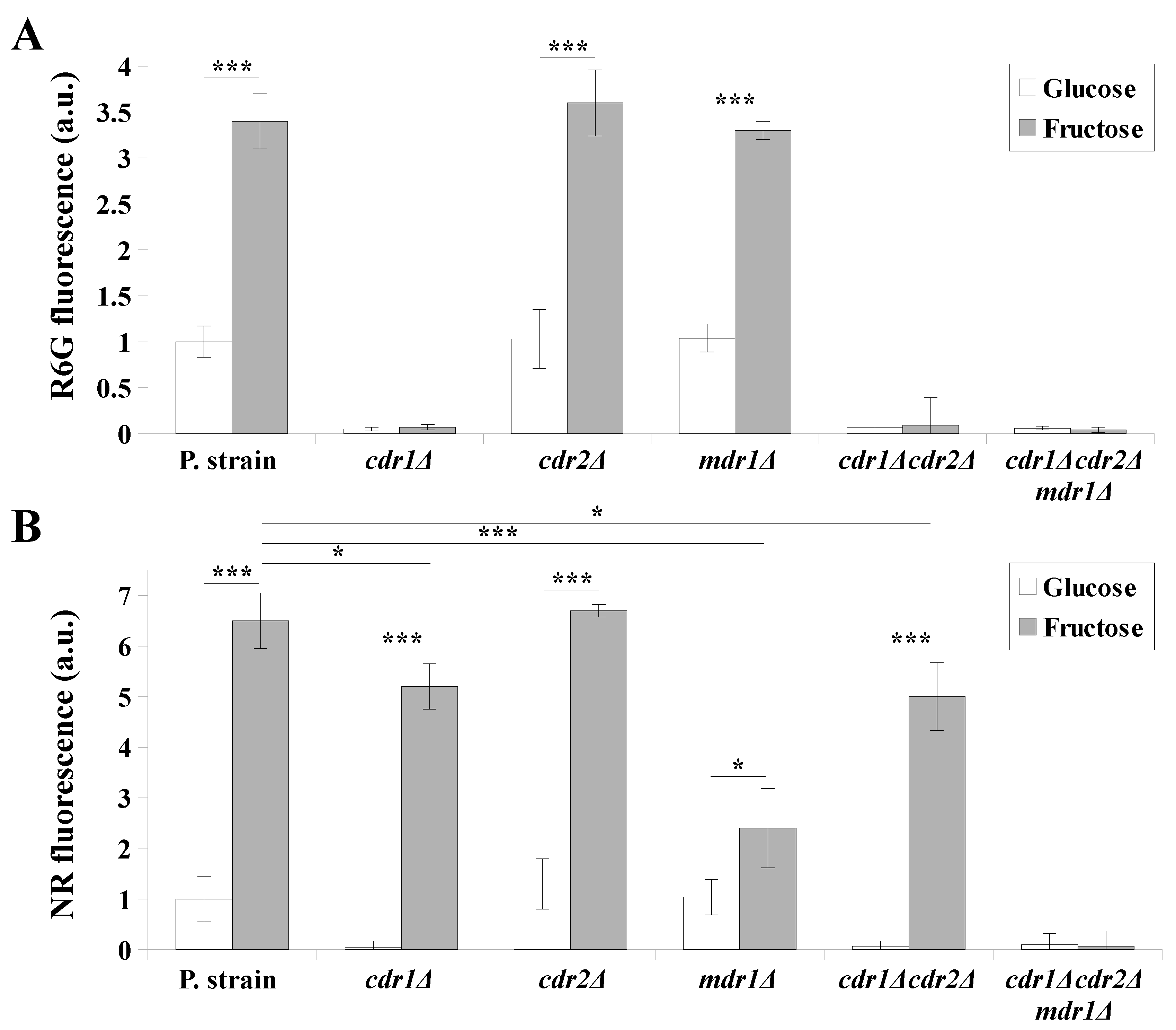
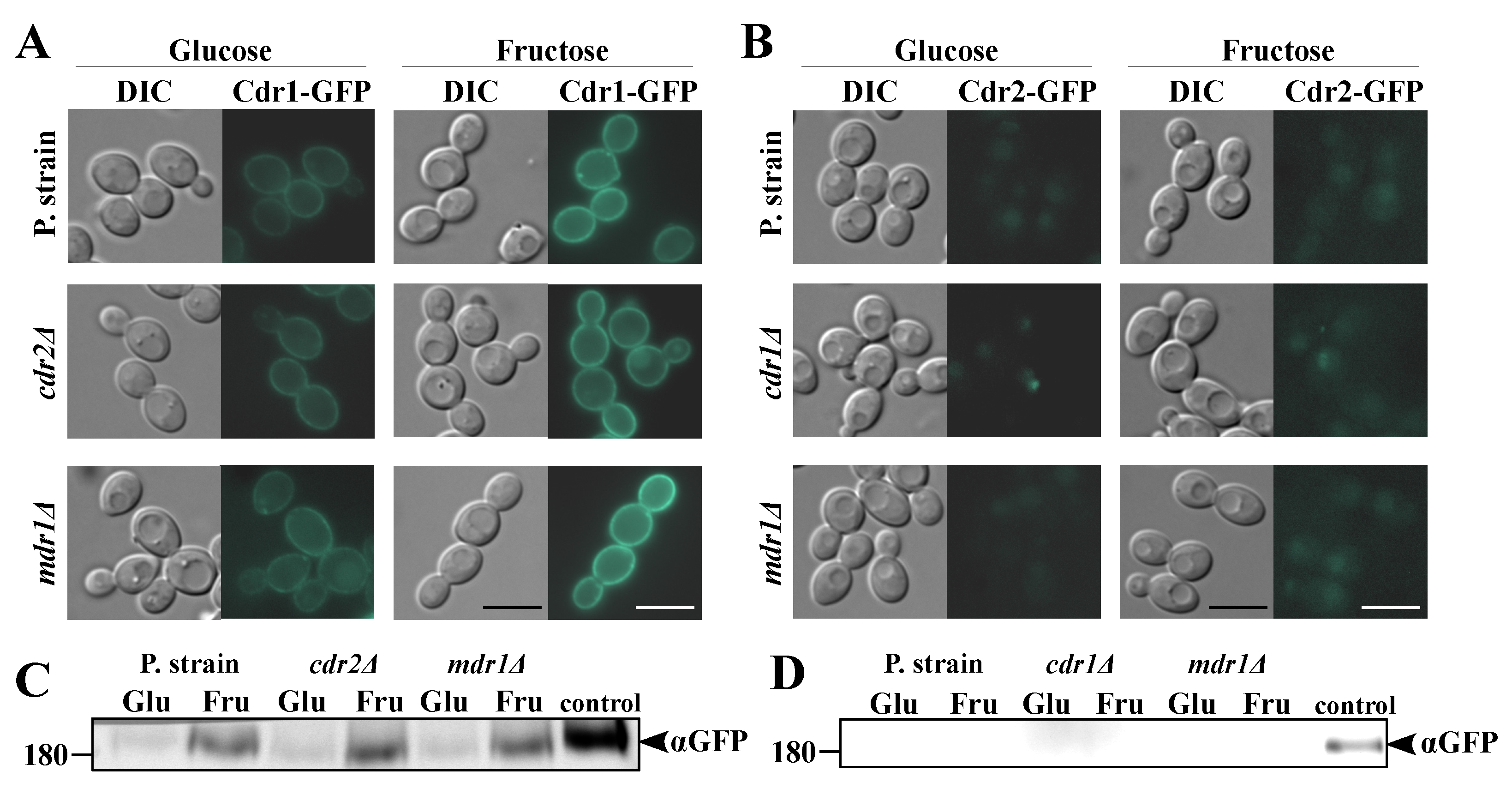
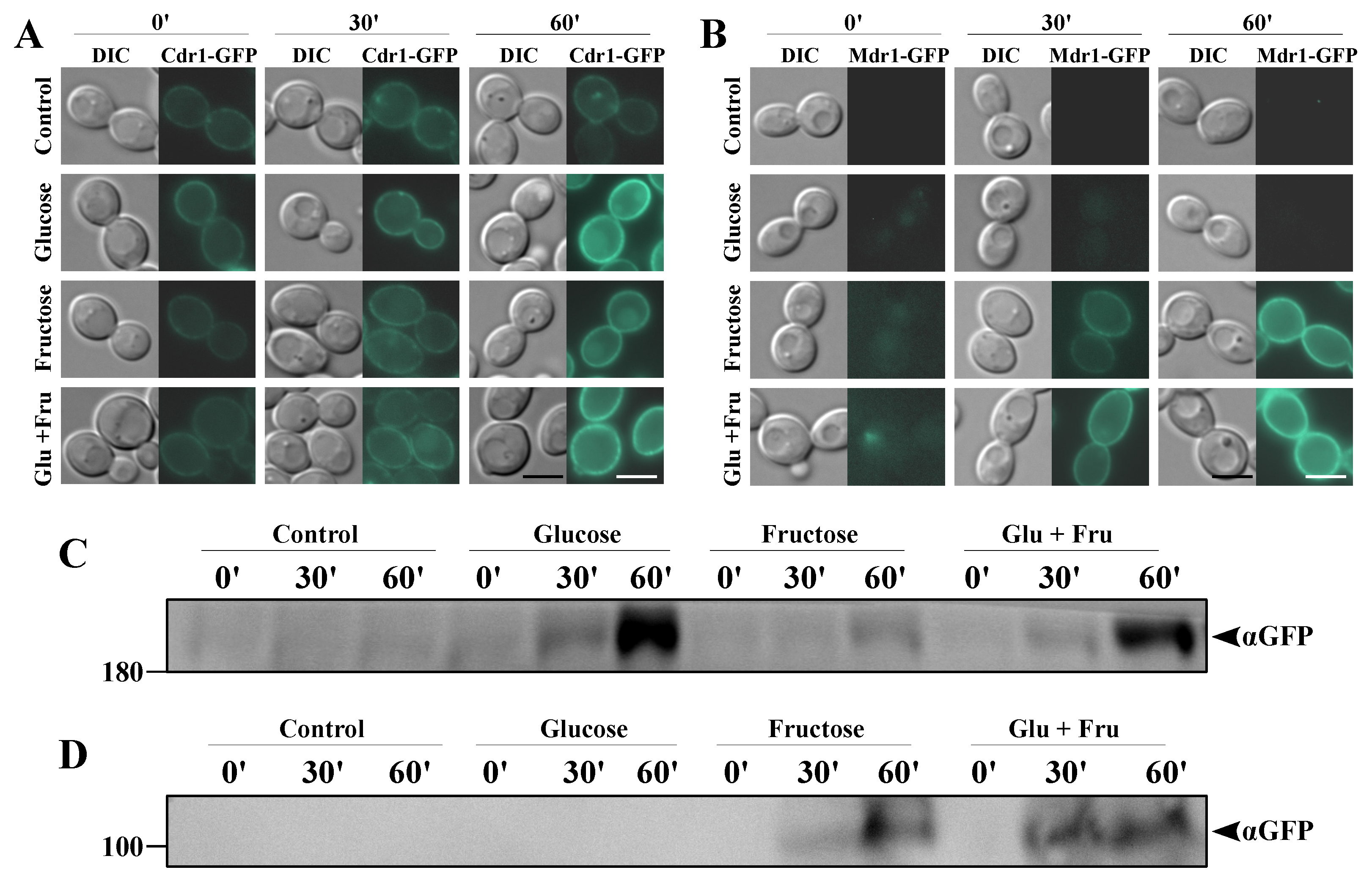
2.2. Fructose-Grown Cells Are Characterized by High Levels of Cdr1p and Mdr1p
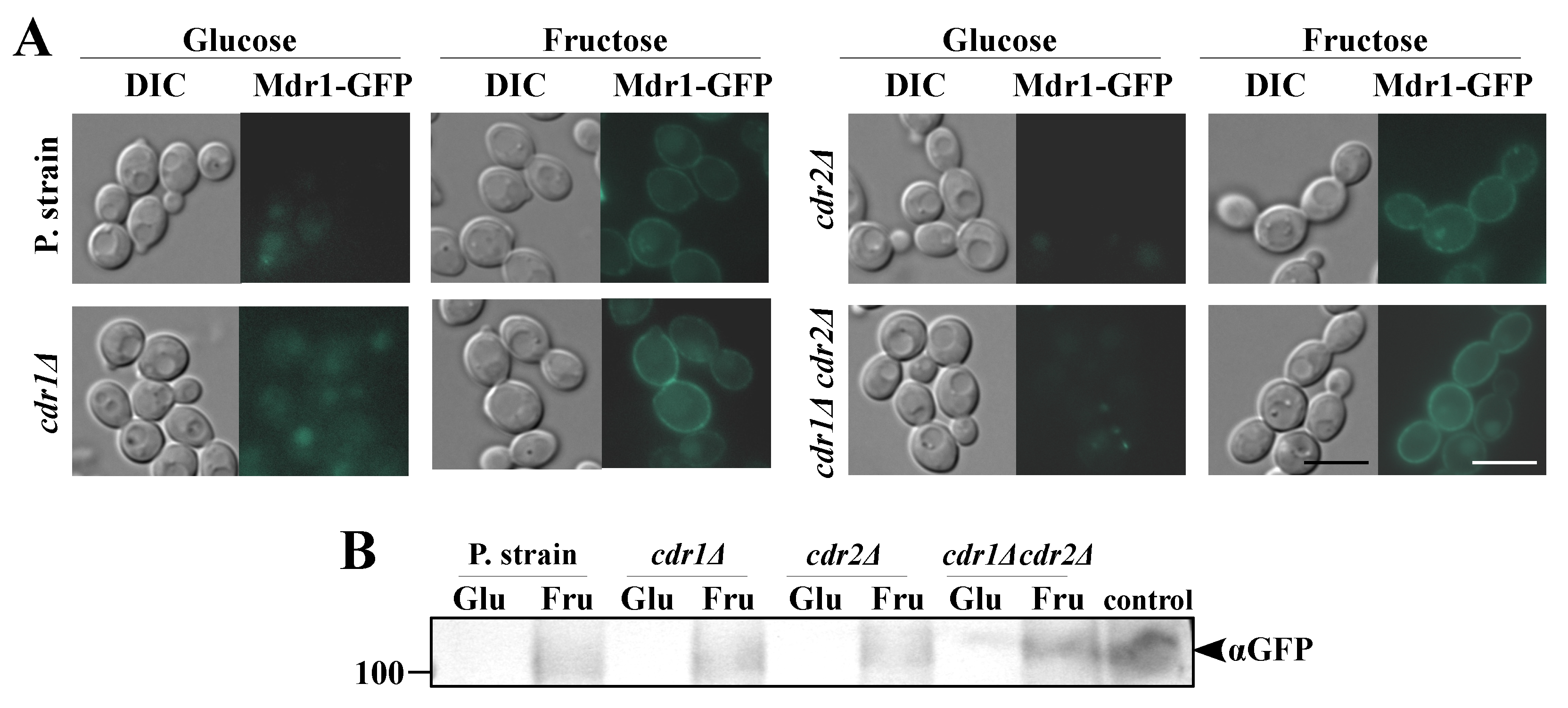
2.3. Serum Levels of Fructose Induces de Novo Synthesis of Mdr1p and Enhanced Synthesis of Cdr1p
3. Conclusions
4. Materials and Methods
4.1. Chemicals
4.2. Strains and Growth Conditions
4.3. Plasmids and Strains Construction
4.4. Percentage of Growth
4.5. Phenotypic Tests
4.6. Efflux Activity of MDR Transporters
4.7. Microscopic Studies
4.8. Western Blotting
4.9. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dadar, M.; Tiwari, R.; Karthik, K.; Chakraborty, S.; Shahali, Y.; Dhama, K. Candida albicans - biology, molecular characterization, pathogenicity, and advances in diagnosis and control – An Update. Microb. Pathog. 2018, 117, 128–138. [Google Scholar] [CrossRef]
- Sanglard, D.; Francoise, I.; Parkinson, T.; Falconer, D.; Jacques, B. Candida albicans Mutations in the Ergosterol Biosynthetic Pathway and Resistance to Several Antifungal Agents. Antimicrob. Agents Chemother. 2003, 47, 2404–2412. [Google Scholar] [CrossRef] [Green Version]
- Beardsley, J.; Halliday, C.L.; Chen, S.C.A.; Sorrell, T.C. Responding to the emergence of antifungal drug resistance: Perspectives from the bench and the bedside. Future Microbiol. 2018, 13, 1175–1191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cortegiani, A.; Misseri, G.; Chowdhary, A. What’s new on emerging resistant Candida species. Intensive Care Med. 2019, 45, 512–515. [Google Scholar] [CrossRef] [PubMed]
- Cannon, R.D.; Lamping, E.; Holmes, A.R.; Niimi, K.; Baret, P.V.; Keniya, M.V.; Tanabe, K.; Niimi, M.; Goffeau, A.; Monk, B.C. Efflux-mediated antifungal drug resistance. Clin. Microbiol. Rev. 2009, 22, 291–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Leon, E.M.; Jacober, S.J.; Sobel, J.D.; Foxman, B. Prevalence and risk factors for vaginal Candida colonization in women with type 1 and type 2 diabetes. BMC Infect. Dis. 2002, 2, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Javed, F.; Ahmed, H.B.; Mehmood, A.; Saeed, A.; Al-Hezaimi, K.; Samaranayake, L.P. Association between glycemic status and oral Candida carriage in patients with prediabetes. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. 2014, 117, 53–58. [Google Scholar] [CrossRef]
- Martinez, R.F.F.; Hernández-Pérez, F.; Miguel, G.F.S.; Jaimes-Aveldañez, A.; Arenas, R. Oral Candida spp carriers: Its prevalence in patients with type 2 Diabetes Mellitus. An. Bras. Dermatol. 2013, 88, 222–225. [Google Scholar] [CrossRef]
- Plotkin, B.J.; Paulson, D.; Chelich, A.; Jurak, D.; Cole, J.; Kasimos, J.; Burdick, J.R.; Casteel, N. Immune responsiveness in a rat model for type II diabetes (Zucker rat, fa/fa): Susceptibility to Candida albicans infection and leucocyte function. J. Med. Microbiol. 1996, 44, 277–283. [Google Scholar] [CrossRef] [Green Version]
- Rodaki, A.; Bohovych, I.M.; Enjalbert, B.; Young, T.; Odds, F.C.; Gow, N.A.R.; Brown, A.J.P. Glucose Promotes Stress Resistance in the Fungal Pathogen Candida albicans. Mol. Biol. Cell 2009, 20, 4845–4855. [Google Scholar] [CrossRef] [Green Version]
- Mandal, S.M.; Mahata, D.; Migliolo, L.; Parekh, A.; Addy, P.S.; Mandal, M.; Basak, A. Glucose directly promotes antifungal resistance in the fungal pathogen, Candida spp. J. Biol. Chem. 2014, 289, 25468–25473. [Google Scholar] [CrossRef] [Green Version]
- Ene, I.V.; Adya, A.K.; Wehmeier, S.; Brand, A.C.; Maccallum, D.M.; Gow, N.A.R.; Brown, A.J.P. Host carbon sources modulate cell wall architecture, drug resistance and virulence in a fungal pathogen. Cell. Microbiol. 2012, 14, 1319–1335. [Google Scholar] [CrossRef] [Green Version]
- Szczepaniak, J.; Łukaszewicz, M.; Krasowska, A. Estimation of Candida albicans ABC transporter behavior in real-time via fluorescence. Front. Microbiol. 2015, 6, E1382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawasaki, T.; Akanuma, H.; Yamanouchi, T. Increased fructose concentrations in blood and urine in patients with diabetes. Diabetes Care 2002, 25, 353–357. [Google Scholar] [CrossRef] [Green Version]
- Kawasaki, T.; Igarashi, K.; Ogata, N.; Oka, Y.; Ichiyanagi, K.; Yamanouchi, T. Markedly increased serum and urinary fructose concentrations in diabetic patients with ketoacidosis or ketosis. Acta Diabetol. 2012, 49, 119–123. [Google Scholar] [CrossRef]
- Hui, H.; Huang, D.; McArthur, D.; Nissen, N.; Boros, L.G.; Heaney, A.P. Direct spectrophotometric determination of serum fructose in pancreatic cancer patients. Pancreas 2009, 38, 706–712. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.J.; Perez-Pozo, S.E.; Sautin, Y.Y.; Manitius, J.; Sanchez-Lozada, L.G.; Feig, D.I.; Shafiu, M.; Segal, M.; Glassock, R.J.; Shimada, M.; et al. Hypothesis: Could excessive fructose intake and uric acid cause type 2 diabetes? Endocr. Rev. 2009, 30, 96–116. [Google Scholar] [CrossRef] [Green Version]
- Abu-Elteen, K.H. The influence of dietary carbohydrates on in vitro adherence of four Candida species to human buccal epithelial cells. Microb. Ecol. Health Dis. 2005, 17, 156–162. [Google Scholar] [CrossRef] [Green Version]
- Van Ende, M.; Wijnants, S.; Van Dijck, P. Sugar sensing and signaling in Candida albicans and Candida glabrata. Front. Microbiol. 2019, 10, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Sandai, D.; Yin, Z.; Selway, L.; Stead, D.; Walker, J.; Leach, M.D.; Bohovych, I.; Ene, I.V.; Kastora, S.; Budge, S.; et al. The Evolutionary Rewiring of Ubiquitination Targets Has Reprogrammed the Regulation of Carbon Assimilation in the Pathogenic Yeast Candida albicans. MBio 2012, 3, e00495-12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleck, C.B.; Schöbel, F.; Brock, M. Nutrient acquisition by pathogenic fungi: Nutrient availability, pathway regulation, and differences in substrate utilization. Int. J. Med. Microbiol. 2011, 301, 400–407. [Google Scholar] [CrossRef]
- Chew, S.Y.; Ho, K.L.; Cheah, Y.K.; Sandai, D.; Brown, A.J.P.; Lung Than, L.T. Physiologically relevant alternative carbon sources modulate biofilm formation, cell wall architecture, and the stress and antifungal resistance of Candida glabrata. Int. J. Mol. Sci. 2019, 20, 3172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, S.; Moye-Rowley, W.S. Multidrug resistance in fungi: Regulation of transporter-encoding gene expression. Front. Physiol. 2014, 5, 143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wirsching, S.; Michel, S.; Morschhäuser, J. Targeted gene disruption in Candida albicans wild-type strains: The role of the MDR1 gene in fluconazole resistance of clinical Candida albicans isolates. Mol. Microbiol. 2000, 36, 856–865. [Google Scholar] [CrossRef] [Green Version]
- Sanglard, D.; Ischer, F.; Monod, M.; Bille, J. Cloning of Candida albicans genes conferring resistance to azole antifungal agents: Characterization of CDR2, a new multidrug ABC transporter gene. Microbiology 1997, 143, 405–416. [Google Scholar] [CrossRef] [Green Version]
- Sanglard, D.; Ischer, F.; Monod, M.; Bille, J. Susceptibilities of Candida albicans multidrug transporter mutants to various antifungal agents and other metabolic inhibitors. Antimicrob. Agents Chemother. 1996, 40, 2300–2305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukherjee, P.K.; Chandra, J.; Kuhn, D.M.; Ghannoum, M.A. Mechanism of fluconazole resistance in Candida albicans biofilms: Phase-specific role of efflux pumps and membrane sterols. Infect. Immun. 2003, 71, 4333–4340. [Google Scholar] [CrossRef] [Green Version]
- De Micheli, M.; Bille, J.; Schueller, C.; Sanglard, D. A common drug-responsive element mediates the upregulation of the Candida albicans ABC transporters CDR1 and CDR2, two genes involved in antifungal drug resistance. Mol. Microbiol. 2002, 43, 1197–1214. [Google Scholar] [CrossRef]
- Rognon, B.; Kozovska, Z.; Coste, A.T.; Pardini, G.; Sanglard, D. Identification of promoter elements responsible for the regulation of MDR1 from Candida albicans, a major facilitator transporter involved in azole resistance. Microbiology 2006, 152, 3701–3722. [Google Scholar] [CrossRef] [Green Version]
- Karababa, M.; Coste, A.T.; Rognon, B.; Bille, J.; Sanglard, D. Comparison of gene expression profiles of Candida albicans azole-resistant clinical isolates and laboratory strains exposed to drugs inducing multidrug transporters. Antimicrob. Agents Chemother. 2004, 48, 3064–3079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, W.; Yang, J.; Yang, L.; Li, Q.; Zhu, X.; Xi, Z.; Qiao, Z.; Cen, W. Research of Mrr1, Cap1 and MDR1 in Candida albicans resistant to azole medications. Exp. Ther. Med. 2018, 15, 1217–1224. [Google Scholar] [CrossRef] [Green Version]
- Ivnitski-Steele, I.; Holmes, A.R.; Lamping, E.; Monk, B.C.; D, R.; Sklar, L.A. Identification of Nile Red as a fluorescent substrate of the Candida albicans ABC transporters Cdr1p and Cdr2p and the MFS transporter Mdr1p. Anal. Biochem. 2009, 394, 87–91. [Google Scholar] [CrossRef] [Green Version]
- Kolaczkowska, A.; Kolaczkowski, M.; Goffeau, A.; Moye-Rowley, W.S. Compensatory activation of the multidrug transporters Pdr5p, Snq2p, and Yor1p by Pdr1p in Saccharomyces cerevisiae. FEBS Lett. 2008, 582, 977–983. [Google Scholar] [CrossRef] [Green Version]
- Martins, M.P.; Franceschini, A.C.C.; Jacob, T.R.; Rossi, A.; Martinez-Rossi, N.M. Compensatory expression of multidrug-resistance genes encoding ABC transporters in dermatophytes. J. Med. Microbiol. 2016, 65, 605–610. [Google Scholar] [CrossRef] [PubMed]
- Nishimoto, A.T.; Zhang, Q.; Hazlett, B.; Morschhäuser, J.; David Rogers, P. Contribution of clinically derived mutations in the gene encoding the zinc cluster transcription factor Mrr2 to fluconazole antifungal resistance and CDR1 expression in Candida albicans. Antimicrob. Agents Chemother. 2019, 63, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.J.; Liu, J.Y.; Shi, C.; Zhao, Y.; Meng, L.N.; Wu, F.; Xiang, M.J. FLO8 deletion leads to azole resistance by upregulating CDR1 and CDR2 in Candida albicans. Res. Microbiol. 2019, 170, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Vogel, M.; Hartmann, T.; Köberle, M.; Treiber, M.; Autenrieth, I.B.; Schumacher, U.K. Rifampicin induces MDR1 expression in Candida albicans. J. Antimicrob. Chemother. 2008, 61, 541–547. [Google Scholar] [CrossRef] [PubMed]
- Semchyshyn, H.M.; Lozinska, L.M. Fructose protects baker’s yeast against peroxide stress: Potential role of catalase and superoxide dismutase. FEMS Yeast Res. 2012, 12, 761–773. [Google Scholar] [CrossRef] [Green Version]
- Harry, J.B.; Oliver, B.G.; Song, J.L.; Silver, P.M.; Little, J.T.; Choiniere, J.; White, T.C. Drug-induced regulation of the MDR1 promoter in Candida albicans. Antimicrob. Agents Chemother. 2005, 49, 2785–2792. [Google Scholar] [CrossRef] [Green Version]
- Masterjohn, C.; Park, Y.; Lee, J.; Noh, S.K.; Koo, S.I.; Bruno, R.S. Dietary fructose feeding increases adipose methylglyoxal accumulation in rats in association with low expression and activity of glyoxalase-2. Nutrients 2013, 5, 3311–3328. [Google Scholar] [CrossRef] [Green Version]
- Biermann, A.R.; Demers, E.G.; Hogan, D.A. Mrr1 regulation of methylglyoxal catabolism and methylglyoxal-induced fluconazole resistance in Candida lusitaniae. Mol. Microbiol. 2020. [Google Scholar] [CrossRef]
- Redhu, A.K.; Shah, A.H.; Prasad, R. MFS transporters of Candida species and their role in clinical drug resistance. FEMS Yeast Res. 2016, 16, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Laughlin, M.R. Normal roles for dietary fructose in carbohydrate metabolism. Nutrients 2014, 6, 3117–3129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suchodolski, J.; Muraszko, J.; Bernat, P.; Krasowska, A. A Crucial Role for Ergosterol in Plasma Membrane Composition, Localisation, and Activity of Cdr1p and H + -ATPase in Candida albicans. Microorganisms 2019, 7, 378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fonzi, W.A.; Irwin, M.Y. Isogenic Strain Construction and Gene Mapping in Candida albicans. Genetis 1993, 134, 717–728. [Google Scholar] [CrossRef]
- Suchodolski, J.; Muraszko, J.; Korba, A.; Bernat, P.; Krasowska, A. Lipid composition and cell surface hydrophobicity of Candida albicans influence the efficacy of fluconazole-gentamicin treatment. Yeast 2020, 37, 117–129. [Google Scholar] [CrossRef]
- Milne, S.W.; Cheetham, J.; Lloyd, D.; Aves, S.; Bates, S. Cassettes for PCR-mediated gene tagging in Candida albicans utilizing nourseothricin resistance. Yeast 2011, 28, 833–841. [Google Scholar] [CrossRef]
- Sasse, C.; Schillig, R.; Dierolf, F.; Weyler, M.; Schneider, S.; Mogavero, S.; Rogers, P.D.; Morschhäuser, J. The transcription factor Ndt80 does not contribute to Mrr1-, Tac1-, and Upc2-mediated fluconazole resistance in Candida albicans. PLoS ONE 2011, 6, E25623. [Google Scholar] [CrossRef]
- CLSI Reference method for broth dilutionantifungal susceptibility testing of yeast. In Approved standard. M27-A3 28, 3rd ed.; CLSI: Annapolis Junction, MD, USA, 2008; p. 604. [CrossRef] [Green Version]
- Szczepaniak, J.; Cieślik, W.; Romanowicz, A.; Musioł, R.; Krasowska, A. Blocking and dislocation of Candida albicans Cdr1p transporter by styrylquinolines. Int. J. Antimicrob. Agents 2017, 50, 171–176. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Parent | Description | Complete Genotype | Reference |
|---|---|---|---|---|
| CAF2-1 | Parental strain | ura3∆::imm434/URA3 | [45] | |
| DSY448 | CAF2-1 | cdr1Δ | ura3Δ::imm434/ura3Δ::imm434 cdr1Δ::hisG/cdr1Δ::hisG-URA3-hisG | [26] |
| DSY653 | CAF2-1 | cdr2Δ | ura3Δ::imm434/ura3Δ::imm434 cdr2Δ::hisG/cdr2Δ::hisG-URA3-hisG | [25] |
| DSY465 | CAF2-1 | mdr1Δ | ura3Δ::imm434/ura3Δ::imm434 mdr1Δ::hisG/mdr1Δ::hisG-URA3-hisG | [26] |
| DSY654 | CAF2-1 | cdr1Δcdr2Δ | ura3Δ::imm434/ura3Δ::imm434 cdr1Δ::hisG/cdr1Δ::hisG cdr2Δ::hisG/cdr2Δ::hisG-URA3-hisG | [25] |
| DSY1050 | CAF2-1 | cdr1Δcdr2Δmdr1Δ | ura3Δ::imm434/ura3Δ::imm434 cdr1Δ::hisG/cdr1Δ::hisG cdr2Δ::hisG/cdr2Δ:: hisG mdr1Δ::hisG/mdr1Δ::hisG-URA3-hisG | [27] |
| KS052 | CAF2-1 | CDR1-GFP | ura3Δ::imm434/URA3 CDR1/CDR1-GFP-NAT1 | [46] |
| KS053 | DSY653 | cdr2Δ CDR1-GFP | ura3Δ::imm434/ura3Δ::imm434 cdr2Δ::hisG/cdr2Δ::hisG-URA3-hisG CDR1/CDR1-GFP-NAT1 | This study |
| KS054 | DSY465 | mdr1Δ CDR1-GFP | ura3Δ::imm434/ura3Δ::imm434 mdr1Δ::hisG/mdr1Δ::hisG-URA3-hisG CDR1/CDR1-GFP-NAT1 | This study |
| KS063 | CAF2-1 | CDR2-GFP | ura3Δ::imm434/URA3 CDR2/CDR2-GFP-NAT1 | [46] |
| KS064 | DSY448 | cdr1Δ CDR2-GFP | ura3Δ::imm434/ura3Δ::imm434 cdr1Δ::hisG/cdr1Δ::hisG-URA3-hisG CDR2/CDR2-GFP-NAT1 | This study |
| KS065 | DSY465 | mdr1Δ CDR2-GFP | ura3Δ::imm434/ura3Δ::imm434 mdr1Δ::hisG/mdr1Δ::hisG-URA3-hisG CDR2/CDR2-GFP-NAT1 | This study |
| KS070 | CAF2-1 | MDR1-GFP | ura3Δ::imm434/URA3 MDR1/MDR1-GFP-NAT1 | This study |
| KS073 | DSY654 | cdr1Δcdr2ΔMDR1-GFP | ura3Δ::imm434/ura3Δ::imm434 cdr1Δ::hisG/cdr1Δ::hisG cdr2Δ::hisG/cdr2Δ::hisG-URA3-hisG MDR1/MDR1-GFP-NAT1 | This study |
| KS074 | DSY653 | cdr2ΔMDR1-GFP | ura3Δ::imm434/ura3Δ::imm434 cdr2Δ::hisG/cdr2Δ::hisG-URA3-hisG MDR1/MDR1-GFP-NAT1 | This study |
| KS075 | DSY448 | cdr1ΔMDR1-GFP | ura3Δ::imm434/ura3Δ::imm434 cdr1Δ::hisG/cdr1Δ::hisG-URA3-hisG MDR1/MDR1-GFP-NAT1 | This study |
| Primer | Sequence 5′–3′ |
|---|---|
| C1_GFPNAT_F | CATTCTTACGGTGATCTTTTATTGGTTAGCTAGAGTTCCAAAGGGTAACA GAGAGAAAAAAAATAAGAAAGGTGGTGGTTCTAAAGGTGAAGAATTATT |
| C1_GFPNAT_R | AACAACAACAATAGTCTAAAAACGTCTATTATATTTTAGACGTTTGAGA TACCACCATGTCAAAAAACAACGTTAGTATCGAATCG ACAGC |
| C2_GFPNAT_F | CATTCTTACTATTTTCTTTTACTGGTTGGCTAGAGTTCCAAAAGGTAATAG AGAAAAGAAGATGAAAAAAGGTGGTGGTTCTAAAG GTGAAGAATTATT |
| C2_GFPNAT_R | ATCAAACAATCACAAATAACGTATAAATAATAATAAGAAAAAAAAAAT ATGAATACTAATTGTAAAATAACGTTAGTATCGAATCG ACAGC |
| M1_GFPNAT_F | TTGTTATGATTGCTATTCCAGTTTTGTTTTACTTGAACGGACCAAAGTTGA GAGCAAGATCTAAGTATGCGGTGGTGGTTCTAAAGGTGAAGAATTATT |
| M1_GFPNAT_R | TCAGTCCTTTTCTCTTTTTAATTATTGATTAATGTATCTATAACACGATATA TCTATAGGAAAACAATGACGTTAGTATCGAATCGACAGC |
| NAT1_F | GCTTATAGATACAGAACTTCTGTTCC |
| NAT1_R | TGAAACCCATTCTTCTATAAGCATG |
| C1NAT1_SF | TCAAGCTATGCTTTCTACTGGA |
| C2NAT1_SF | GTATTGGCTGGTCCTAATGTG |
| M1NAT1_SF | TATTGGTATTGTCATTGCTGCC |
| GFP_N1_SR2 | AATTCTTCACCTTTAGAACCACC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suchodolski, J.; Krasowska, A. Fructose Induces Fluconazole Resistance in Candida albicans through Activation of Mdr1 and Cdr1 Transporters. Int. J. Mol. Sci. 2021, 22, 2127. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22042127
Suchodolski J, Krasowska A. Fructose Induces Fluconazole Resistance in Candida albicans through Activation of Mdr1 and Cdr1 Transporters. International Journal of Molecular Sciences. 2021; 22(4):2127. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22042127
Chicago/Turabian StyleSuchodolski, Jakub, and Anna Krasowska. 2021. "Fructose Induces Fluconazole Resistance in Candida albicans through Activation of Mdr1 and Cdr1 Transporters" International Journal of Molecular Sciences 22, no. 4: 2127. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22042127