
Anatomy and Histochemistry of Seed Coat Development of Wild (Pisum sativum subsp. elatius (M. Bieb.) Asch. et Graebn. and Domesticated Pea (Pisum sativum subsp. sativum L.)



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
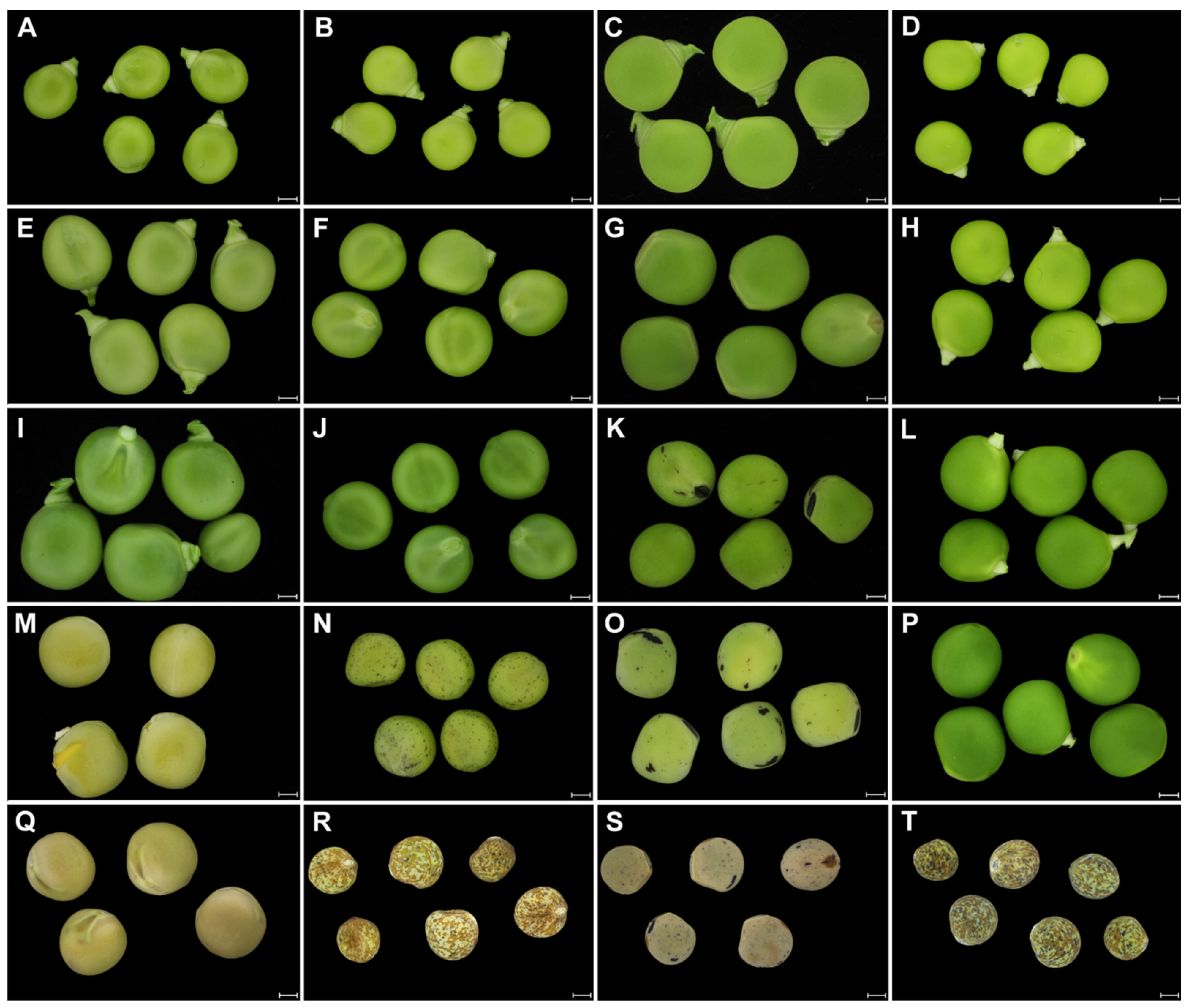
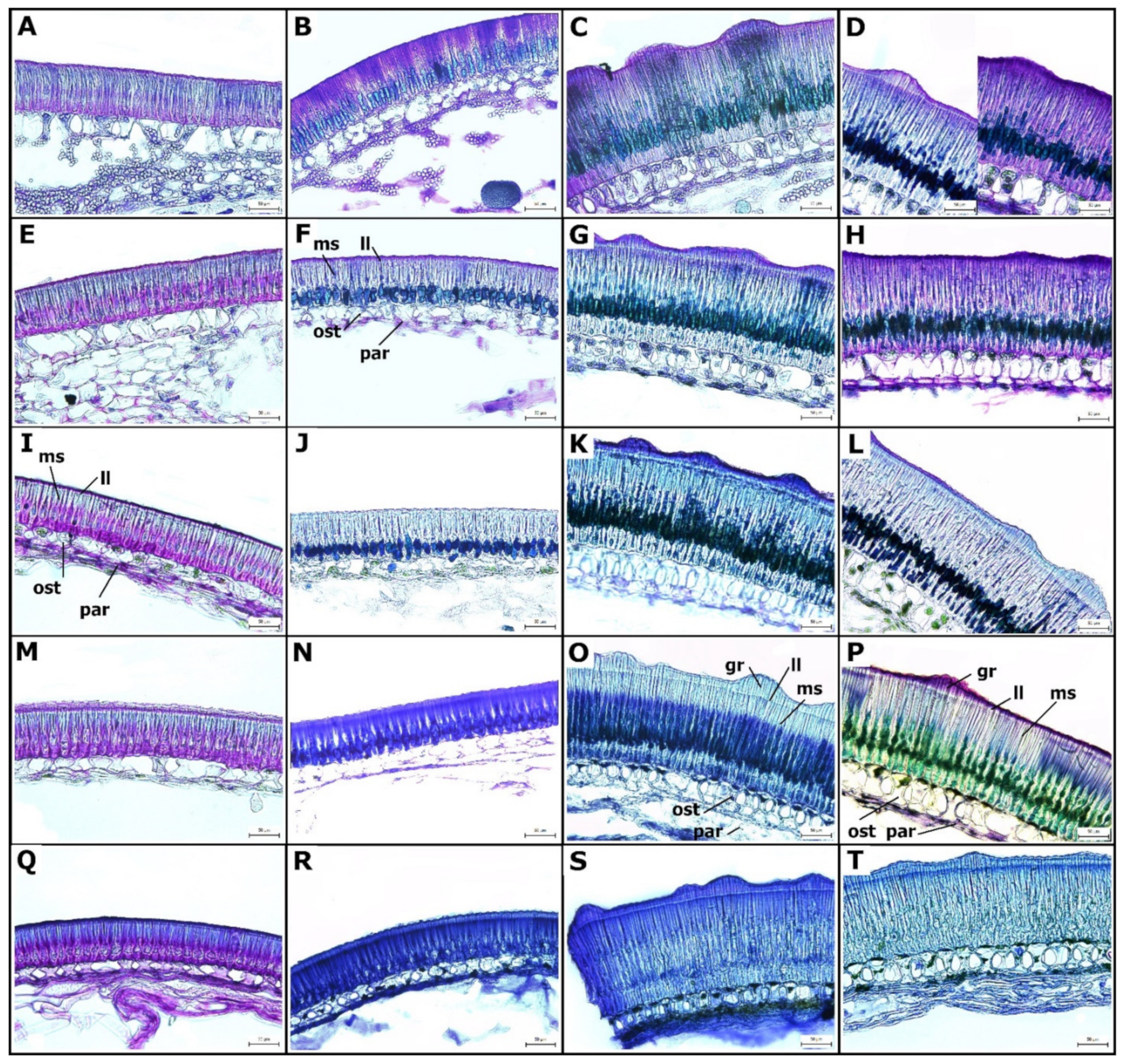
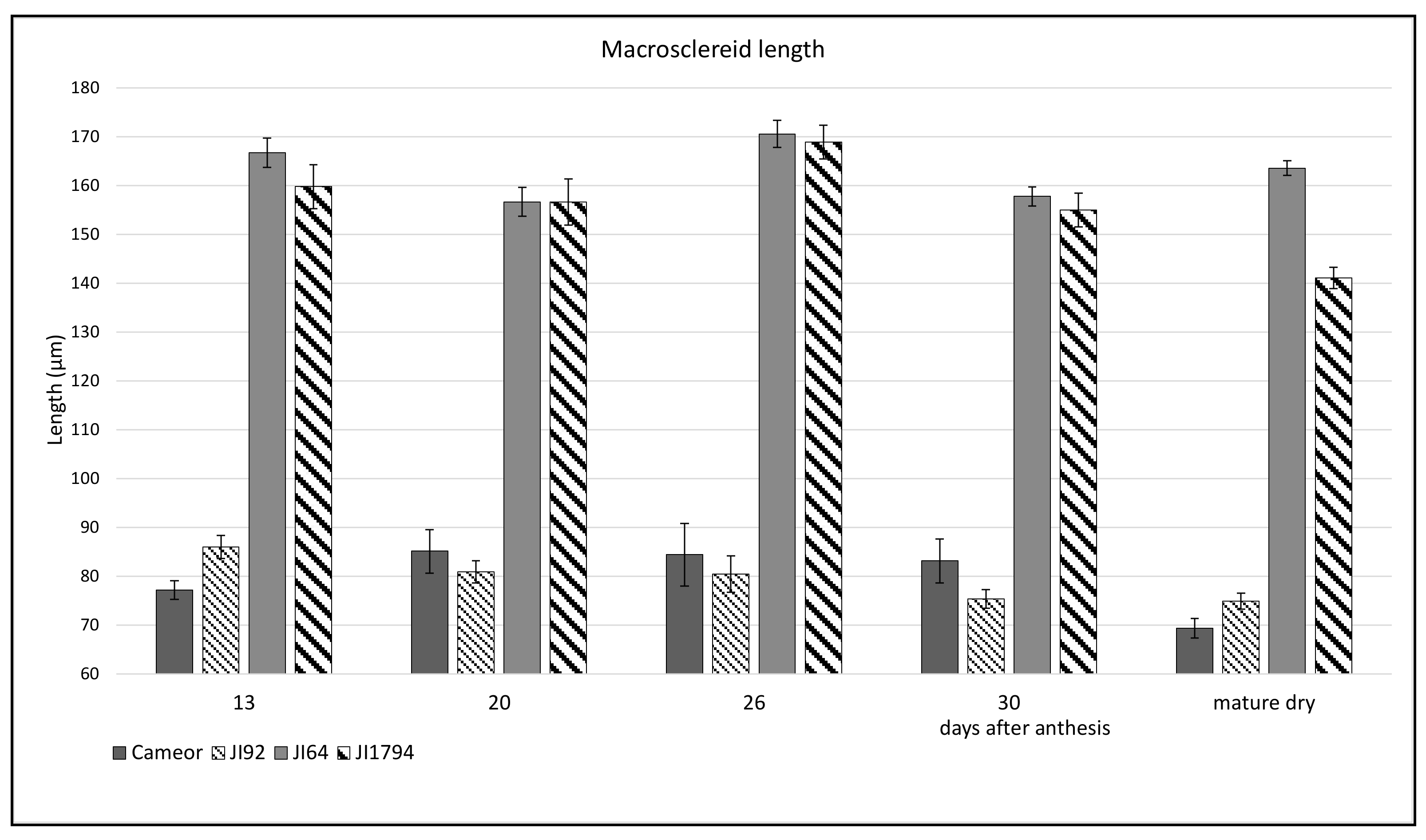
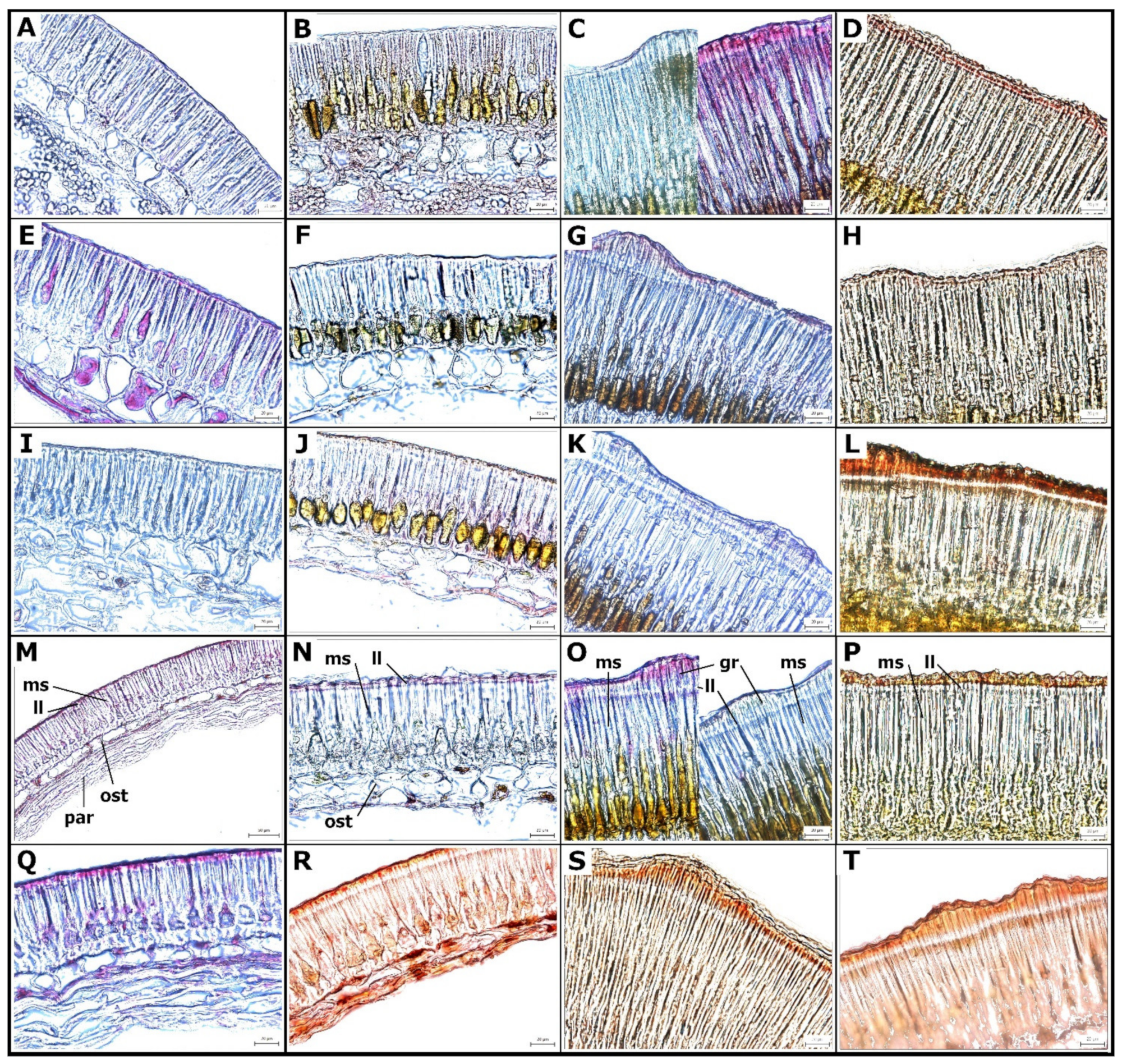
2. Results
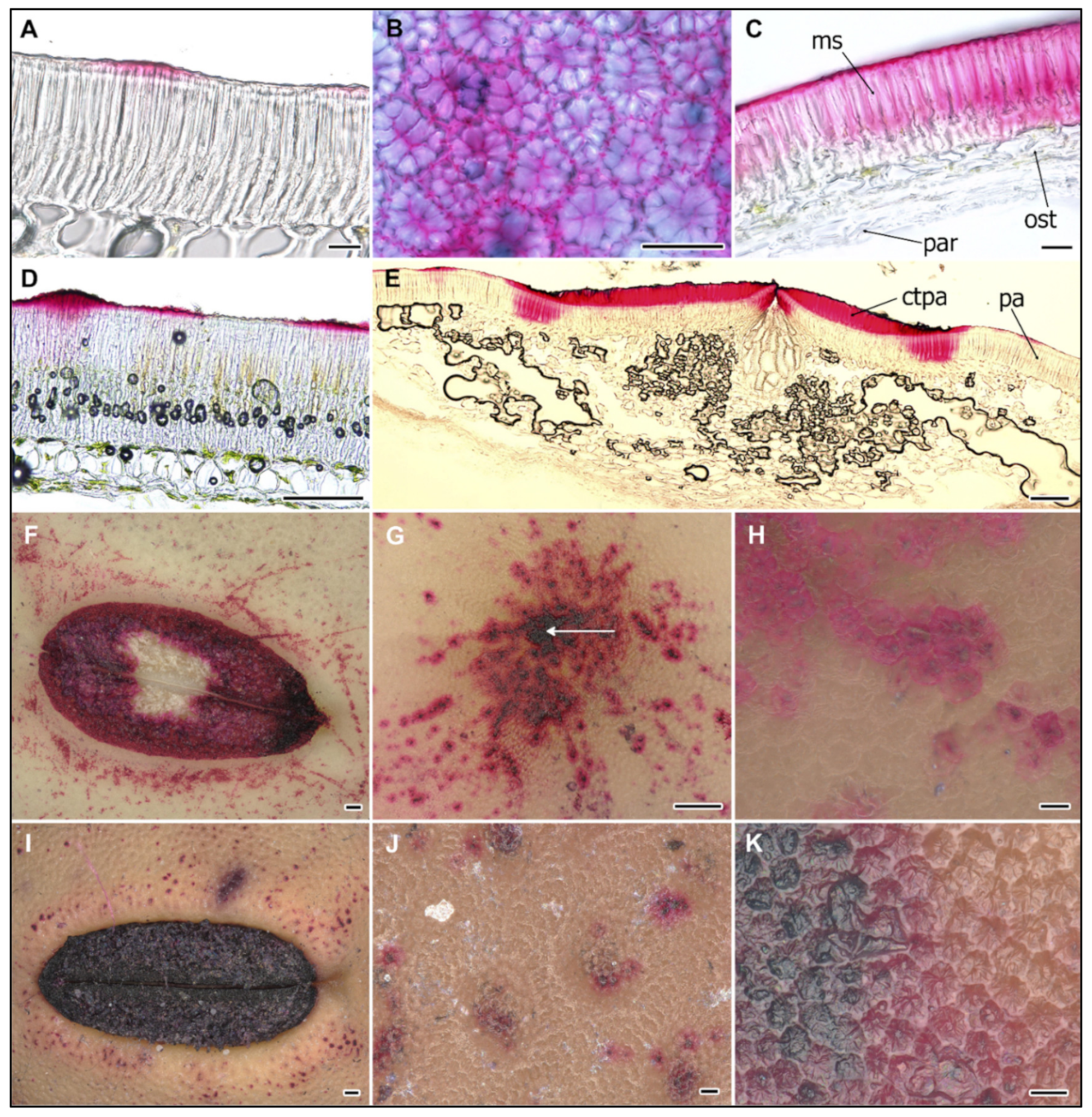
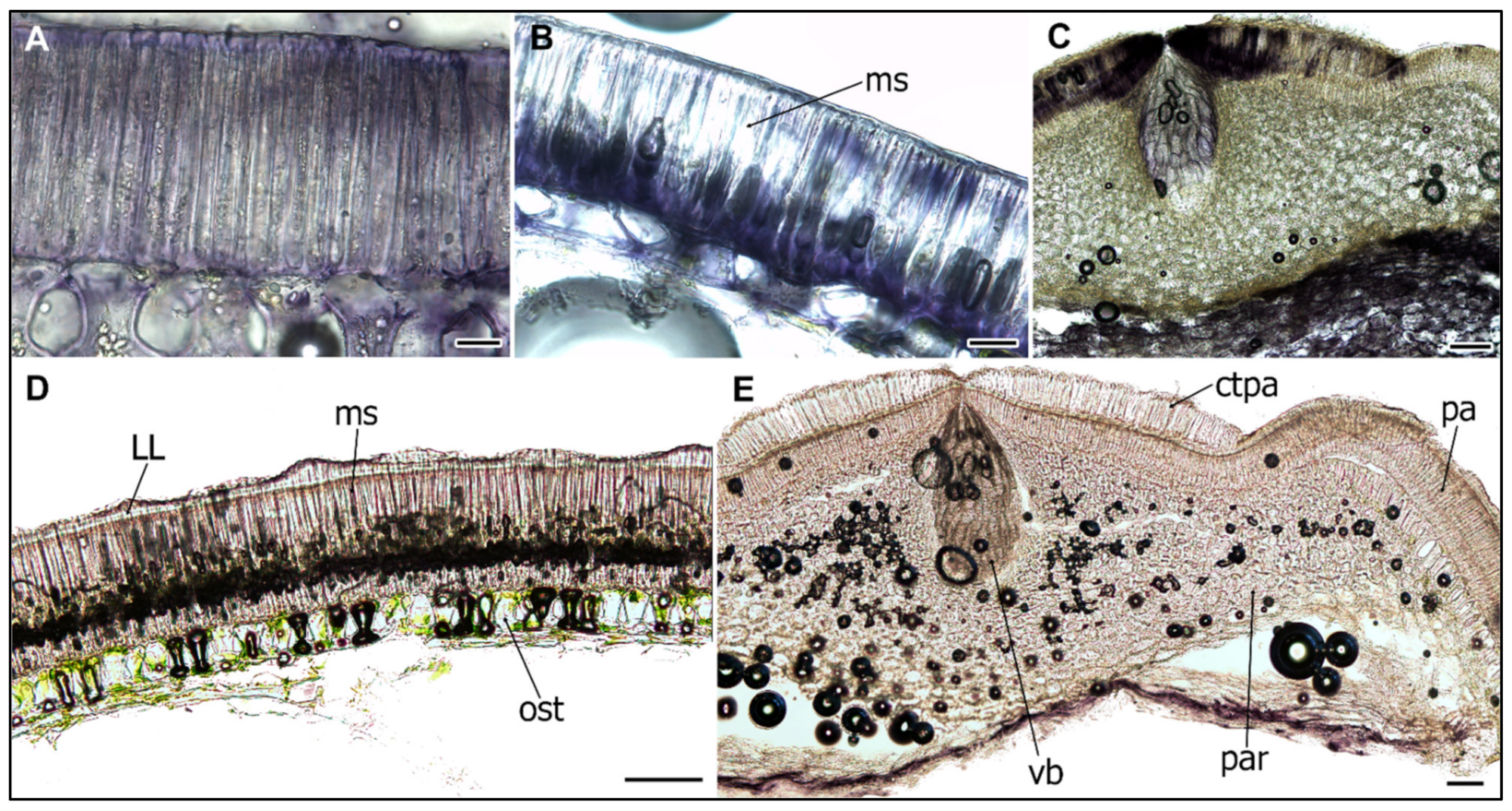
2.1. Structure of Wild and Cultivated Pea Seed Coat Differ during Development
2.2. Cell Wall Lipids Staining with Sudan Red
2.3. Ruthenium Red Staining of Polyanionic Substances
2.4. Peroxidase Activity during the SC Development
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Plant Material
5.2. Developmental Stages Labeling
5.3. Anatomical and Histochemical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bewley, J.D.; Bradford, K.; Hilhorst, H.; Nonogaki, H. Seeds: Physiology of Development, Germination and Dormancy, 3rd ed.; Springer: New York, NY, USA, 2013. [Google Scholar]
- Baroux, C.; Grossniklaus, U. Seeds-an evolutionary innovation underlying reproductive success in flowering plants. Curr. Top. Dev. Biol. 2019, 131, 605–642. [Google Scholar] [CrossRef]
- Linkies, A.; Graeber, K.; Knight, C.; Leubner-Metzger, G. The evolution of seeds. New Phytol. 2010, 186, 817–831. [Google Scholar] [CrossRef] [PubMed]
- Matilla, A.J. Seed coat formation: Its evolution and regulation. Seed Sci. Res. 2019, 29, 215–226. [Google Scholar] [CrossRef]
- Coen, O.; Magnani, E. Seed coat thickness in the evolution of Angiosperms. Cell. Mol. Life Sci. 2018, 75, 2509–2518. [Google Scholar] [CrossRef] [Green Version]
- Haughn, G.; Chaudhury, A. Genetic analysis of seed coat development in Arabidopsis. Trends Plant Sci. 2005, 10, 472–477. [Google Scholar] [CrossRef]
- Coen, O.; Fiume, E.; Xu, W.; De Vos, D.; Lu, J.; Pechoux, C.; Lepiniec, L.; Magnani, E. Developmental patterning of the sub-epidermal integument cell layer in Arabidopsis seeds. Development 2017, 144, 1490–1497. [Google Scholar] [CrossRef] [Green Version]
- Debeaujon, I.; Léon-Kloosterziel, K.M.; Koornneef, M. Influence of the testa on seed dormancy, germination, and longevity in Arabidopsis. Plant Physiol. 2000, 122, 403–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bentsink, L.; Koornneef, M. Seed Dormancy and Germination. Arab. Book 2008, 6, e0119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinbrecher, T.; Leubner-Metzger, G. Tissue and cellular mechanics of seeds. Curr. Opin. Genet. Dev. 2018, 51, 1–10. [Google Scholar] [CrossRef]
- Verdier, J.; Dessaint, F.; Schneider, C.; Abirached-Darmency, M. A combined histology and transcriptome analysis unravels novel questions on Medicago truncatula seed coat. J. Exp. Bot. 2013, 64, 459–470. [Google Scholar] [CrossRef]
- Radchuk, V.; Borisjuk, L. Physical, metabolic and developmental functions of the seed coat. Front. Plant Sci. 2014, 5, 510. [Google Scholar] [CrossRef] [Green Version]
- Smýkal, P.; Vernoud, V.; Blair, M.W.; Soukup, A.; Thompson, R.D. The role of the testa during development and in establishment of dormancy of the legume seed. Front. Plant Sci. 2014, 5, 351. [Google Scholar]
- Gunn, C.R. Seed Topography in the Fabaceae. Seed Sci. Technol. 1981, 9, 733–737. [Google Scholar]
- Baskin, C.; Baskin, J.M. Seed Ecology, Biogeography, and Evolution of Dormancy and Germination, 2nd ed.; Academic Press; Elsevier: London, UK, 2014. [Google Scholar]
- Moïse, J.A.; Han, S.; Gudynaitę-Savitch, L.; Johnson, D.A.; Miki, B.L.A. Seed Coats: Structure, development, composition, and biotechnology. In Vitro Cell. Dev. Biol. Plant 2005, 41, 620–644. [Google Scholar] [CrossRef]
- Gallardo, K.; Firnhaber, C.; Zuber, H.; Héricher, D.; Belghazi, M.; Henry, C.; Küster, H.; Thompson, R. A Combined proteome and transcriptome analysis of developing Medicago truncatula seeds: Evidence for metabolic specialization of maternal and filial tissues. Mol. Cell. Proteom. 2007, 6, 2165–2179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.L.; Grusak, M.A. Structure and development of Medicago truncatula pod wall and seed coat. Ann. Bot. 2005, 95, 737–747. [Google Scholar] [CrossRef] [Green Version]
- Fu, F.; Zhang, W.; Li, Y.Y.; Wang, H.L. Establishment of the model system between phytochemicals and gene expression profiles in macrosclereid cells of Medicago truncatula. Sci. Rep. 2017, 7, 2580. [Google Scholar] [CrossRef] [PubMed]
- Qutob, D.Q.; Ma, F.M.; Peterson, C.A.P.A.; Bernards, M.A.B.A.; Gijzen, M.G. Structural and permeability properties of the soybean seed coat. Botany 2008, 86, 219–227. [Google Scholar] [CrossRef]
- Miller, S.S.; Bowman, L.-A.A.; Gijzen, M.; Miki, B.L.A. Early development of the seed coat of soybean (Glycine max). Ann. Bot. 1999, 84, 297–304. [Google Scholar] [CrossRef] [Green Version]
- Miller, S.S.; Jin, Z.; Schnell, J.A.; Romero, M.C.; Brown, D.C.W.; Johnson, D.A. Hourglass Cell Development in the Soybean Seed Coat. Ann. Bot. 2010, 106, 235–242. [Google Scholar] [CrossRef] [Green Version]
- Ranathunge, K.; Shao, S.; Qutob, D.; Gijzen, M.; Peterson, C.A.; Bernards, M.A. Properties of the soybean seed coat cuticle change during development. Planta 2010, 231, 1171–1188. [Google Scholar] [CrossRef]
- Reeve, R.M. Ontogeny of the sclereids in the integument of Pisum sativum L. Am. J. Bot. 1946, 33, 806–816. [Google Scholar] [CrossRef]
- Spurný, M. Cell wall structure of the epidermal cells of the pea seed coat Pisum sativum L. Preslia 1954, 26, 79–88. [Google Scholar]
- Spurný, M. The light line of the epidermal cells of the pea seed coat Pisum sativum L. Preslia 1954, 26, 139–142. [Google Scholar]
- Spurný, M. Cell wall structure of epidermal cells of the pea seed coat (Pisum sativum L.) studied by microcinematography. Mikroskopie 1963, l8, 272–279. [Google Scholar]
- Spurný, M. Changes in the permeability of the seed coat in connection with the development of suberin adcrustations of the macrosclereids from the seed coat of the pea (Pisum sativum L.). Flora 1964, 154, 547–567. [Google Scholar] [CrossRef]
- Marbach, I.; Mayer, A.M. Permeability of seed coats to water as related to drying conditions and metabolism of phenolics 1. Plant Physiol. 1974, 54, 817–820. [Google Scholar] [CrossRef] [PubMed]
- Marbach, I.; Mayer, A.M. Changes in catechol oxidase and permeability to water in seed coats of Pisum elatius during seed development and maturation. Plant Physiol. 1975, 56, 93–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werker, E.; Marbach, I.; Mayer, A.M. Relation between the anatomy of the testa, water permeability and the presence of phenolics in the genus Pisum. Ann. Bot. 1979, 43, 765–771. [Google Scholar] [CrossRef]
- Harris, W.M. On the Development of macrosclereids in seed coats of Pisum sativum L. Am. J. Bot. 1983, 70, 1528–1535. [Google Scholar] [CrossRef]
- Harris, W.M. On the development of osteosclereids in seed coats of Pisum sativum L. New Phytol. 1984, 98, 135–141. [Google Scholar] [CrossRef]
- Van Dongen, J.T.; Ammerlaan, A.M.H.; Wouterlood, M.; Van Aelst, A.C.; Borstlap, A.C. Structure of the developing pea seed coat and the post-phloem transport pathway of nutrients. Ann. Bot. 2003, 91, 729–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, H.; Borisjuk, L.; Wobus, U. Molecular physiology of legume seed development. Annu. Rev. Plant Biol. 2005, 56, 253–279. [Google Scholar] [CrossRef] [PubMed]
- Kigel, J.; Rosental, L.; Fait, A. Seed physiology and germination of grain legumes. In Plant Breeding; Handbook of Plant Breeding; De Ron, A.M., Ed.; Springer: New York, NY, USA, 2015; pp. 327–363. [Google Scholar]
- Janská, A.; Pecková, E.; Sczepaniak, B.; Smýkal, P.; Soukup, A. The role of the testa during the establishment of physical dormancy in the pea seed. Ann. Bot. 2019, 123, 815–829. [Google Scholar] [CrossRef] [PubMed]
- Godwin, J.; Raviv, B.; Grafi, G. Dead pericarps of dry fruits function as long-term storage for active hydrolytic enzymes and other substances that affect germination and microbial growth. Plants 2017, 6, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raviv, B.; Aghajanyan, L.; Granot, G.; Makover, V.; Frenkel, O.; Gutterman, Y.; Grafi, G. The dead seed coat functions as a long-term storage for active hydrolytic enzymes. PLoS ONE 2017, 12, e0181102. [Google Scholar] [CrossRef] [Green Version]
- Smýkal, P.; Nelson, M.N.; Berger, J.D.; Von Wettberg, E.J.B. The impact of genetic changes during crop domestication. Agronomy 2018, 8, 119. [Google Scholar] [CrossRef] [Green Version]
- Hradilová, I.; Trněný, O.; Válková, M.; Cechová, M.; Janská, A.; Prokešová, L.; Aamir, K.; Krezdorn, N.; Rotter, B.; Winter, P.; et al. A combined comparative transcriptomic, metabolomic, and anatomical analyses of two key domestication traits: Pod dehiscence and seed dormancy in pea (Pisum sp.). Front. Plant Sci. 2017, 8, 542. [Google Scholar] [CrossRef] [Green Version]
- Blixt, S. Pea. In Handbook of Genetics; King, R.C., Ed.; Plenum Press: New York, NY, USA, 1974. [Google Scholar]
- Soukup, A. Selected simple methods of plant cell wall histochemistry and staining for light microscopy. In Plant Cell Morphogenesis: Methods and Protocols; Methods in Molecular Biology; Cvrčková, F., Žárský, V., Eds.; Springer: New York, NY, USA, 2019; pp. 27–42. [Google Scholar]
- Watson, D.P. Structure of the testa and its relation to germination in the Papilionaceae tribes Trifoliae and Lotea. Ann. Bot. 1948, 12, 385–409. [Google Scholar] [CrossRef]
- Corner, E.J.H. The Leguminous Seed. Phytomorphology 1951, 1, 117–150. [Google Scholar]
- Rolston, M.P. Water Impermeable Seed Dormancy. Bot. Rev. 1978, 44, 365–396. [Google Scholar] [CrossRef]
- Werker, E. Seed dormancy as explained by anatomy of embryo envelopes. Isr. J. Bot. 1980, 29, 22–44. [Google Scholar]
- Nadeau, C.D.; Ozga, J.A.; Kurepin, L.V.; Jin, A.; Pharis, R.P.; Reinecke, D.M. Tissue-specific regulation of gibberellin biosynthesis in developing pea seeds. Plant Physiol. 2011, 156, 897–912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chai, M.; Zhou, C.; Molina, I.; Fu, C.; Nakashima, J.; Li, G.; Zhang, W.; Park, J.; Tang, Y.; Jiang, Q.; et al. A class II KNOX gene, KNOX4, controls seed physical dormancy. Proc. Natl. Acad. Sci. USA 2016, 113, 6997–7002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCartney, L.; Knox, J.P. Regulation of pectic polysaccharide domains in relation to cell development and cell properties in the pea testa. J. Exp. Bot. 2002, 53, 707–713. [Google Scholar] [CrossRef] [Green Version]
- Cechová, M.; Válková, M.; Hradilová, I.; Janská, A.; Soukup, A.; Smýkal, P.; Bednář, P. Towards better understanding of pea seed dormancy using Laser Desorption/Ionization Mass Spectrometry. Int. J. Mol. Sci. 2017, 18, 2196. [Google Scholar] [CrossRef] [Green Version]
- Cavazza, L. Recherches sur l’impermeabilite des grains dures chez les legumineuses. Ber. Schweiz Bot. Ges. 1950, 60, 596–610. [Google Scholar]
- Hamly, D.H. Softening of the Seeds of Melilotus alba. Bot. Gaz. 1932, 93, 345–375. [Google Scholar] [CrossRef]
- Bevilacqua, L.R.; Roti-Michelozzi, G.; Modenesi, P. The watertight dormancy of Melilotus alba seeds: Further observations on the palisade cell wall. Can. J. Bot. 1989, 67, 3453–3456. [Google Scholar] [CrossRef]
- Martens, H.; Jakobsen, H.B.; Lyshede, O.B. Development of the strophiole in seeds of white clover (Trifolium repens L.). Seed Sci. Res. 1995, 5, 171–176. [Google Scholar] [CrossRef]
- Manning, J.C.; van Staden, J. The development and ultrastructure of the testa and tracheid bar in Erythrina lysistemon Hutch. (Leguminosae: Papilionoideae). Protoplasma 1985, 129, 157–167. [Google Scholar] [CrossRef]
- De Jong, A.; Koerselman-Kooij, J.W.; Schuurmans, J.A.M.J.; Borstlap, A.C. The mechanism of amino acid efflux from seed coats of developing pea seeds as revealed by uptake experiments. Plant Physiol. 1997, 114, 731–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Souza, F.H.D.D.; Marcos-Filho, J. The seed coat as a modulator of seed-environment relationships in Fabaceae. Brazil. J. Bot. 2001, 24, 365–375. [Google Scholar] [CrossRef] [Green Version]
- Egley, G.H.; Paul, R.N.; Duke, S.O.; Vaughn, K.C. Peroxidase involvement in lignification in water-impermeable seed coats of weedy leguminous and malvaceous species. Plant Cell Environ. 1985, 8, 253–260. [Google Scholar] [CrossRef]
- Egley, G.H.; Paul, R.N.; Vaughn, K.C.; Duke, S.O. Role of peroxidase in the development of water-impermeable seed coats in Sida spinosa L. Planta 1983, 157, 224–232. [Google Scholar] [CrossRef]
- Gijzen, M.; Van Huystee, R.; Buzzell, R.I. Soybean seed coat peroxidase (A comparison of high-activity and low-activity genotypes). Plant Physiol. 1993, 103, 1061–1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmitz, N.; van Huystee, R.; Gijzen, M. Characterization of anionic soybean (Glycine max) seed coat peroxidase. Can. J. Bot. 1997, 75, 1336–1341. [Google Scholar] [CrossRef]
- Serrato-Valenti, G.; Cornara, L.; Ferrando, M.; Modenesi, P. Structural and histochemical features of Stylosanthes scabra (Leguminosae; Papilionoideae) seed coat as related to water entry. Can. J. Bot. 1993, 71, 834–840. [Google Scholar] [CrossRef]
- Hyde, E.O.C. The Function of the hilum in some Papilionaceae in relation to the ripening of the seed and the permeability of the testa. Ann. Bot. 1954, 18, 241–256. [Google Scholar] [CrossRef]
- Hanma, A.; Denna, D. Moisture movement in impermeable snap bean seed. Quart. Bull. Mich. Agr. Exp. Sta. 1962, 44, 726–730. [Google Scholar]
- Lush, W.M.; Evans, L.T. The seed coats of cowpeas and other grain legumes: Structure in relation to function. Field Crops Res. 1980, 3, 267–286. [Google Scholar] [CrossRef]
- Gillikin, J.; Graham, J.S. Purification and developmental analysis of the major anionic peroxidase from the seed coat of Glycine max. Plant Physiol. 1991, 96, 214–220. [Google Scholar] [CrossRef] [Green Version]
- Chou, Y.E.; Schuetz, M.; Hoffmann, N.; Watanabe, Y.; Sibout, R.; Samuels, A.L. Distribution, mobility, and anchoring of lignin-related oxidative enzymes in Arabidopsis secondary cell walls. J. Exp. Bot. 2018, 69, 1849–1859. [Google Scholar] [CrossRef] [PubMed]
- Renard, J.; Martínez-Almonacid, I.; Sonntag, A.; Molina, I.; Moya-Cuevas, J.; Bissoli, G.; Muñoz-Bertomeu, J.; Faus, I.; Niñoles, R.; Shigeto, J.; et al. PRX2 and PRX25, Peroxidases regulated by cog1, are involved in seed longevity in Arabidopsis. Plant Cell Environ. 2020, 43, 315–326. [Google Scholar] [CrossRef]
- Müller, K.; Linkies, A.; Vreeburg, R.A.M.; Fry, S.C.; Krieger-Liszkay, A.; Leubner-Metzger, G. In vivo cell wall loosening by hydroxyl radicals during cress seed germination and elongation growth. Plant Physiol. 2009, 150, 1855–1865. [Google Scholar] [CrossRef] [Green Version]
- Pourcel, L.; Routaboul, J.-M.; Kerhoas, L.; Caboche, M.; Lepiniec, L.; Debeaujon, I. TRANSPARENT TESTA10 encodes a laccase-like enzyme involved in oxidative polymerization of flavonoids in Arabidopsis seed coat. Plant Cell 2005, 17, 2966–2980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renzi, J.P.; Duchoslav, M.; Brus, J.; Hradilová, I.; Pechanec, V.; Václavek, T.; Machalová, J.; Hron, K.; Verdier, J.; Smýkal, P. Physical dormancy release in Medicago truncatula seeds is related to environmental variations. Plants 2020, 9, 503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, T.J. ImageJ for Microscopy. BioTechniques 2007, 43, S25–S30. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zablatzká, L.; Balarynová, J.; Klčová, B.; Kopecký, P.; Smýkal, P. Anatomy and Histochemistry of Seed Coat Development of Wild (Pisum sativum subsp. elatius (M. Bieb.) Asch. et Graebn. and Domesticated Pea (Pisum sativum subsp. sativum L.). Int. J. Mol. Sci. 2021, 22, 4602. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094602
Zablatzká L, Balarynová J, Klčová B, Kopecký P, Smýkal P. Anatomy and Histochemistry of Seed Coat Development of Wild (Pisum sativum subsp. elatius (M. Bieb.) Asch. et Graebn. and Domesticated Pea (Pisum sativum subsp. sativum L.). International Journal of Molecular Sciences. 2021; 22(9):4602. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094602
Chicago/Turabian StyleZablatzká, Lenka, Jana Balarynová, Barbora Klčová, Pavel Kopecký, and Petr Smýkal. 2021. "Anatomy and Histochemistry of Seed Coat Development of Wild (Pisum sativum subsp. elatius (M. Bieb.) Asch. et Graebn. and Domesticated Pea (Pisum sativum subsp. sativum L.)" International Journal of Molecular Sciences 22, no. 9: 4602. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094602