
Rhizobial Exopolysaccharides: Genetic Regulation of Their Synthesis and Relevance in Symbiosis with Legumes

 , and
, and 
Abstract
:1. Introduction
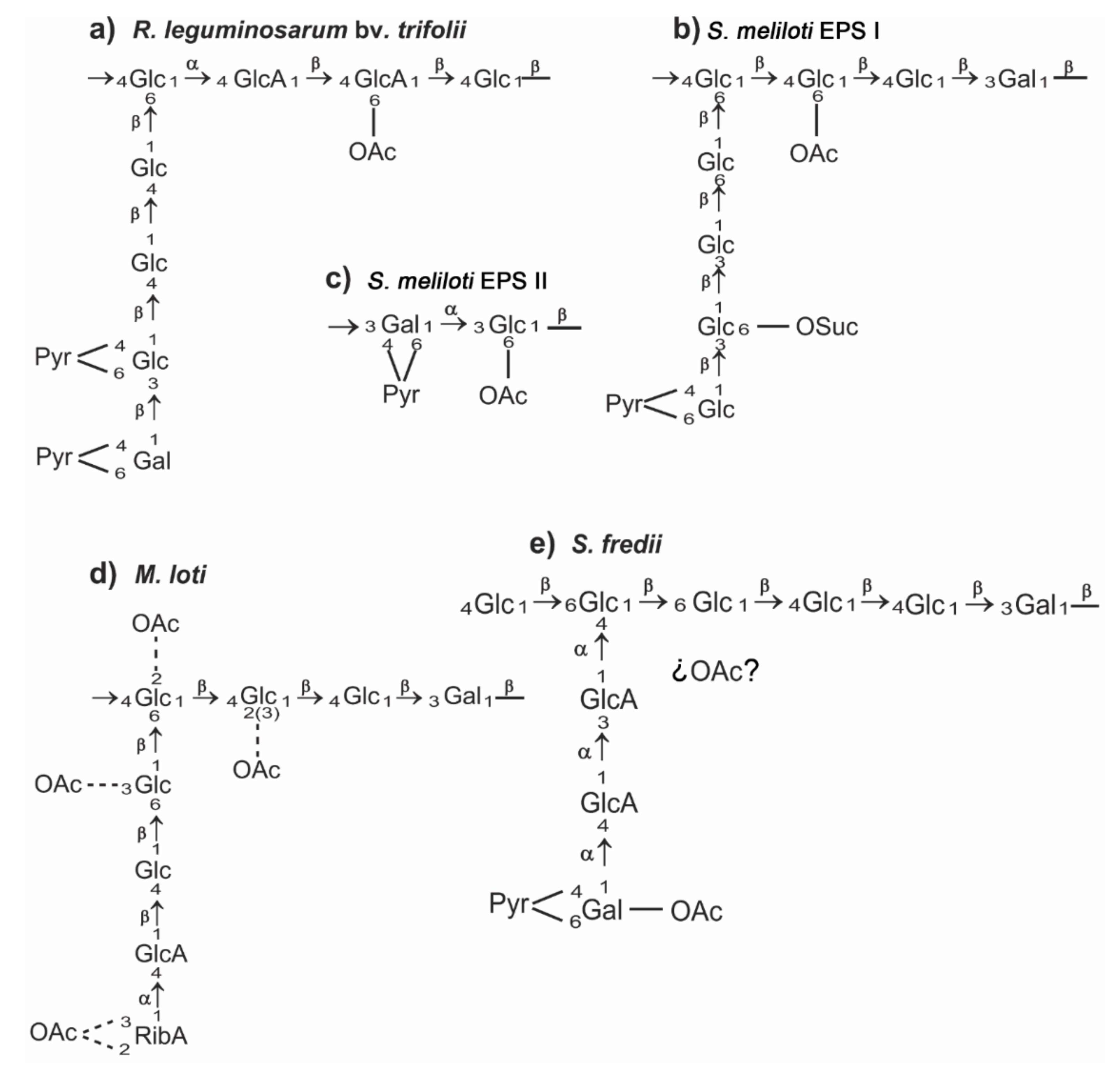
2. A General View of Rhizobial Exopolysaccharides
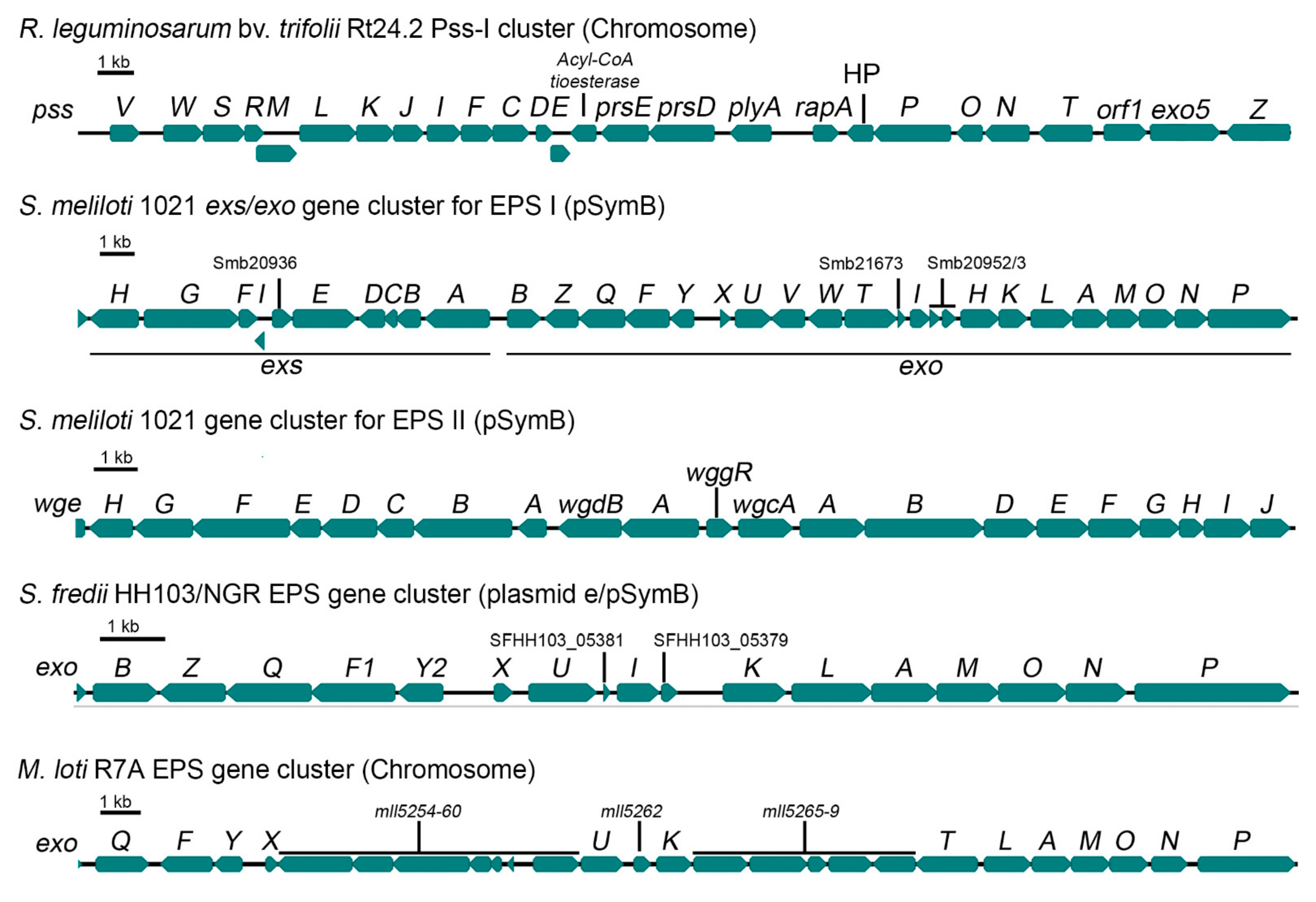
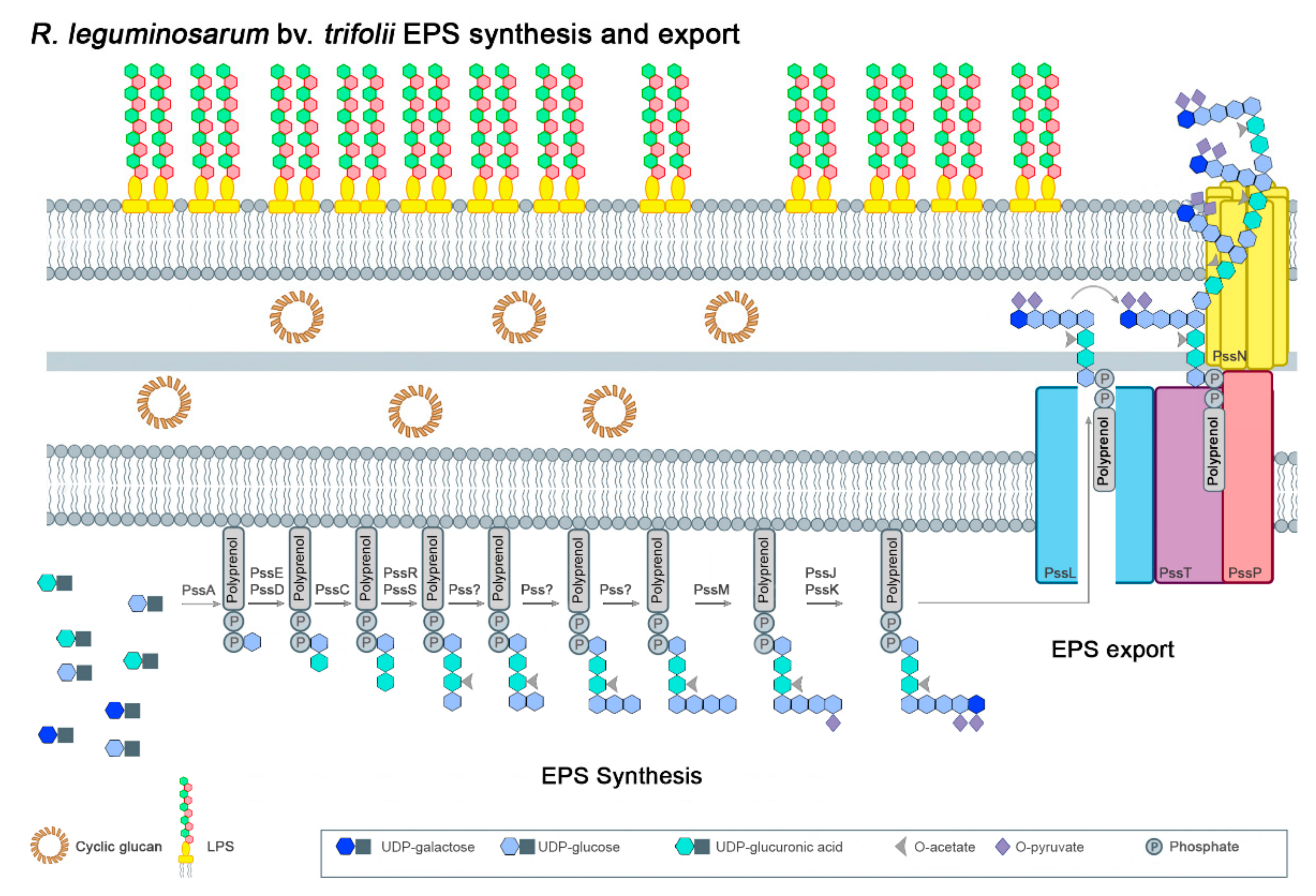
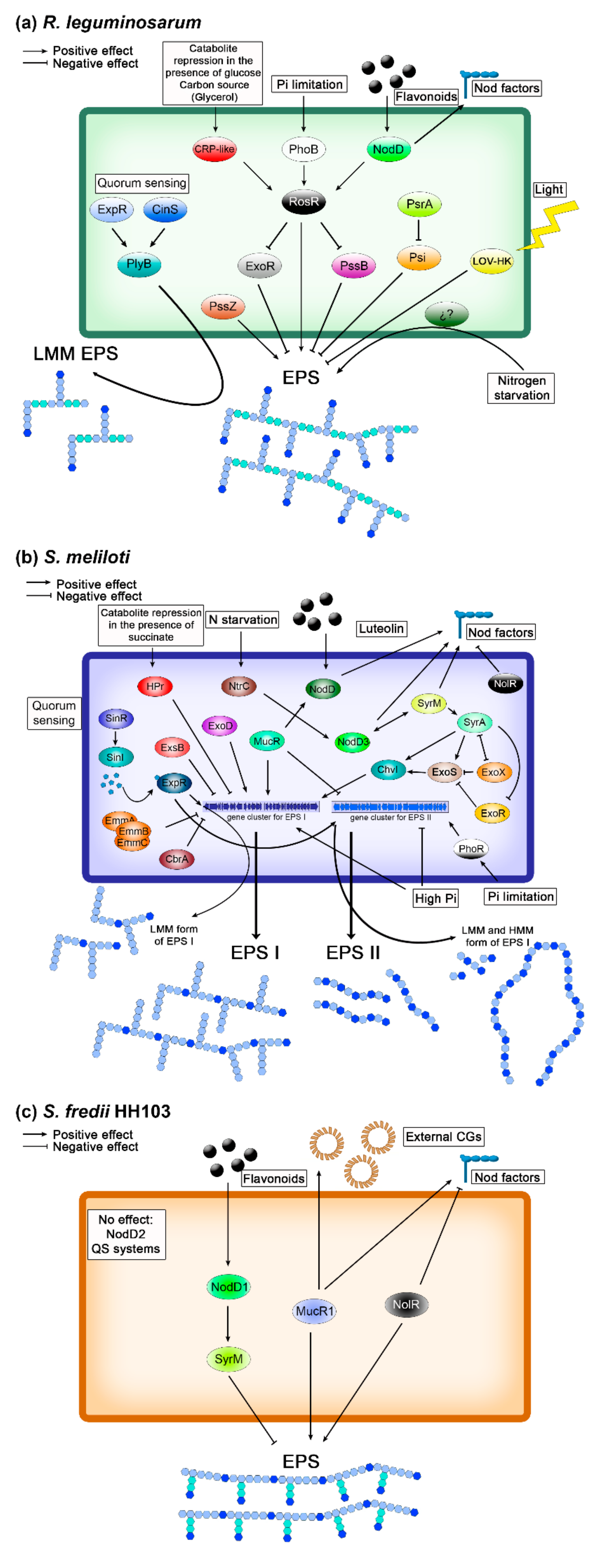
3. Rhizobium leguminosarum
4. Sinorhizobium meliloti
5. Sinorhizobium fredii
6. Mesorhizobium loti
7. Conclusions and Perspectives
- (1)
- Rhizobial EPS production has been found to be extremely complex in all the rhizobial strains in which this issue has been addressed. Usually, it involves the participation of different regulatory systems, either positive or negative ones, including the participation of global regulatory proteins and, in some cases, that of QS systems.
- (2)
- In many rhizobia-legume symbioses, EPS is absolutely essential or very important for a successful interaction. This is especially true in symbioses forming indeterminate nodules, although the interactions between S. fredii HH103 and G. uralensis and C. cajan are a clear exception. Also, the fact that EPS plays a very important role in the success of rhizobial infection in the M. loti/L. japonicus couple, which forms determinate nodules, makes it difficult to formulate a general rule. Interestingly, EPS appears to be crucial for the symbiotic performance of several rhizobia exhibiting a narrow host range (such as S. meliloti, R. leguminosarum, and M. loti).
- (3)
- In some rhizobia, EPS production is connected with the nod regulon, but the type of connection (positive or negative) varies depending on the rhizobial strain. Thus, in S. meliloti and R. leguminosarum bv. trifolii, appropriate flavonoids induce Nod factor production and stimulates EPS biosynthesis, which is enhanced in parallel to that of Nod factors, whereas in S. fredii HH103, appropriate flavonoids induce Nod factor production but suppress EPS biosynthesis.
- (4)
- Rhizobial EPS, in addition to being crucial for biofilm formation and for passive protection against abiotic stresses, may play a crucial role as a molecular signal required for suppressing plant defence responses and, thus, allowing a successful infection process. There is clear evidence that, at least in S. meliloti, R. leguminosarum bv. trifolii and S. fredii NGR234, this signalling function is carried out by oligosaccharides derived from EPS. The recent discovery of the Lotus japonicus EPR3 receptor as a detector of the presence of the “correct” oligosaccharides for granting a successful infection process supports this idea. However, in this context, it is surprising that S. fredii HH103, a broad host-range rhizobial strain, represses the production of its EPS in the presence of any nod genes inducing flavonoids and, thus, in the early steps of its symbiotic interactions with any potential host legume. Might this repression be a strategy for nodulating potential host legumes that would stop infection when they perceive the HH103 EPS to inadequate? Clearly, more research is required to elucidate this issue.
- (5)
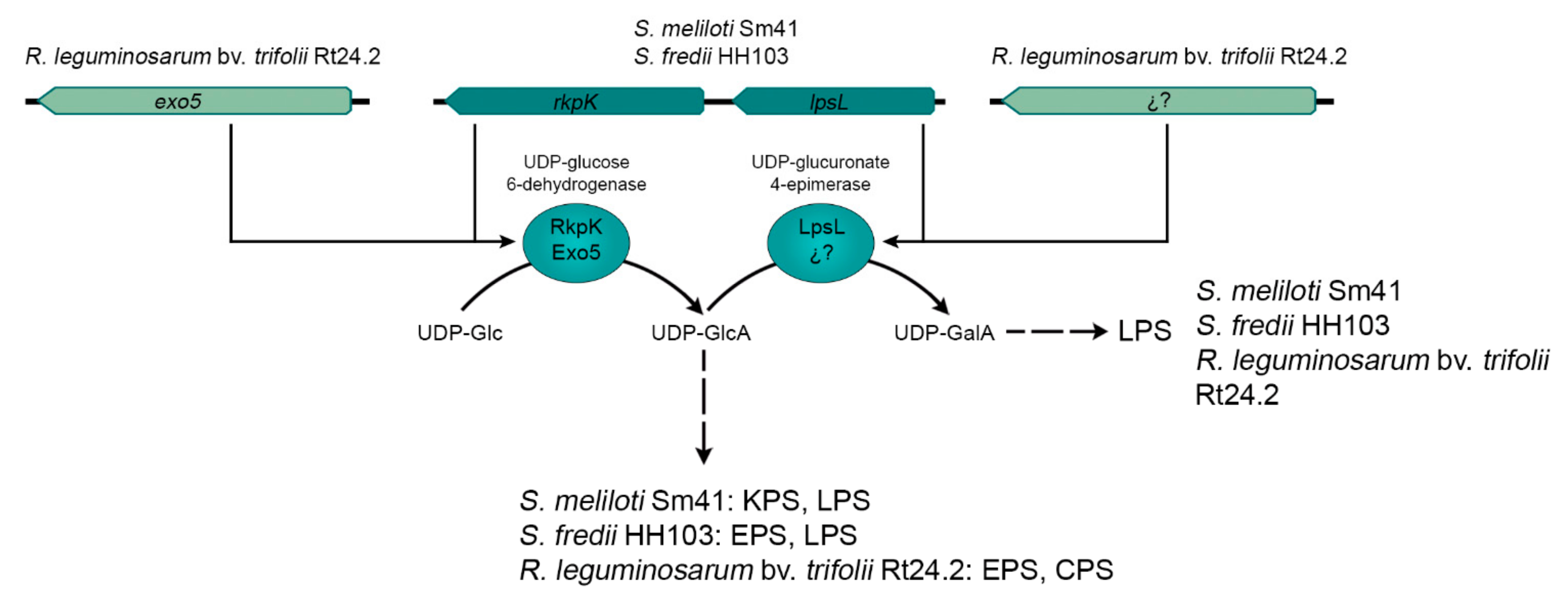
- Rhizobial EPS production may be interconnected with the biosynthesis of other symbiotic relevant surface polysaccharides. For example, in R. leguminosarum bv. trifolii and S. fredii, the synthesis of EPS and LPS are interconnected through the rkp-2 region, whereas in S. meliloti, this genetic region participates in the production of KPS and LPS. At least in S. fredii, the production of EPS and extracellular CG is simultaneously controlled by the MucR1 regulator.
- (6)
- Rhizobial EPS symbiotic function can overlap with that of other surface polysaccharides, such KPS in the case of the interaction between S. meliloti and Medicago. However, this is not a general rule, as S. fredii mutants lacking EPS and KPS can still induce the formation of nitrogen-fixing nodules on G. uralensis and C. cajan.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
| AHL | N-acyl-homoserine lactone(s) |
| CG | Cyclic glucans |
| CPS | Capsular polysaccharide |
| EPS | Exopolysaccharide |
| GlcA | Glucuronic acid |
| GalA | Galacturonic acid |
| HMM | High molecular mass |
| IRLC | Inverting repeat-lacking clade |
| IT | Infection thread(s) |
| KPS | K-antigen capsular polysaccharide |
| LMM | Low molecular mass |
| LOV-domain | Light, oxygen, and voltage domain |
| LPS | Lipopolysaccharide |
| QS | Quorum sensing |
| TM | Transmembrane |
References
- Poole, P.; Ramachandran, V.; Terpolilli, J. Rhizobia: From saprophytes to endosymbionts. Nat. Rev. Microbiol. 2018, 16, 291–303. [Google Scholar] [CrossRef] [PubMed]
- Tsyganova, A.V.; Kitaeva, A.B.; Tsyganov, V.E. Cell differentiation in nitrogen-fixing nodules hosting symbiosomes. Funct. Plant Biol. 2017, 45, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Coba de la Peña, T.; Fedorova, E.; Pueyo, J.J.; Lucas, M.M. The symbiosome: Legume and rhizobia co-evolution toward a nitrogen-fixing organelle? Front. Plant Sci. 2018, 8, 2229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oldroyd, G.E.D.; Dixon, R. Biotechnological solutions to the nitrogen problem. Curr. Opin. Biotechnol. 2014, 26, 19–24. [Google Scholar] [CrossRef] [Green Version]
- Santos, M.S.; Nogueira, M.A.; Hungria, M. Microbial inoculants: Reviewing the past, discussing the present and previewing an outstanding future for the use of beneficial bacteria in agriculture. AMB Express 2019, 9, 205. [Google Scholar] [CrossRef]
- Walker, L.; Lagunas, B.; Gifford, M.L. Determinants of host range specificity in legume-rhizobia symbiosis. Front. Microbiol. 2020, 11, 3028. [Google Scholar] [CrossRef]
- Janczarek, M.; Rachwał, K.; Marzec, A.; Grzadziel, J.; Palusińska-Szysz, M. Signal molecules and cell-surface components involved in early stages of the legume-rhizobium interactions. Appl. Soil Ecol. 2015, 85, 94–113. [Google Scholar] [CrossRef]
- López-Baena, F.J.; Ruiz-Sainz, J.E.; Rodríguez-Carvajal, M.A.; Vinardell, J.M. Bacterial molecular signals in the Sinorhizobium fredii-soybean symbiosis. Int. J. Mol. Sci. 2016, 17, 755. [Google Scholar] [CrossRef] [Green Version]
- Oldroyd, G.E. Speak, friend, and enter: Signalling systems that promote beneficial symbiotic associations in plants. Nat. Rev. Microbiol. 2013, 11, 252–263. [Google Scholar] [CrossRef]
- Roy, S.; Liu, W.; Nandety, R.S.; Crook, A.; Mysore, K.S.; Pislariu, C.I.; Frugoli, J.; Dickstein, R.; Udvardi, M.K. Celebrating 20 Years of Genetic Discoveries in Legume Nodulation and Symbiotic Nitrogen Fixation. Plant Cell 2020, 32, 15–41. [Google Scholar] [CrossRef] [Green Version]
- Staehelin, C.; Krishnan, H.B. Nodulation outer proteins: Double-edged swords of symbiotic rhizobia. Biochem. J. 2015, 470, 263–274. [Google Scholar] [CrossRef] [Green Version]
- Downie, J.A. The roles of extracellular proteins, polysaccharides and signals in the interactions of rhizobia with legume roots. FEMS Microbiol. Rev. 2010, 34, 150–170. [Google Scholar] [CrossRef]
- Wanke, A.; Malisic, M.; Wawra, S.; Zuccaro, A. Unraveling the sugar code: The role of microbial extracellular glycans in plant-microbe interactions. J. Exp. Bot. 2021, 72, 15–35. [Google Scholar] [CrossRef]
- Laus, M.C.; Logman, T.J.; van Brussel, A.A.N.; Carlson, R.W.; Azadi, P.; Gao, M.; Kijne, J.W. Involvement of exo5 in production of surface polysaccharides in Rhizobium leguminosarum and its role in nodulation of Vicia sativa subsp. nigra. J. Bacteriol. 2004, 186, 6617–6625. [Google Scholar] [CrossRef] [Green Version]
- Crespo-Rivas, J.C.; Margaret, I.; Hidalgo, A.; Buendía-Clavería, A.M.; Ollero, F.J.; López-Baena, F.J.; del Socorro Murdoch, P.; Rodríguez-Carvajal, M.A.; Soria-Díaz, M.E.; Reguera, M.; et al. Sinorhizobium fredii HH103 cgs mutants are unable to nodulate determinate- and indeterminate nodule-forming legumes and overproduce an altered EPS. Mol. Plant Microbe Interact. 2009, 22, 575–588. [Google Scholar] [CrossRef] [Green Version]
- Fraysse, N.; Couderc, F.; Poinsot, V. Surface polysaccharide involvement in establishing the rhizobium-legume symbiosis. Eur. J. Biochem. 2003, 270, 1365–1380. [Google Scholar] [CrossRef]
- Jones, K.M.; Kobayashi, H.; Davies, B.W.; Taga, M.E.; Walker, G.C. How rhizobial symbionts invade plants: The Sinorhizobium-Medicago model. Nat. Rev. Microbiol. 2007, 5, 619–633. [Google Scholar] [CrossRef] [Green Version]
- Kawaharada, Y.; Kelly, S.; Nielsen, M.W.; Hjuler, C.T.; Gysel, K.; Muszynski, A.; Carlson, R.W.; Thygesen, M.B.; Sandal, N.; Asmussen, M.H.; et al. Receptor-mediated exopolysaccharide perception controls bacterial infection. Nature 2015, 523, 308–312. [Google Scholar] [CrossRef] [Green Version]
- Kawaharada, Y.; Nielsen, M.; Kelly, S.; James, E.; Andersen, K.; Rasmussen, S.; Füchtbauer, W.; Madsen, L.; Heckmann, A.; Radutoiu, S.; et al. Differential regulation of the Epr3 receptor coordinates membrane-restricted rhizobial colonization of root nodule primordia. Nat. Commun. 2017, 8, 14534–14545. [Google Scholar] [CrossRef]
- Becker, A.; Pühler, A. Production of exopolysaccharides. In The Rhizobiaceae. Molecular Biology of Model Plant-Associated Bacteria; Spaink, H.P., Kondorosi, A., Hooykaas, J.J., Eds.; Springer: Dordrecht, The Netherlands, 1998; pp. 97–118. [Google Scholar] [CrossRef]
- Muszyński, A.; Heiss, C.; Hjuler, C.T.; Sullivan, J.T.; Kelly, S.J.; Thygesen, M.B.; Stougaard, J.; Azadi, P.; Carlson, R.W.; Ronson, C.W. Structures of exopolysaccharides involved in receptor-mediated perception of Mesorhizobium loti by Lotus japonicus. J. Biol. Chem. 2016, 291, 20946–20961. [Google Scholar] [CrossRef] [Green Version]
- Stacey, G.; So, J.S.; Roth, L.E.; Lakshmi, S.K.B.; Carlson, R.W. A lipopolysaccharide mutant of Bradyrhizobium japonicum that uncouples plant from bacterial differentiation. Mol. Plant Microbe Interact. 1991, 4, 332–340. [Google Scholar] [CrossRef] [PubMed]
- Maroti, G.; Kondorosi, E. Nitrogen-fixing Rhizobium-legume symbiosis: Are polyploidy and host peptide-governed symbiont differentiation general principles of endosymbiosis? Front. Microbiol. 2014, 5, 326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferguson, B.J.; Indrasumunar, A.; Hayashi, S.; Lin, M.H.; Lin, Y.H.; Reid, D.E.; Gresshoff, P.M. Molecular analysis of legume nodule development and autoregulation. J. Integr. Plant Biol. 2010, 52, 61–76. [Google Scholar] [CrossRef] [PubMed]
- Robertson, B.K.; Aman, P.; Darvill, A.G.; McNeil, M.; Albersheim, P. Host-symbiont interactions. The structure of acidic extracellular polysaccharides secreted by Rhizobium leguminosarum and Rhizobium trifolii. Plant Physiol. 1981, 67, 389–400. [Google Scholar] [CrossRef] [Green Version]
- Breedveld, M.W.; Canter Cremers, H.C.J.; Batley, M.; Posthumus, M.A.; Zevenhuizen, L.P.T.M.; Wijfelman, C.A.; Zehnder, A.J.B. Polysaccharide synthesis in relation to nodulation behaviour of Rhizobium leguminosarum. J. Bacteriol. 1993, 175, 750–757. [Google Scholar] [CrossRef] [Green Version]
- McNeil, M.; Darvill, J.; Darvill, A.; Albersheim, P.; van Veen, R.; Hooykaas, P.; Schilperoort, R.; Dell, A. The discernible structural features of the acidic exopolysaccharides secreted by different Rhizobium species are the same. Carbohydr. Res. 1986, 146, 307–326. [Google Scholar] [CrossRef]
- Spaink, H.P. Root nodulation and infection factors produced by rhizobial bacteria. Annu. Rev. Microbiol. 2000, 54, 257–288. [Google Scholar] [CrossRef] [Green Version]
- Schmid, J. Recent insights in microbial exopolysaccharide biosynthesis and engineering strategies. Curr. Opin. Biotechnol. 2018, 53, 130–136. [Google Scholar] [CrossRef]
- Finan, T.M.; Weidner, S.; Womg, K.; Buhrmester, J.; Chain, P.; Vorhölter, F.J.; Hernandez-Lucas, I.; Becker, A.; Cowie, A.; Gouzy, J.; et al. The complete sequence of the 1,683-kb pSymB megaplasmid from the N2-fixing endosymbiont Sinorhizobium meliloti. Proc. Natl. Acad. Sci. USA 2001, 98, 9889–9894. [Google Scholar] [CrossRef] [Green Version]
- Young, J.P.W.; Crossman, L.C.; Johnston, A.W.B.; Thomson, N.R.; Ghazoui, Z.F.; Hull, K.H.; Wexler, M.; Curson, A.R.; Todd, J.D.; Poole, P.S.; et al. The genome of Rhizobium leguminosarum has recognizable core and accessory components. Genome Biol. 2006, 7, R34. [Google Scholar] [CrossRef] [Green Version]
- González, V.; Santamaría, R.I.; Bustos, P.; Hernández-González, I.; Medrano-Soto, A.; Moreno-Hagelsieb, G.; Janga, S.C.; Ramírez, M.A.; Jiménez-Jacinto, V.; Collado-Vides, J.; et al. The partitioned Rhizobium etli genome: Genetic and metabolic redundancy in seven interacting replicons. Proc. Natl. Acad. Sci. USA 2006, 103, 3834–3839. [Google Scholar] [CrossRef] [Green Version]
- Kaneko, T.; Nakamura, Y.; Sato, S.; Minamisawa, K.; Uchiumi, T.; Sasamoto, S.; Watanabe, A.; Idesawa, K.; Iriguchi, M.; Kawashima, K.; et al. Complete genomic sequence of nitrogen-fixing symbiotic bacterium Bradyrhizobium japonicum USDA110. DNA Res. 2002, 9, 225–256. [Google Scholar] [CrossRef] [Green Version]
- Król, J.E.; Mazur, A.; Marczak, M.; Skorupska, A. Syntenic arrangements of the surface polysaccharide biosynthesis genes in Rhizobium leguminosarum. Genomics 2007, 89, 237–247. [Google Scholar] [CrossRef] [Green Version]
- Janczarek, M.; Rachwał, K.; Kopcińska, J. Genetic characterization of the Pss region and the role of PssS in exopolysaccharide production and symbiosis of Rhizobium leguminosarum bv. trifolii with clover. Plant Soil 2015, 396, 257–275. [Google Scholar] [CrossRef] [Green Version]
- Janczarek, M.; Rachwal, K.; Turska-Szewczuk, A. A mutation in pssE affects exopolysaccharide synthesis by Rhizobium leguminosarum bv. trifolii, its surface properties, and symbiosis with clover. Plant Soil 2017, 417, 331–347. [Google Scholar] [CrossRef]
- Rachwał, K.; Matczyńska, E.; Janczarek, M. Transcriptome profiling of a Rhizobium leguminosarum bv. trifolii rosR mutant reveals the role of the transcriptional regulator RosR in motility, synthesis of cell-surface components, and other cellular processes. BMC Genom. 2015, 16, 1111. [Google Scholar] [CrossRef] [Green Version]
- Reeve, W.; O’Hara, G.; Chain, P.; Ardley, J.; Bräu, L.; Nandesena, K.; Tiwari, R.; Copeland, A.; Nolan, M.; Han, C.; et al. Complete genome sequence of Rhizobium leguminosarum bv. trifolii strain WSM1325, an effective microsymbiont of annual Mediterranean clovers. Stand. Genom. Sci. 2010, 2, 347–356. [Google Scholar] [CrossRef]
- Reeve, W.; O’Hara, G.; Chain, P.; Ardley, J.; Bräu, L.; Nandesena, K.; Tiwari, R.; Malfatti, S.; Kiss, H.; Lapidus, A.; et al. Complete genome sequence of Rhizobium leguminosarum bv. trifolii strain WSM2304, an effective microsymbiont of the South American clover Trifolium polymorphum. Stand. Genom. Sci. 2010, 2, 66–76. [Google Scholar] [CrossRef]
- Sadykov, M.R.; Ivashina, T.V.; Kanapin, A.A.; Shlyapnikov, M.G.; Ksenzenko, V.N. Structural and functional organization of the exopolysaccharide biosynthesis genes in Rhizobium leguminosarum bv. viciae VF39. Mol. Biol. 1998, 32, 665–671. [Google Scholar]
- Bonomi, H.R.; Posadas, D.M.; Paris, G.; Carrica, M.D.; Frederickson, M.; Pietrasanta, L.I.; Bogomolni, R.A.; Zorreguieta, A.; Goldbaum, F.A. Light regulates attachment, exopolysaccharide production, and nodulation in Rhizobium leguminosarum through a LOV-histidine kinase photoreceptor. Proc. Natl. Acad. Sci. USA 2012, 109, 12135–12140. [Google Scholar] [CrossRef] [Green Version]
- Cieśla, J.; Kopycińska, M.; Łukowska, M.; Bieganowski, A.; Janczarek, M. Surface properties of wild-type Rhizobium leguminosarum bv. trifolii strain 24.2 and its derivatives with different extracellular polysaccharide content. PLoS ONE 2016, 11, e0165080. [Google Scholar] [CrossRef]
- Kopycińska, M.; Lipa, P.; Cieśla, J.; Kozieł, M.; Janczarek, M. Extracellular polysaccharide protects Rhizobium leguminosarum cells against zinc stress in vitro and during symbiosis with clover. Environ. Microbiol. Rep. 2018, 10, 355–368. [Google Scholar] [CrossRef] [PubMed]
- Janczarek, M.; Urbanik-Sypniewska, T. Expression of the Rhizobium leguminosarum bv. trifolii pssA gene involved in exopolysaccharide synthesis is regulated by RosR, phosphate and the carbon source. J. Bacteriol. 2013, 195, 3412–3423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janczarek, M.; Rachwał, K.; Cieśla, J.; Ginalska, G.; Bieganowski, J. Production of exopolysaccharide by Rhizobium leguminosarum bv. trifolii and its role in bacterial attachment and surface properties. Plant Soil 2015, 388, 211–227. [Google Scholar] [CrossRef] [Green Version]
- Jaszek, M.; Janczarek, M.; Kuczyński, K.; Piersiak, T.; Grzywnowicz, K. The response of the Rhizobium leguminosarum bv. trifolii wild-type and exopolysaccharide-deficient mutants to oxidative stress. Plant Soil 2014, 376, 75–94. [Google Scholar] [CrossRef] [Green Version]
- Muszyński, A.; Laus, M.; Kijne, J.W.; Carlson, R.W. The structures of the lipopolysaccharides from Rhizobium leguminosarum RBL5523 and its UDP-glucose dehydrogenase mutant (exo5). Glycobiology 2011, 21, 55–68. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Navarro, D.N.; Rodriguez-Carvajal, M.A.; Acosta-Jurado, S.; Soto, M.J.; Margaret, I.; Crespo-Rivas, J.C.; Sanjuan, J.; Temprano, F.; Gil-Serrano, A.; Ruiz-Sainz, J.E.; et al. Structure and biological roles of Sinorhizobium fredii HH103 exopolysaccharide. PLoS ONE 2014, 9, e115391. [Google Scholar] [CrossRef] [Green Version]
- Djordjevic, S.P.; Batley, M.; Redmond, J.W.; Rolfe, B.G. The structure of the exopolysaccharide from Rhizobium sp. strain ANU280 (NGR234). Carbohydr. Res. 1986, 148, 87–99. [Google Scholar] [CrossRef]
- Kelly, S.J.; Muszyński, A.; Kawaharada, Y.; Hubber, A.M.; Sullivan, J.T.; Sandal, N.; Carlson, R.W.; Stougaard, J.; Ronson, C.W. Conditional requirement for exopolysaccharide in the Mesorhizobium–Lotus symbiosis. Mol. Plant-Microbe Interact. 2013, 26, 319–329. [Google Scholar] [CrossRef] [Green Version]
- Canter Cremers, H.C.J.; Batley, M.; Redmond, J.W.; Eysdems, L.; Breedveld, M.W.; Zevenhuizen, L.P.T.M.; Pees, E.; Wijffelman, C.A.; Lugtenberg, B.J.J. Rhizobium leguminosarum exoB mutants are deficient in the synthesis of UDP-glucose 4′-epimerase. J. Biol. Chem. 1990, 265, 21122–21127. [Google Scholar] [CrossRef]
- Sánchez-Andújar, B.; Coronado, C.; Philip-Hollingsworth, S.; Dazzo, F.B.; Palomares, A.J. Structure and role in symbiosis of the exoB gene of Rhizobium leguminosarum bv. trifolii. Mol. Gen. Genet. 1997, 255, 131–140. [Google Scholar] [CrossRef]
- Pollock, T.J.; Workum, W.A.; Thorne, L.; Mikolajczak, M.J.; Yamazaki, M.; Kijne, J.W.; Armentrout, R.W. Assignment of biochemical functions to glycosyl transferase genes which are essential for biosynthesis of exopolysaccharides in Sphingomonas strain S88 and Rhizobium leguminosarum. J. Bacteriol. 1998, 180, 586–593. [Google Scholar] [CrossRef] [Green Version]
- Rolfe, B.G.; Carlson, R.W.; Ridge, R.W.; Dazzo, F.B.; Mateos, P.F.; Pankhurst, C.E. Defective infection and nodulation of clovers by exopolysaccharide mutants of Rhizobium leguminosarum bv. trifolii. Aust. J. Plant Physiol. 1996, 23, 285–303. [Google Scholar] [CrossRef]
- van Workum, W.A.; Canter Cremers, H.C.J.; Wijfjes, A.H.M.; van der Kolk, C.; Wijffelman, C.A.; Kijne, J.W. Cloning and characterization of four genes of Rhizobium leguminosarum bv. trifolii involved in exopolysaccharide production and nodulation. Mol. Plant Microbe Interact. 1997, 10, 290–301. [Google Scholar] [CrossRef]
- Ivashina, T.V.; Khmelnitsky, M.I.; Shlyapnikov, M.G.; Kanapin, A.A.; Ksenzenko, V.N. The pss4 gene from Rhizobium leguminosarum biovar viciae VF39: Cloning, sequence and the possible role in polysaccharide production and nodule formation. Gene 1994, 50, 111–116. [Google Scholar] [CrossRef]
- Janczarek, M.; Król, J.; Skorupska, A. The pssB gene product of Rhizobium leguminosarum bv. trifolii is homologous to a family of inositol monophosphatases. FEMS Microbiol. Lett. 1999, 173, 319–325. [Google Scholar] [CrossRef] [Green Version]
- Janczarek, M.; Rachwał, K. Mutation in the pssA gene involved in exopolysaccharide synthesis leads to several physiological and symbiotic defects in Rhizobium leguminosarum bv. trifolii. Int. J. Mol. Sci. 2013, 14, 3711–3735. [Google Scholar] [CrossRef] [Green Version]
- Latchford, J.W.; Borthakur, D.; Johnston, A.W.B. The products of Rhizobium genes, psi and pss, which affect exopolysaccharide production, are associated with the bacterial cell surface. Mol. Microbiol. 1991, 5, 2107–2114. [Google Scholar] [CrossRef]
- Janczarek, M.; Skorupska, A. Regulation of pssA and pssB gene expression in R. leguminosarum bv. trifolii in response to environmental factors. Antonie Van Leeuwenhoek 2004, 85, 217–227. [Google Scholar] [CrossRef]
- Lipa, P.; Vinardell, J.M.; Janczarek, M. Transcriptomic studies reveal that the Rhizobium leguminosarum Serine/Threonine Protein Phosphatase PssZ has a role in the synthesis of cell-surface components, nutrient utilization, and other cellular processes. Int. J. Mol. Sci. 2019, 20, 2905. [Google Scholar] [CrossRef] [Green Version]
- Król, J.; Wielbo, J.; Mazur, A.; Kopcińska, J.; Łotocka, B.; Golinowski, W.; Skorupska, A. Molecular characterization of pssCDE Genes of Rhizobium leguminosarum bv. trifolii strain TA1: pssD mutant is affected in exopolysaccharide synthesis and endocytosis of bacteria. Mol. Plant Microbe Interact. 1998, 11, 1142–1148. [Google Scholar] [CrossRef] [PubMed]
- Ivashina, T.V.; Ksenzenko, V.N. Exopolysaccharide biosynthesis in Rhizobium leguminosarum: From genes to functions. In The Complex World of Polysaccharides; Karunaratne, D.N., Ed.; InTech: Rijeka, Croatia, 2012; pp. 99–126. [Google Scholar] [CrossRef] [Green Version]
- Marczak, M.; Mazur, A.; Koper, P.; Żebracki, K.; Skorupska, A. Synthesis of rhizobial exopolysaccharides and their importance for symbiosis with legume plants. Genes 2017, 8, 360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marczak, M.; Wójcik, M.; Żebracki, K.; Turska-Szewczuk, A.; Talarek, K.; Nowak, D.; Wawiórka, L.; Sieńczyk, M.; Łupicka-Słowik, A.; Bobrek, K.; et al. PssJ is a terminal galactosyltransferase involved in the assembly of the exopolysaccharide subunit in Rhizobium leguminosarum bv. trifolii. Int. J. Mol. Sci. 2020, 21, 7764. [Google Scholar] [CrossRef] [PubMed]
- Ivashina, T.V.; Fedorova, E.E.; Ashina, N.P.; Kalinchuk, N.A.; Druzhinina, T.N.; Shashkov, A.S.; Shibaev, V.N.; Ksenzenko, V.N. Mutation in the pssM gene encoding ketal pyruvate transferase leads to disruption of Rhizobium leguminosarum bv. viciae-Pisum sativum symbiosis. J. Appl. Microbiol. 2010, 109, 731–742. [Google Scholar] [CrossRef]
- Mazur, A.; Król, J.E.; Marczak, M.; Skorupska, A. Membrane topology of PssT, the transmembrane protein component of type I exopolysaccharide transport system in the Rhizobium leguminosarum bv. trifolii strain TA1. J. Bacteriol. 2003, 185, 2503–2511. [Google Scholar] [CrossRef] [Green Version]
- Mazur, A.; Król, J.E.; Wielbo, J.; Urbanik-Sypniewska, T.; Skorupska, A. Rhizobium leguminosarum bv. trifolii PssP protein is required for exopolysaccharide biosynthesis and polymerization. Mol. Plant Microbe Interact. 2002, 15, 388–397. [Google Scholar] [CrossRef] [Green Version]
- Grangeasse, C.; Terreux, R.; Nessler, S. Bacterial tyrosine-kinases: Structure-function analysis and therapeutic potential. Biochim. Biophys. Acta 2010, 1804, 628–634. [Google Scholar] [CrossRef]
- Janczarek, M.; Vinardell, J.M.; Lipa, P.; Karaś, M. Hanks-type serine/threonine protein kinases and phosphatases in bacteria: Roles in signaling and adaptation to various environments. Int. J. Mol. Sci. 2018, 19, 2872. [Google Scholar] [CrossRef] [Green Version]
- Lipa, M.; Janczarek, M. Phosphorylation systems in symbiotic nitrogen-fixing bacteria and their role in bacterial adaptation to various environmental stresses. PeerJ. 2020, 8, e8466. [Google Scholar] [CrossRef] [Green Version]
- Collins, R.F.; Derrick, J.P. Wza: A new structural paradigm for outer membrane secretory proteins? Trends Microbiol. 2007, 15, 96–100. [Google Scholar] [CrossRef]
- Marczak, M.; Mazur, A.; Król, J.E.; Gruszecki, W.I.; Skorupska, A. Lipoprotein PssN of Rhizobium leguminosarum bv. trifolii: Subcellular localization and possible involvement in exopolysaccharide export. J. Bacteriol. 2006, 188, 6943–6952. [Google Scholar] [CrossRef] [Green Version]
- Zorreguieta, A.; Finnie, C.; Downie, J.A. Extracellular glycanases of Rhizobium leguminosarum are activated on the cell surface by an exopolysaccharide-related component. J. Bacteriol. 2000, 182, 1304–1312. [Google Scholar] [CrossRef] [Green Version]
- Finnie, C.; Hartley, N.M.; Findlay, K.C.; Downie, J.A. The Rhizobium leguminosarum prsDE genes are required for secretion of several proteins, some of which influence nodulation, symbiotic nitrogen fixation and exopolysaccharide modification. Mol. Microbiol. 1997, 25, 135–146. [Google Scholar] [CrossRef]
- Finnie, C.; Zorreguieta, A.; Hartley, N.M.; Downie, J.A. Characterization of Rhizobium leguminosarum exopolysaccharide glycanases that are secreted via a type I exporter and have a novel heptapeptide repeat motif. J. Bacteriol. 1998, 180, 1691–1969. [Google Scholar] [CrossRef] [Green Version]
- Ausmees, N.; Jacobsson, K.; Lindberg, M. A unipolarly located, cell-surface-associated agglutinin, RapA, belongs to a family of Rhizobium-adhering proteins (Rap) in Rhizobium leguminosarum bv. trifolii. Microbiology 2001, 147, 549–559. [Google Scholar] [CrossRef] [Green Version]
- Krehenbrink, M.; Downie, J.A. Identification of protein secretion systems and novel secreted proteins in Rhizobium leguminosarum bv. viciae. BMC Genom. 2008, 9, 55. [Google Scholar] [CrossRef] [Green Version]
- Lipa, P.; Vinardell, J.M.; Kopcińska, J.; Zdybicka-Barabas, A.; Janczarek, M. Mutation in the pssZ gene negatively impacts exopolysaccharide synthesis, surface properties, and symbiosis of Rhizobium leguminosarum bv. trifolii with clover. Genes 2018, 9, 369. [Google Scholar] [CrossRef] [Green Version]
- Mijakovic, I.; Grangeasse, C.; Turgay, K. Exploring the diversity of protein modifications: Special bacterial phosphorylation systems. FEMS Microbiol. Rev. 2016, 40, 398–417. [Google Scholar] [CrossRef] [Green Version]
- Janczarek, M.; Skorupska, A. The Rhizobium leguminosarum bv. trifolii RosR: Transcriptional regulator involved in exopolysaccharide production. Mol. Plant Microbe Interact. 2007, 20, 867–881. [Google Scholar] [CrossRef] [Green Version]
- Janczarek, M.; Skorupska, A. Rhizobium leguminosarum bv. trifolii rosR gene expression is regulated by catabolic repression. FEMS Microbiol. Lett. 2009, 291, 112–119. [Google Scholar] [CrossRef]
- Janczarek, M.; Kalita, M.; Skorupska, A. New taxonomic markers for identification of Rhizobium leguminosarum and discrimination between closely related species. Arch. Microbiol. 2009, 191, 207–219. [Google Scholar] [CrossRef] [PubMed]
- Bittinger, M.A.; Handelsman, J. Identification of genes in the RosR regulon of Rhizobium etli. J. Bacteriol. 2000, 182, 1706–1713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chou, A.Y.; Archdeacon, J.; Kado, C.I. Agrobacterium transcriptional regulator Ros is a prokaryotic zinc finger protein that regulates the plant oncogene ipt. Proc. Natl. Acad. Sci. USA 1998, 95, 5293–5298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keller, M.; Roxlau, A.; Wenig, W.M.; Schmidt, M.; Quandt, J.; Niehaus, K.; Jording, D.; Arnold, W.; Pühler, A. Molecular analysis of the Rhizobium meliloti mucR gene regulating the biosynthesis of the exopolysaccharides succinoglycan and galactoglucan. Mol. Plant Microbe Interact. 1995, 8, 267–277. [Google Scholar] [CrossRef]
- Acosta-Jurado, S.; Alias-Villegas, C.; Navarro-Gomez, P.; Zehner, S.; Murdoch, P.D.; Rodriguez-Carvajal, M.A.; Soto, M.J.; Ollero, F.J.; Ruiz-Sainz, J.E.; Gottfert, M.; et al. The Sinorhizobium fredii HH103 MucR1 global regulator is connected with the nod regulon and is required for efficient symbiosis with Lotus burttii and Glycine max cv. Williams. Mol. Plant Microbe Interact. 2016, 29, 700–712. [Google Scholar] [CrossRef] [Green Version]
- Jiao, J.; Wu, L.J.; Zhang, B.; Hu, Y.; Li, Y.; Zhang, X.X.; Guo, H.J.; Liu, L.X.; Chen, W.X.; Zhang, Z.; et al. MucR is required for transcriptional activation of conserved ion transporters to support nitrogen fixation of Sinorhizobium fredii in soybean nodules. Mol. Plant Microbe Interact. 2016, 29, 352–361. [Google Scholar] [CrossRef] [Green Version]
- Pirone, L.; Pitzer, J.E.; D’Abrosca, G.; Fattorusso, R.; Malgieri, G.; Pedone, E.M.; Pedone, P.V.; Roop, R.M., II; Baglivo, I. Identifying the region responsible for Brucella abortus MucR higher-order oligomer formation and examining its role in gene regulation. Sci. Rep. 2018, 8, 17238. [Google Scholar] [CrossRef] [Green Version]
- Janczarek, M.; Kutkowska, J.; Piersiak, T.; Skorupska, A. Rhizobium leguminosarum bv. trifolii rosR is required for interaction with clover, biofilm formation and adaptation to the environment. BMC Microbiol. 2010, 10, 284. [Google Scholar] [CrossRef] [Green Version]
- Rachwał, K.; Boguszewska, A.; Kopcińska, J.; Karaś, M.; Tchórzewski, M.; Janczarek, M. The regulatory protein RosR affects Rhizobium leguminosarum bv. trifolii protein profiles, cell surface properties, and symbiosis with clover. Front. Microbiol. 2016, 7, 1302. [Google Scholar] [CrossRef] [Green Version]
- Janczarek, M.; Jaroszuk-Ściseł, J.; Skorupska, A. Multiple copies of rosR and pssA genes enhance exopolysaccharide production, symbiotic competitiveness and clover nodulation in Rhizobium leguminosarum bv. trifolii. Antonie Van Leeuwenhoek 2009, 96, 471–486. [Google Scholar] [CrossRef]
- Janczarek, M.; Skorupska, A. Modulation of rosR expression and exopolysaccharide production in Rhizobium leguminosarum bv. trifolii by phosphate and clover root exudates. Int. J. Mol. Sci. 2011, 12, 4132–4155. [Google Scholar] [CrossRef] [Green Version]
- Rachwał, K.; Lipa, P.; Wojda, I.; Vinardell, J.-M.; Janczarek, M. Regulatory elements located in the upstream region of the Rhizobium leguminosarum rosR global regulator are essential for its transcription and mRNA stability. Genes 2017, 8, 388. [Google Scholar] [CrossRef] [Green Version]
- Wielbo, J.; Mazur, A.; Krol, J.E.; Marczak, M.; Skorupska, A. Environmental modulation of the pssTNOP gene expression in Rhizobium leguminosarum bv. trifolii. Can. J. Microbiol. 2004, 50, 201–211. [Google Scholar] [CrossRef]
- Mueller, K.; González, J.E. Complex regulation of symbiotic functions is coordinated by MucR and quorum sensing in Sinorhizobium meliloti. J. Bact. 2011, 193, 485–496. [Google Scholar] [CrossRef] [Green Version]
- Reeve, W.G.; Dilworth, M.J.; Tiwari, R.P.; Glenn, A.R. Regulation of exopolysaccharide production in Rhizobium leguminosarum biovar viciae WSM710 involves exoR. Microbiology 1997, 143, 1951–1958. [Google Scholar] [CrossRef] [Green Version]
- Ivashina, T.; Sadykov, M.P.; Thatujev, B.M.; Kanapin, A.A.; Shlapnikov, M.G.; Ksenzenko, W. The Rhizobium leguminosarum bv viciae VF39 pssB gene product participates in symbiotic fixation of molecular nitrogen. Dokl. Akad. Nauk. 1996, 350, 712–715. [Google Scholar]
- Kutkowska, J.; Janczarek, M.; Kopcińska, J.; Urbanik-Sypniewska, T.; Skorupska, A. Effects of pssB mutation on surface polysaccharides and symbiotic phenotype of Rhizobium leguminosarum bv. trifolii. Acta Biol. Cracov. Bot. 2007, 49, 81–89. [Google Scholar]
- Janczarek, M.; Skorupska, A. The Rhizobium leguminosarum bv. trifolii pssB gene product is an inositol monophosphatase that influences exopolysaccharide synthesis. Arch. Microbiol. 2001, 175, 143–151. [Google Scholar] [CrossRef]
- Borthakur, D.; Johnston, A.W.B. Sequence of psi, a gene of the symbiotic plasmid of Rhizobium phaseoli which inhibits exopolysaccharide synthesis and nodulation and demonstration that its transcription is inhibited by psr, another gene on the symbiotic plasmid. Mol. Gen. Genet. 1987, 207, 149–154. [Google Scholar] [CrossRef]
- Mimmack, M.L.; Borthakur, D.; Jones, M.A.; Downie, J.A.; Johnston, A.W.B. The psi operon of Rhizobium leguminosarum biovar phaseoli: Identification of two genes whose products are located at the bacterial cell surface. Microbiology 1994, 140, 1223–1229. [Google Scholar] [CrossRef] [Green Version]
- Mimmack, M.L.; Hong, G.F.; Johnston, A.W.B. Sequence and regulation of psrA, a gene on the Sym plasmid of Rhizobium leguminosarum biovar phaseoli which inhibits transcription of the psi genes. Microbiology 1994, 140, 455–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wisniewski-Dyé, F.; Downie, J.A. Quorum sensing in Rhizobium. Antonie Van Leeuwenhoek 2002, 81, 397–407. [Google Scholar] [CrossRef] [PubMed]
- Edwards, A.; Frederix, M.; Wisniewski-Dyé, F.; Jones, J.; Zorreguieta, A.; Downie, J.A. The cin and rai quorum-sensing regulatory systems in Rhizobium leguminosarum are coordinated by ExpR and CinS, a small regulatory protein coexpressed with CinI. J. Bacteriol. 2009, 191, 3059–3067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frederix, M.; Edwards, A.; McAnulla, C.; Downie, J.A. Co-ordination of quorum-sensing regulation in Rhizobium leguminosarum by induction of an anti-repressor. Mol. Microbiol. 2011, 81, 994–1007. [Google Scholar] [CrossRef]
- Bogino, P.C.; Oliva, M.M.; Sorroche, F.G.; Giordano, W. The role of bacterial biofilms and surface components in plant-bacterial associations. Int. J. Mol. Sci. 2013, 14, 5838–5859. [Google Scholar] [CrossRef] [Green Version]
- D’Haeze, W.; Glushka, J.; De Rycke, R.; Holsters, M.; Carlson, R.W. Structural characterization of extracellular polysaccharides of Azorhizobium caulinodans and importance for nodule initiation on Sesbania rostrata. Mol. Microbiol. 2004, 52, 485–500. [Google Scholar] [CrossRef]
- Jiménez-Guerrero, I.; Acosta-Jurado, S.; del Cerro, P.; Navarro-Gómez, P.; López-Baena, F.J.; Ollero, F.J.; Vinardell, J.M.; Pérez-Montaño, F. Transcriptomic studies of the effect of nod gene-inducing molecules in rhizobia: Different weapons, one purpose. Genes 2018, 9, 1. [Google Scholar] [CrossRef] [Green Version]
- Galibert, F.; Finan, T.M.; Long, S.R.; Puhler, A.; Abola, P.; Ampe, F.; Barloy-Hubler, F.; Barnett, M.J.; Becker, A.; Boistard, P.; et al. The composite genome of the legume symbiont Sinorhizobium meliloti. Science 2001, 293, 668–672. [Google Scholar] [CrossRef] [Green Version]
- Barnett, M.J.; Long, S.R. The Sinorhizobium meliloti SyrM regulon: Effects on global gene expression are mediated by syrA and nodD3. J. Bacteriol. 2015, 197, 1792–1806. [Google Scholar] [CrossRef] [Green Version]
- Barnett, M.J.; Swanson, J.A.; Long, S.R. Multiple genetic controls on Rhizobium meliloti syrA, a regulator of exopolysaccharide abundance. Genetics 1998, 148, 19–32. [Google Scholar] [CrossRef]
- Cren, M.; Kondorosi, A.; Kondorosi, E. NolR controls expression of the Rhizobium meliloti nodulation genes involved in the core Nod factor synthesis. Mol. Microbiol. 1995, 15, 733–747. [Google Scholar] [CrossRef]
- Becker, A.; Rüberg, S.; Baumgarth, B.; Bertram-Drogatz, P.A.; Quester, I.; Pühler, A. Regulation of succinoglycan and galactoglucan biosynthesis in Sinorhizobium meliloti. J. Mol. Microbiol. Biotechnol. 2002, 4, 187–190. [Google Scholar]
- Janczarek, M. Environmental signals and regulatory pathways that influence exopolysaccharide production in rhizobia. Int. J. Mol. Sci. 2011, 12, 7898–7933. [Google Scholar] [CrossRef] [Green Version]
- Glenn, S.A.; Gurich, N.; Feeney, M.A.; Gonzalez, J.E. The ExpR/Sin quorum-sensing system controls succinoglycan production in Sinorhizobium meliloti. J. Bacteriol. 2007, 189, 7077–7088. [Google Scholar] [CrossRef] [Green Version]
- Zhan, H.J.; Lee, C.C.; Leigh, J.A. Induction of the second exopolysaccharide (EPSb) in Rhizobium meliloti SU47 by low phosphate concentrations. J. Bacteriol. 1991, 173, 7391–7394. [Google Scholar] [CrossRef] [Green Version]
- Leigh, J.A.; Walker, G.C. Exopolysaccharides of Rhizobium: Synthesis, regulation and symbiotic function. Trends Genet. 1994, 10, 63–67. [Google Scholar] [CrossRef]
- Mendrygal, K.E.; González, J.E. Environmental regulation of exopolysaccharide production in Sinorhizobium meliloti. J. Bacteriol. 2000, 182, 599–606. [Google Scholar] [CrossRef] [Green Version]
- Krol, E.; Becker, A. Global transcriptional analysis of the phosphate starvation response in Sinorhizobium meliloti strains 1021 and 2011. Mol. Genet. Genom. 2004, 272, 1–17. [Google Scholar] [CrossRef]
- Yao, S.Y.; Luo, L.; Har, K.J.; Becker, A.; Ruberg, S.; Yu, G.Q.; Zhu, J.B.; Cheng, H.P. Sinorhizobium meliloti ExoR and ExoS proteins regulate both succinoglycan and flagellum production. J. Bacteriol. 2004, 186, 6042–6049. [Google Scholar] [CrossRef] [Green Version]
- Wells, D.H.; Chen, E.J.; Fisher, R.F.; Long, S.R. ExoR is genetically coupled to the ExoS-ChvI two-component system and located in the periplasm of Sinorhizobium meliloti. Mol. Microbiol. 2007, 64, 647–664. [Google Scholar] [CrossRef]
- Chen, E.J.; Fisher, R.F.; Perovich, V.M.; Sabio, E.A.; Long, S.R. Identification of direct transcriptional target genes of ExoS/ChvI two-component signaling in Sinorhizobium meliloti. J. Bacteriol. 2009, 191, 6833–6842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Kemp, J.; Da Fonseca, I.O.; Equi, R.C.; Sheng, X.; Charles, T.C.; Sobral, B.W.S. Sinorhizobium meliloti 1021 loss-of-function deletion mutation in chvI and its phenotypic characteristics. Mol. Plant Microbe Interact. 2010, 23, 153–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibson, K.E.; Barnett, M.J.; Toman, C.J.; Long, S.R.; Walker, G.C. The symbiosis regulator CbrA modulates a complex regulatory network affecting the flagellar apparatus and cell envelope proteins. J. Bacteriol. 2007, 189, 3591–3602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, J.; González, J.E. The novel genes emmABC are associated with exopolysaccharide production, motility, stress adaptation, and symbiosis in Sinorhizobium meliloti. J. Bacteriol. 2009, 191, 5890–5900. [Google Scholar] [CrossRef] [Green Version]
- Reed, J.W.; Glazebrook, J.; Walker, G.C. The exoR gene of Rhizobium meliloti affects RNA levels of other exo genes but lacks homology to known transcriptional regulators. J. Bacteriol. 1991, 173, 3789–3794. [Google Scholar] [CrossRef] [Green Version]
- Becker, A.; Küster, H.; Niehaus, K.; Pühler, A. Extension of the Rhizobium meliloti succinoglycan biosynthesis gene cluster: Identification of the exsA gene encoding an ABC transporter protein, and the exsB gene, which probably codes for a regulator of succinoglycan biosynthesis. Mol. Gen. Genet. 1995, 249, 487–497. [Google Scholar] [CrossRef]
- Pinedo, C.A.; Gage, D.J. HPrK regulates succinate-mediated catabolite repression in the gram-negative symbiont Sinorhizobium meliloti. J. Bacteriol. 2009, 191, 298–309. [Google Scholar] [CrossRef] [Green Version]
- Bahlawane, C.; Baumgarth, B.; Serrania, J.; Rüberg, S.; Becker, A. Fine-Tuning of galactoglucan biosynthesis in Sinorhizobium meliloti by differential WggR (ExpG)-, PhoB-, and MucR-dependent regulation of two promoters. J. Bacteriol. 2008, 190, 3456–3466. [Google Scholar] [CrossRef] [Green Version]
- Calatrava-Morales, N.; McIntosh, M.; Soto, M.J. Regulation mediated by N-acyl homoserine lactone quorum sensing signals in the rhizobium-legume symbiosis. Genes 2018, 9, 263. [Google Scholar] [CrossRef] [Green Version]
- Hoang, H.H.; Becker, A.; Gonzalez, J.E. The LuxR homolog ExpR, in combination with the Sin quorum sensing system, plays a central role in Sinorhizobium meliloti gene expression. J. Bacteriol. 2004, 186, 5460–5472. [Google Scholar] [CrossRef] [Green Version]
- Pellock, B.J.; Teplitski, M.; Boinay, R.P.; Bauer, W.D.; Walker, G.C. A LuxR homolog controls production of symbiotically active extracellular polysaccharide II by Sinorhizobium meliloti. J. Bacteriol. 2002, 184, 5067–5076. [Google Scholar] [CrossRef] [Green Version]
- McIntosh, M.; Krol, E.; Becker, A. Competitive and cooperative effects in quorum-sensing-regulated galactoglucan biosynthesis in Sinorhizobium meliloti. J. Bacteriol. 2008, 190, 5308–5317. [Google Scholar] [CrossRef] [Green Version]
- Buendia, A.M.; Enenkel, B.; Köplin, R.; Niehaus, K.; Arnold, W.; Pühler, A. The Rhizobium meliloti exoZ/exoB fragment of megaplasmid 2: ExoB functions as a UDP-glucose 4-epimerase and ExoZ shows homology to NodX of Rhizobium leguminosarum biovar viciae strain TOM. Mol. Microbiol. 1991, 5, 1519–1530. [Google Scholar] [CrossRef]
- Müller, P.; Hynes, M.; Kapp, D.; Niehaus, K.; Pühler, A. Two classes of Rhizobium meliloti infection mutants differ in exopolysaccharide production and in coinoculation properties with nodulation mutants. Mol. Gen. Genet. 1988, 211, 17–26. [Google Scholar] [CrossRef]
- Kapp, D.; Niehaus, K.; Quandt, J.; Muller, P.; Pühler, A. Cooperative action of Rhizobium meliloti nodulation and infection mutants during the process of forming mixed infected alfalfa nodules. Plant Cell 1990, 2, 139–151. [Google Scholar] [CrossRef] [Green Version]
- Becker, A.; Kleickmann, A.; Küster, H.; Keller, M.; Arnold, W.; Pühler, A. Analysis of the Rhizobium meliloti genes exoU, exoV, exoW, exoT, and exoI involved in exopolysaccharide biosynthesis and nodule invasion: exoU and exoW probably encode glucosyltransferases. Mol. Plant Microbe Interact. 1993, 6, 735–744. [Google Scholar] [CrossRef]
- Pellock, B.J.; Cheng, H.P.; Walker, G.C. Alfalfa root nodule invasion efficiency is dependent on Sinorhizobium meliloti polysaccharides. J. Bacteriol. 2000, 182, 4310–4318. [Google Scholar] [CrossRef] [Green Version]
- Fraysse, N.; Lindner, B.; Kaczynski, Z.; Sharypova, L.; Holst, O.; Niehaus, K.; Poinsot, V. Sinorhizobium meliloti strain 1021 produces a low-molecular-mass capsular polysaccharide that is a homopolymer of 3-deoxy-D-manno-oct-2-ulosonic acid harboring a phospholipid anchor. Glycobiology 2005, 15, 101–108. [Google Scholar] [CrossRef]
- Putnoky, P.; Petrovics, G.; Kereszt, A.; Grosskopf, E.; Ha, D.T.; Bánfalvi, Z.; Kondorosi, A. Rhizobium meliloti lipopolysaccharide and exopolysaccharide can have the same function in the plant-bacterium interaction. J. Bacteriol. 1990, 172, 5450–5458. [Google Scholar] [CrossRef] [Green Version]
- Jones, K.M.; Sharopova, N.; Lohar, D.P.; Zhang, J.Q.; VandenBosch, K.A.; Walker, G.C. Differential response of the plant Medicago truncatula to its symbiont Sinorhizobium meliloti or an exopolysaccharide- deficient mutant. Proc. Natl. Acad. Sci. USA 2008, 105, 704–709. [Google Scholar] [CrossRef] [Green Version]
- Barnett, M.J.; Long, S.R. Novel genes and regulators that influence production of cell surface exopolysaccharides in Sinorhizobium meliloti. J. Bacteriol. 2018, 200, e00501-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, R.; Herouart, D.; Sigaud, S.; Touati, D.; Puppo, A. Oxidative burst in alfalfa–Sinorhizobium meliloti symbiotic interaction. Mol. Plant Microbe Interact. 2001, 14, 86–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maroti, G.; Downie, J.A.; Kondorosi, E. Plant cysteine-rich peptides that inhibit pathogen growth and control rhizobial differentiation in legume nodules. Curr. Opin. Plant Biol. 2015, 26, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Rinaudi, L.V.; Giordano, W. An integrated view of biofilm formation in rhizobia. FEMS Microbiol. Lett. 2010, 304, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Margaret, I.; Becker, A.; Blom, J.; Bonilla, I.; Goesmann, A.; Göttfert, M.; Lloret, J.; Mittard-Runte, V.; Rückert, C.; Ruiz-Sainz, J.E.; et al. Symbiotic properties and first analyses of the genomic sequence of the fast growing model strain Sinorhizobium fredii HH103 nodulating soybean. J. Biotechnol. 2011, 155, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Dowdle, S.F.; Bohlool, B.B. Predominance of fast-growing Rhizobium japonicum in a soybean field in the People’s Republic of China. Appl. Environ. Microbiol. 1985, 50, 1171–1176. [Google Scholar] [CrossRef] [Green Version]
- Pueppke, S.G.; Broughton, W.J. Rhizobium sp. strain NGR234 and R. fredii USDA257 share exceptionally broad: Nested host ranges. Mol. Plant Microbe Interact. 1999, 12, 293–318. [Google Scholar] [CrossRef] [Green Version]
- Temprano-Vera, F.; Rodríguez-Navarro, D.N.; Acosta-Jurado, S.; Perret, X.; Fossou, R.K.; Navarro-Gómez, P.; Zhen, T.; Yu, D.; An, Q.; Buendía-Clavería, A.M.; et al. Sinorhizobium fredii Strains HH103 and NGR234 form nitrogen fixing nodules with diverse wild soybeans (Glycine soja) from Central China but are ineffective on Northern China accessions. Front. Microbiol. 2018, 9, 2843. [Google Scholar] [CrossRef]
- Tian, C.F.; Zhou, Y.J.; Zhang, Y.M.; Li, Q.Q.; Zhang, Y.Z.; Li, D.F.; Wang, S.; Wang, J.; Gilbert, L.B.; Li, Y.R.; et al. Comparative genomics of rhizobia nodulating soybean suggests extensive recruitment of lineage-specific genes in adaptations. Proc. Natl. Acad. Sci. USA 2012, 109, 8629–8634. [Google Scholar] [CrossRef] [Green Version]
- Weidner, S.; Becker, A.; Bonilla, I.; Jaenicke, S.; Lloret, J.; Margaret, I.; Pühler, A.; Ruiz-Sainz, J.E.; Schneiker-Bekel, S.; Szczepanowski, R.; et al. Genome sequence of the soybean symbiont Sinorhizobium fredii HH103. J. Bacteriol. 2012, 194, 1617–1618. [Google Scholar] [CrossRef] [Green Version]
- Vinardell, J.M.; Acosta-Jurado, S.; Zehner, S.; Gottfert, M.; Becker, A.; Baena, I.; Blom, J.; Crespo-Rivas, J.C.; Goesmann, A.; Jaenicke, S.; et al. The Sinorhizobium fredii HH103 genome: A comparative analysis with S. fredii strains differing in their symbiotic behavior with soybean. Mol. Plant Microbe Interact. 2015, 28, 811–824. [Google Scholar] [CrossRef] [Green Version]
- Perez-Montano, F.; Jimenez-Guerrero, I.; Acosta-Jurado, S.; Navarro-Gomez, P.; Ollero, F.J.; Ruiz-Sainz, J.E.; Lopez-Baena, F.J.; Vinardell, J.M. A transcriptomic analysis of the effect of genistein on Sinorhizobium fredii HH103 reveals novel rhizobial genes putatively involved in symbiosis. Sci. Rep. 2016, 7, 31592. [Google Scholar] [CrossRef] [Green Version]
- Acosta-Jurado, S.; Alias-Villegas, C.; Navarro-Gomez, P.; Almozara, A.; Rodriguez-Carvajal, M.A.; Medina, C.; Vinardell, J.M. Sinorhizobium fredii HH103 syrM inactivation affects the expression of a large number of genes, impairs nodulation with soybean and extends the host-range to Lotus japonicus. Environ. Microbiol. 2020, 22, 1104–1124. [Google Scholar] [CrossRef]
- Machado, D.; Pueppke, S.G.; Vinardell, J.M.; Ruiz-Sainz, J.E.; Krishnan, H.B. Expression of nodD1 and nodD2 in Sinorhizobium fredii, a nitrogen-fixing symbiont of soybean and other legumes. Mol. Plant Microbe Interact. 1998, 11, 375–382. [Google Scholar] [CrossRef] [Green Version]
- Vinardell, J.M.; Ollero, F.J.; Hidalgo, A.; López-Baena, F.J.; Medina, C.; Ivanov-Vangelov, K.; Parada, M.; Madinabeitia, N.; Espuny, M.R.; Bellogín, R.A.; et al. NolR regulates diverse symbiotic signals of Sinorhizobium fredii HH103. Mol. Plant Microbe Interact. 2004, 17, 676–685. [Google Scholar] [CrossRef] [Green Version]
- Acosta-Jurado, S.; Rodriguez-Navarro, D.N.; Kawaharada, Y.; Rodriguez-Carvajal, M.A.; Gil-Serrano, A.; Soria-Diaz, M.E.; Pérez-Montaño, F.; Fernández-Perea, J.; Niu, Y.; Alias-Villegas, C.; et al. Sinorhizobium fredii HH103 nolR and nodD2 mutants gain capacity for infection thread invasion of Lotus japonicus Gifu and Lotus burttii. Environ. Microbiol. 2019, 21, 1718–1739. [Google Scholar] [CrossRef]
- Acosta-Jurado, S.; Navarro-Gomez, P.; Murdoch, P.S.; Crespo-Rivas, J.C.; Jie, S.; Cuesta-Berrio, L.; Ruiz-Sainz, J.E.; Rodriguez-Carvajal, M.A.; Vinardell, J.M. Exopolysaccharide production by Sinorhizobium fredii HH103 is repressed by genistein in a NodD1-dependent manner. PLoS ONE 2016, 11, e0160499. [Google Scholar] [CrossRef]
- Streit, W.R.; Schmitz, R.A.; Perret, X.; Staehelin, C.; Deakin, W.J.; Raasch, C.; Liesegang, H.; Broughton, W.J. An evolutionary hot spot: The pNGR234b replicon of Rhizobium sp. strain NGR234. J. Bacteriol. 2004, 186, 535–542. [Google Scholar] [CrossRef] [Green Version]
- Acosta-Jurado, S.; Navarro-Gómez, P.; Crespo-Rivas, J.C.; Medina, C.; del Socorro Murdoch, P.; Cuesta-Berrio, L.; Rodríguez-Carvajal, M.A.; Ruiz-Sainz, J.E.; Vinardell, J.M. The Sinorhizobium (Ensifer) fredii HH103 rkp-2 region is involved in the biosynthesis of lipopolysaccharide and exopolysaccharide but not in K-antigen polysaccharide production. Plant Soil 2017, 417, 415–431. [Google Scholar] [CrossRef]
- Di Lorenzo, F.; Speciale, I.; Silipo, A.; Alías-Villegas, C.; Acosta-Jurado, S.; Rodríguez-Carvajal, M.Á.; Dardanelli, M.S.; Palmigiano, A.; Garozzo, D.; Ruiz-Sainz, J.E.; et al. Structure of the unusual Sinorhizobium fredii HH103 lipopolysaccharide and its role in symbiosis. J. Biol. Chem. 2020, 295, 10969–10987. [Google Scholar] [CrossRef]
- Kereszt, A.; Kiss, E.; Reuhs, B.L.; Carlson, R.W.; Kondorosi, A.; Putnoky, P. Novel rkp gene clusters of Sinorhizobium meliloti involved in capsular polysaccharide production and invasion of the symbiotic nodule: The rkpK gene encodes a UDP-glucose dehydrogenase. J. Bacteriol. 1998, 180, 5426–5431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunn, M.F.; Pueppke, A.G.; Krishnan, H.B. The nod gene inducer genistein alters the composition and molecular mass distribution of extracellular polysaccharides produced by Rhizobium fredii USDA 193. FEMS Microbiol. Lett. 1992, 97, 107–112. [Google Scholar] [CrossRef] [Green Version]
- Acosta-Jurado, S.; Alías-Villegas, C.; Almozara, A.; Espuny, M.R.; Vinardell, J.M.; Pérez-Montaño, F. Deciphering the symbiotic significance of quorum sensing systems of Sinorhizobium fredii HH103. Microorganisms 2020, 8, 68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krysciak, D.; Grote, J.; Rodriguez Orbegoso, M.; Utpatel, C.; Förstner, K.U.; Li, L.; Schmeisser, C.; Krishnan, H.B.; Streit, W.R. RNA sequencing analysis of the broad-host-range strain Sinorhizobium fredii NGR234 identifies a large set of genes linked to quorum sensing-dependent regulation in the background of a traI and ngrI deletion mutant. Appl. Environ. Microbiol. 2014, 80, 5655–5671. [Google Scholar] [CrossRef] [Green Version]
- Staehelin, C.; Forsberg, L.S.; D’Haeze, W.; Gao, M.Y.; Carlson, R.W.; Xie, Z.P.; Pellock, B.J.; Jones, K.M.; Walker, G.C.; Streit, W.R.; et al. Exo-oligosaccharides of Rhizobium sp. strain NGR234 are required for symbiosis with various legumes. J. Bacteriol. 2006, 188, 6168–6178. [Google Scholar] [CrossRef] [Green Version]
- Parada, M.; Vinardell, J.M.; Ollero, F.J.; Hidalgo, A.; Guitierrez, R.; Buendia-Claveria, A.M.; Lei, W.; Margaret, I.; Lopez-Baena, F.J.; Gil-Serrano, A.M.; et al. Sinorhizobium fredii HH103 mutants affected in capsular polysaccharide (KPS) are impaired for nodulation with soybean and Cajanus cajan. Mol. Plant Microbe Interact. 2006, 19, 43–52. [Google Scholar] [CrossRef] [Green Version]
- Hidalgo, A.; Margaret, I.; Crespo-Rivas, J.C.; Parada, M.; Murdoch, P.S.; Lopez, A.; Buendia-Claveria, A.M.; Moreno, J.; Albareda, M.; Gil- Serrano, A.M.; et al. The rkpU gene of Sinorhizobium fredii HH103 is required for bacterial K- antigen polysaccharide production and for efficient nodulation with soybean but not with cowpea. Microbiology 2010, 156, 3398–3411. [Google Scholar] [CrossRef]
- Acosta-Jurado, S.; Rodríguez-Navarro, D.N.; Kawaharada, Y.; Perea, J.F.; Gil-Serrano, A.; Jin, H.; An, Q.; Rodríguez-Carvajal, M.A.; Andersen, S.U.; Sandal, N.; et al. Sinorhizobium fredii HH103 invades Lotus burttii by crack entry in a Nod factor- and surface polysaccharide-dependent manner. Mol. Plant Microbe Interact. 2016, 29, 925–937. [Google Scholar] [CrossRef] [Green Version]
- Margaret-Oliver, I.; Lei, W.; Parada, M.; Rodríguez-Carvajal, M.A.; Crespo-Rivas, J.C.; Hidalgo, Á.; Gil-Serrano, A.; Moreno, J.; Rodríguez-Navarro, D.N.; Buendía-Clavería, A.; et al. Sinorhizobium fredii HH103 does not strictly require KPS and/or EPS to nodulate Glycyrrhiza uralensis, an indeterminate nodule-forming legume. Arch. Microbiol. 2012, 194, 87–102. [Google Scholar] [CrossRef]
- Crespo-Rivas, J.C.; Guefrachi, I.; Mok, K.C.; Villaécija-Aguilar, J.A.; Acosta-Jurado, S.; Pierre, O.; Ruiz-Sainz, J.E.; Taga, M.E.; Mergaert, P.; Vinardell, J.M. Sinorhizobium fredii HH103 bacteroids are not terminally differentiated and show altered O-antigen in nodules of the Inverted Repeat-Lacking Clade legume Glycyrrhiza uralensis. Environ. Microbiol. 2016, 18, 2392–2404. [Google Scholar] [CrossRef]
- Martínez-Hidalgo, P.; Ramírez-Bahena, M.H.; Flores-Félix, J.D.; Igual, J.M.; Sanjuán, J.; León-Barrios, M.; Peix, A.; Velázquez, E. Reclassification of strains MAFF 303099T and R7A into Mesorhizobium japonicum sp. nov. Int. J. Syst. Evol. Microbiol. 2016, 66, 4936–4941. [Google Scholar] [CrossRef]
- Sullivan, J.T.; Patrick, H.N.; Lowther, W.L.; Scott, D.B.; Ronson, C.W. Nodulating strain of Rhizobium loti arise through chromosomal symbiotic gene transfer in the environment. Proc. Natl. Acad. Sci. USA 1995, 92, 8985–8989. [Google Scholar] [CrossRef] [Green Version]
- Saeki, K.; Kouchi, H. The Lotus symbiont, Mesorhizobium loti: Molecular genetic techniques and application. J. Plant Res. 2000, 113, 457–465. [Google Scholar] [CrossRef]
- López-Lara, I.M.; van den Berg, J.D.J.; Thomas-Oates, J.E.; Glushka, J.; Lugtenberg, B.J.J.; Spaink, H.P. Structural identification of the lipo-chitin oligosaccharide nodulation signals of Rhizobium loti. Mol. Microbiol. 1995, 15, 627–638. [Google Scholar] [CrossRef]
- Imaizumi-Anraku, H.; Kawaguchi, M.; Koiwa, H.; Akao, S.; Syōno, K. Two ineffective-nodulating mutants of Lotus japonicus-different phenotypes caused by the blockage of endocytotic bacterial release and nodule maturation. Plant Cell Physiol. 1997, 38, 871–881. [Google Scholar] [CrossRef] [Green Version]
- Kaneko, T.; Nakamura, Y.; Sato, S.; Asamizu, E.; Kato, T.; Sasamoto, S.; Watanabe, A.; Idesawa, K.; Ishikawa, A.; Kawashima, K.; et al. Complete genome structure of the nitrogen-fixing symbiotic bacterium Mesorhizobium loti. DNA Res. 2000, 7, 331–338. [Google Scholar] [CrossRef] [Green Version]
- Kelly, S.; Sullivan, J.; Ronson, C.; Tian, R.; Bräu, L.; Munk, C.; Goodwin, L.; Han, C.; Woyke, T.; Reddy, T.; et al. Genome sequence of the Lotus spp. microsymbiont Mesorhizobium loti strain R7A. Stand. Genom. Sci. 2014, 9, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, J.T.; Trzebiatowski, J.R.; Cruickshank, R.W.; Gouzy, J.; Brown, S.D.; Elliot, R.M.; Fleetwood, D.J.; McCallum, N.G.; Rossbach, U.; Stuart, G.S.; et al. Comparative sequence analysis of the symbiosis island of Mesorhizobium loti strain R7A. J. Bacteriol. 2002, 184, 3086–3095. [Google Scholar] [CrossRef] [Green Version]
- Hubber, A.; Vergunst, A.C.; Sullivan, J.T.; Hooykaas, P.J.; Ronson, C.W. Symbiotic phenotypes and translocated effector proteins of the Mesorhizobium loti strain R7A VirB/D4 type IV secretion system. Mol. Microbiol. 2004, 54, 561–574. [Google Scholar] [CrossRef]
- Hotter, G.S.; Scott, D.B. Exopolysaccharide mutants of Rhizobium loti are fully effective on a determinate nodulating host but are ineffective on an indeterminate nodulating host. J. Bacteriol. 1991, 173, 851–859. [Google Scholar] [CrossRef] [Green Version]
- Mishima, E.; Hosokawa, A.; Imaizumi-Anraku, H.; Saito, K.; Kawaguchi, M.; Saeki, K. Requirement for Mesorhizobium loti ornithine transcarbamoylase for successful symbiosis with Lotus japonicus as revealed by an unexpected long-range genome deletion. Plant Cell Physiol. 2008, 49, 301–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radutoiu, S.; Madsen, L.H.; Madsen, E.B.; Felle, H.H.; Umehara, Y.; Gronlund, M.; Sato, S.; Nakamura, Y.; Tabata, S.; Sandal, N.; et al. Plant recognition of symbiotic bacteria requires two LysM receptor-like kinases. Nature 2003, 425, 585–592. [Google Scholar] [CrossRef] [PubMed]
- Broghammer, A.; Krusell, L.; Blaise, M.; Sauer, J.; Sullivan, J.T.; Maolanon, N.; Vinther, M.; Lorentzen, A.; Madsen, E.B.; Jensen, K.J.; et al. Legume receptors perceive the rhizobial lipochitin oligosaccharide signal molecules by direct binding. Proc. Natl. Acad. Sci. USA 2012, 109, 13859–13864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, J.E.M.M.; Gysel, K.; Birkefeldt, T.G.; Vinther, M.; Muszyński, A.; Azadi, P.; Laursen, N.S.; Sullivan, J.T.; Ronson, C.W.; Stougaard, J.; et al. Structural signatures in EPR3 define a unique class of plant carbohydrate receptors. Nature 2020, 11, 3797. [Google Scholar] [CrossRef]
- Maillet, F.; Fournier, J.; Mendis, H.C.; Tadege, M.; Wen, J.; Ratet, P.; Mysore, K.S.; Gough, C.; Jones, K.M. Sinorhizobium meliloti succinylated high-molecular-weight succinoglycan and the Medicago truncatula LysM receptor-like kinase MtLYK10 participate independently in symbiotic infection. Plant J. 2020, 102, 311–326. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Acosta-Jurado, S.; Fuentes-Romero, F.; Ruiz-Sainz, J.-E.; Janczarek, M.; Vinardell, J.-M. Rhizobial Exopolysaccharides: Genetic Regulation of Their Synthesis and Relevance in Symbiosis with Legumes. Int. J. Mol. Sci. 2021, 22, 6233. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22126233
Acosta-Jurado S, Fuentes-Romero F, Ruiz-Sainz J-E, Janczarek M, Vinardell J-M. Rhizobial Exopolysaccharides: Genetic Regulation of Their Synthesis and Relevance in Symbiosis with Legumes. International Journal of Molecular Sciences. 2021; 22(12):6233. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22126233
Chicago/Turabian StyleAcosta-Jurado, Sebastián, Francisco Fuentes-Romero, Jose-Enrique Ruiz-Sainz, Monika Janczarek, and José-María Vinardell. 2021. "Rhizobial Exopolysaccharides: Genetic Regulation of Their Synthesis and Relevance in Symbiosis with Legumes" International Journal of Molecular Sciences 22, no. 12: 6233. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22126233