
Murine Esophagus Expresses Glial-Derived Central Nervous System Antigens

 ,
, 
 ,
,  , and
, and 
Abstract
:
1. Introduction
2. Results
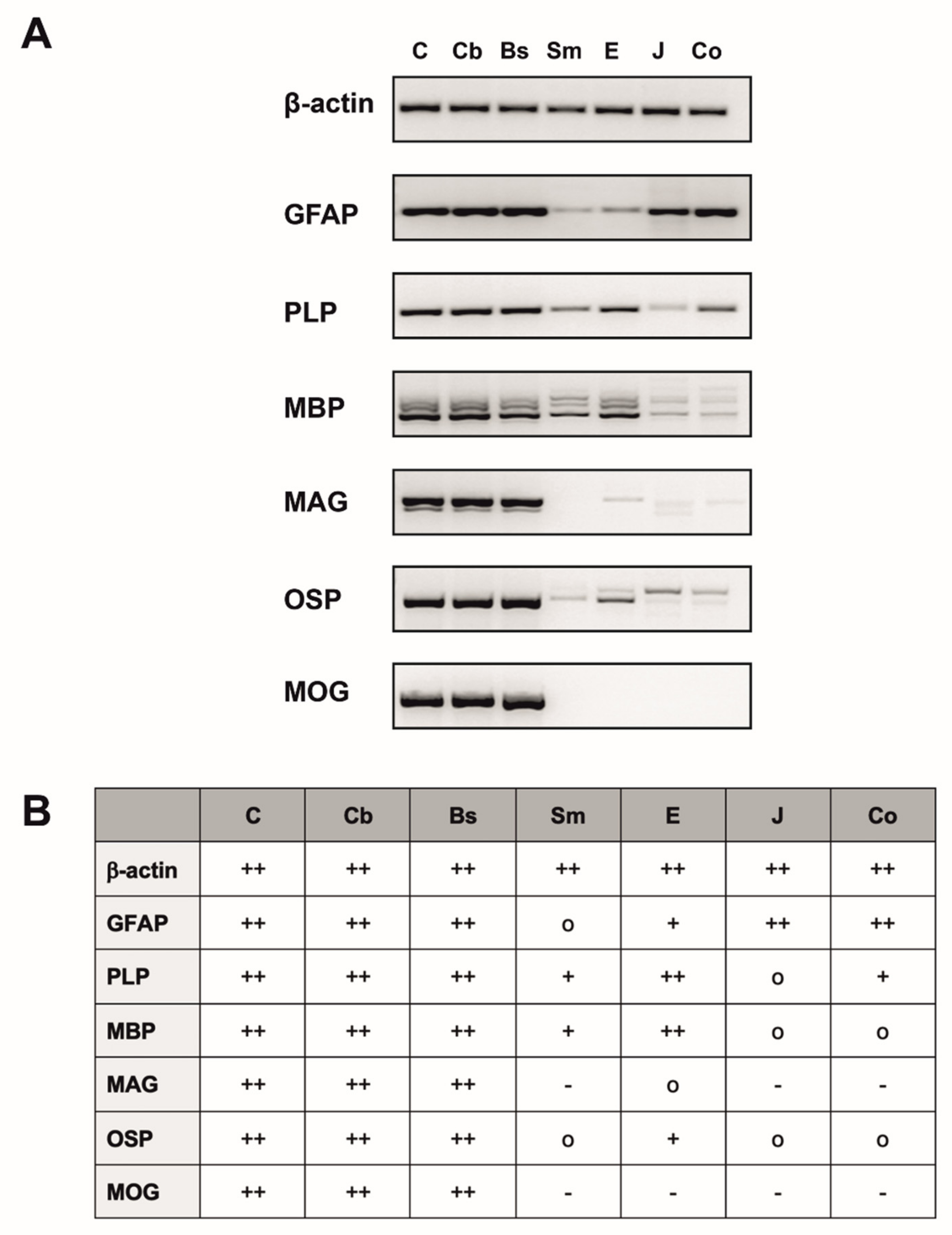
2.1. The Esophagus Expresses CNS Typical Markers at the mRNA-Level
2.2. Distribution of MBP, PLP and GFAP in the Cerebellum
2.3. Glial Cells in Neuromuscular Junctions of the Tibialis Anterior Muscle
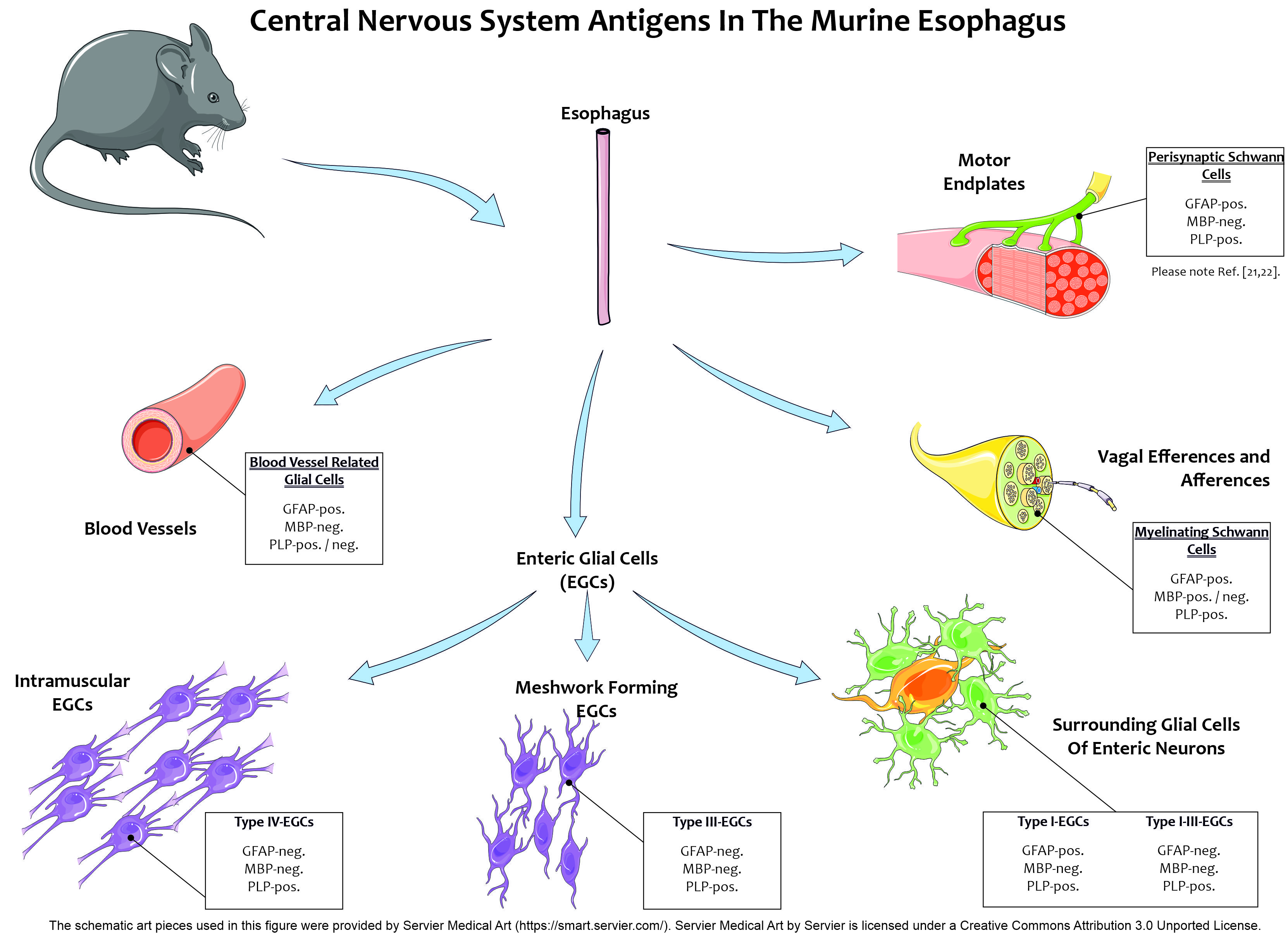
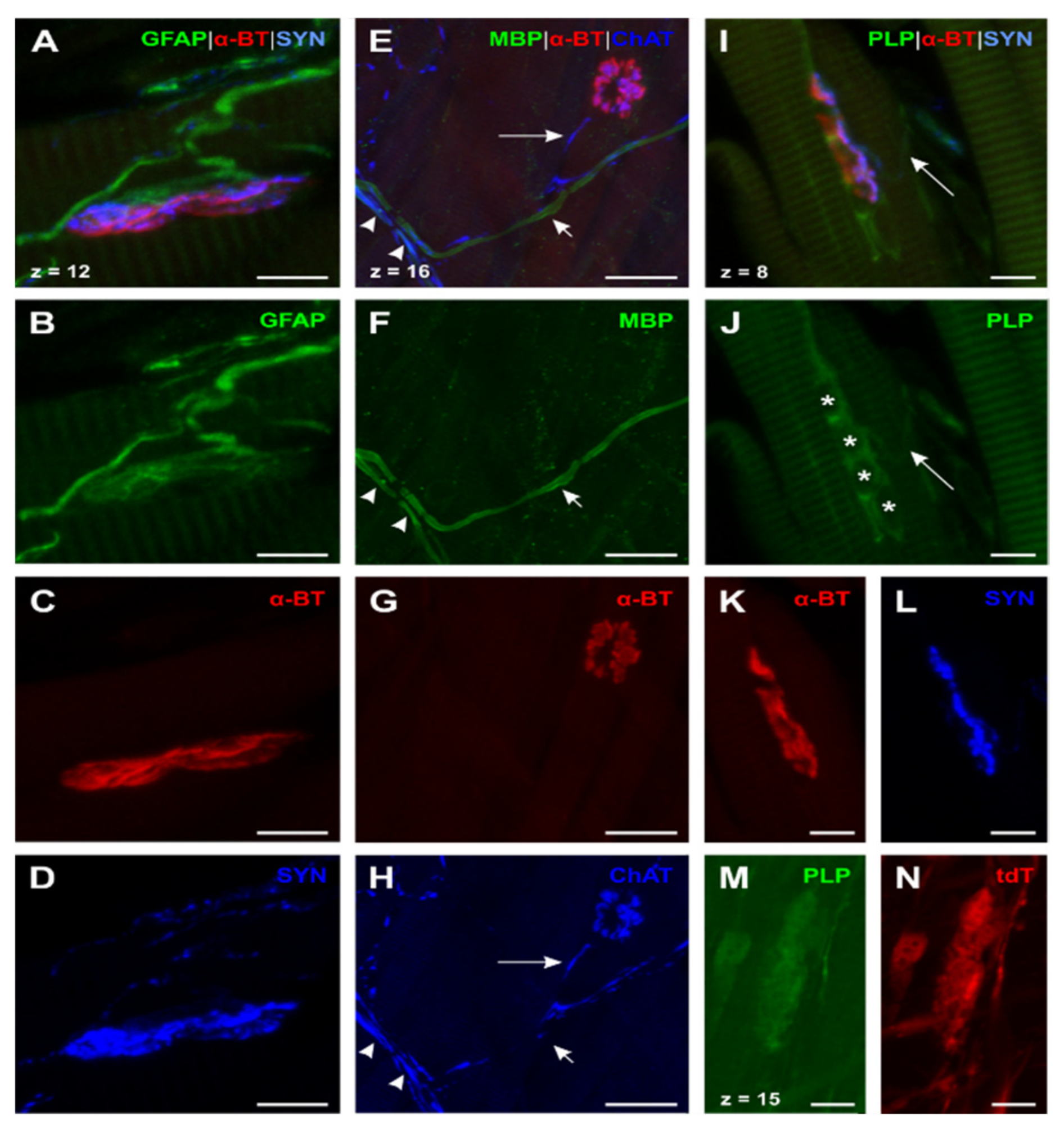
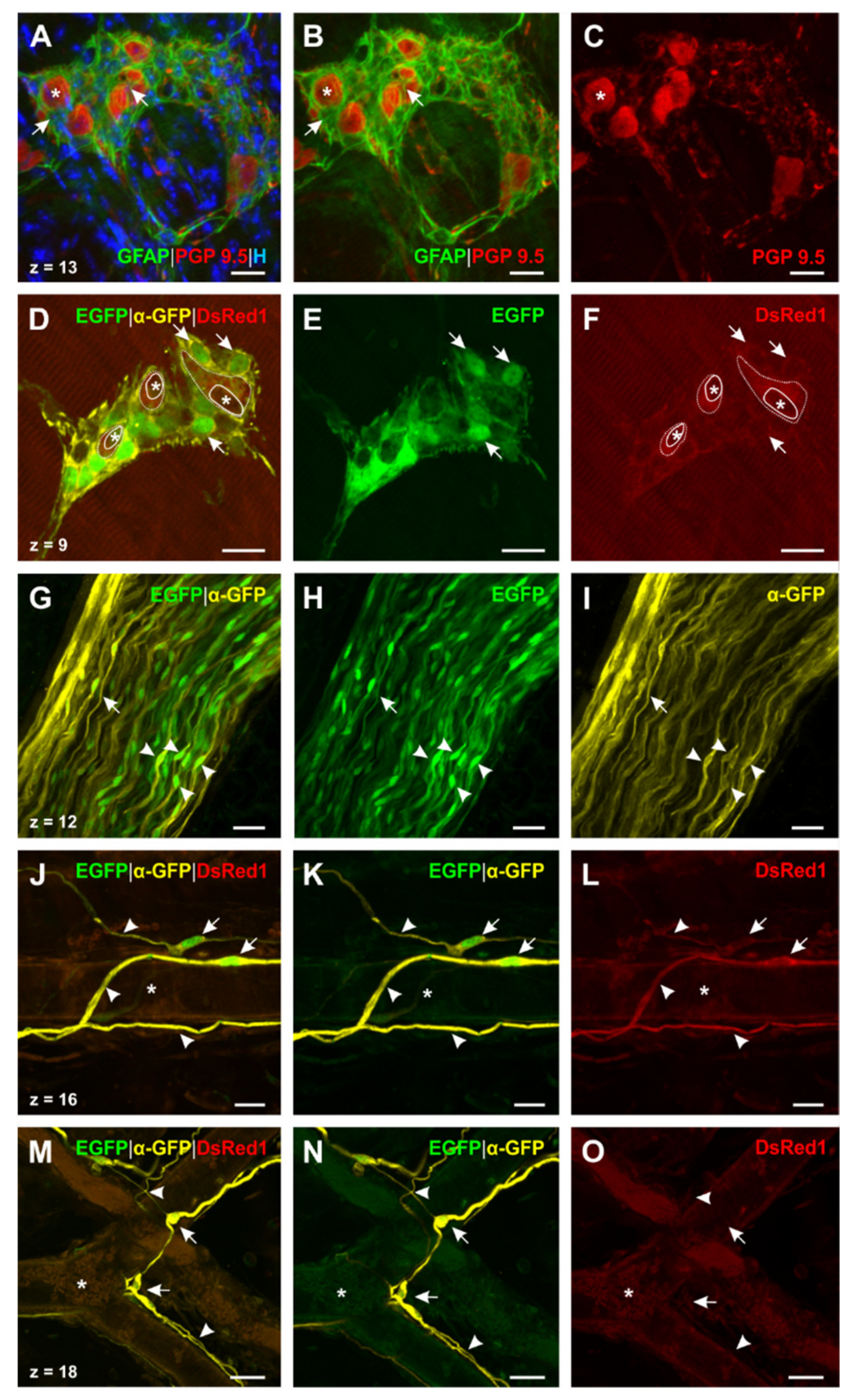
2.4. Glial Cells in Neuromuscular Junctions of the Esophagus
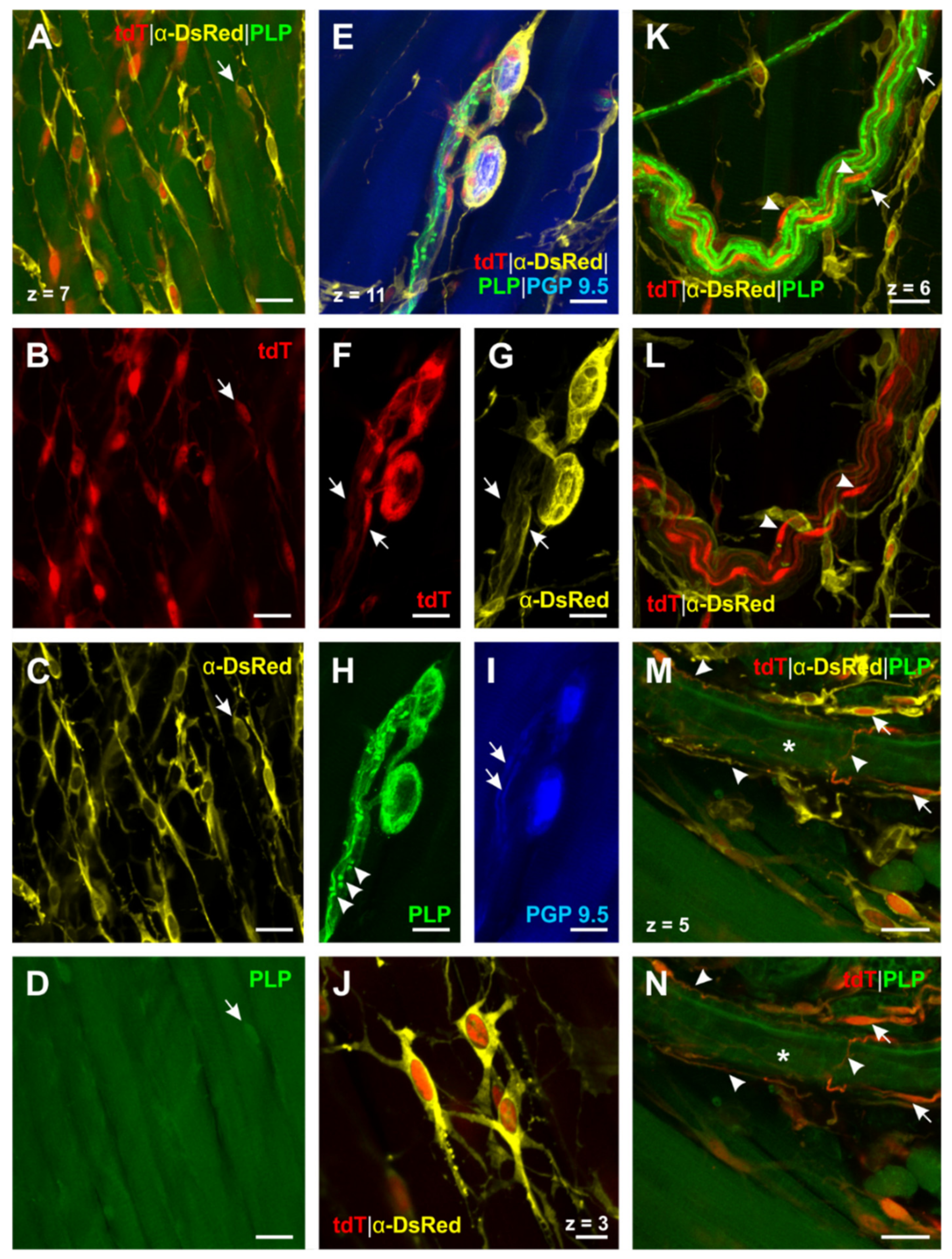
2.5. Transgenic Mice Reveal the Presence of Different Types of Enteric Glial Cells in the Esophagus
3. Discussion
3.1. Presence of Glial Cell Specific Markers in the Esophageal Neuromuscular Junction
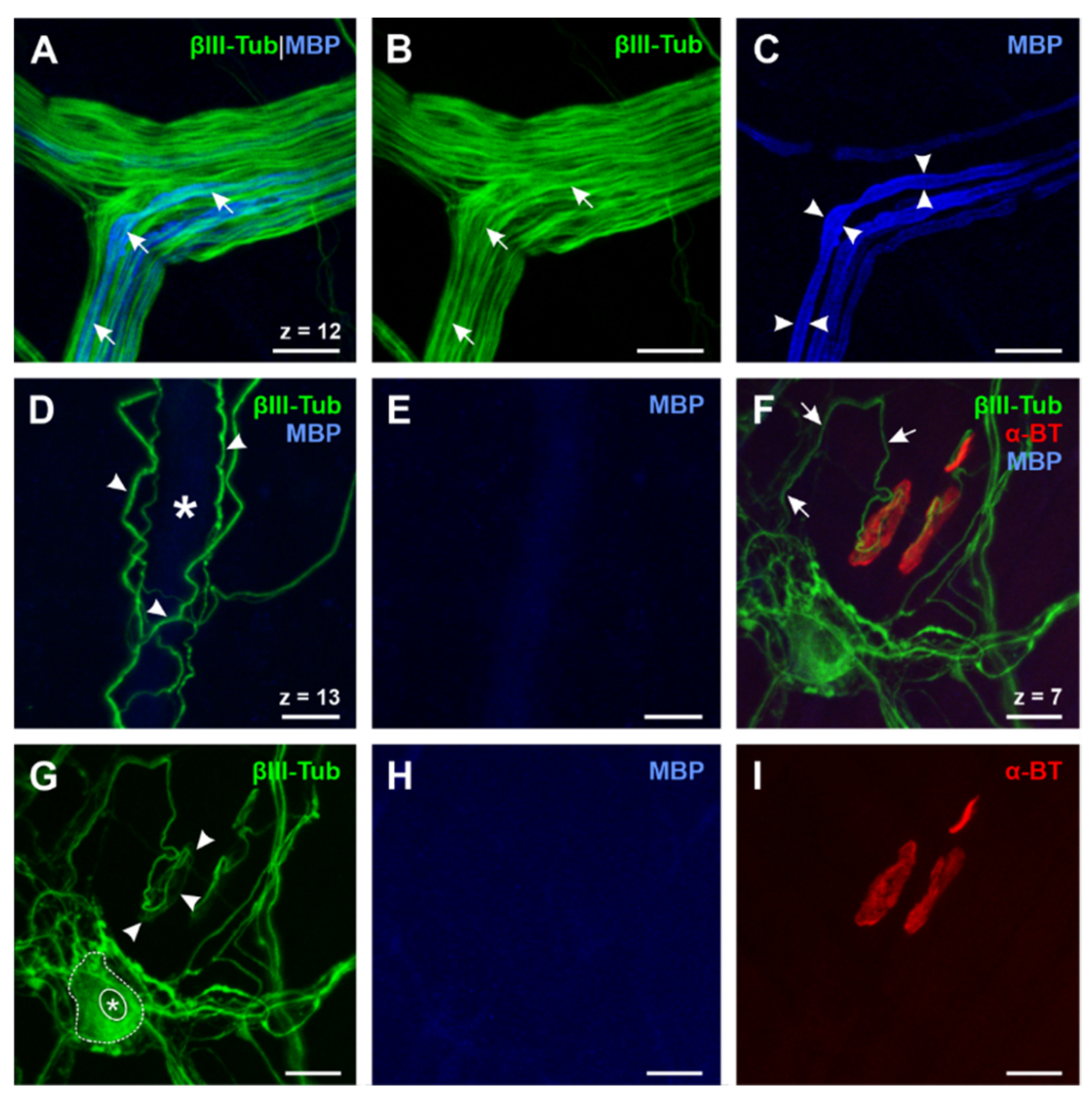
3.2. Different Degrees of Myelination in the Myenteric Plexus
3.3. Differentiation of Esophageal Enteric Glial Cells by the Use of Glial Markers
3.4. Enteric Glial Cells as an Immunological Target
4. Materials and Methods
4.1. Mice
4.2. RT-PCR
4.2.1. Tissue Preparation
4.2.2. Processing
4.3. Immunohistochemistry
4.3.1. Tissue Preparation and Fixation
4.3.2. Frozen Sections
4.3.3. Whole Mounts
4.3.4. Control Experiments
4.4. Image Aquisition
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BSA | Bovine serum albumin |
| CALB | Calbindin (D28k) |
| cDNA | Complementary DNA |
| ChAT | Choline Acetyltransferase |
| CNS | Central nervous system |
| DEPC | Diethyl pyrocarbonate |
| DsRed | Discosoma sp. red fluorescent protein |
| EAE | Experimental autoimmune encephalomyelitis |
| EDTA | Ethylenediaminetetraacetic acid |
| EGC | Enteric glial cell |
| EGFP | Enhanced green fluorescent protein |
| ENS | Enteric nervous system |
| GFAP | Glial fibrillary acidic protein |
| GFP | Green fluorescent protein |
| MAG | Myelin-associated glycoprotein (also: Siglec-4) |
| MBP | Myelin basic protein |
| MOG | Myelin oligodendrocyte glycoprotein |
| MS | Multiple sclerosis |
| NMJ | Neuromuscular junction |
| OSP | Oligodendrocyte-specific protein (also: Claudin-11) |
| PETZ | Präklinisches Experimentelles Tierzentrum; animal facility of the University Erlangen-Nürnberg |
| PGP 9.5 | Protein gene product 9.5 |
| PLP / PLP1 | Proteolipid protein 1 (also: lipohilin) |
| PNS | Peripheral nervous system |
| PSC | Perisynaptic Schwann cell |
| RT-PCR | Reverse transcriptase polymerase chain reaction |
| TAE | Tris/acetate/EDTA-buffer |
| TBS | Tris-buffered saline |
| tdT | tdTomato |
| α-BT | α-Bungarotoxin |
References
- Compston, A.; Coles, A. Multiple sclerosis. Lancet 2008, 372, 1502–1517. [Google Scholar] [CrossRef]
- Mahad, D.H.; Trapp, B.D.; Lassmann, H. Pathological mechanisms in progressive multiple sclerosis. Lancet Neurol. 2015, 14, 183–193. [Google Scholar] [CrossRef]
- Katz Sand, I. Classification, diagnosis, and differential diagnosis of multiple sclerosis. Curr. Opin. Neurol. 2015, 28, 193–205. [Google Scholar] [CrossRef] [PubMed]
- Weber, M.S.; Hemmer, B.; Cepok, S. The role of antibodies in multiple sclerosis. Biochim. Biophys. Acta 2011, 1812, 239–245. [Google Scholar] [CrossRef] [Green Version]
- Wunsch, M.; Jabari, S.; Voussen, B.; Enders, M.; Srinivasan, S.; Cossais, F.; Wedel, T.; Boettner, M.; Schwarz, A.; Weyer, L.; et al. The enteric nervous system is a potential autoimmune target in multiple sclerosis. Acta Neuropathol. 2017, 134, 281–295. [Google Scholar] [CrossRef]
- Spear, E.T.; Holt, E.A.; Joyce, E.J.; Haag, M.M.; Mawe, S.M.; Hennig, G.W.; Lavoie, B.; Applebee, A.M.; Teuscher, C.; Mawe, G.M. Altered gastrointestinal motility involving autoantibodies in the experimental autoimmune encephalomyelitis model of multiple sclerosis. Neurogastroenterol. Motil. 2018, 30, e13349. [Google Scholar] [CrossRef] [PubMed]
- Levinthal, D.J.; Rahman, A.; Nusrat, S.; O’Leary, M.; Heyman, R.; Bielefeldt, K. Adding to the burden: Gastrointestinal symptoms and syndromes in multiple sclerosis. Mult. Scler. Int. 2013, 2013, 319201. [Google Scholar] [CrossRef] [PubMed]
- Levinthal, D.J.; Bielefeldt, K. Systematic review and meta-analysis: Gastric electrical stimulation for gastroparesis. Auton. Neurosci. 2017, 202, 45–55. [Google Scholar] [CrossRef]
- Preziosi, G.; Raptis, D.A.; Raeburn, A.; Thiruppathy, K.; Panicker, J.; Emmanuel, A. Gut dysfunction in patients with multiple sclerosis and the role of spinal cord involvement in the disease. Eur. J. Gastroenterol. Hepatol. 2013, 25, 1044–1050. [Google Scholar] [CrossRef] [Green Version]
- Calcagno, P.; Ruoppolo, G.; Grasso, M.G.; de Vincentiis, M.; Paolucci, S. Dysphagia in multiple sclerosis - prevalence and prognostic factors. Acta Neurol. Scand. 2002, 105, 40–43. [Google Scholar] [CrossRef] [PubMed]
- Tassorelli, C.; Bergamaschi, R.; Buscone, S.; Bartolo, M.; Furnari, A.; Crivelli, P.; Alfonsi, E.; Alberici, E.; Bertino, G.; Sandrini, G.; et al. Dysphagia in multiple sclerosis: From pathogenesis to diagnosis. Neurol. Sci. 2008, 29 (Suppl. 4), S360–S363. [Google Scholar] [CrossRef] [PubMed]
- De Pauw, A.; Dejaeger, E.; D’hooghe, B.; Carton, H. Dysphagia in multiple sclerosis. Clin. Neurol. Neurosurg. 2002, 104, 345–351. [Google Scholar] [CrossRef]
- Alali, D.; Ballard, K.; Vucic, S.; Bogaardt, H. Dysphagia in Multiple Sclerosis: Evaluation and Validation of the DYMUS Questionnaire. Dysphagia 2018, 33, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Barkhof, F.; Koeller, K.K. Demyelinating Diseases of the CNS (Brain and Spine). In Diseases of the Brain, Head and Neck, Spine 2020-2023: Diagnostic Imaging, 1st ed.; Hodler, J., Kubik-Huch, R.A., von Schulthess, G.K., Eds.; Springer International Publishing: Cham, Germany, 2020; pp. 165–176. ISBN 978-3-030-38490-6. [Google Scholar]
- Love, S. Demyelinating diseases. J. Clin. Pathol. 2006, 59, 1151–1159. [Google Scholar] [CrossRef] [PubMed]
- Alali, D.; Ballard, K.; Bogaardt, H. Treatment Effects for Dysphagia in Adults with Multiple Sclerosis: A Systematic Review. Dysphagia 2016, 31, 610–618. [Google Scholar] [CrossRef]
- Kim, J.-S.; Sung, H.-Y. Gastrointestinal Autonomic Dysfunction in Patients with Parkinson’s Disease. J. Mov. Disord. 2015, 8, 76–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verne, G.N.; Sallustio, J.E.; Eaker, E.Y. Anti-myenteric neuronal antibodies in patients with achalasia. A prospective study. Dig. Dis. Sci. 1997, 42, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Neuhuber, W.L.; Wörl, J.; Berthoud, H.R.; Conte, B. NADPH-diaphorase-positive nerve fibers associated with motor endplates in the rat esophagus: New evidence for co-innervation of striated muscle by enteric neurons. Cell Tissue Res. 1994, 276, 23–30. [Google Scholar] [CrossRef]
- Wörl, J.; Mayer, B.; Neuhuber, W.L. Nitrergic innervation of the rat esophagus: Focus on motor endplates. J. Auton. Nerv. Syst. 1994, 49, 227–233. [Google Scholar] [CrossRef]
- Wörl, J.; Neuhuber, W.L. Enteric co-innervation of motor endplates in the esophagus: State of the art ten years after. Histochem. Cell Biol. 2005, 123, 117–130. [Google Scholar] [CrossRef]
- Neuhuber, W.L.; Wörl, J. Enteric co-innervation of striated muscle in the esophagus: Still enigmatic? Histochem. Cell Biol. 2016, 146, 721–735. [Google Scholar] [CrossRef] [PubMed]
- Kuramoto, H.; Yoshimura, R.; Sakamoto, H.; Kadowaki, M. Regional variations in the number distribution of intrinsic myenteric neurons and coinnervated motor endplates on the striated muscles in the rat esophagus. Auton. Neurosci. 2019, 219, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Furness, J.B. The enteric nervous system and neurogastroenterology. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 286–294. [Google Scholar] [CrossRef] [PubMed]
- Chadi, G.; Gomide, V.C.; Rodrigues de Souza, R.; Scabello, R.T.; Maurício da Silva, C. Basic fibroblast growth factor, neurofilament, and glial fibrillary acidic protein immunoreactivities in the myenteric plexus of the rat esophagus and colon. J. Morphol. 2004, 261, 323–333. [Google Scholar] [CrossRef]
- Raab, M.; Neuhuber, W.L. Intraganglionic laminar endings and their relationships with neuronal and glial structures of myenteric ganglia in the esophagus of rat and mouse. Histochem. Cell Biol. 2004, 122, 445–459. [Google Scholar] [CrossRef]
- Kuerten, S.; Lichtenegger, F.S.; Faas, S.; Angelov, D.N.; Tary-Lehmann, M.; Lehmann, P.V. MBP-PLP fusion protein-induced EAE in C57BL/6 mice. J. Neuroimmunol. 2006, 177, 99–111. [Google Scholar] [CrossRef]
- Robinson, A.P.; Harp, C.T.; Noronha, A.; Miller, S.D. The experimental autoimmune encephalomyelitis (EAE) model of MS: Utility for understanding disease pathophysiology and treatment. Handb. Clin. Neurol. 2014, 122, 173–189. [Google Scholar] [CrossRef] [Green Version]
- Mørkholt, A.S.; Kastaniegaard, K.; Trabjerg, M.S.; Gopalasingam, G.; Niganze, W.; Larsen, A.; Stensballe, A.; Nielsen, S.; Nieland, J.D. Identification of brain antigens recognized by autoantibodies in experimental autoimmune encephalomyelitis-induced animals treated with etomoxir or interferon-β. Sci. Rep. 2018, 8, 7092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, M.; Nelms, B.D.; Dong, L.; Salinas-Rios, V.; Rutlin, M.; Gershon, M.D.; Corfas, G. Enteric glia express proteolipid protein 1 and are a transcriptionally unique population of glia in the mammalian nervous system. Glia 2015, 63, 2040–2057. [Google Scholar] [CrossRef]
- Grundmann, D.; Loris, E.; Maas-Omlor, S.; Huang, W.; Scheller, A.; Kirchhoff, F.; Schäfer, K.-H. Enteric Glia: S100, GFAP, and Beyond. Anat. Rec. 2019, 302, 1333–1344. [Google Scholar] [CrossRef] [PubMed]
- Mayer, M.C.; Meinl, E. Glycoproteins as targets of autoantibodies in CNS inflammation: MOG and more. Ther. Adv. Neurol. Disord. 2012, 5, 147–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quarles, R.H. Myelin-associated glycoprotein (MAG): Past, present and beyond. J. Neurochem. 2007, 100, 1431–1448. [Google Scholar] [CrossRef]
- Pascual-Goñi, E.; Martín-Aguilar, L.; Lleixà, C.; Martínez-Martínez, L.; Simón-Talero, M.J.; Díaz-Manera, J.; Cortés-Vicente, E.; Rojas-García, R.; Moga, E.; Juárez, C.; et al. Clinical and laboratory features of anti-MAG neuropathy without monoclonal gammopathy. Sci. Rep. 2019, 9, 6155. [Google Scholar] [CrossRef]
- Bronstein, J.M.; Micevych, P.E.; Chen, K. Oligodendrocyte-specific protein (OSP) is a major component of CNS myelin. J. Neurosci. Res. 1997, 50, 713–720. [Google Scholar] [CrossRef]
- Yu, Y.; Fu, Y.; Watson, C. The inferior olive of the C57BL/6J mouse: A chemoarchitectonic study. Anat. Rec. 2014, 297, 289–300. [Google Scholar] [CrossRef]
- Grosche, J.; Matyash, V.; Möller, T.; Verkhratsky, A.; Reichenbach, A.; Kettenmann, H. Microdomains for neuron-glia interaction: Parallel fiber signaling to Bergmann glial cells. Nat. Neurosci. 1999, 2, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Triolo, D.; Dina, G.; Lorenzetti, I.; Malaguti, M.; Morana, P.; Del Carro, U.; Comi, G.; Messing, A.; Quattrini, A.; Previtali, S.C. Loss of glial fibrillary acidic protein (GFAP) impairs Schwann cell proliferation and delays nerve regeneration after damage. J. Cell Sci. 2006, 119, 3981–3993. [Google Scholar] [CrossRef] [Green Version]
- Ko, C.-P.; Sugiura, Y.; Feng, Z. The biology of perisynaptic (terminal) Schwann cells. In The Biology of Schwann Cells; Armati, P., Ed.; Cambridge University Press: Cambridge, UK, 2007; pp. 72–99. ISBN 9780511541605. [Google Scholar]
- Barik, A.; Li, L.; Sathyamurthy, A.; Xiong, W.-C.; Mei, L. Schwann Cells in Neuromuscular Junction Formation and Maintenance. J. Neurosci. 2016, 36, 9770–9781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armati, P.J.; Mathey, E.K. An update on Schwann cell biology--immunomodulation, neural regulation and other surprises. J. Neurol. Sci. 2013, 333, 68–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugiura, Y.; Lin, W. Neuron-glia interactions: The roles of Schwann cells in neuromuscular synapse formation and function. Biosci. Rep. 2011, 31, 295–302. [Google Scholar] [CrossRef] [Green Version]
- Feng, Z.; Ko, C.-P. Schwann cells promote synaptogenesis at the neuromuscular junction via transforming growth factor-beta1. J. Neurosci. 2008, 28, 9599–9609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Auld, D.S.; Robitaille, R. Perisynaptic Schwann cells at the neuromuscular junction: Nerve- and activity-dependent contributions to synaptic efficacy, plasticity, and reinnervation. Neuroscientist 2003, 9, 144–157. [Google Scholar] [CrossRef]
- Gruber, H. Uber Struktur und Innervation der quergestreiften Muskulatur des Oesophagus der Ratte. Z. Zellforsch. Mikrosk. Anat. 1968, 91, 236–247. [Google Scholar] [CrossRef]
- Samarasinghe, D.D. Some observations on the innervation of the striated muscle in the mouse oesophagus–an electron microscopy study. J. Anat. 1972, 112, 173–184. [Google Scholar] [PubMed]
- Berthoud, H.R.; Patterson, L.M.; Neumann, F.; Neuhuber, W.L. Distribution and structure of vagal afferent intraganglionic laminar endings (IGLEs) in the rat gastrointestinal tract. Anat. Embryol. 1997, 195, 183–191. [Google Scholar] [CrossRef]
- Powley, T.L.; Baronowsky, E.A.; Gilbert, J.M.; Hudson, C.N.; Martin, F.N.; Mason, J.K.; McAdams, J.L.; Phillips, R.J. Vagal afferent innervation of the lower esophageal sphincter. Auton. Neurosci. 2013, 177, 129–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neuhuber, W.L. Sensory vagal innervation of the rat esophagus and cardia: A light and electron microscopic anterograde tracing study. J. Auton. Nerv. Syst. 1987, 20, 243–255. [Google Scholar] [CrossRef]
- Sengupta, J.N. Esophageal sensory physiology. GI Motil. Online 2006. [Google Scholar] [CrossRef]
- Clouse, R.E.; Richter, J.E.; Heading, R.C.; Janssens, J.; Wilson, J.A. Functional esophageal disorders. Gut 1999, 45 (Suppl. 2), II31–II36. [Google Scholar] [CrossRef] [Green Version]
- Richter, J.E.; Barish, C.F.; Castell, D.O. Abnormal sensory perception in patients with esophageal chest pain. Gastroenterology 1986, 91, 845–852. [Google Scholar] [CrossRef]
- Hanani, M.; Reichenbach, A. Morphology of horseradish peroxidase (HRP)-injected glial cells in the myenteric plexus of the guinea-pig. Cell Tissue Res. 1994, 278, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Gulbransen, B.D.; Sharkey, K.A. Novel functional roles for enteric glia in the gastrointestinal tract. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 625–632. [Google Scholar] [CrossRef] [PubMed]
- Cornet, A.; Savidge, T.C.; Cabarrocas, J.; Deng, W.L.; Colombel, J.F.; Lassmann, H.; Desreumaux, P.; Liblau, R.S. Enterocolitis induced by autoimmune targeting of enteric glial cells: A possible mechanism in Crohn’s disease? Proc. Natl. Acad. Sci. USA 2001, 98, 13306–13311. [Google Scholar] [CrossRef] [Green Version]
- Olsson, T.; Baig, S.; Höjeberg, B.; Link, H. Antimyelin basic protein and antimyelin antibody-producing cells in multiple sclerosis. Ann. Neurol. 1990, 27, 132–136. [Google Scholar] [CrossRef] [PubMed]
- Link, H.; Baig, S.; Olsson, O.; Jiang, Y.-P.; Höjeberg, B.; Olsson, T. Persistent anti-myelin basic protein IgG antibody response in multiple sclerosis cerebrospinal fluid. J. Neuroimmunol. 1990, 28, 237–248. [Google Scholar] [CrossRef]
- Sellebjerg, F.T.; Frederiksen, J.L.; Olsson, T. Anti-myelin basic protein and anti-proteolipid protein antibody-secreting cells in the cerebrospinal fluid of patients with acute optic neuritis. Arch. Neurol. 1994, 51, 1032–1036. [Google Scholar] [CrossRef]
- Sun, J.B.; Olsson, T.; Wang, W.Z.; Xiao, B.G.; Kostulas, V.; Fredrikson, S.; Ekre, H.P.; Link, H. Autoreactive T and B cells responding to myelin proteolipid protein in multiple sclerosis and controls. Eur. J. Immunol. 1991, 21, 1461–1468. [Google Scholar] [CrossRef] [PubMed]
- Abdelhak, A.; Huss, A.; Kassubek, J.; Tumani, H.; Otto, M. Serum GFAP as a biomarker for disease severity in multiple sclerosis. Sci. Rep. 2018, 8, 14798. [Google Scholar] [CrossRef]
- Kassubek, R.; Gorges, M.; Schocke, M.; Hagenston, V.A.M.; Huss, A.; Ludolph, A.C.; Kassubek, J.; Tumani, H. GFAP in early multiple sclerosis: A biomarker for inflammation. Neurosci. Lett. 2017, 657, 166–170. [Google Scholar] [CrossRef]
- Kuerten, S.; Javeri, S.; Tary-Lehmann, M.; Lehmann, P.V.; Angelov, D.N. Fundamental differences in the dynamics of CNS lesion development and composition in MP4- and MOG peptide 35-55-induced experimental autoimmune encephalomyelitis. Clin. Immunol. 2008, 129, 256–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuerten, S.; Kostova-Bales, D.A.; Frenzel, L.P.; Tigno, J.T.; Tary-Lehmann, M.; Angelov, D.N.; Lehmann, P.V. MP4- and MOG:35-55-induced EAE in C57BL/6 mice differentially targets brain, spinal cord and cerebellum. J. Neuroimmunol. 2007, 189, 31–40. [Google Scholar] [CrossRef] [Green Version]
- Recks, M.S.; Stormanns, E.R.; Bader, J.; Arnhold, S.; Addicks, K.; Kuerten, S. Early axonal damage and progressive myelin pathology define the kinetics of CNS histopathology in a mouse model of multiple sclerosis. Clin. Immunol. 2013, 149, 32–45. [Google Scholar] [CrossRef]
- Becker, K.J.; Tanzi, P.; Zierath, D.; Buckwalter, M.S. Antibodies to myelin basic protein are associated with cognitive decline after stroke. J. Neuroimmunol. 2016, 295–296, 9–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gee, J.M.; Kalil, A.; Thullbery, M.; Becker, K.J. Induction of immunologic tolerance to myelin basic protein prevents central nervous system autoimmunity and improves outcome after stroke. Stroke 2008, 39, 1575–1582. [Google Scholar] [CrossRef] [Green Version]
- Shibata, D.; Cain, K.; Tanzi, P.; Zierath, D.; Becker, K. Myelin basic protein autoantibodies, white matter disease and stroke outcome. J. Neuroimmunol. 2012, 252, 106–112. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Zoltewicz, J.S.; Mondello, S.; Newsom, K.J.; Yang, Z.; Yang, B.; Kobeissy, F.; Guingab, J.; Glushakova, O.; Robicsek, S.; et al. Human traumatic brain injury induces autoantibody response against glial fibrillary acidic protein and its breakdown products. PLoS ONE 2014, 9, e92698. [Google Scholar] [CrossRef] [Green Version]
- Hergenroeder, G.W.; Moore, A.N.; Schmitt, K.M.; Redell, J.B.; Dash, P.K. Identification of autoantibodies to glial fibrillary acidic protein in spinal cord injury patients. Neuroreport 2016, 27, 90–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunchok, A.; Zekeridou, A.; McKeon, A. Autoimmune glial fibrillary acidic protein astrocytopathy. Curr. Opin. Neurol. 2019, 32, 452–458. [Google Scholar] [CrossRef]
- Leone, D.P.; Genoud, S.; Atanasoski, S.; Grausenburger, R.; Berger, P.; Metzger, D.; Macklin, W.B.; Chambon, P.; Suter, U. Tamoxifen-inducible glia-specific Cre mice for somatic mutagenesis in oligodendrocytes and Schwann cells. Mol. Cell Neurosci. 2003, 22, 430–440. [Google Scholar] [CrossRef]
- Madisen, L.; Zwingman, T.A.; Sunkin, S.M.; Oh, S.W.; Zariwala, H.A.; Gu, H.; Ng, L.L.; Palmiter, R.D.; Hawrylycz, M.J.; Jones, A.R.; et al. A robust and high-throughput Cre reporting and characterization system for the whole mouse brain. Nat. Neurosci. 2010, 13, 133–140. [Google Scholar] [CrossRef] [Green Version]
- Lalo, U.; Pankratov, Y.; Kirchhoff, F.; North, R.A.; Verkhratsky, A. NMDA receptors mediate neuron-to-glia signaling in mouse cortical astrocytes. J. Neurosci. 2006, 26, 2673–2683. [Google Scholar] [CrossRef] [Green Version]
- Hirrlinger, P.G.; Scheller, A.; Braun, C.; Quintela-Schneider, M.; Fuss, B.; Hirrlinger, J.; Kirchhoff, F. Expression of reef coral fluorescent proteins in the central nervous system of transgenic mice. Mol. Cell. Neurosci. 2005, 30, 291–303. [Google Scholar] [CrossRef] [PubMed]
- Jahn, H.M.; Kasakow, C.V.; Helfer, A.; Michely, J.; Verkhratsky, A.; Maurer, H.H.; Scheller, A.; Kirchhoff, F. Refined protocols of tamoxifen injection for inducible DNA recombination in mouse astroglia. Sci. Rep. 2018, 8, 5913. [Google Scholar] [CrossRef]
- Kilkenny, C.; Browne, W.J.; Cuthill, I.C.; Emerson, M.; Altman, D.G. Improving bioscience research reporting: The ARRIVE guidelines for reporting animal research. PLoS Biol. 2010, 8, e1000412. [Google Scholar] [CrossRef]
- Shi, X.; Yan, C.; Liu, B.; Yang, C.; Nie, X.; Wang, X.; Zheng, J.; Wang, Y.; Zhu, Y. miR-381 Regulates Neural Stem Cell Proliferation and Differentiation via Regulating Hes1 Expression. PLoS ONE 2015, 10, e0138973. [Google Scholar] [CrossRef] [PubMed]
- Chew, L.-J.; Coley, W.; Cheng, Y.; Gallo, V. Mechanisms of regulation of oligodendrocyte development by p38 mitogen-activated protein kinase. J. Neurosci. 2010, 30, 11011–11027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, P.; Bagnell, R.; D’Ercole, A.J. Mouse NG2 + Oligodendrocyte Precursors Express mRNA for Proteolipid Protein But Not Its DM-20 Variant: A Study of Laser Microdissection-Captured NG2 + Cells. J. Neurosci. 2003, 23, 4401–4405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hellani, A.; Ji, J.; Mauduit, C.; Deschildre, C.; Tabone, E.; Benahmed, M. Developmental and hormonal regulation of the expression of oligodendrocyte-specific protein/claudin 11 in mouse testis. Endocrinology 2000, 141, 3012–3019. [Google Scholar] [CrossRef] [PubMed]
- Delarasse, C.; Daubas, P.; Mars, L.T.; Vizler, C.; Litzenburger, T.; Iglesias, A.; Bauer, J.; Della Gaspera, B.; Schubart, A.; Decker, L.; et al. Myelin/oligodendrocyte glycoprotein–deficient (MOG-deficient) mice reveal lack of immune tolerance to MOG in wild-type mice. J. Clin. Investig. 2003, 112, 544–553. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N° | Staining Protocol | Antibody Setup |
|---|---|---|
| ① | Quadruple staining of PLP, Calbindin D28k, MBP and Hoechst for Cerebellar Tissue | Primary Antibodies |
| Rat anti-PLP Guinea pig anti-Calbindin D28k Rabbit anti-MBP | ||
| Secondary Antibodies | ||
| Donkey anti-rat Alexa 488 Goat anti-guinea pig Alexa 555 Donkey anti-rabbit Alexa 647 Hoechst | ||
| ② | Quadruple staining of PLP, Calbindin D28k, GFAP and Hoechst for Cerebellar Tissue | Primary Antibodies |
| Rat anti-PLP Guinea pig anti-Calbindin D28k Rabbit anti-GFAP | ||
| Secondary Antibodies | ||
| Donkey anti-rat Alexa 488 Goat anti-guinea pig Alexa 555 Donkey anti-rabbit Alexa 647 Hoechst | ||
| ③ | Triple Staining of GFAP, Synaptophysin and α-BT for Skeletal Muscle and Esophagus | Primary Antibodies |
| Rabbit anti-GFAP Guinea pig anti-Synaptophysin | ||
| Secondary Antibodies and Toxins | ||
| Donkey anti-rabbit Alexa 488 Donkey anti-guinea pig Alexa 647 α-Bungarotoxin Alexa 555 | ||
| ④ | Triple staining of MBP, Synaptophysin and α-BT for Skeletal Muscle | Primary Antibodies |
| Rabbit anti-MBP Guinea pig anti-Synaptophysin | ||
| Secondary Antibodies and Toxins | ||
| Donkey anti-rabbit Alexa 488 Donkey anti-guinea pig Alexa 647 α-Bungarotoxin Alexa 555 | ||
| ⑤ | Quadruple staining of PLP, Synaptophysin, α-BT and Hoechst for Skeletal Muscle and Esophagus | Primary Antibodies |
| Rat anti-PLP Guinea pig anti-Synaptophysin | ||
| Secondary Antibodies and Toxins | ||
| Donkey anti-rat Alexa 488 Donkey anti-guinea pig Alexa 647 α-Bungarotoxin Alexa 555 Hoechst | ||
| ⑥ | Triple Staining of ChAT, MBP and α-BT for Skeletal Muscle | Primary Antibodies |
| Goat anti-ChAT Rabbit anti-MBP | ||
| Secondary Antibodies and Toxins | ||
| Donkey anti-goat Alexa 647 Donkey anti-rabbit Alexa 488 α-Bungarotoxin Alexa 555 | ||
| ⑦ | Triple Staining of ChAT, Synaptophysin and α-BT for Skeletal Muscle | Primary Antibodies |
| Goat anti-ChAT Guinea pig anti-Synaptophysin | ||
| Secondary Antibodies and Toxins | ||
| Donkey anti-goat Alexa 488 Donkey anti-guinea pig Alexa 647 α-Bungarotoxin Alexa 555 | ||
| ⑧ | Triple Staining of βIII-tubulin, MBP and α-BT for Skeletal Muscle | Primary Antibodies |
| Rabbit anti-βIII-tubulin Chicken anti-MBP | ||
| Secondary Antibodies and Toxins | ||
| Donkey anti-rabbit Alexa 488 Goat anti-chicken Alexa 647 α-Bungarotoxin Alexa 555 |
| N° | Staining Protocol | Antibody Setup |
|---|---|---|
| ① | Quadruple Staining of MBP, ChAT, α-BT and Hoechst | Primary Antibodies |
| Rabbit anti-MBP Goat anti-ChAT | ||
| Secondary Antibodies and Toxins | ||
| Donkey anti-rabbit Alexa 488 Donkey anti-Goat Alexa 647 α-Bungarotoxin Alexa 555 Hoechst | ||
| ② | Triple Staining of DsRed, PLP and Protein Gene Product 9.5 (PGP 9.5) in PLP-CreERT2 x tdT mice | Primary Antibodies |
| Rabbit anti-DsRed (This antibody also binds to tdTomato) Rat anti-PLP Guinea pig anti-PGP 9.5 | ||
| Secondary Antibodies | ||
| Donkey anti-rabbit Alexa 647 Donkey anti-rat Alexa 488 Donkey anti-guinea pig DYE 405 | ||
| ③ | Triple Staining of GFAP, PGP 9.5 and Hoechst | Primary Antibodies |
| Rabbit anti-GFAP Guinea pig anti-PGP 9.5 | ||
| Secondary Antibodies and Toxins | ||
| Donkey anti-rabbit Alexa 488 Goat anti-guinea pig Alexa 555 Hoechst | ||
| ④ | Double Staining of GFP and DsRed in GFAP-EGFP x PLP-DsRed1 Mice | Primary Antibodies |
| Chicken anti-GFP Rabbit anti-DsRed | ||
| Secondary Antibodies and Toxins | ||
| Goat anti-chicken Alexa 647 Donkey anti-rabbit DYE 405 | ||
| ⑤ | Triple staining of βIII-Tubulin, MBP and α-BT | Primary Antibodies |
| Rabbit anti- βIII-tubulin Chicken anti-MBP | ||
| Secondary Antibodies and Toxins | ||
| Donkey anti-rabbit Alexa 488 Donkey anti-chicken Alexa 647 α-Bungarotoxin Alexa 555 |
| Gene | Forward Primer Reverse Primer Primer Reference | Cycle Conditions | |||
|---|---|---|---|---|---|
| β-Actin (154 bp) |
(F) 5′-GGCTGTATTCCCCTCCATCG-3′ (R) 5′-CCAGTTGGTAACAATGCCATGT-3′ Self-Designed | Initial Denaturation | 10 min | 95 °C | |
| 35 Cycles | Denaturation | 45 s | 95 °C | ||
| Annealing | 30 s | 57 °C | |||
| Extension | 45 s | 72 °C | |||
| Final extension | 10 min | 72 °C | |||
| GFAP (199 bp) | (F) 5′-CAACGTTAAGCTAGCCCTGGACAT-3′ (R) 5′-CTCACCATCCCGCATCTCCACAGT-3′ Shi et al. [77] | Initial denaturation | 10 min | 95 °C | |
| 35 Cycles | Denaturation | 45 s | 95 °C | ||
| Annealing | 30 s | 60 °C | |||
| Extension | 45 s | 72 °C | |||
| Final extension | 10 min | 72 °C | |||
| PLP (218 bp) | (F) 5′-AGCGGGTGTGTCATTGTTTGGGAA-3′ (R) 5′-ACCATACATTCTGGCATCAGCGCA-3′ Chew et al. [78] | Initial denaturation | 10 min | 95 °C | |
| 35 Cycles | Denaturation | 45 s | 95 °C | ||
| Annealing | 30 s | 58 °C | |||
| Extension | 45 s | 72 °C | |||
| Final extension | 10 min | 72 °C | |||
| MBP (342–642 bp) | (F) 5′-ATGGCATCACAGAAGAGACC-3′ (R) 5′-CATGGGAGATCCAGAGCGGC-3′ Ye et al. [79] | Initial denaturation | 10 min | 95 °C | |
| 35 Cycles | Denaturation | 45 s | 95 °C | ||
| Annealing | 30 s | 56 °C | |||
| Extension | 45 s | 72 °C | |||
| Final extension | 10 min | 72 °C | |||
| MAG (355–400 bp) | (F) 5′-CTCTATGGCACCCAGAGCCT-3′ (R) 5′-TGTCCTTGGTGGGTCGTTTT-3′ Ye et al. [79] | Initial denaturation | 10 min | 95 °C | |
| 35 Cycles | Denaturation | 45 s | 95 °C | ||
| Annealing | 30 s | 56 °C | |||
| Extension | 45 s | 72 °C | |||
| Final extension | 10 min | 72 °C | |||
| OSP (339 bp) | (F) 5′-GATTGGCATCATCGTCACAACG-3′ (R) 5′-AGCCAGCAGAATAAGGAGCACC-3′ Hellani et al. [80] | Initial denaturation | 10 min | 95 °C | |
| 35 Cycles | Denaturation | 45 s | 95 °C | ||
| Annealing | 30 s | 50 °C | |||
| Extension | 45 s | 72 °C | |||
| Final extension | 10 min | 72 °C | |||
| MOG (841 bp) | (F) 5′-GACCTCAGCTTGGCCTGACCC-3′ (R) 5′-TGCTGGGCTCTCCTTCCGC-3′ Delarasse et al. [81] | Initial denaturation | 5 min | 94 °C | |
| 35 Cycles | Denaturation | 1 min | 95 °C | ||
| Annealing | 1 min | 66 °C | |||
| Extension | 3 min | 72 °C | |||
| Final extension | 5 min | 72 °C | |||
| Primary Antibodies | Host Species | Dilution | Source (Catalogue Number) |
|---|---|---|---|
| Calbindin D28k | Guinea pig | 1:100 | Synaptic Systems Göttingen, Germany (214 004) |
| ChAT | Goat | 1:40 | Millipore Temecula, CA, USA (AB144P–1ML) |
| DsRed | Rabbit | 1:1000 | Takara Bio Mountain View, CA, USA (632496) |
| GFAP | Rabbit | 1:800–1:2000 | Dako Glostrup, Denmark (Z0334) |
| GFP | Chicken | 1:1000 | ThermoFisher Waltham, MA, USA (A10262) |
| MBP | Rabbit | 1:200–1:500 | Abcam Cambridge, UK (ab40390) |
| MBP | Chicken | 1:500–1:3000 | Abcam Cambridge, UK (ab134018) |
| PGP 9.5 | Guinea pig | 1:500 | Fitzgerald Ind. Acton, MA, USA (20R–PG011) |
| PLP (PLP1) | Rat | 1:1000 | Kind gift from Wendy B. Macklin |
| Synaptophysin | Guinea pig | 1:1000 | Synaptic Systems Göttingen, Germany (101 004) |
| βIII-Tubulin | Rabbit | 1:500–1:4000 | Abcam Cambridge, UK (ab18207) |
| Secondary Antibodies and Toxins | Dilution | Source (Catalogue Number) | |
| Donkey Anti-Chicken Alexa 647 | 1:1000 | Jackson ImmunoResearch West Grove, PA, USA (703–605–155) | |
| Goat Anti-Chicken Alexa 647 | 1:1000 | Jackson ImmunoResearch West Grove, PA, USA (103–605–155) | |
| Donkey Anti-Goat Alexa 488 | 1:1000 | Molecular Probes Eugene, OR, USA (A11055) | |
| Donkey Anti-Goat Alexa 647 | 1:1000 | Molecular Probes Eugene, OR, USA (A21447) | |
| Donkey Anti-Guinea Pig DYE 405 | 1:200 | Jackson ImmunoResearch West Grove, PA, USA (706–475–148) | |
| Goat Anti-Guinea Pig Alexa 555 | 1:1000 | Molecular Probes Eugene, OR, USA (A–21435) | |
| Donkey Anti-Guinea Pig Alexa 647 | 1:1000 | Jackson ImmunoResearch West Grove, PA, USA (706–605–148) | |
| Donkey Anti-Rabbit DYE 405 | 1:200 | Jackson ImmunoResearch West Grove, PA, USA (711–475–152) | |
| Donkey Anti-Rabbit Alexa 488 | 1:1000 | ThermoFisher Waltham, MA, USA (A–21206) | |
| Donkey Anti-Rabbit Alexa 647 | 1:1000 | Molecular Probes Eugene, OR, USA (A31573) | |
| Donkey Anti-Rat Alexa 488 | 1:1000 | ThermoFisher Waltham, MA, USA (A–21208) | |
| α-Bungarotoxin Alexa 555 | 1:1000 | Molecular Probes Eugene, OR, USA (B35451) | |
| Hoechst | 1:1000 | Sigma-Aldrich St. Louis, MO, USA (H6024) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kapitza, C.; Chunder, R.; Scheller, A.; Given, K.S.; Macklin, W.B.; Enders, M.; Kuerten, S.; Neuhuber, W.L.; Wörl, J. Murine Esophagus Expresses Glial-Derived Central Nervous System Antigens. Int. J. Mol. Sci. 2021, 22, 3233. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22063233
Kapitza C, Chunder R, Scheller A, Given KS, Macklin WB, Enders M, Kuerten S, Neuhuber WL, Wörl J. Murine Esophagus Expresses Glial-Derived Central Nervous System Antigens. International Journal of Molecular Sciences. 2021; 22(6):3233. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22063233
Chicago/Turabian StyleKapitza, Christopher, Rittika Chunder, Anja Scheller, Katherine S. Given, Wendy B. Macklin, Michael Enders, Stefanie Kuerten, Winfried L. Neuhuber, and Jürgen Wörl. 2021. "Murine Esophagus Expresses Glial-Derived Central Nervous System Antigens" International Journal of Molecular Sciences 22, no. 6: 3233. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22063233