
Proinflammatory Interleukin-33 Induces Dichotomic Effects on Cell Proliferation in Normal Gastric Epithelium and Gastric Cancer

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. IL-33 Is Expressed in Gastric Cell Lines and Its Expression Is Increased in Chronic Gastritis
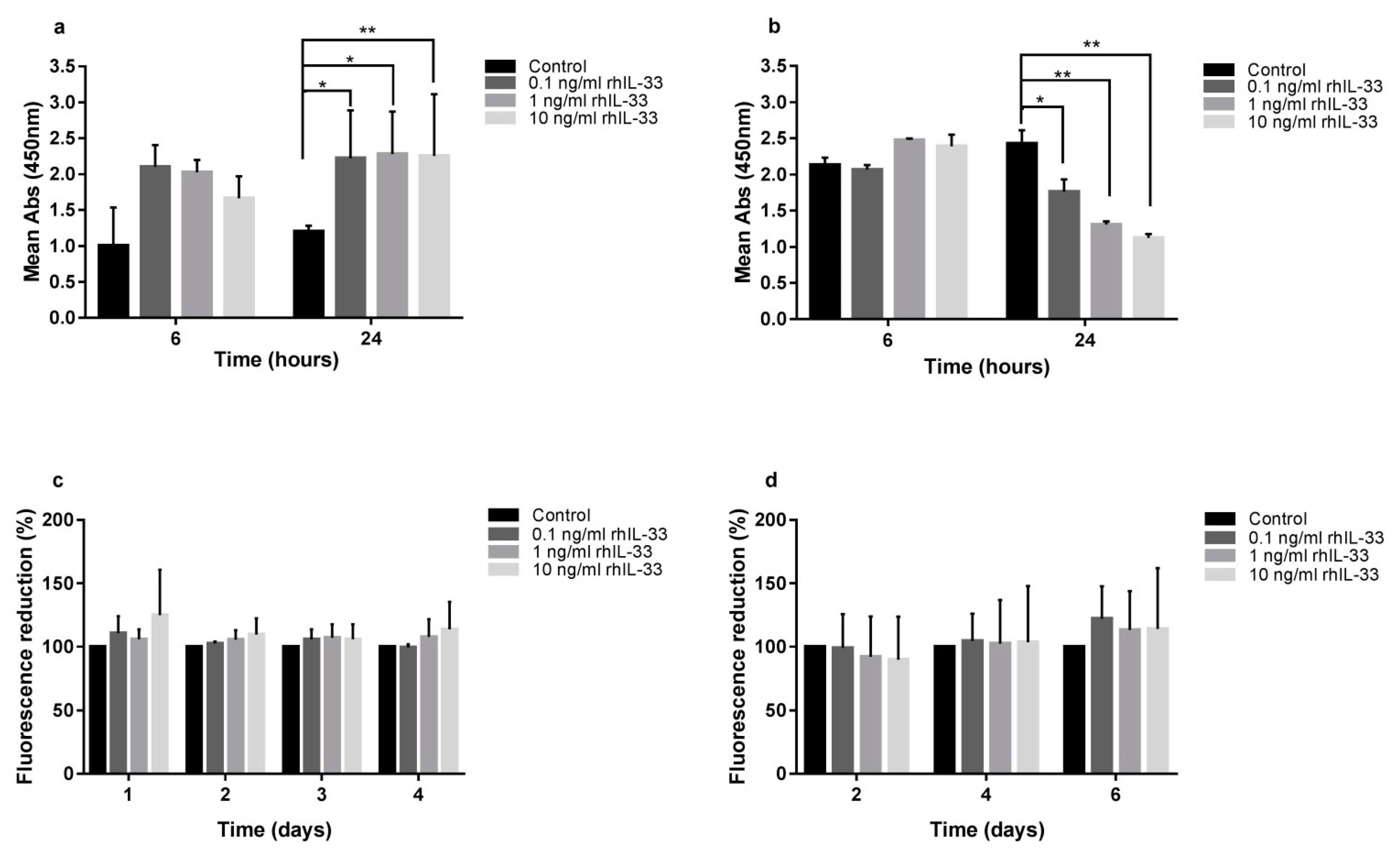
2.2. IL-33 Differentially Modulates Gastric Cell Proliferation and Migration
2.3. IL-33 Induces Apoptosis in Gastric Cancer Cell Lines, but Not in Non-Neoplastic Cells
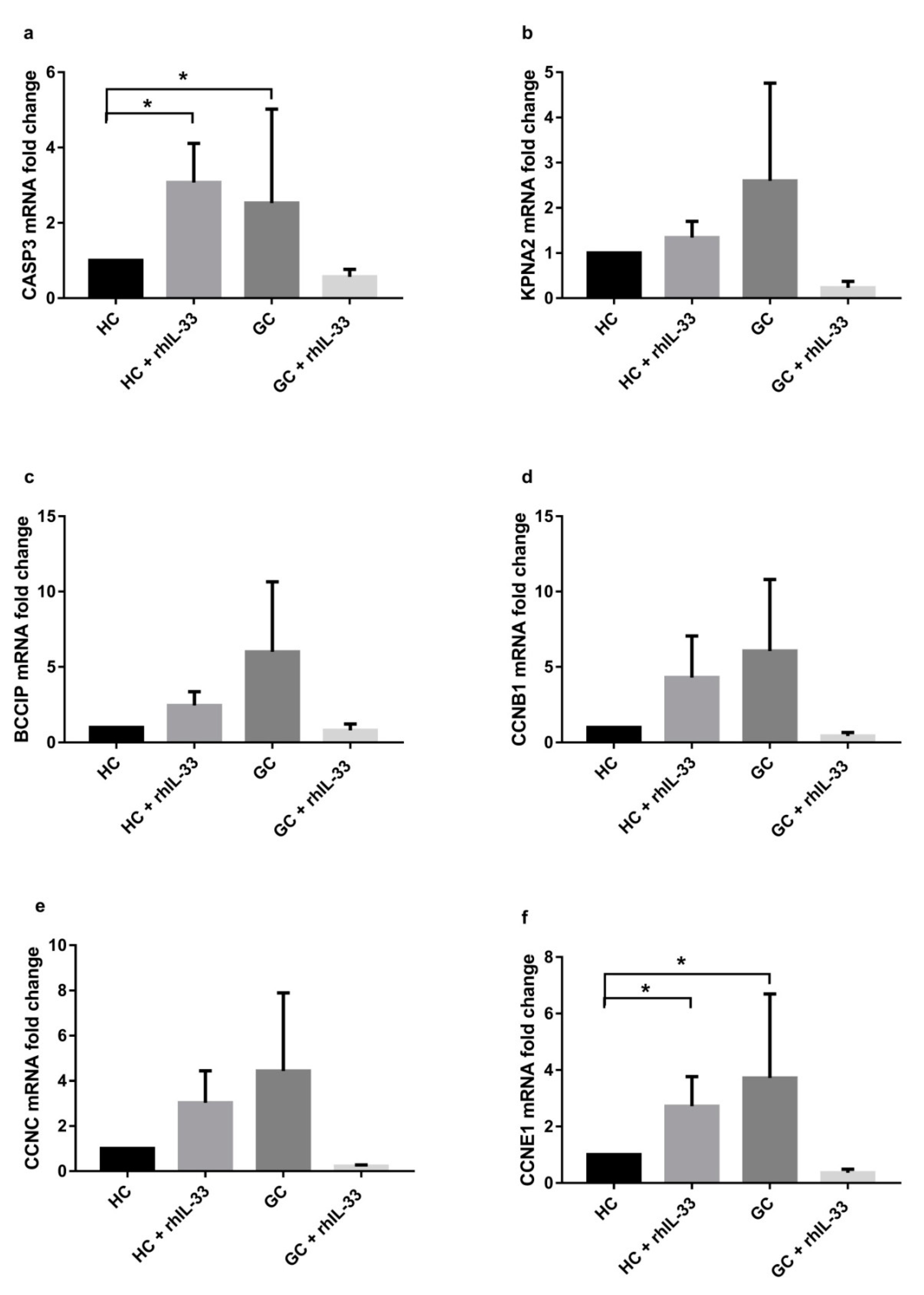
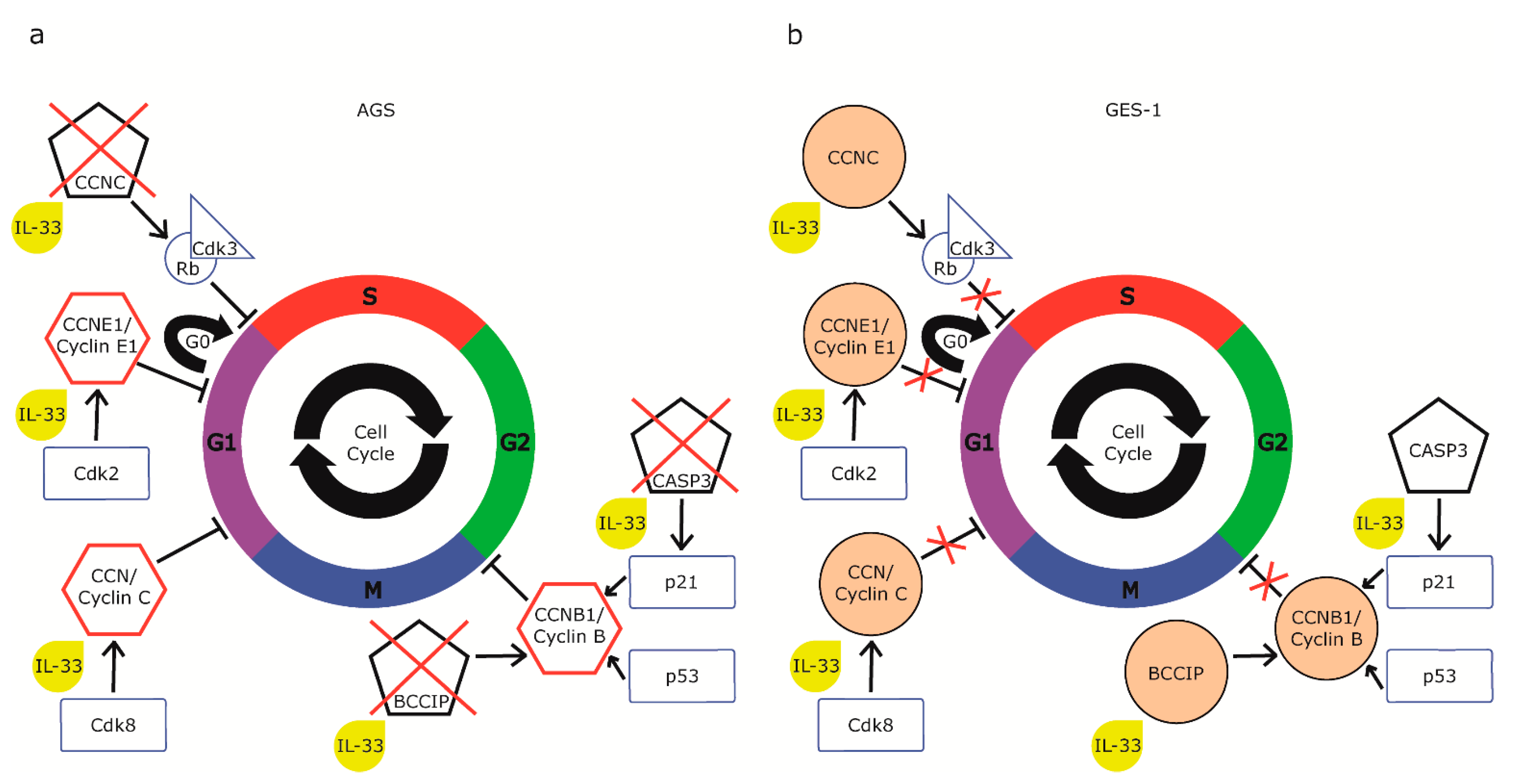
2.4. IL-33 Acts on Cell Cycle Gene Expression in Gastric Cell Lines
2.5. IL-33-Stimulation of Human Gastric Biopsies and Surgical Specimens Modifies Cell Cycle Gene Expression
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Treatments
4.2. Wound Healing
4.3. Evaluation of Cell Proliferation
4.4. Cell Cycle Analysis by Propidium Iodine (PI) Staining
4.5. Analysis of Cell Cycle by RT2 Profiler PCR Array for Cell Cycle
4.6. Evaluation of Caspase-3 and Caspase-7 Activities
4.7. Annexin V Apoptosis Detection
4.8. Gastric Tissue Sample
4.9. Reverse Transcription and Real-Time PCR
4.10. Statistical Analysis
4.11. Ethical Committee and Informed Consent Statement
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carcas, L.P. Gastric cancer review. J. Carcinog 2014, 13, 14. [Google Scholar] [CrossRef]
- Machlowska, J.; Baj, J.; Sitarz, M.; Maciejewski, R.; Sitarz, R. Gastric Cancer: Epidemiology, Risk Factors, Classification, Genomic Characteristics and Treatment Strategies. Int. J. Mol. Sci. 2020, 21, 4012. [Google Scholar] [CrossRef]
- Nagini, S. Carcinoma of the stomach: A review of epidemiology, pathogenesis, molecular genetics and chemoprevention. World J. Gastrointest. Oncol. 2012, 4, 156–169. [Google Scholar] [CrossRef]
- Wasmer, M.H.; Krebs, P. The Role of IL-33-Dependent Inflammation in the Tumor Microenvironment. Front. Immunol. 2016, 7, 682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petersen, C.P.; Meyer, A.R.; De Salvo, C.; Choi, E.; Schlegel, C.; Petersen, A.; Engevik, A.C.; Prasad, N.; Levy, S.E.; Peebles, R.S.; et al. A signalling cascade of IL-33 to IL-13 regulates metaplasia in the mouse stomach. Gut 2017, 67, 805–817. [Google Scholar] [CrossRef]
- Schmitz, J.; Owyang, A.; Oldham, E.; Song, Y.; Murphy, E.; McClanahan, T.K.; Zurawski, G.; Moshrefi, M.; Qin, J.; Li, X.; et al. IL-33, an interleukin-1-like cytokine that signals via the IL-1 receptor-related protein ST2 and induces T helper type 2-associated cytokines. Immunity 2005, 23, 479–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tu, S.; Bhagat, G.; Cui, G.; Takaishi, S.; Kurt-Jones, E.A.; Rickman, B.; Betz, K.S.; Penz-Oesterreicher, M.; Bjorkdahl, O.; Fox, J.G.; et al. Overexpression of interleukin-1beta induces gastric inflammation and cancer and mobilizes myeloid-derived suppressor cells in mice. Cancer Cell 2008, 14, 408–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fagundes, C.T.; Amaral, F.A.; Souza, A.L.; Vieira, A.T.; Xu, D.; Liew, F.Y.; Souza, D.G.; Teixeira, M.M. ST2, an IL-1R family member, attenuates inflammation and lethality after intestinal ischemia and reperfusion. J. Leukoc. Biol. 2007, 81, 492–499. [Google Scholar] [CrossRef]
- Lopetuso, L.R.; De Salvo, C.; Pastorelli, L.; Rana, N.; Senkfor, H.N.; Petito, V.; Di Martino, L.; Scaldaferri, F.; Gasbarrini, A.; Cominelli, F.; et al. IL-33 promotes recovery from acute colitis by inducing miR-320 to stimulate epithelial restitution and repair. Proc. Natl. Acad. Sci. USA 2018, 115, E9362–E9370. [Google Scholar] [CrossRef] [Green Version]
- Monticelli, L.A.; Osborne, L.C.; Noti, M.; Tran, S.V.; Zaiss, D.M.; Artis, D. IL-33 promotes an innate immune pathway of intestinal tissue protection dependent on amphiregulin-EGFR interactions. Proc. Natl. Acad. Sci. USA 2015, 112, 10762–10767. [Google Scholar] [CrossRef] [Green Version]
- Miller, A.M. Role of IL-33 in inflammation and disease. J. Inflamm. 2011, 8, 22. [Google Scholar] [CrossRef] [Green Version]
- Pastorelli, L.; Garg, R.R.; Hoang, S.B.; Spina, L.; Mattioli, B.; Scarpa, M.; Fiocchi, C.; Vecchi, M.; Pizarro, T.T. Epithelial-derived IL-33 and its receptor ST2 are dysregulated in ulcerative colitis and in experimental Th1/Th2 driven enteritis. Proc. Natl. Acad. Sci. USA 2010, 107, 8017–8022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Humphreys, N.E.; Xu, D.; Hepworth, M.R.; Liew, F.Y.; Grencis, R.K. IL-33, a potent inducer of adaptive immunity to intestinal nematodes. J. Immunol. 2008, 180, 2443–2449. [Google Scholar] [CrossRef]
- Neill, D.R.; Wong, S.H.; Bellosi, A.; Flynn, R.J.; Daly, M.; Langford, T.K.; Bucks, C.; Kane, C.M.; Fallon, P.G.; Pannell, R.; et al. Nuocytes represent a new innate effector leukocyte that mediates type-2 immunity. Nature 2010, 464, 1367–1370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Salvo, C.; Pastorelli, L.; Petersen, C.P.; Buttò, L.F.; Buela, K.A.; Omenetti, S.; Locovei, S.A.; Ray, S.; Friedman, H.R.; Duijser, J.; et al. IL-33 triggers early eosinophil-dependent events leading to metaplasia in a chronic model of gastritis-prone mice. Gastroenterology 2020, 160, 302–316. [Google Scholar] [CrossRef]
- Lu, B.; Yang, M.; Wang, Q. Interleukin-33 in tumorigenesis, tumor immune evasion, and cancer immunotherapy. J. Mol. Med. 2016, 94, 535–543. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.X.; Hu, Z.; Shen, X.; Dong, L.Y.; Zhou, W.Z.; Hu, W.H. IL-33 Promotes Gastric Cancer Cell Invasion and Migration Via ST2-ERK1/2 Pathway. Dig. Dis. Sci. 2015, 60, 1265–1272. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.L.; Zhao, Y.R.; Weng, G.B.; Chen, Y.C.; Wei, X.N.; Shao, J.P.; Ji, H. IL-33-induced JNK pathway activation confers gastric cancer chemotherapy resistance. Oncol. Rep. 2015, 33, 2746–2752. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Li, X.; Li, Q.; Tan, Y.; Xu, B.; Xie, Q.; Deng, X.; Lu, B.; Jiang, J.; Wu, C. Interleukin-33 Expression does not Correlate with Survival of Gastric Cancer Patients. Pathol. Oncol. Res. 2017, 23, 615–619. [Google Scholar] [CrossRef] [PubMed]
- Eissmann, M.F.; Dijkstra, C.; Wouters, M.A.; Baloyan, D.; Mouradov, D.; Nguyen, P.M.; Davalos-Salas, M.; Putoczki, T.L.; Sieber, O.M.; Mariadason, J.M.; et al. Interleukin 33 Signaling Restrains Sporadic Colon Cancer in an Interferon-γ-Dependent Manner. Cancer Immunol. Res. 2018, 6, 409–421. [Google Scholar] [CrossRef] [Green Version]
- Beltrán, C.J.; Núñez, L.E.; Díaz-Jiménez, D.; Farfan, N.; Candia, E.; Heine, C.; López, F.; González, M.J.; Quera, R.; Hermoso, M.A. Characterization of the novel ST2/IL-33 system in patients with inflammatory bowel disease. Inflamm. Bowel Dis. 2010, 16, 1097–1107. [Google Scholar] [CrossRef]
- Buzzelli, J.N.; Chalinor, H.V.; Pavlic, D.I.; Sutton, P.; Menheniott, T.R.; Giraud, A.S.; Judd, L.M. IL33 Is a Stomach Alarmin That Initiates a Skewed Th2 Response to Injury and Infection. Cell Mol. Gastroenterol. Hepatol. 2015, 1, 203–221.e203. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Wang, X.; Yang, Q.; Zhao, X.; Wen, W.; Li, G.; Lu, J.; Qin, W.; Qi, Y.; Xie, F.; et al. Tumoral expression of IL-33 inhibits tumor growth and modifies the tumor microenvironment through CD8+ T and NK cells. J. Immunol. 2015, 194, 438–445. [Google Scholar] [CrossRef] [Green Version]
- Tong, X.; Barbour, M.; Hou, K.; Gao, C.; Cao, S.; Zheng, J.; Zhao, Y.; Mu, R.; Jiang, H.R. Interleukin-33 predicts poor prognosis and promotes ovarian cancer cell growth and metastasis through regulating ERK and JNK signaling pathways. Mol. Oncol. 2016, 10, 113–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bianchetti, L.; Marini, M.A.; Isgrò, M.; Bellini, A.; Schmidt, M.; Mattoli, S. IL-33 promotes the migration and proliferation of circulating fibrocytes from patients with allergen-exacerbated asthma. Biochem. Biophys. Res. Commun. 2012, 426, 116–121. [Google Scholar] [CrossRef]
- Saluja, R.; Hawro, T.; Eberle, J.; Church, M.K.; Maurer, M. Interleukin-33 promotes the proliferation of mouse mast cells through ST2/MyD88 and p38 MAPK-dependent and Kit-independent pathways. J. Biol. Regul. Homeost. Agents 2014, 28, 575–585. [Google Scholar]
- Tominaga, S.; Tago, K.; Tsuda, H.; Komine, M. Dual function of IL-33 on proliferation of NIH-3T3 cells. Cytokine 2015, 72, 105–108. [Google Scholar] [CrossRef]
- Fang, Y.; Zhao, L.; Xiao, H.; Cook, K.M.; Bai, Q.; Herrick, E.J.; Chen, X.; Qin, C.; Zhu, Z.; Wakefield, M.R.; et al. IL-33 acts as a foe to MIA PaCa-2 pancreatic cancer. Med. Oncol. 2017, 34, 23. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.G.; Walker, C.L. Cyclins and cell cycle checkpoints. Annu. Rev. Pharmacol. Toxicol. 1999, 39, 295–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sage, J. Cyclin C makes an entry into the cell cycle. Dev. Cell 2004, 6, 607–608. [Google Scholar] [CrossRef] [Green Version]
- Sage, J.; Miller, A.L.; Pérez-Mancera, P.A.; Wysocki, J.M.; Jacks, T. Acute mutation of retinoblastoma gene function is sufficient for cell cycle re-entry. Nature 2003, 424, 223–228. [Google Scholar] [CrossRef]
- Yang, Z.P.; Ling, D.Y.; Xie, Y.H.; Wu, W.X.; Li, J.R.; Jiang, J.; Zheng, J.L.; Fan, Y.H.; Zhang, Y. The Association of Serum IL-33 and sST2 with Breast Cancer. Dis. Markers 2015, 2015, 516895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kania, J.; Konturek, S.J.; Marlicz, K.; Hahn, E.G.; Konturek, P.C. Expression of survivin and caspase-3 in gastric cancer. Dig. Dis. Sci. 2003, 48, 266–271. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Chen, X.Y.; Liu, J.; Cheng, X.X.; Wang, X.W.; Kong, Q.Y.; Li, H. Differential caspase-3 expression in noncancerous, premalignant and cancer tissues of stomach and its clinical implication. Cancer Detect. Prev. 2006, 30, 168–173. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Ni, F.; Yu, F.; Cui, Z.; Zhu, X.; Chen, J. Prognostic significance of mRNA expression of CASPs in gastric cancer. Oncol. Lett. 2019, 18, 4535–4554. [Google Scholar] [CrossRef]
- Huang, K.H.; Fang, W.L.; Li, A.F.; Liang, P.H.; Wu, C.W.; Shyr, Y.M.; Yang, M.H. Caspase-3, a key apoptotic protein, as a prognostic marker in gastric cancer after curative surgery. Int. J. Surg. 2018, 52, 258–263. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Jeung, H.C.; Jeong, H.J.; Choi, Y.H.; Kim, J.E.; Jung, J.J.; Rha, S.Y.; Yang, W.I.; Chung, H.C. Identification of genes with correlated patterns of variations in DNA copy number and gene expression level in gastric cancer. Genomics 2007, 89, 451–459. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Lahti, J.M.; Valentine, M.; Saito, M.; Reed, S.I.; Look, A.T.; Kidd, V.J. Molecular cloning and chromosomal localization of the human cyclin C (CCNC) and cyclin E (CCNE) genes: Deletion of the CCNC gene in human tumors. Genomics 1996, 32, 253–259. [Google Scholar] [CrossRef]
- O’Donnell, C.; Mahmoud, A.; Keane, J.; Murphy, C.; White, D.; Carey, S.; O’Riordain, M.; Bennett, M.W.; Brint, E.; Houston, A. An antitumorigenic role for the IL-33 receptor, ST2L, in colon cancer. Br. J. Cancer 2016, 114, 37–43. [Google Scholar] [CrossRef] [Green Version]
- Maywald, R.L.; Doerner, S.K.; Pastorelli, L.; De Salvo, C.; Benton, S.M.; Dawson, E.P.; Lanza, D.G.; Berger, N.A.; Markowitz, S.D.; Lenz, H.J.; et al. IL-33 activates tumor stroma to promote intestinal polyposis. Proc. Natl Acad Sci USA 2015, 112, E2487–E2496. [Google Scholar] [CrossRef] [Green Version]
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Cancer incidence and mortality worldwide: Sources, methods and major patterns in GLOBOCAN 2012. Int. J. Cancer 2015, 136, E359–E386. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | N | Gender | n (%) | Age (Mean ± SD) |
|---|---|---|---|---|
| Healthy Controls (HC) | 48 | F M | 33 (68.8) 15 (31.3) | 60.7 ± 15.1 |
| Gastritis H.pylori-negative (G Hp−) | 21 | F M | 10 (47.6) 11 (52.4) | 71.5 ± 14.1 |
| Gastritis H.pylori-positive (G Hp+) | 34 | F M | 21 (61.8) 13 (38.2) | 66.2 ± 18.5 |
| GeneSymbol | GES-1 (Fold Change) | GES-1 + rhIL-33 (Fold Change) | p-Value | AGS (Fold Change) | p-Value | AGS + rhIL-33 (Fold Change) | p-Value |
|---|---|---|---|---|---|---|---|
| BCCIP | 1.000 | 1.453 | 0.281 | 2.395 | 0.002 | 1.649 | 0.019 |
| CASP3 | 1.000 | 1.818 | 0.141 | 24.084 | 0.00003 | 10.566 | 0.010 |
| CCNB1 | 1.000 | 1.446 | 0.284 | 2.389 | 0.002 | 1.653 | 0.019 |
| CCNC | 1.000 | 1.473 | 0.265 | 6.105 | 0.053 | 3.313 | 0.0004 |
| CCNE1 | 1.000 | 1.153 | 0.576 | 6.035 | 0.0001 | 4.145 | 0.0001 |
| KPNA2 | 1.000 | 1.456 | 0.006 | 3.046 | 0.0002 | 1.039 | 0.744 |
| Group | N | Gender | n (%) | Age (Mean ± SD) |
|---|---|---|---|---|
| Healthy Controls (HC) | 48 | F M | 33 (68.8) 15 (31.3) | 60.7 ± 15.1 |
| Gastric cancer (GC) | 16 | F M | 10 (62.5) 6 (37.5) | 75.6 ± 8.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pisani, L.F.; Tontini, G.E.; Gentile, C.; Marinoni, B.; Teani, I.; Nandi, N.; Creo, P.; Asti, E.; Bonavina, L.; Vecchi, M.; et al. Proinflammatory Interleukin-33 Induces Dichotomic Effects on Cell Proliferation in Normal Gastric Epithelium and Gastric Cancer. Int. J. Mol. Sci. 2021, 22, 5792. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115792
Pisani LF, Tontini GE, Gentile C, Marinoni B, Teani I, Nandi N, Creo P, Asti E, Bonavina L, Vecchi M, et al. Proinflammatory Interleukin-33 Induces Dichotomic Effects on Cell Proliferation in Normal Gastric Epithelium and Gastric Cancer. International Journal of Molecular Sciences. 2021; 22(11):5792. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115792
Chicago/Turabian StylePisani, Laura Francesca, Gian Eugenio Tontini, Carmine Gentile, Beatrice Marinoni, Isabella Teani, Nicoletta Nandi, Pasquale Creo, Emanuele Asti, Luigi Bonavina, Maurizio Vecchi, and et al. 2021. "Proinflammatory Interleukin-33 Induces Dichotomic Effects on Cell Proliferation in Normal Gastric Epithelium and Gastric Cancer" International Journal of Molecular Sciences 22, no. 11: 5792. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115792