
A Novel Hexose Transporter ChHxt6 Is Required for Hexose Uptake and Virulence in Colletotrichum higginsianum

 , , and
, , and 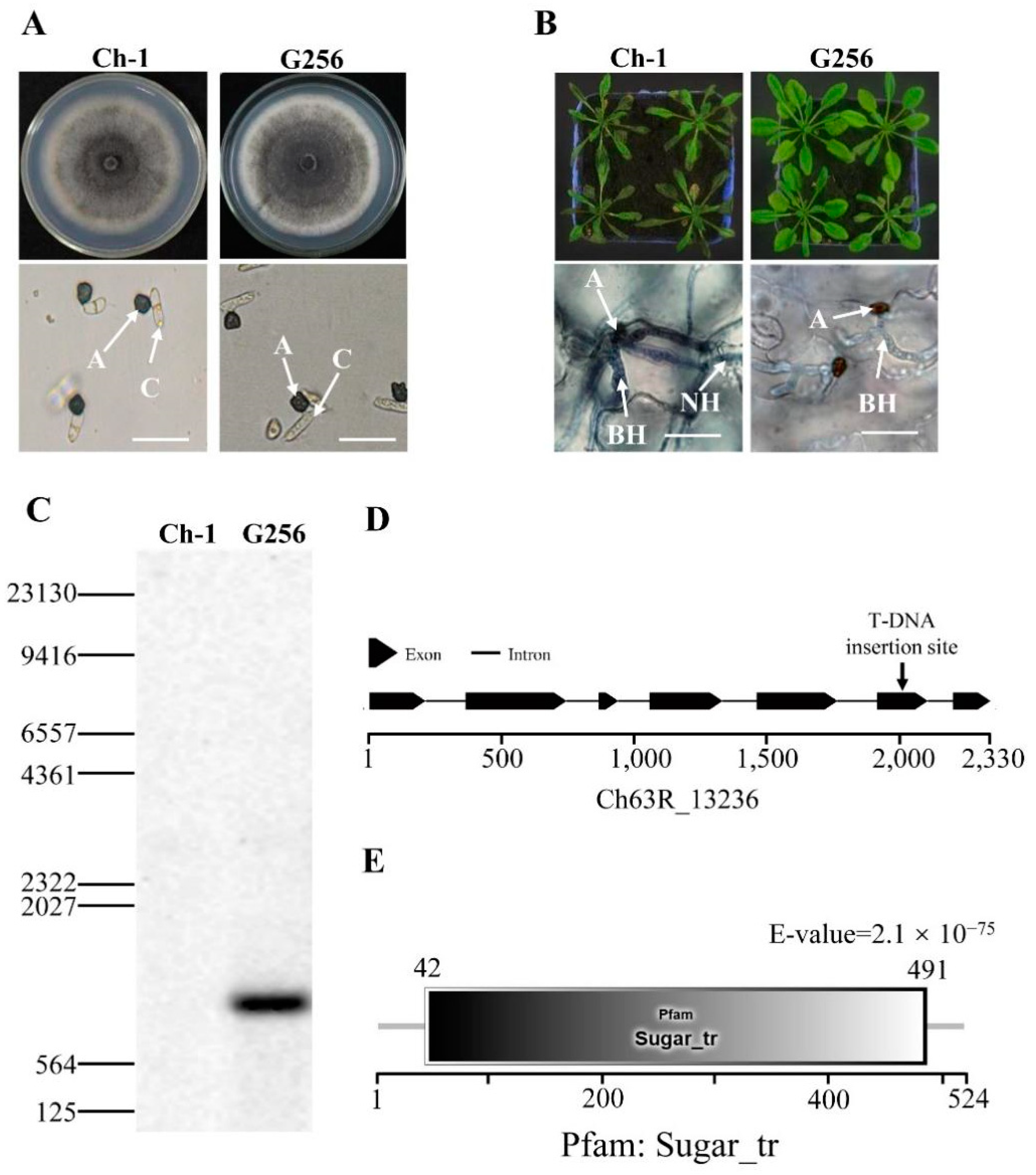{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
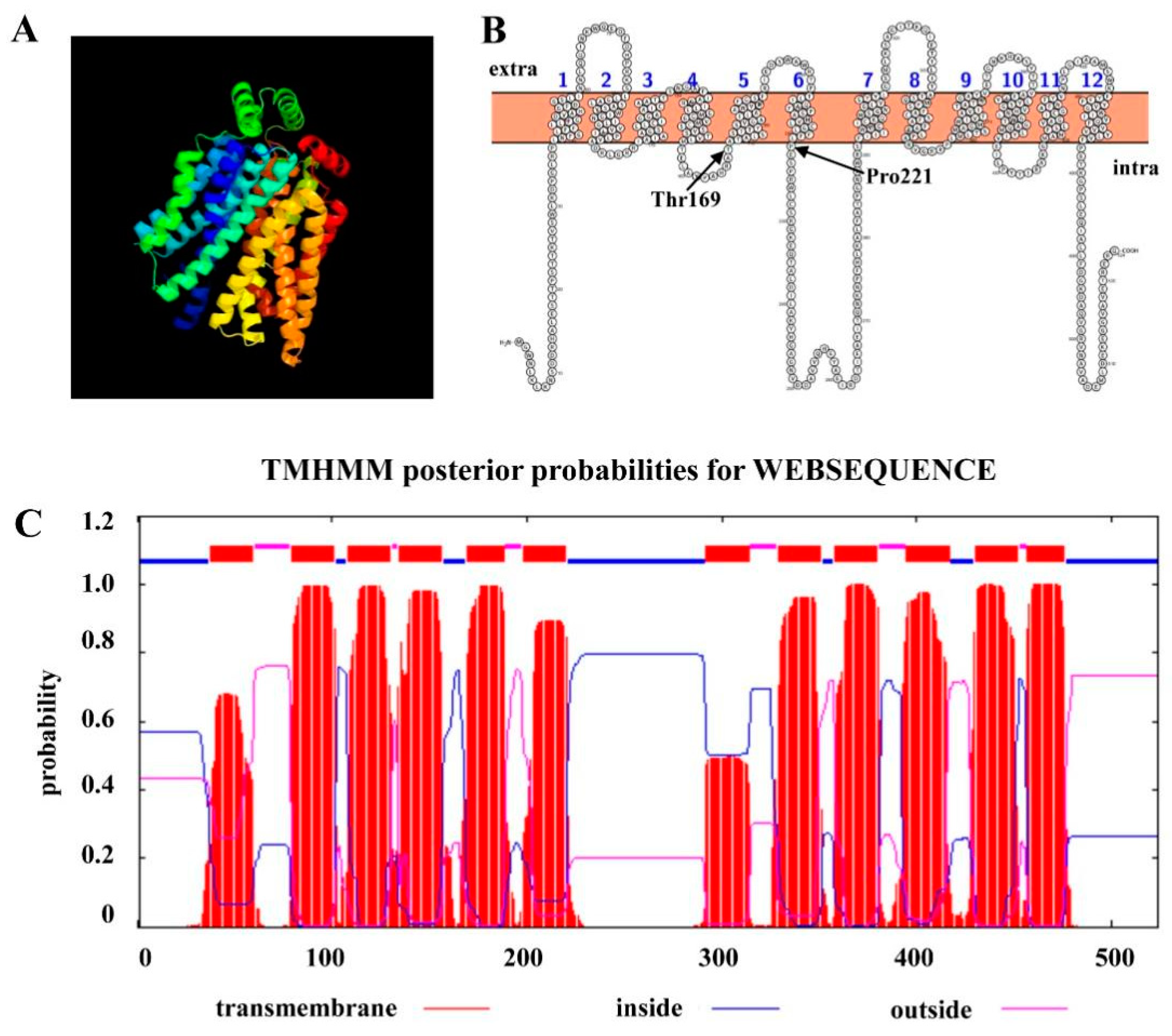
2.1. Identification of a New Hexose Transporter Gene in C. higginsianum
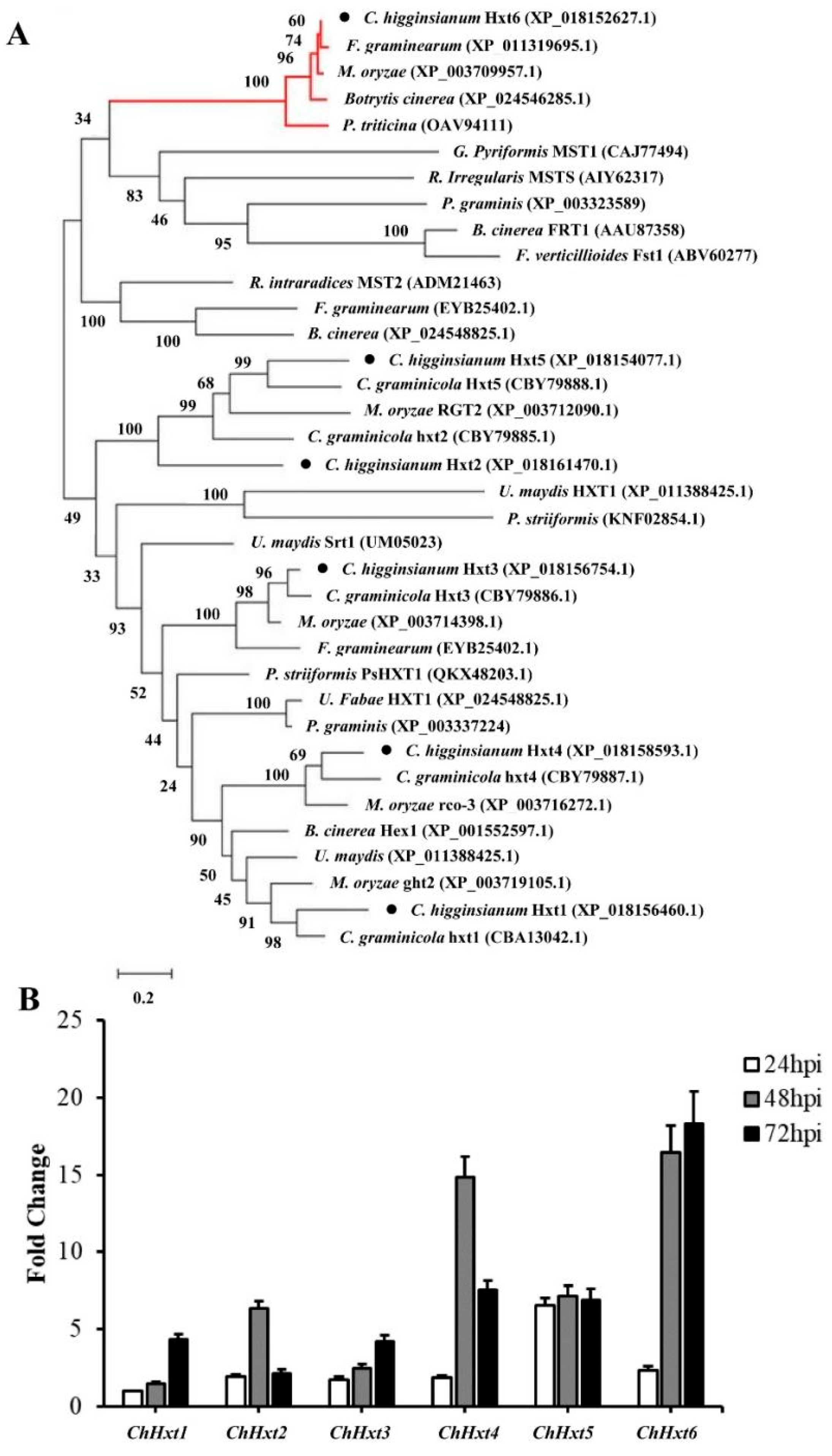
2.2. Characterizations of Six Hexose Transporter Genes in C. higginsianum
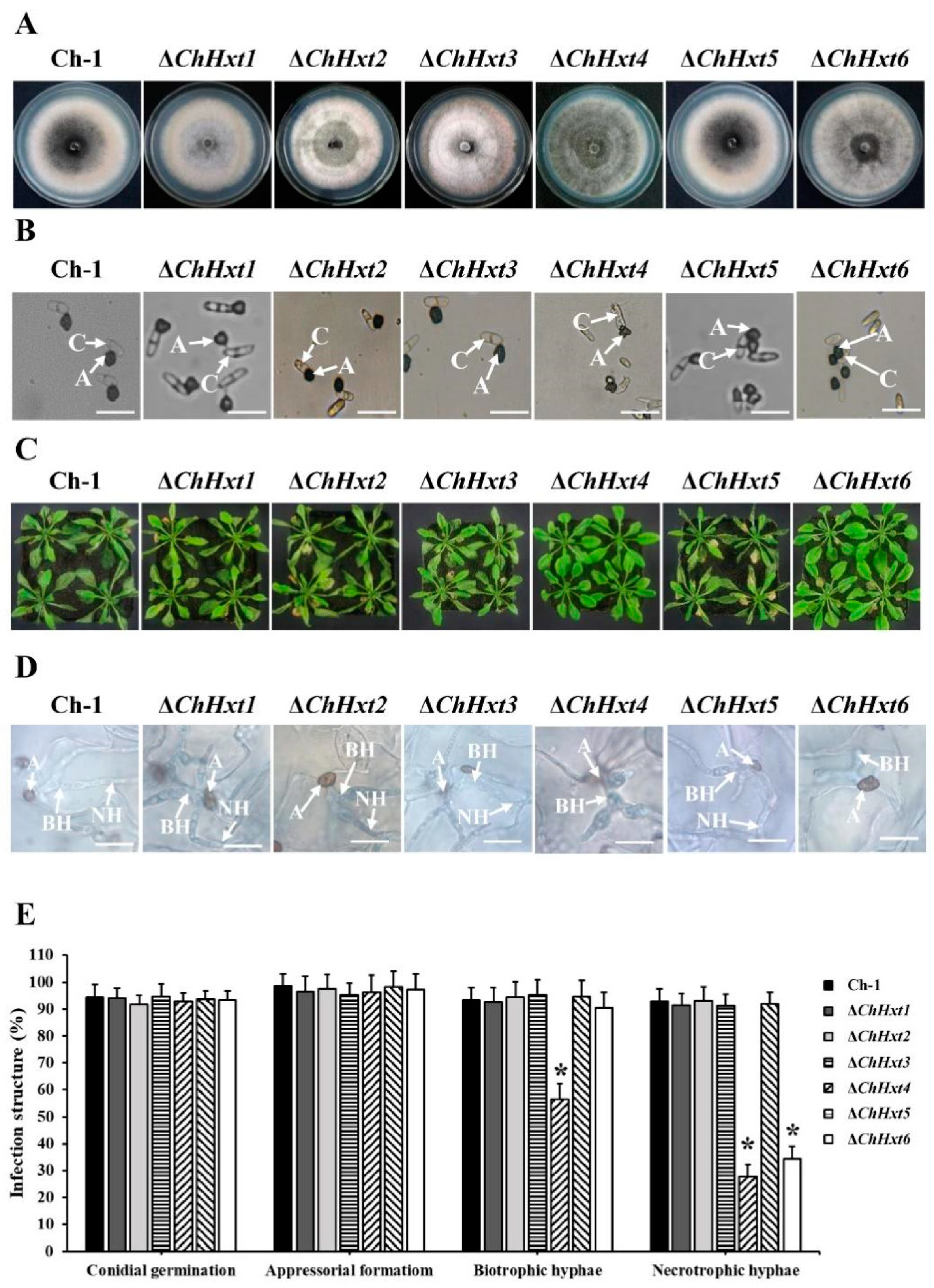
2.3. Deletion of the Six Hexose Transporter Genes in C. higginsianum
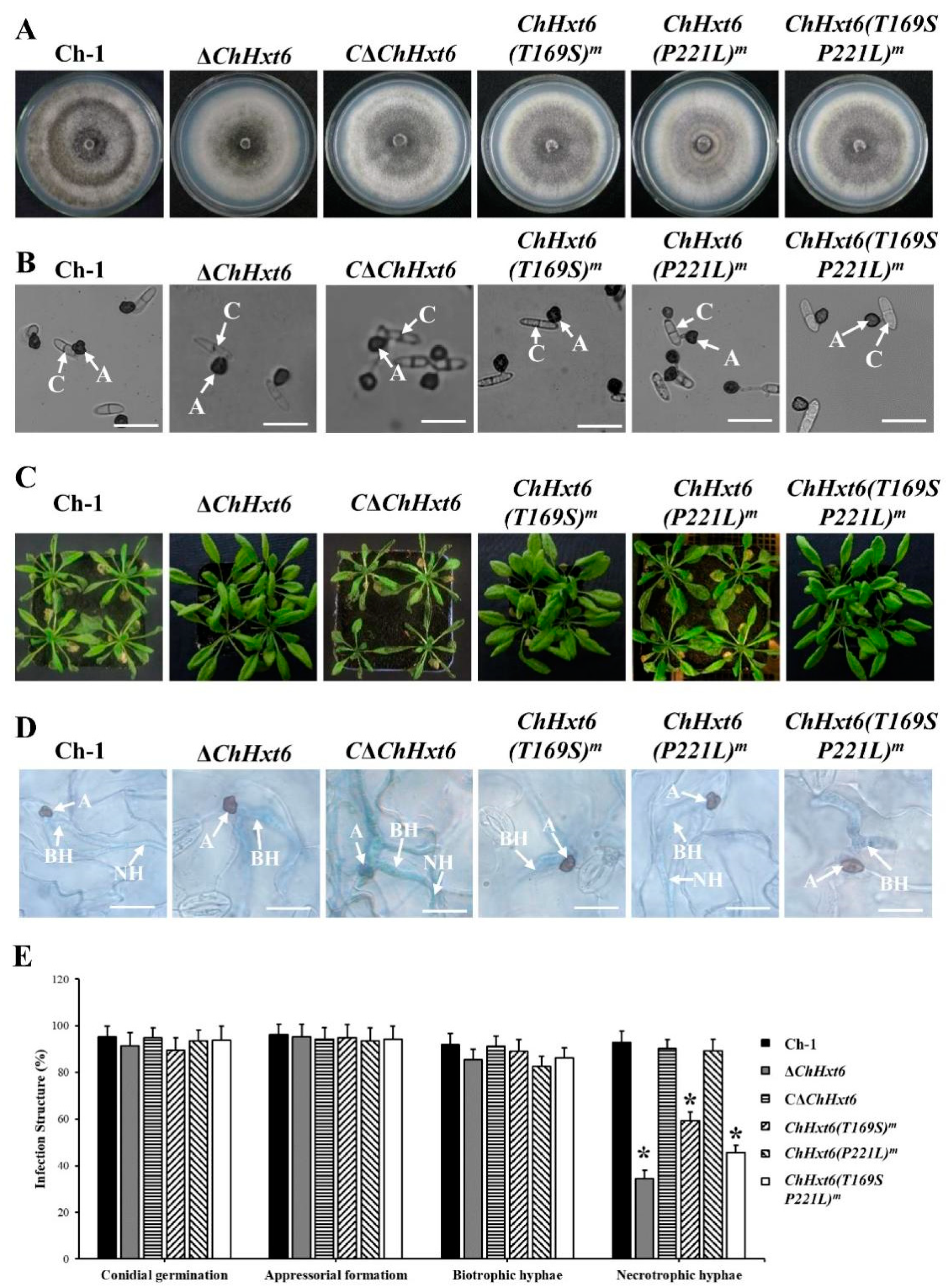
2.4. ChHxt4 and ChHxt6 Are Important for Virulence in C. higginsianum
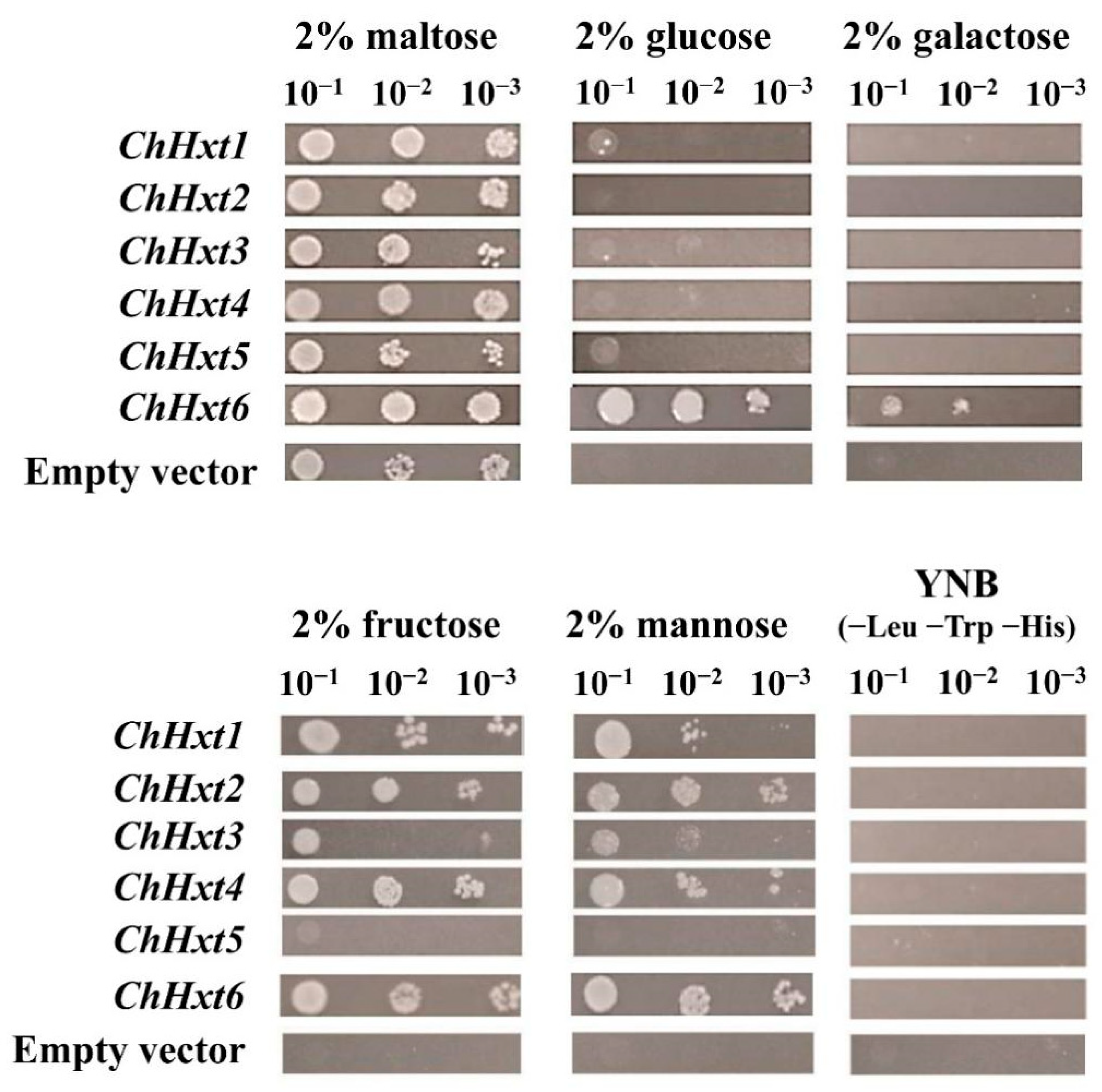
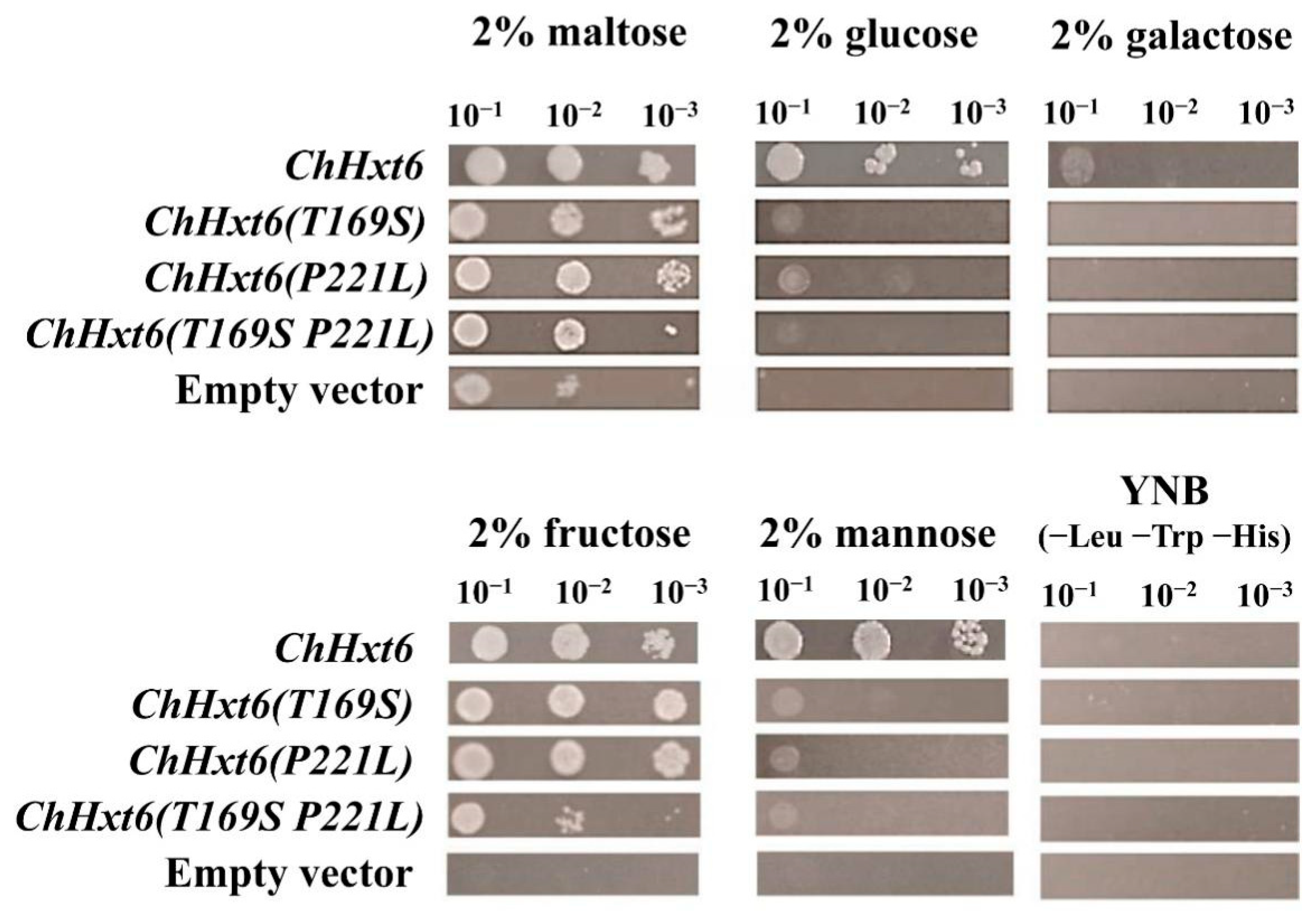
2.5. ChHxt6 Is a Central Monosaccharide Transporter for Hexose Uptake
2.6. Thr169 Position in ChHxt6 Is Vital for Hexose Transport and Virulence
3. Discussion
4. Materials and Methods
4.1. Strains, Vectors and Plants
4.2. Molecular Identification of T-DNA Insertion Site from a Virulence-Deficient Mutant
4.3. Deletion and Complementation of ChHxt Genes
4.4. Phenotype Analysis
4.5. Virulence Assay and Plant Infection Observation
4.6. Site-Directed Mutagenesis
4.7. Expression of ChHxts in Yeast
4.8. DNA/RNA Manipulation, Southern Blotting, RT-PCR and RT-qPCR
4.9. Bioinformatics
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hyde, K.D.; Cai, L.; Cannon, P.F.; Crouch, J.A.; Crous, P.W.; Damm, U.; Goodwin, P.H.; Chen, H.; Johnston, P.R.; Jones, E.B.G.; et al. Colletotrichum-names in current use. Fungal Divers. 2009, 39, 147–182. [Google Scholar] [CrossRef]
- O’Connell, R.; Herbert, C.; Sreenivasaprasad, S.; Khatib, M.; Esquerré-Tugayé, M.T.; Dumas, B. A novel Arabidopsis-Colletotrichum pathosystem for the molecular dissection of plant-fungal interactions. Mol. Plant Microbe Interact. 2004, 17, 272–282. [Google Scholar] [CrossRef] [Green Version]
- Bisson, L.F.; Neigeborn, L.; Carlson, M.; Fraenkel, D.G. The SNF3 gene is required for high-affinity glucose transport in Saccharomyces cerevisiae. J. Bacteriol. 1987, 169, 1656–1662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.Q.; Cheung, L.S.; Feng, L.; Tanner, W.; Formmer, W.B. Transport of sugars. Annu. Rev. Biochem. 2015, 84, 865–894. [Google Scholar] [CrossRef] [PubMed]
- Lewis, D.A.; Bisson, L.F. The HXT1 gene product of Saccharomyces cerevisiae is a new member of the family of hexose transporters. Mol. Cell Biol. 1991, 11, 3804–3813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lagunas, R. Sugar transport in Saccharomyces cerevisiae. FEMS Microbiol. Rev. 1993, 10, 229–242. [Google Scholar] [CrossRef] [PubMed]
- Voegele, R.T.; Struck, C.; Hahn, M.; Mendgen, K. The role of haustoria in sugar supply during infection of broad bean by the rust fungus Uromyces fabae. Proc. Natl. Acad. Sci. USA 2001, 98, 8133–8138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voegele, R.T.; Wirsel, S.; Möll, U.; Lechner, M.; Mendgen, K. Cloning and characterization of a novel invertase from the obligate biotroph Uromyces fabae and analysis of expression patterns of host and pathogen invertases in the course of infection. Mol. Plant Microbe Interact. 2006, 19, 625–634. [Google Scholar] [CrossRef] [Green Version]
- Wahl, R.; Wippel, K.; Goos, S.; Kämper, J.; Sauer, N. A novel high-affinity sucrose transporter is required for virulence of the plant pathogen Ustilago maydis. PLoS Biol. 2010, 8, e1000303. [Google Scholar] [CrossRef] [Green Version]
- Schuler, D.; Wahl, R.; Wippel, K.; Vranes, M.; Munsterkotter, M.; Sauer, N.; Kämper, J. Hxt1, a monosaccharide transporter and sensor required for virulence of the maize pathogen Ustilago maydis. New Phytol. 2015, 206, 1086–1100. [Google Scholar] [CrossRef]
- Chang, Q.; Liu, J.; Lin, X.H.; Hu, S.J.; Yang, Y.; Li, D.; Chen, L.Y.; Huai, B.Y.; Huang, L.L.; Voegele, R.T.; et al. A unique invertase is important for sugar absorption of an obligate biotrophic pathogen during infection. New Phytol. 2017, 215, 1548–1561. [Google Scholar] [CrossRef] [Green Version]
- Chang, Q.; Lin, X.H.; Yao, M.; Liu, P.; Guo, J.; Huang, L.L.; Voegele, R.T.; Kang, Z.S.; Liu, J. Hexose transporter PsHXT1-mediated sugar uptake is required for pathogenicity of wheat stripe rust. Plant Biotechnol. J. 2020, 18, 2367–2369. [Google Scholar] [CrossRef] [PubMed]
- Schüssler, A.; Martin, H.; Cohen, D.; Fitz, M.; Wipf, D. Characterization of a carbohydrate transporter from symbiotic glomeromycotan fungi. Nature 2006, 444, 933–936. [Google Scholar] [CrossRef] [PubMed]
- Lingner, U.; Münch, S.; Deising, H.B.; Sauer, N. Hexose transporters of a hemibiotrophic plant pathogen: Functional variations and regulatory differences at different stages of infection. J. Biol. Chem. 2011, 286, 20913–20922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saitoh, H.; Hirabuchi, A.; Fujisawa, S.; Mitsuoka, C.; Terauchi, R.; Takano, Y. MoST1 encoding a hexose transporter-like protein is involved in both conidiation and mycelial melanization of Magnaporthe oryzae. FEMS Microbiol. Lett. 2014, 352, 104–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Connell, R.J.; Thon, M.R.; Hacquard, S.; Amyotte, S.G.; Kleemann, J.; Torres, M.F.; Damm, U.; Buiate, E.A.; Epstein, L.; Alkan, N.; et al. Lifestyle transitions in plant pathogenic Colletotrichum fungi deciphered by genome and transcriptome analyses. Nat. Genet. 2012, 44, 1060–1065. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.P.; Zhao, D.; Zheng, L.; Hsiang, T.; Wei, Y.D.; Fu, Y.P.; Huang, J.B. Identification of virulence genes in the crucifer anthracnose fungus Colletotrichum higginsianum by insertional mutagenesis. Microb. Pathog. 2013, 64, 6–17. [Google Scholar] [CrossRef]
- Batista, A.S.; Miletti, L.C.; Stambuk, B.U. Sucrose fermentation by Saccharomyces cerevisiae lacking hexose transport. J. Mol. Microbiol. Biotechnol. 2004, 8, 26–33. [Google Scholar] [CrossRef]
- Sambrook, J.; Russell, D.W. Molecular cloning: A Laboratory Manual. In Cold Spring Harbor, 3rd ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001. [Google Scholar]
- Yuan, Q.F.; Chen, M.J.; Yan, Y.Q.; Gu, Q.N.; Huang, J.B.; Zheng, L. ChSte7 is required for vegetative growth and various plant infection processes in Colletotrichum higginsianum. Biomed Res. Int. 2016, 2016, e7496569. [Google Scholar] [CrossRef] [Green Version]
- Carrigan, P.E.; Ballar, P.; Tuzmen, S. Site-directed mutagenesis. Methods Mol. Biol. 2011, 700, 107–124. [Google Scholar] [CrossRef]
- Gietz, R.D.; Schiestl, R.H. High-efficiency yeast transformation using the LiAc/SS carrier DNA/PEG method. Nat. Protoc. 2007, 2, 31–34. [Google Scholar] [CrossRef]
- Gu, Q.N.; Yuan, Q.F.; Zhao, D.; Huang, J.B.; Hsiang, T.; Wei, Y.D.; Zheng, L. Acetyl-coenzyme A synthetase gene ChAcs1 is essential for lipid metabolism, carbon utilization and virulence of the hemibiotrophic fungus Colletotrichum higginsianum. Mol. Plant Pathol. 2019, 20, 107–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [Green Version]
- Solovyev, V.; Kosarev, P.; Seledsov, I.; Vorobyev, D. Automatic annotation of eukaryotic genes, pseudogenes and promoters. Genome Biol. 2006, 7, 10.1–10.12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Letunic, I.; Khedkar, S.; Bork, P. SMART: Recent updates, new developments and status in 2020. Nucleic Acids Res. 2021, 49, 458–460. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Dobson, L.; Reményi, I.; Tusnády, G.E. CCTOP: A Consensus Constrained TOPology prediction web server. Nucleic Acids Res. 2015, 43, 408–412. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, Q.; Yan, Y.; Sohail, M.A.; Liu, H.; Huang, J.; Hsiang, T.; Zheng, L. A Novel Hexose Transporter ChHxt6 Is Required for Hexose Uptake and Virulence in Colletotrichum higginsianum. Int. J. Mol. Sci. 2021, 22, 5963. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115963
Yuan Q, Yan Y, Sohail MA, Liu H, Huang J, Hsiang T, Zheng L. A Novel Hexose Transporter ChHxt6 Is Required for Hexose Uptake and Virulence in Colletotrichum higginsianum. International Journal of Molecular Sciences. 2021; 22(11):5963. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115963
Chicago/Turabian StyleYuan, Qinfeng, Yaqin Yan, Muhammad Aamir Sohail, Hao Liu, Junbin Huang, Tom Hsiang, and Lu Zheng. 2021. "A Novel Hexose Transporter ChHxt6 Is Required for Hexose Uptake and Virulence in Colletotrichum higginsianum" International Journal of Molecular Sciences 22, no. 11: 5963. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115963