
Agrobacterium-Mediated Capsicum annuum Gene Editing in Two Cultivars, Hot Pepper CM334 and Bell Pepper Dempsey

Abstract
:1. Introduction
2. Results
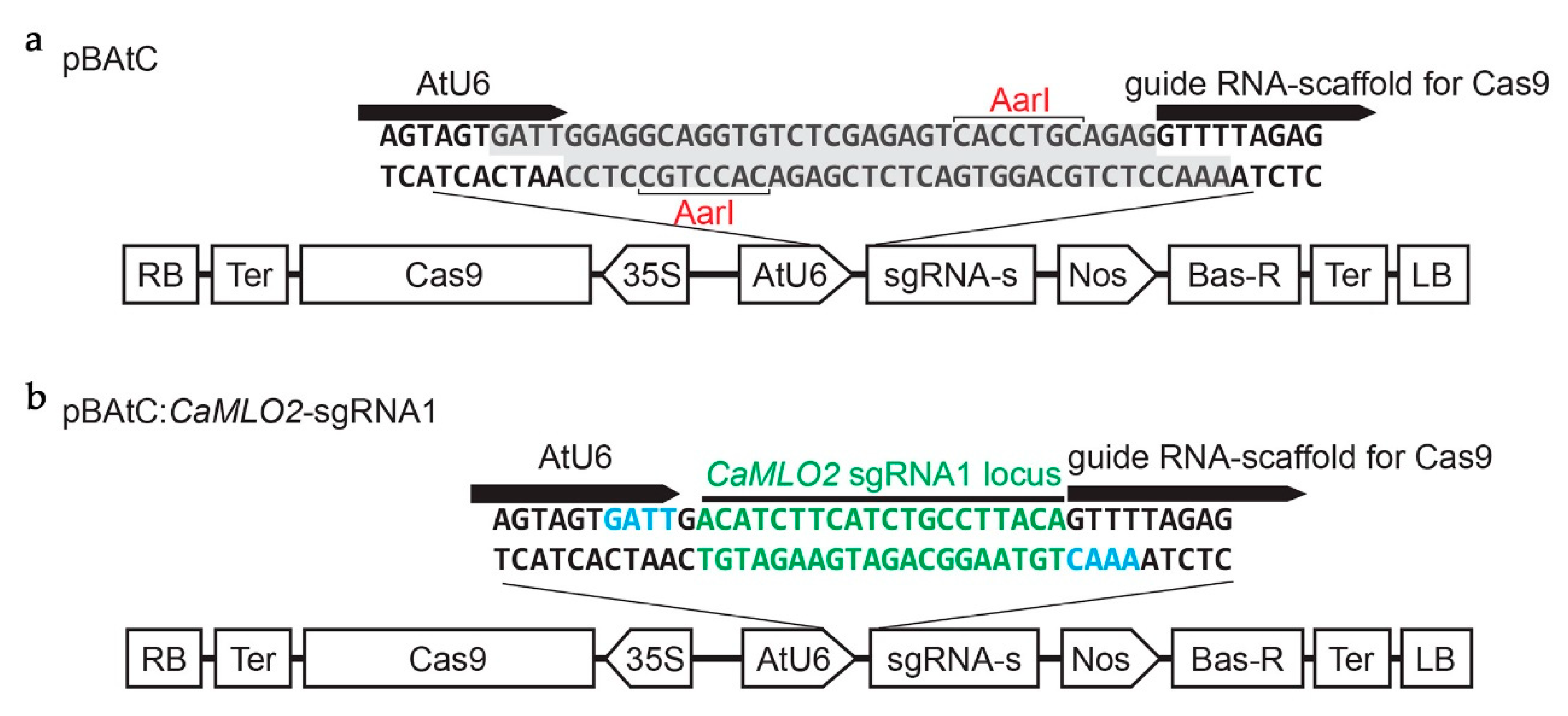
2.1. Vector Construction and Agrobacterium-Mediated Transformation
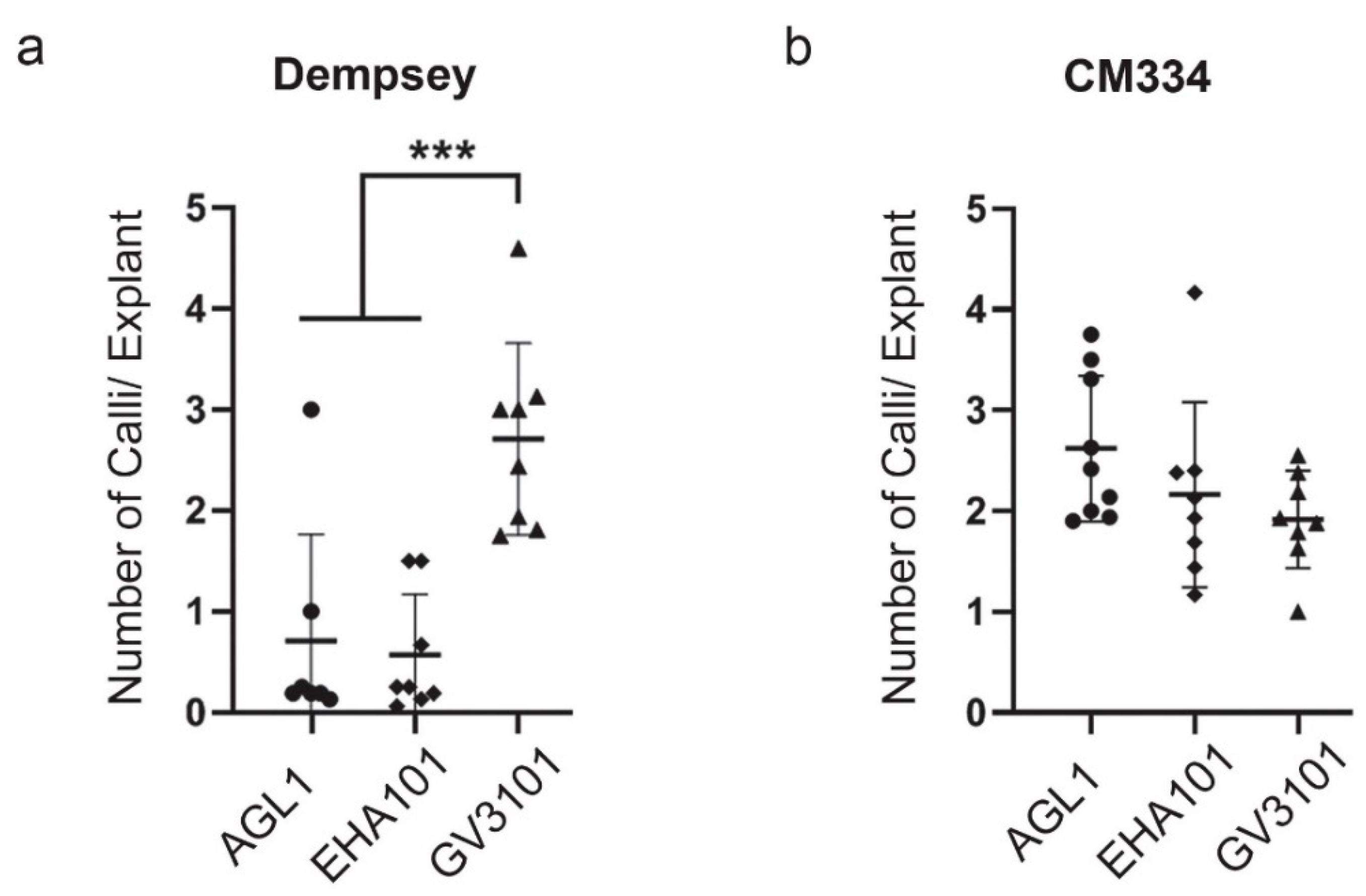
2.2. Comparison of Agrobacterium-Mediated Callus Induction in cv. Dempsey and CM334
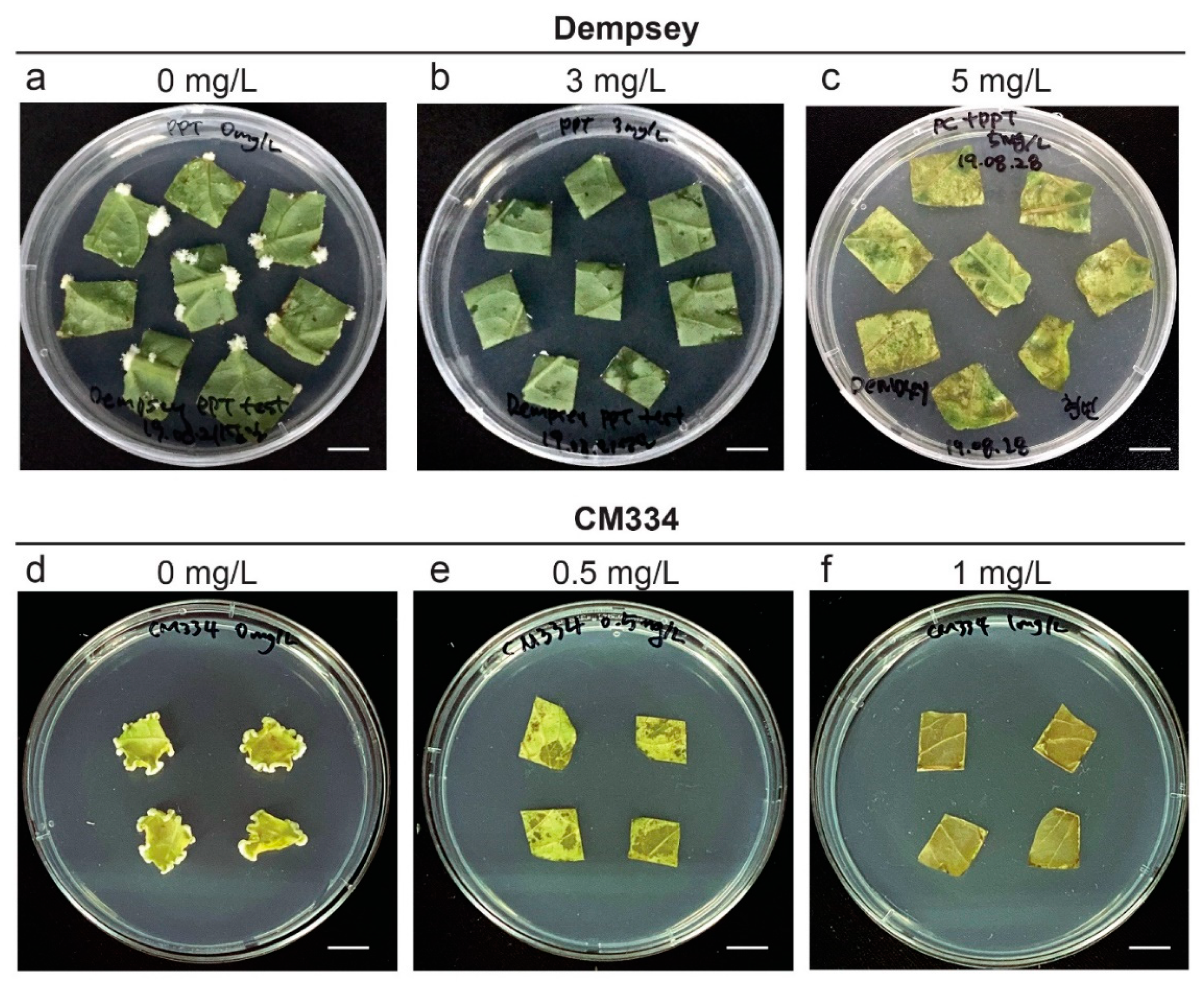
2.3. Suitable Phosphinothricin (PPT) Concentration for Screening Transformants in cv. Dempsey and CM334
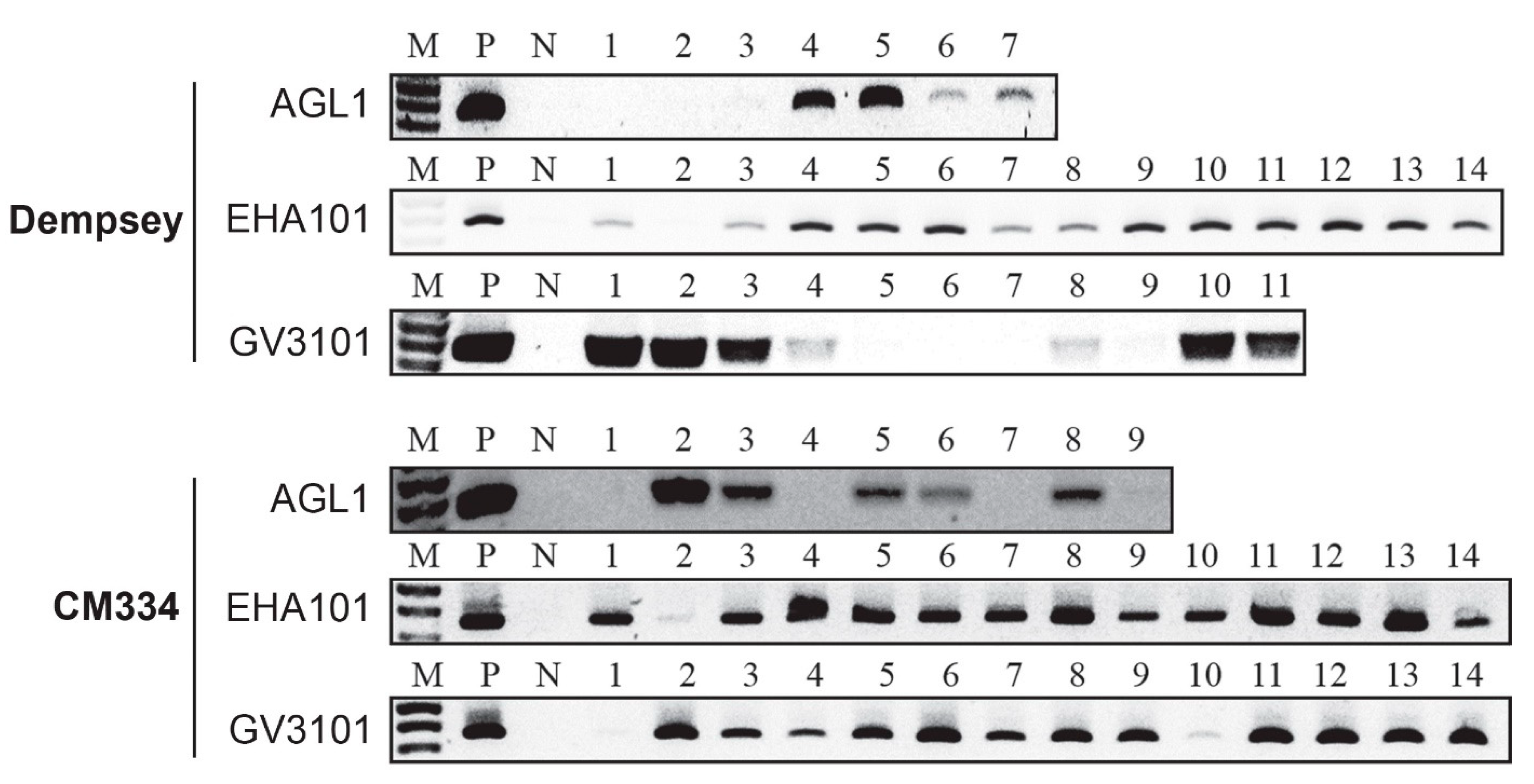
2.4. Evaluation of PPT-Selected Transformants in cv. Dempsey and CM334
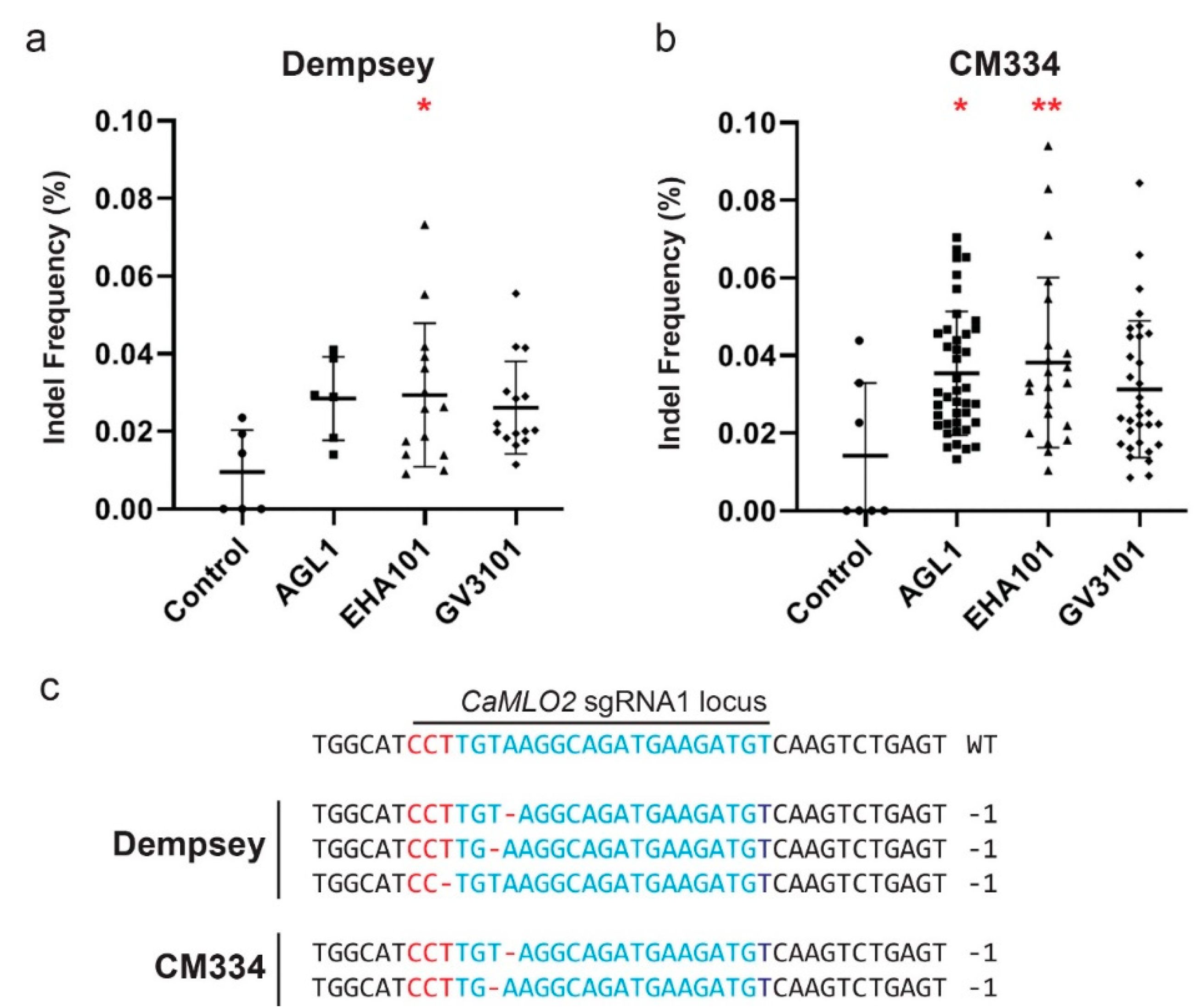
2.5. Analysis of CRISPR/Cas9 Indel Frequencies of Positive Transformants in cv. Dempsey and CM334
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Plasmid Construction
4.3. Agrobacterium-Mediated Transformation
4.4. Callus Induction
4.5. Antibiotic Selection
4.6. Genomic DNA (gDNA) Extraction from Pepper and PCR Analysis
4.7. Targeted Deep Sequencing
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Elad, Y.; Messika, Y.; Brand, M.; David, D.R.; Sztejnberg, A. Effect of colored shade nets on pepper powdery mildew (Leveillula taurica). Phytoparasitica 2007, 35, 285–299. [Google Scholar] [CrossRef]
- Lyngkjær, M.F.; Newton, A.C.; Atzema, J.L.; Baker, S.J. The Barley mlo-gene: An important powdery mildew resistance source. Agronomie 2000, 20, 745–756. [Google Scholar] [CrossRef] [Green Version]
- Acevedo-Garcia, J.; Gruner, K.; Reinstädler, A.; Kemen, A.; Kemen, E.; Cao, L.; Takken, F.L.W.; Reitz, M.U.; Schäfer, P.; O’Connell, R.J.; et al. The powdery mildew-resistant Arabidopsis mlo2 mlo6 mlo12 triple mutant displays altered infection phenotypes with diverse types of phytopathogens. Sci. Rep. 2017, 7, 9319. [Google Scholar] [CrossRef]
- Büschges, R.; Hollricher, K.; Panstruga, R.; Simons, G.; Wolter, M.; Frijters, A.; van Daelen, R.; van der Lee, T.; Diergaarde, P.; Groenendijk, J.; et al. The barley mlo gene: A novel control element of plant pathogen resistance. Cell 1997, 88, 695–705. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Cheng, X.; Shan, Q.; Zhang, Y.; Liu, J.; Gao, C.; Qiu, J.L. Simultaneous editing of three homoeoalleles in hexaploid bread wheat confers heritable resistance to powdery mildew. Nat. Biotechnol. 2014, 32, 947–951. [Google Scholar] [CrossRef] [PubMed]
- Qin, C.; Yu, C.; Shen, Y.; Fang, X.; Chen, L.; Min, J.; Cheng, J.; Zhao, S.; Xu, M.; Luo, Y.; et al. Whole-genome sequencing of cultivated and wild peppers provides insights into Capsicum domestication and specialization. Proc. Natl. Acad. Sci. USA 2014, 111, 5135–5140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Park, M.; Yeom, S.I.; Kim, Y.M.; Lee, J.M.; Lee, H.A.; Seo, E.; Choi, J.; Cheong, K.; Kim, K.T.; et al. Genome sequence of the hot pepper provides insights into the evolution of pungency in Capsicum species. Nat. Genet. 2014, 46, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Seo, E.; Kim, S.; Yeom, S.I.; Choi, D. Genome-wide comparative analyses reveal the dynamic evolution of nucleotide-binding Leucine-rich repeat gene family among Solanaceae plants. Front. Plant Sci. 2016, 7, 1205. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Park, J.; Yeom, S.I.; Kim, Y.M.; Seo, E.; Kim, K.T.; Kim, M.S.; Lee, J.M.; Cheong, K.; Shin, H.S.; et al. New reference genome sequences of hot pepper reveal the massive evolution of plant disease-resistance genes by retroduplication. Genome Biol. 2017, 18, 210. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Choi, J.; Won, K.H. A stable DNA-free screening system for CRISPR/RNPs-mediated gene editing in hot and sweet cultivars of Capsicum annuum. BMC Plant Biol. 2020, 20, 449. [Google Scholar] [CrossRef]
- Nester, E.W. Agrobacterium: Nature’s genetic engineer. Front. Plant Sci. 2015, 5, 730. [Google Scholar] [CrossRef] [Green Version]
- Kyndt, T.; Quispe, D.; Zhai, H.; Jarret, R.; Ghislain, M.; Liu, Q.; Gheysen, G.; Kreuze, J.F. The genome of cultivated sweet potato contains Agrobacterium T-DNAs with expressed genes: An example of a naturally transgenic food crop. Proc. Natl. Acad. Sci. USA 2015, 112, 5844–5849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fraley, R.T.; Rogers, S.G.; Horsch, R.B.; Sanders, P.R.; Flick, J.S.; Adams, S.P.; Bittner, M.L.; Brand, L.A.; Fink, C.L.; Fry, J.S.; et al. Expression of bacterial genes in plant cells. Proc. Natl. Acad. Sci. USA 1983, 80, 4803–4807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrera-Estrella, L.; De Block, M.; Messens, E.; Hernalsteens, J.P.; Van Montagu, M.; Schell, J. Chimeric genes as dominant selectable markers in plant cells. EMBO J. 1983, 2, 987–995. [Google Scholar] [CrossRef] [PubMed]
- Sardesai, N.; Subramanyam, S. Agrobacterium: A genome-editing tool-delivery system. Curr. Top. Microbiol. Immunol. 2018, 418, 463–488. [Google Scholar] [CrossRef]
- Park, S.H.; Morris, J.L.; Park, J.E.; Hirschi, K.D.; Smith, R.H. Efficient and genotype-independent Agrobacterium-mediated tomato transformation. J. Plant Physiol. 2003, 160, 1253–1257. [Google Scholar] [CrossRef] [Green Version]
- Cortina, C.; Culiáñez-Macià, F.A. Tomato transformation and transgenic plant production. Plant Cell Tissue Organ Cult. 2004, 76, 269–275. [Google Scholar] [CrossRef]
- Shan, Q.; Wang, Y.; Li, J.; Zhang, Y.; Chen, K.; Liang, Z.; Zhang, K.; Liu, J.; Xi, J.J.; Qiu, J.L.; et al. Targeted genome modification of crop plants using a CRISPR-Cas system. Nat. Biotechnol. 2013, 31, 686–688. [Google Scholar] [CrossRef]
- Brooks, C.; Nekrasov, V.; Lippman, Z.B.; Van Eck, J. Efficient gene editing in tomato in the first generation using the clustered regularly interspaced short palindromic repeats/CRISPR-associated9 system. Plant Physiol. 2014, 166, 1292–1297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butler, N.M.; Baltes, N.J.; Voytas, D.F.; Douches, D.S. Geminivirus-mediated genome editing in potato (Solanum tuberosum L.) using sequence-specific nucleases. Front. Plant Sci. 2016, 7, 1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gammoudi, N.; Pedro, T.S.; Ferchichi, A.; Gisbert, C. Improvement of regeneration in pepper: A recalcitrant species. In Vitro Cell. Dev. Biol. Plant 2018, 54, 145–153. [Google Scholar] [CrossRef]
- Xu, J.; Kang, B.C.; Naing, A.H.; Bae, S.J.; Kim, J.S.; Kim, H.; Kim, C.K. CRISPR/Cas9-mediated editing of 1-aminocyclopropane-1-carboxylate oxidase1 enhances Petunia flower longevity. Plant Biotechnol. J. 2020, 18, 287–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, K.; Jang, S.; Lee, J.H.; Lee, D.G.; Kwon, J.K.; Kang, B.C. A MYB transcription factor is a candidate to control pungency in Capsicum annuum. Theor. Appl. Genet. 2019, 132, 1235–1246. [Google Scholar] [CrossRef] [PubMed]
- Pickersgill, B. Genetic resources and breeding of Capsicum spp. Euphytica 1997, 96, 129–133. [Google Scholar] [CrossRef]
- Li, D.; Zhao, K.; Xie, B.; Zhang, B.; Luo, K. Establishment of a highly efficient transformation system for pepper (Capsicum annuum L.). Plant Cell Rep. 2003, 21, 785–788. [Google Scholar] [CrossRef]
- Lee, Y.H.; Kim, H.S.; Kim, J.Y.; Jung, M.; Park, Y.S.; Lee, J.S.; Choi, S.H.; Her, N.H.; Lee, J.H.; Hyung, N.I.; et al. A new selection method for pepper transformation: Callus-mediated shoot formation. Plant Cell Rep. 2004, 23, 50–58. [Google Scholar] [CrossRef]
- Kumar, R.V.; Sharma, V.K.; Chattopadhyay, B.; Chakraborty, S. An improved plant regeneration and Agrobacterium-mediated transformation of red pepper (Capsicum annuum L.). Physiol. Mol. Biol. Plants 2012, 18, 357–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bagga, S.; Lucero, Y.; Apodaca, K.; Rajapakse, W.; Lujan, P.; Ortega, J.L.; Sengupta-Gopalan, C. Chile (Capsicum annuum) plants transformed with the RB gene from Solanum bulbocastanum are resistant to Phytophthora capsici. PLoS ONE 2019, 14, e0223213. [Google Scholar] [CrossRef]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A programmable dual RNA-guided DNA endonuclease in adaptive bacterial immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef]
- Zetsche, B.; Gootenberg, J.S.; Abudayyeh, O.O.; Slaymaker, I.M.; Makarova, K.S.; Essletzbichler, P.; Volz, S.E.; Joung, J.; van der Oost, J.; Regev, A.; et al. Cpf1 is a single RNA-guided endonuclease of a class 2 CRISPR-Cas system. Cell 2015, 163, 759–771. [Google Scholar] [CrossRef] [Green Version]
- Gaudelli, N.M.; Komor, A.C.; Rees, H.A.; Packer, M.S.; Badran, A.H.; Bryson, D.I.; Liu, D.R. Programmable base editing of A•T to G•C in genomic DNA without DNA cleavage. Nature 2017, 551, 464–471. [Google Scholar] [CrossRef]
- Anzalone, A.V.; Randolph, P.B.; Davis, J.R.; Sousa, A.A.; Koblan, L.W.; Levy, J.M.; Chen, P.J.; Wilson, C.; Newby, G.A.; Raguram, A.; et al. Search-and-replace genome editing without double-strand breaks or donor DNA. Nature 2019, 576, 149–157. [Google Scholar] [CrossRef]
- Holsters, M.; Silva, B.; Van Vliet, F.; Genetello, C.; De Block, M.; Dhaese, P.; Depicker, A.; Inzé, D.; Engler, G.; Villarroel, R.; et al. The functional organization of the nopaline A. tumefaciens plasmid pTiC58. Plasmid 1980, 3, 212–230. [Google Scholar] [CrossRef]
- Hoekema, A.; Hirsch, P.R.; Hooykaas, P.J.J.; Schilperoort, R.A. A binary plant vector strategy based on separation of vir- and T-region of the Agrobacterium tumefaciens Ti-plasmid. Nature 1983, 303, 179–180. [Google Scholar] [CrossRef]
- Hood, E.E.; Helmer, G.L.; Fraley, R.T.; Chilton, M.D. The hypervirulence of Agrobacterium tumefaciens A281 is encoded in a region of pTiBo542 outside of T-DNA. J. Bacteriol. 1986, 168, 1291–1301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hood, E.E.; Gelvin, S.B.; Melchers, L.S.; Hoekema, A. New Agrobacterium helper plasmids for gene transfer to plants. Transgenic Res. 1993, 2, 208–218. [Google Scholar] [CrossRef]
- Hellens, R.; Mullineaux, P.; Klee, H. Technical focus: A guide to Agrobacterium binary Ti vectors. Trends Plant Sci. 2000, 5, 446–451. [Google Scholar] [CrossRef]
- El-Nagar, M.M. Somatic embryogenesis of pepper (Capsicum annuum L.) and regeneration of transgenic plants after Agrobacterium-mediated transformation. J. Appl. Sci. Res. 2012, 8, 5550–5563. [Google Scholar]
- Mihálka, V.; Fári, M.; Szász, A.; Balázs, E.; Nagy, I. Optimized protocols for efficient plant regeneration and gene transfer in pepper (Capsicum annuum L.). J. Plant Biotechnol. 2000, 2, 143–149. [Google Scholar]
- Kim, H.; Kim, S.T.; Ryu, J.; Choi, M.K.; Kweon, J.; Kang, B.C.; Ahn, H.M.; Bae, S.; Kim, J.; Kim, J.S.; et al. A simple, flexible and high-throughput cloning system for plant genome editing via CRISPR-Cas system. J. Integr. Plant Biol. 2016, 58, 705–712. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Lim, J. Leaf-induced callus formation in two cultivars: Hot pepper ’CM334’ and bell pepper ‘Dempsey’. Plant Signal. Behav. 2019, 14, 1604016. [Google Scholar] [CrossRef] [PubMed]
- Hwang, H.H.; Wu, E.T.; Liu, S.Y.; Chang, S.C.; Tzeng, K.C.; Kado, C.I. Characterization and host range of five tumorigenic Agrobacterium tumefaciens strains and possible application in plant transient transformation assays. Plant Pathol. 2013, 62, 1384–1397. [Google Scholar] [CrossRef]
- Dabauza, M.; Peña, L. Response of sweet pepper (Capsicum annuum L.) genotypes to Agrobacterium tumefaciens as a means of selecting proper vectors for genetic transformation. J. Hortic. Sci. Biotechnol. 2003, 78, 39–45. [Google Scholar] [CrossRef]
- Gelvin, S.B. Agrobacterium transformation of Arabidopsis thaliana roots: A quantitative assay. Methods Mol. Biol. 2006, 343, 105–114. [Google Scholar] [CrossRef]
- Olhoft, P.M.; Donovan, C.M.; Somers, D.A. Soybean (Glycine max) transformation using mature cotyledonary node explants. Methods Mol. Biol. 2006, 343, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.S.; Hwang, B.K. The pepper MLO gene, CaMLO2, is involved in the susceptibility cell-death response and bacterial and oomycete proliferation. Plant J. 2012, 72, 843–855. [Google Scholar] [CrossRef]
- Lane, R.P.; McCarter, S.M.; Kuhn, C.W.; Deom, C.M. ’Dempsey’, a virus- and bacterial spot-resistant bell pepper. Hort. Sci. 1997, 32, 333–334. [Google Scholar] [CrossRef] [Green Version]
- Porebski, S.; Bailey, L.G.; Baum, B.R. Modification of a CTAB DNA extraction protocol for plants containing high polysaccharide and polyphenol components. Plant Mol. Biol. Rep. 1997, 15, 8–15. [Google Scholar] [CrossRef]
- Park, J.; Lim, K.; Kim, J.S.; Bae, S. Cas-analyzer: An online tool for assessing genome editing results using NGS data. Bioinformatics 2017, 33, 286–288. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | Dempsey | CM334 | |||||
|---|---|---|---|---|---|---|---|
| Agrobacterium | Number of PCR-Applied Calli among PPT Selected | Number of PCR-Positive Calli | Percentage of both Positives (PPT and PCR) | Number of PCR-Applied Calli among PPT Selected | Number of PCR-Positive Calli | Percentage of both Positives (PPT and PCR) | |
| AGL1 | 29 | 23 | 79.3 | 37 | 28 | 75.7 | |
| EHA101 | 36 | 22 | 61.1 | 35 | 30 | 85.7 | |
| GV3101 | 37 | 19 | 51.4 | 35 | 33 | 94.3 | |
| Primer | Sequence |
|---|---|
| CaMLO2 F | ATGGCTAAAGAACGGTCGAT |
| CaMLO2 R | ATGGAGCTGGTGTATTGCAT |
| Primary F | TGGGATTCATATCATTGTTGTTG |
| Primary R | CCGAATGTGTCTCAGCCTTT |
| Secondary F | ACACTCTTTCCCTACACGACGCTCTTCCGATCTTGGGATTCATATCATTGTTGTTG |
| Secondary R | ACTGGAGTTCAGAGTGTGCTCTTCCGATCTCCGAATGTGTCTCAGCCTTT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, S.-i.; Kim, H.-B.; Jeon, H.-J.; Kim, H. Agrobacterium-Mediated Capsicum annuum Gene Editing in Two Cultivars, Hot Pepper CM334 and Bell Pepper Dempsey. Int. J. Mol. Sci. 2021, 22, 3921. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22083921
Park S-i, Kim H-B, Jeon H-J, Kim H. Agrobacterium-Mediated Capsicum annuum Gene Editing in Two Cultivars, Hot Pepper CM334 and Bell Pepper Dempsey. International Journal of Molecular Sciences. 2021; 22(8):3921. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22083921
Chicago/Turabian StylePark, Sung-il, Hyun-Bin Kim, Hyun-Ji Jeon, and Hyeran Kim. 2021. "Agrobacterium-Mediated Capsicum annuum Gene Editing in Two Cultivars, Hot Pepper CM334 and Bell Pepper Dempsey" International Journal of Molecular Sciences 22, no. 8: 3921. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22083921