
Foxp3 Silencing with Antisense Oligonucleotide Improves Immunogenicity of an Adjuvanted Recombinant Vaccine against Sporothrix schenckii

 ,
,  , , , , , , , and
, , , , , , , and
Abstract
:1. Introduction
2. Results
2.1. Primary Sequence of the Foxp3 Gene in FASTA Format and Target Region for Foxp3 Silencing
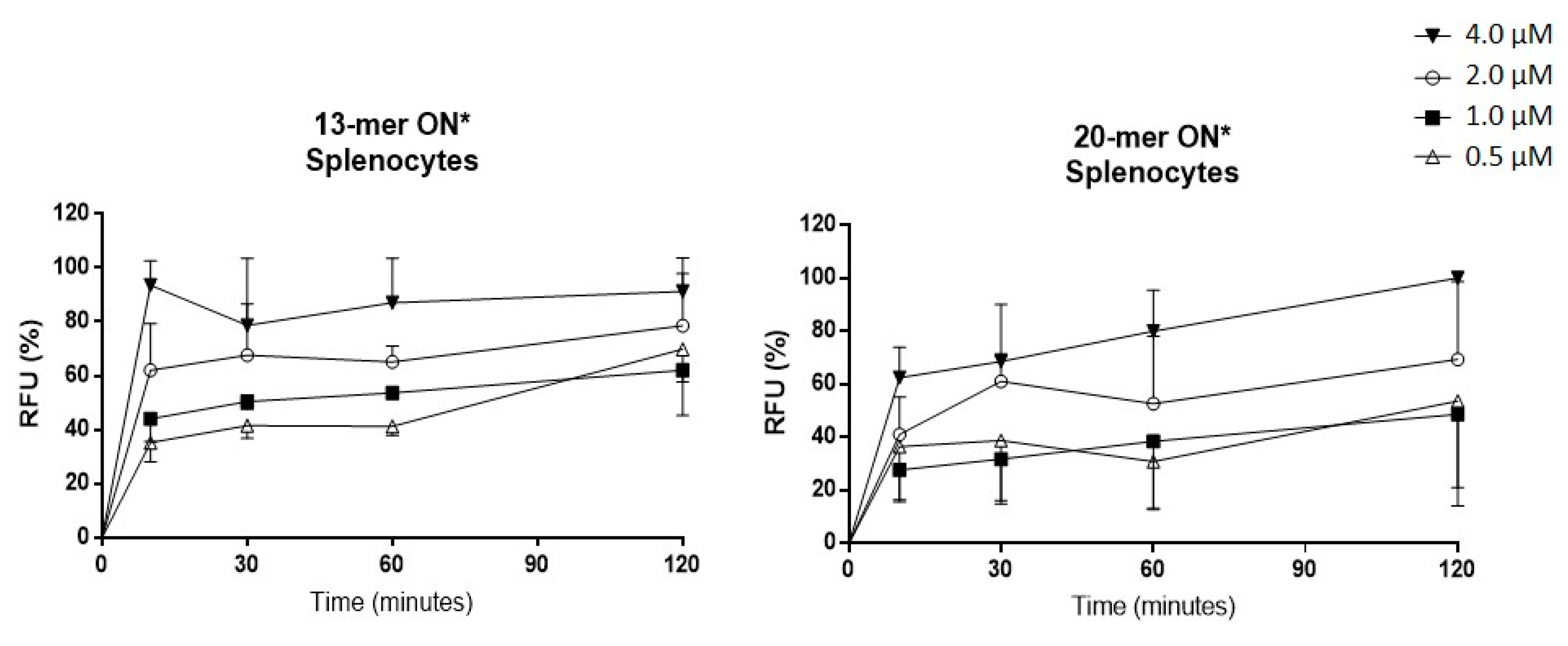
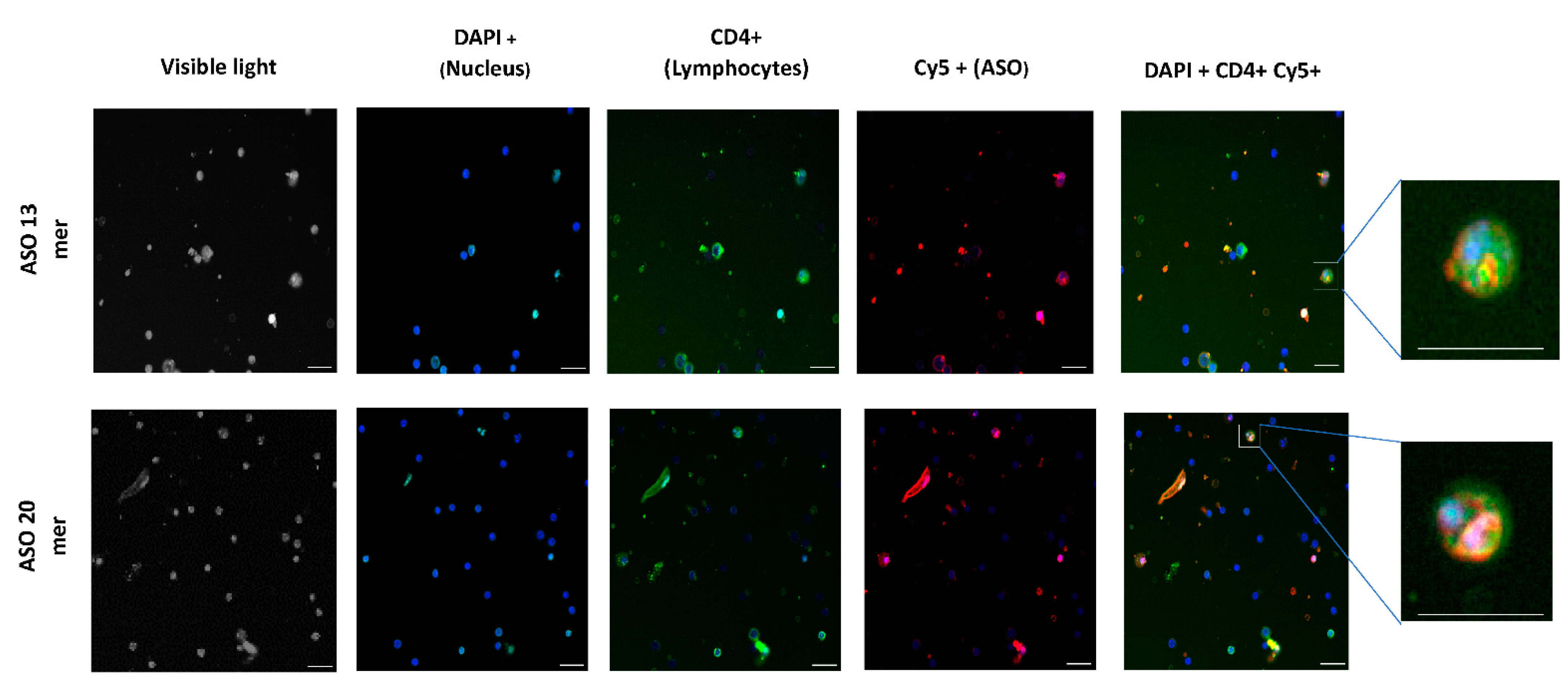
2.2. Oligonucleotide (ON) Uptake Kinetics in Murine Splenocytes
2.3. Biological Activity of the ASO Foxp3
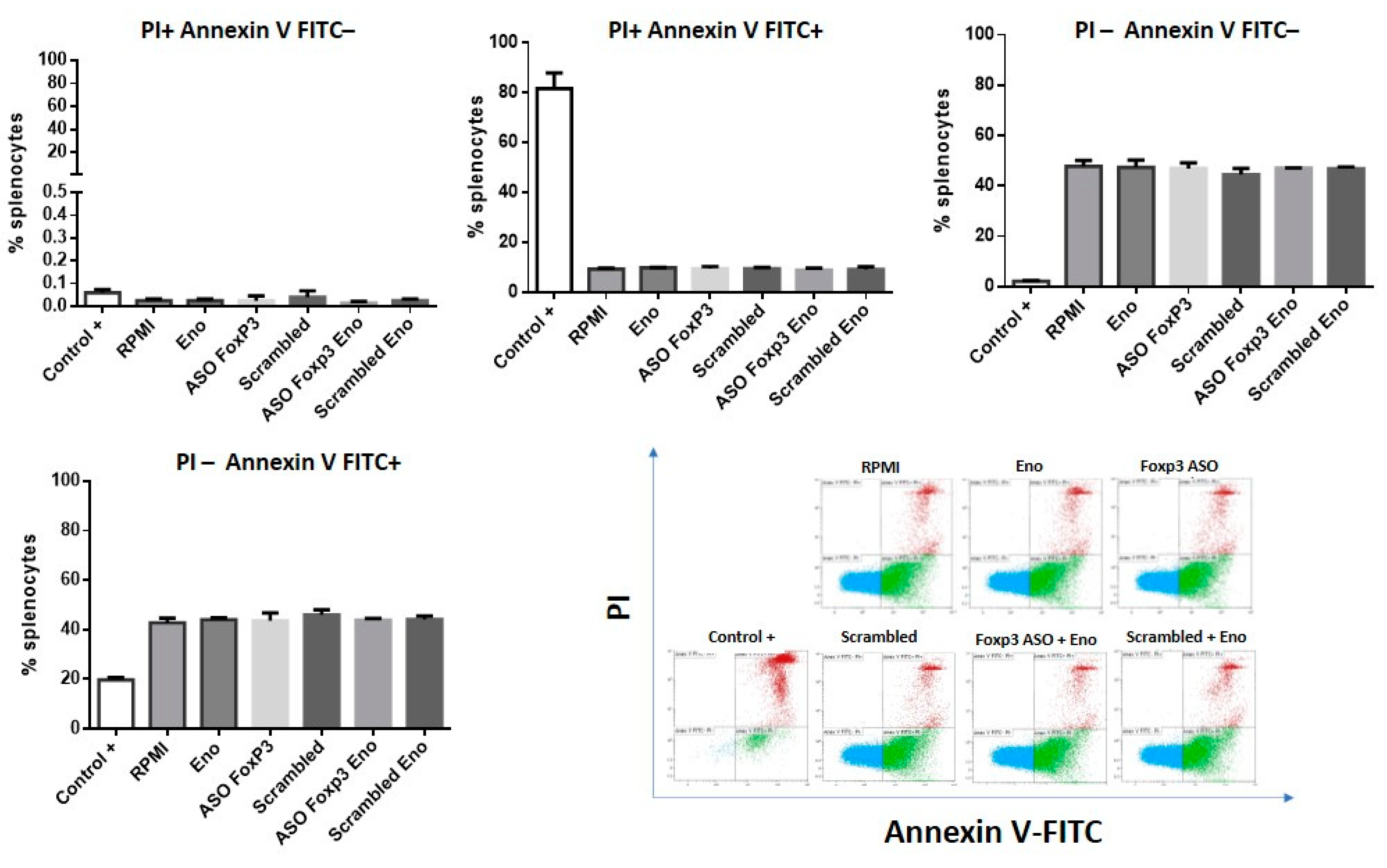
2.4. Cellular Viability
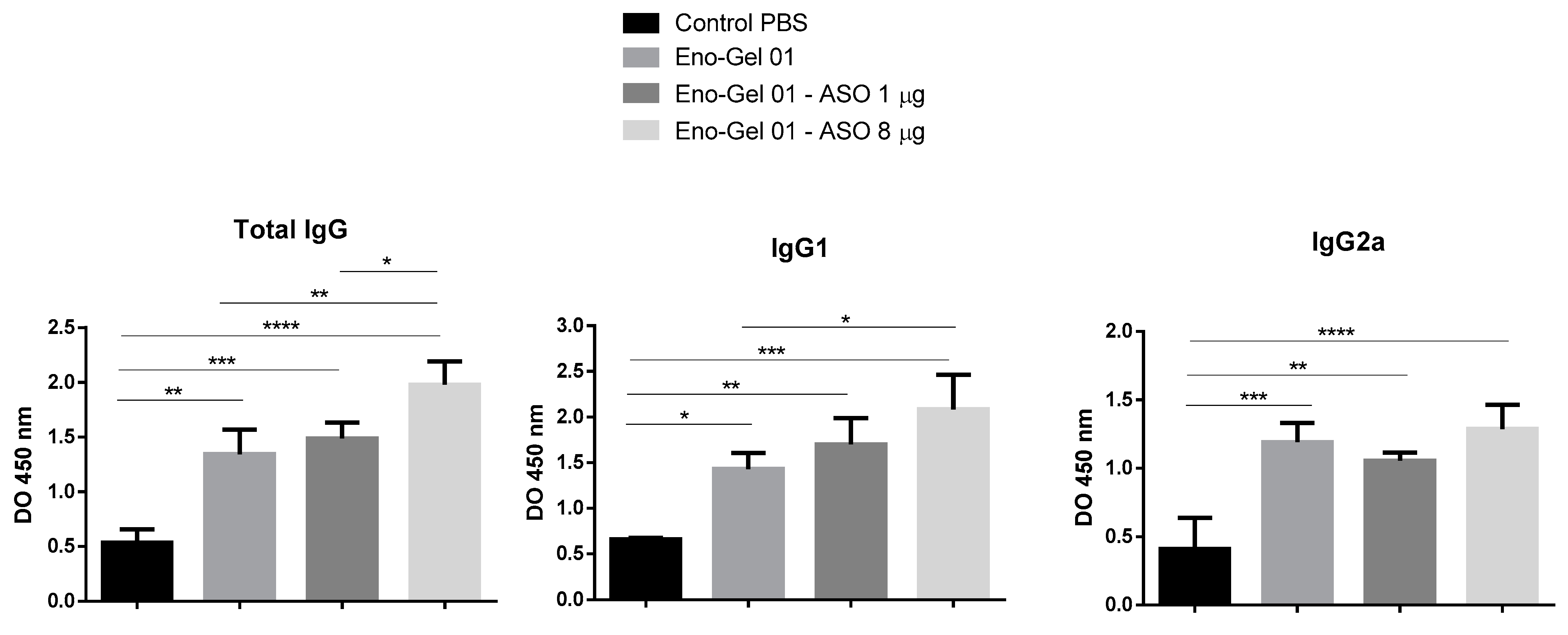
2.5. Immunogenicity Study
2.5.1. Anti-Eno Antibodies
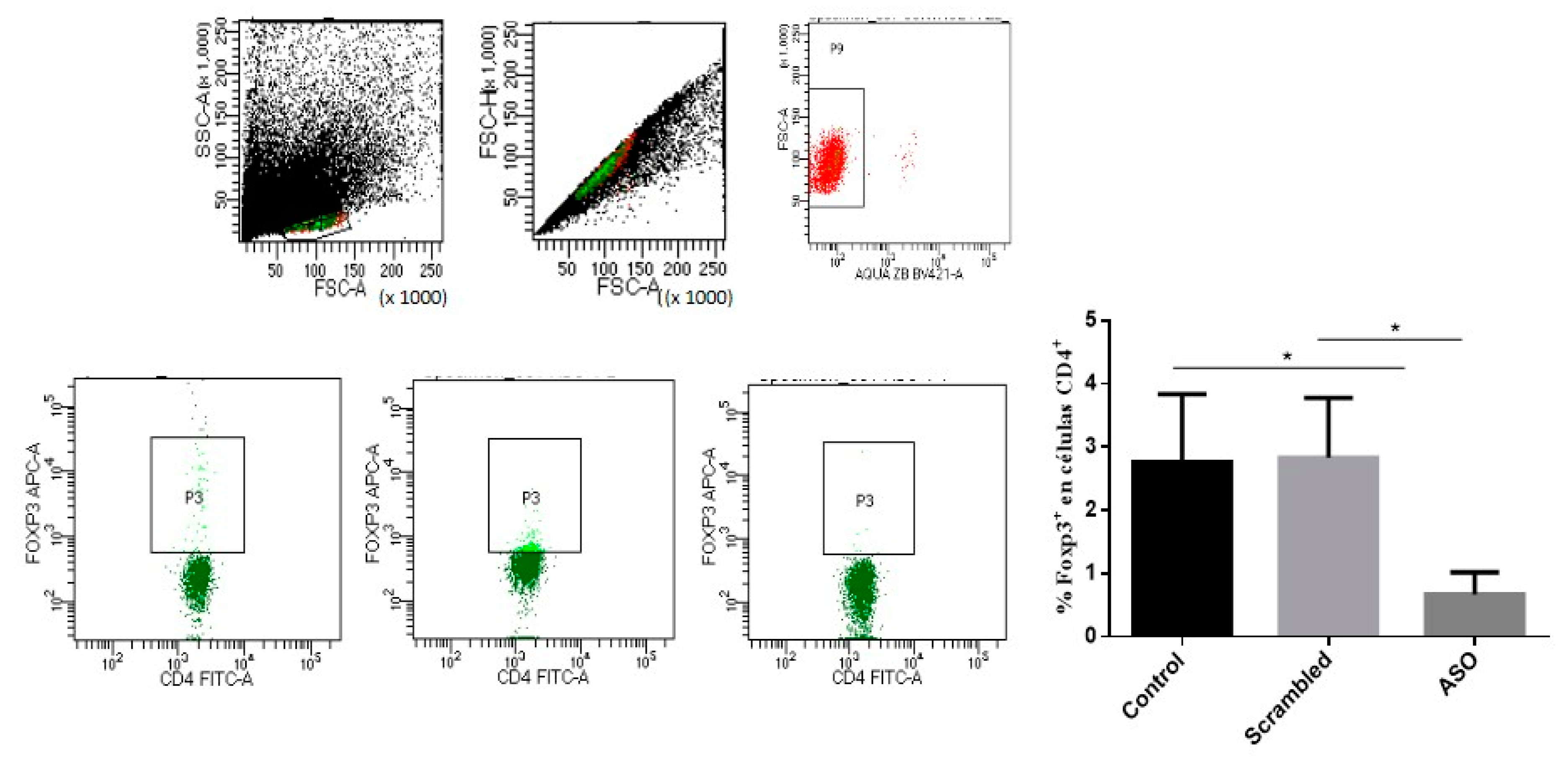
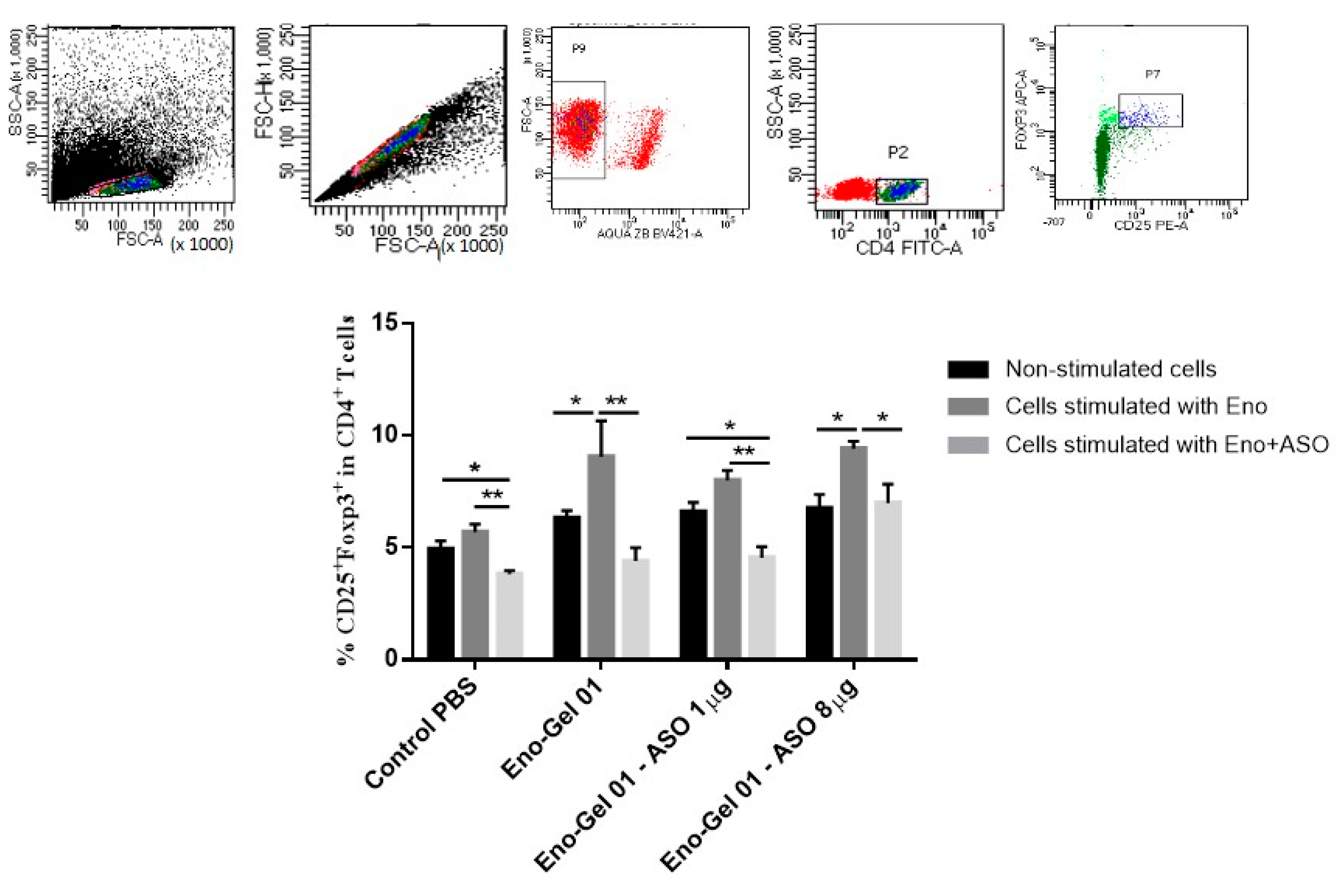
2.5.2. CD4+ CD25+ Foxp3 T Cells
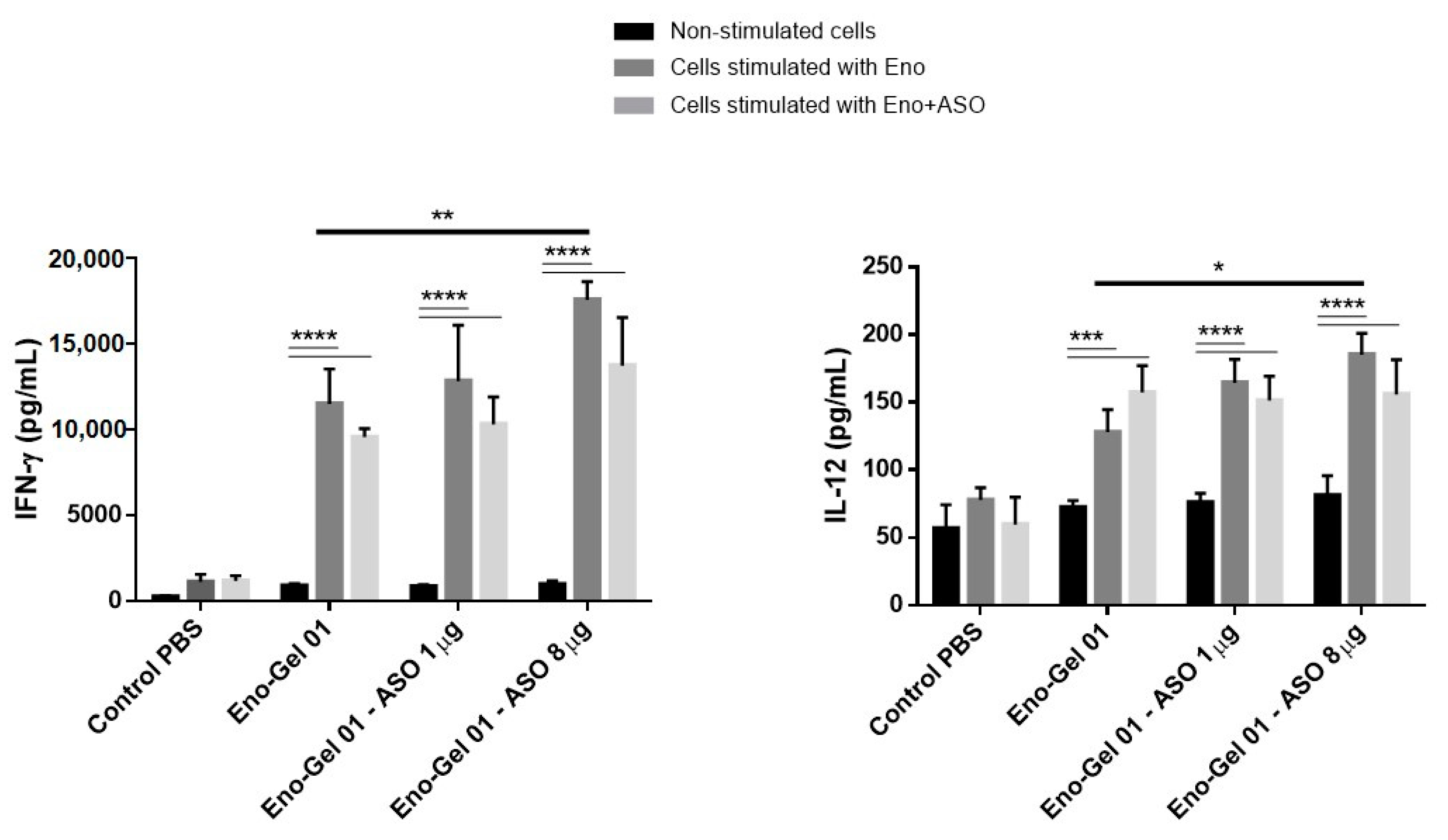
2.5.3. IFN-γ and IL-12 Production
3. Discussion
4. Materials and Methods
4.1. Oligonucleotides (ONs)
4.2. ON Uptake Kinetics in Murine Splenocytes
4.3. Biological Activity of the ASO Foxp3
4.4. Recombinant Sporothrix schenckii Enolase
4.5. Cytotoxicity of ASO Foxp3 and Eno
4.6. Adjuvants and Vaccine Formulation
4.7. Immunization Schedule
4.8. Quantification of Anti-Eno Antibody Response by ELISA
4.9. Quantification of IFN-γ and IL-12 in Splenocyte Culture Supernatant
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Brunkow, M.E.; Jeffery, E.W.; Hjerrild, K.A.; Paeper, B.; Clark, L.B.; Yasayko, S.A.; Wilkinson, J.E.; Galas, D.; Ziegler, S.F.; Ramsdell, F. Disruption of a new forkhead/winged-helix protein, scurfin, results in the fatal lymphoproliferative disorder of the scurfy mouse. Nat. Genet. 2001, 27, 68–73. [Google Scholar] [CrossRef]
- Hori, S.; Nomura, T.; Sakaguchi, S. Control of regulatory T cell development by the transcription factor Foxp3. Science 2003, 299, 1057–1061. [Google Scholar] [CrossRef] [Green Version]
- Fontenot, J.D.; Gavin, M.A.; Rudensky, A.Y. Foxp3 programs the development and function of CD4+CD25+ regulatory T cells. Nat. Immunol. 2003, 4, 330–336. [Google Scholar] [CrossRef] [PubMed]
- Netea, M.G.; Sutmuller, R.; Hermann, C.; Van der Graaf, C.A.; Van der Meer, J.W.; van Krieken, J.H.; Hartung, T.; Adema, G.; Kullberg, B.J. Toll-like receptor 2 suppresses immunity against Candida albicans through induction of IL-10 and regulatory T cells. J. Immunol. 2004, 172, 3712–3718. [Google Scholar] [CrossRef] [Green Version]
- Felonato, M.; Pina, A.; de Araujo, E.F.; Loures, F.V.; Bazan, S.B.; Feriotti, C.; Calich, V.L. Anti-CD25 treatment depletes Treg cells and decreases disease severity in susceptible and resistant mice infected with Paracoccidioides brasiliensis. PLoS ONE 2012, 7, e51071. [Google Scholar] [CrossRef]
- Galdino, N.; Loures, F.V.; de Araújo, E.F.; da Costa, T.A.; Preite, N.W.; Calich, V. Depletion of regulatory T cells in ongoing paracoccidioidomycosis rescues protective Th1/Th17 immunity and prevents fatal disease outcome. Sci. Rep. 2018, 8, 16544. [Google Scholar] [CrossRef]
- Whibley, N.; Maccallum, D.M.; Vickers, M.A.; Zafreen, S.; Waldmann, H.; Hori, S.; Gaffen, S.L.; Gow, N.A.; Barker, R.N.; Hall, A.M. Expansion of Foxp3(+) T-cell populations by Candida albicans enhances both Th17-cell responses and fungal dissemination after intravenous challenge. Eur. J. Immunol. 2014, 44, 1069–1083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batista-Duharte, A.; Téllez-Martínez, D.; de Andrade, C.R.; Polesi, M.C.; Portuondo, D.L.; Carlos, I.Z. Transient Foxp3(+) regulatory T-cell depletion enhances protective Th1/Th17 immune response in murine sporotrichosis caused by Sporothrix schenckii. Immunobiology 2020, 225, 151993. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, N.G.; Chattopadhyay, S.; Mehrotra, S.; Chhabra, A.; Mukherji, B. Regulatory T-cell response and tumor vaccine-induced cytotoxic T lymphocytes in human melanoma. Hum. Immunol. 2004, 65, 794–802. [Google Scholar] [CrossRef] [PubMed]
- Klages, K.; Mayer, C.T.; Lahl, K.; Loddenkemper, C.; Teng, M.W.; Ngiow, S.F.; Smyth, M.J.; Hamann, A.; Huehn, J.; Sparwasser, T. Selective depletion of Foxp3+ regulatory T cells improves effective therapeutic vaccination against established melanoma. Cancer Res. 2010, 70, 7788–7799. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Tan, D.; Liu, H.; Li, K. CD4(+) CD25(+) FoxP3(+) T regulatory cells in subjects responsive or unresponsive to hepatitis B vaccination. Zhong Nan da Xue Xue Bao. Yi Xue Ban 2011, 36, 1046–1051. [Google Scholar] [CrossRef]
- Stein, P.; Weber, M.; Prüfer, S.; Schmid, B.; Schmitt, E.; Probst, H.C.; Waisman, A.; Langguth, P.; Schild, H.; Radsak, M.P. Regulatory T cells and IL-10 independently counterregulate cytotoxic T lymphocyte responses induced by transcutaneous immunization. PLoS ONE 2011, 6, e27911. [Google Scholar] [CrossRef] [Green Version]
- Espinoza Mora, M.R.; Steeg, C.; Tartz, S.; Heussler, V.; Sparwasser, T.; Link, A.; Fleischer, B.; Jacobs, T. Depletion of regulatory T cells augments a vaccine-induced T effector cell response against the liver-stage of malaria but fails to increase memory. PLoS ONE 2014, 9, e104627. [Google Scholar] [CrossRef] [Green Version]
- Mousavi Niri, N.; Memarnejadian, A.; Pilehvar-Soltanahmadi, Y.; Agha Sadeghi, M.; Mahdavi, M.; Kheshtchin, N.; Arab, S.; Namdar, A.; Jadidi, F.; Zarghami, N.; et al. Improved anti-treg vaccination targeting Foxp3 efficiently decreases regulatory T cells in mice. J. Immunother. 2016, 39, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Batista-Duharte, A.; Téllez-Martínez, D.; Fuentes, D.; Carlos, I.Z. Molecular adjuvants that modulate regulatory T cell function in vaccination: A critical appraisal. Pharmacol. Res. 2018, 129, 237–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batista-Duharte, A.; Sendra, L.; Herrero, M.J.; Téllez-Martínez, D.; Carlos, I.Z.; Aliño, S.F. Progress in the use of antisense oligonucleotides for vaccine improvement. Biomolecules 2020, 10, 316. [Google Scholar] [CrossRef] [Green Version]
- Crooke, S.T. Molecular mechanisms of antisense oligonucleotides. Nucleic Acid Ther. 2017, 27, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Dhuri, K.; Bechtold, C.; Quijano, E.; Pham, H.; Gupta, A.; Vikram, A.; Bahal, R. Antisense oligonucleotides: An emerging area in drug discovery and development. J. Clin. Med. 2020, 9, 2004. [Google Scholar] [CrossRef]
- Lu, L.; Barbi, J.; Pan, F. The regulation of immune tolerance by FOXP3. Nat. Rev. Immunol. 2017, 17, 703–717. [Google Scholar] [CrossRef] [PubMed]
- Nair, S.; Boczkowski, D.; Fassnacht, M.; Pisetsky, D.; Gilboa, E. Vaccination against the forkhead family transcription factor Foxp3 enhances tumor immunity. Cancer Res. 2007, 67, 371–380. [Google Scholar] [CrossRef] [Green Version]
- Mousavi Niri, N.; Memarnejadian, A.; Hadjati, J.; Aghasadeghi, M.R.; Shokri, M.; Pilehvar-Soltanahmadi, Y.; Akbarzadeh, A.; Zarghami, N. Construction and production of Foxp3-Fc (IgG) DNA vaccine/fusion protein. Avicenna J. Med. Biotechnol. 2016, 8, 57–64. [Google Scholar]
- Casares, N.; Rudilla, F.; Arribillaga, L.; Llopiz, D.; Riezu-Boj, J.I.; Lozano, T.; López-Sagaseta, J.; Guembe, L.; Sarobe, P.; Prieto, J.; et al. A peptide inhibitor of FOXP3 impairs regulatory T cell activity and improves vaccine efficacy in mice. J. Immunol. 2010, 185, 5150–5159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lozano, T.; Villanueva, L.; Durántez, M.; Gorraiz, M.; Ruiz, M.; Belsúe, V.; Riezu-Boj, J.I.; Hervás-Stubbs, S.; Oyarzábal, J.; Bandukwala, H.; et al. Inhibition of FOXP3/NFAT interaction enhances T cell function after TCR stimulation. J. Immunol. 2015, 195, 3180–3189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lozano, T.; Soldevilla, M.M.; Casares, N.; Villanueva, H.; Bendandi, M.; Lasarte, J.J.; Pastor, F. Targeting inhibition of Foxp3 by a CD28 2′-Fluro oligonucleotide aptamer conjugated to P60-peptide enhances active cancer immunotherapy. Biomaterials 2016, 91, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Miguel, A.; Sendra, L.; Noé, V.; Ciudad, C.J.; Dasí, F.; Hervas, D.; Herrero, M.J.; Aliño, S.F. Silencing of Foxp3 enhances the antitumor efficacy of GM -CSF genetically modified tumor cell vaccine against B16 melanoma. Onco Targets Ther. 2017, 10, 503–514. [Google Scholar] [CrossRef] [Green Version]
- Hagedorn, P.H.; Pontoppidan, M.; Bisgaard, T.S.; Berrera, M.; Dieckmann, A.; Ebeling, M.; Møller, M.R.; Hudlebusch, H.; Jensen, M.L.; Hansen, H.F.; et al. Identifying and avoiding off-target effects of RNase H-dependent antisense oligonucleotides in mice. Nucleic Acids Res. 2018, 46, 5366–5380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Portuondo, D.L.; Dores-Silva, P.R.; Ferreira, L.S.; de Oliveira, C.S.; Téllez-Martínez, D.; Marcos, C.M.; de Aguiar Loesch, M.L.; Guzmán, F.; Gava, L.M.; Borges, J.C.; et al. Immunization with recombinant enolase of Sporothrix spp. (rSsEno) confers effective protection against sporotrichosis in mice. Sci. Rep. 2019, 9, 17179. [Google Scholar] [CrossRef] [Green Version]
- Téllez-Martínez, D.; Leandro Portuondo, D.; Loesch, M.L.; Batista-Duharte, A.; Zeppone Carlos, I. A Recombinant enolase-Montanide™ PetGel a vaccine promotes a protective Th1 immune response against a highly virulent Sporothrix schenckii by toluene exposure. Pharmaceutics 2019, 11, 144. [Google Scholar] [CrossRef] [Green Version]
- Ripple, M.J.; You, D.; Honnegowda, S.; Giaimo, J.D.; Sewell, A.B.; Becnel, D.M.; Cormier, S.A. Immunomodulation with IL-4Rα antisense oligonucleotide prevents respiratory syncytial virus-mediated pulmonary disease. J. Immunol. 2010, 185, 4804–4811. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Liu, N.; Lu, Y.; Huang, Z.; Zang, Y.; Chen, J.; Zhang, J.; Ding, Z. Phosphorothioated antisense oligodeoxynucleotide suppressing interleukin-10 is a safe and potent vaccine adjuvant. Vaccine 2019, 37, 4081–4088. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yang, L.; Zhao, P.; Yao, Y.; Lu, F.; Tu, L.; Liu, J.; Li, Z.; Yu, Y.; Wang, L. Adjuvanticity of a CTLA-4 3’ UTR complementary oligonucleotide for emulsion formulated recombinant subunit and inactivated vaccines. Vaccine 2017, 35, 2379–2389. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Song, Y.; Cui, C.; Lan, Y.; Li, X.; Liu, Y.; Lu, F.; Zhang, Y.; Yu, Y.; Wang, L. A LAG3-interfering oligonucleotide acts as an adjuvant to enhance the antibody responses induced by recombinant protein vaccines and inactivated influenza virus vaccines. Appl. Microbiol. Biotechnol. 2019, 103, 6543–6557. [Google Scholar] [CrossRef] [PubMed]
- Maia, D.C.; Sassá, M.F.; Placeres, M.C.; Carlos, I.Z. Influence of Th1/Th2 cytokines and nitric oxide in murine systemic infection induced by Sporothrix schenckii. Mycopathologia 2006, 161, 11–19. [Google Scholar] [CrossRef]
- Batista-Duharte, A.; Téllez-Martínez, D.; Roberto de Andrade, C.; Portuondo, D.L.; Jellmayer, J.A.; Polesi, M.C.; Carlos, I.Z. Sporothrix brasiliensis induces a more severe disease associated with sustained Th17 and regulatory T cells responses than Sporothrix schenckii sensu stricto in mice. Fung. Biol. 2018, 122, 1163–1170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Xu, P.; Liu, H.; Fang, C.; Guo, H.; Chen, X.; Tan, M.; Zhang, Y.; Min, W. Silencing IDO2 in dendritic cells: A novel strategy to strengthen cancer immunotherapy in a murine lung cancer model. Int. J. Oncol. 2020, 57, 587–597. [Google Scholar] [CrossRef]
- Colamatteo, A.; Carbone, F.; Bruzzaniti, S.; Galgani, M.; Fusco, C.; Maniscalco, G.T.; Di Rella, F.; de Candia, P.; De Rosa, V. Molecular mechanisms controlling Foxp3 expression in health and autoimmunity: From epigenetic to post-translational regulation. Front. Immunol. 2020, 10, 3136. [Google Scholar] [CrossRef]
- Lahl, K.; Sparwasser, T. In vivo depletion of FoxP3+ Tregs using the DEREG mouse model. Methods Mol. Biol. 2011, 707, 157–172. [Google Scholar] [CrossRef]
- van Elsas, A.; Sutmuller, R.P.; Hurwitz, A.A.; Ziskin, J.; Villasenor, J.; Medema, J.P.; Overwijk, W.W.; Restifo, N.P.; Melief, C.J.; Offringa, R.; et al. Elucidating the autoimmune and antitumor effector mechanisms of a treatment based on cytotoxic T lymphocyte antigen-4 blockade in combination with a B16 melanoma vaccine: Comparison of prophylaxis and therapy. J. Exp. Med. 2001, 194, 481–489. [Google Scholar] [CrossRef]
- Kähler, K.C.; Hassel, J.C.; Heinzerling, L.; Loquai, C.; Thoms, K.M.; Ugurel, S.; Zimmer, L.; Gutzmer, R. Side effect management during immune checkpoint blockade using CTLA-4 and PD-1 antibodies for metastatic melanoma—An update. J. Dtsch. Dermatol. Ges. 2020, 18, 582–609. [Google Scholar] [CrossRef]
- Corsello, S.M.; Barnabei, A.; Marchetti, P.; De Vecchis, L.; Salvatori, R.; Torino, F. Endocrine side effects induced by immune checkpoint inhibitors. J. Clin. Endocrinol. Metab. 2013, 98, 1361–1375. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, L.S.; Gonçalves, A.C.; Portuondo, D.L.; Maia, D.C.; Placeres, M.C.; Batista-Duharte, A.; Carlos, I.Z. Optimal clearance of Sporothrix schenckii requires an intact Th17 response in a mouse model of systemic infection. Immunobiology 2015, 220, 985–992. [Google Scholar] [CrossRef] [PubMed]
- Roberts, T.C.; Langer, R.; Wood, M.J.A. Advances in oligonucleotide drug delivery. Nat. Rev. Drug Discov. 2020, 19, 673–694. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vaccine Formulations/100 µL/Mouse |
|---|
| PBS (Control) |
| 100 µg of Eno in PBS |
| 100 µg of Eno in PBS + 5% Montanide Gel 01 adjuvant (Gel 01) kindly provided by Seppic (Paris, France). |
| 100 µg of Eno in PBS + 5% Gel 01 + ASO anti-Foxp3, 1 µg |
| 100 µg of Eno in PBS + 5% Gel 01 + ASO anti-Foxp3, 8 µg. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Batista-Duharte, A.; Sendra, L.; Herrero, M.J.; Portuondo, D.L.; Téllez-Martínez, D.; Olivera, G.; Fernández-Delgado, M.; Javega, B.; Herrera, G.; Martínez, A.; et al. Foxp3 Silencing with Antisense Oligonucleotide Improves Immunogenicity of an Adjuvanted Recombinant Vaccine against Sporothrix schenckii. Int. J. Mol. Sci. 2021, 22, 3470. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073470
Batista-Duharte A, Sendra L, Herrero MJ, Portuondo DL, Téllez-Martínez D, Olivera G, Fernández-Delgado M, Javega B, Herrera G, Martínez A, et al. Foxp3 Silencing with Antisense Oligonucleotide Improves Immunogenicity of an Adjuvanted Recombinant Vaccine against Sporothrix schenckii. International Journal of Molecular Sciences. 2021; 22(7):3470. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073470
Chicago/Turabian StyleBatista-Duharte, Alexander, Luis Sendra, Maria José Herrero, Deivys Leandro Portuondo, Damiana Téllez-Martínez, Gladys Olivera, Manuel Fernández-Delgado, Beatriz Javega, Guadalupe Herrera, Alicia Martínez, and et al. 2021. "Foxp3 Silencing with Antisense Oligonucleotide Improves Immunogenicity of an Adjuvanted Recombinant Vaccine against Sporothrix schenckii" International Journal of Molecular Sciences 22, no. 7: 3470. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073470