
In Vitro Characterization of Guanylyl Cyclase BdPepR2 from Brachypodium distachyon Identified through a Motif-Based Approach

 , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
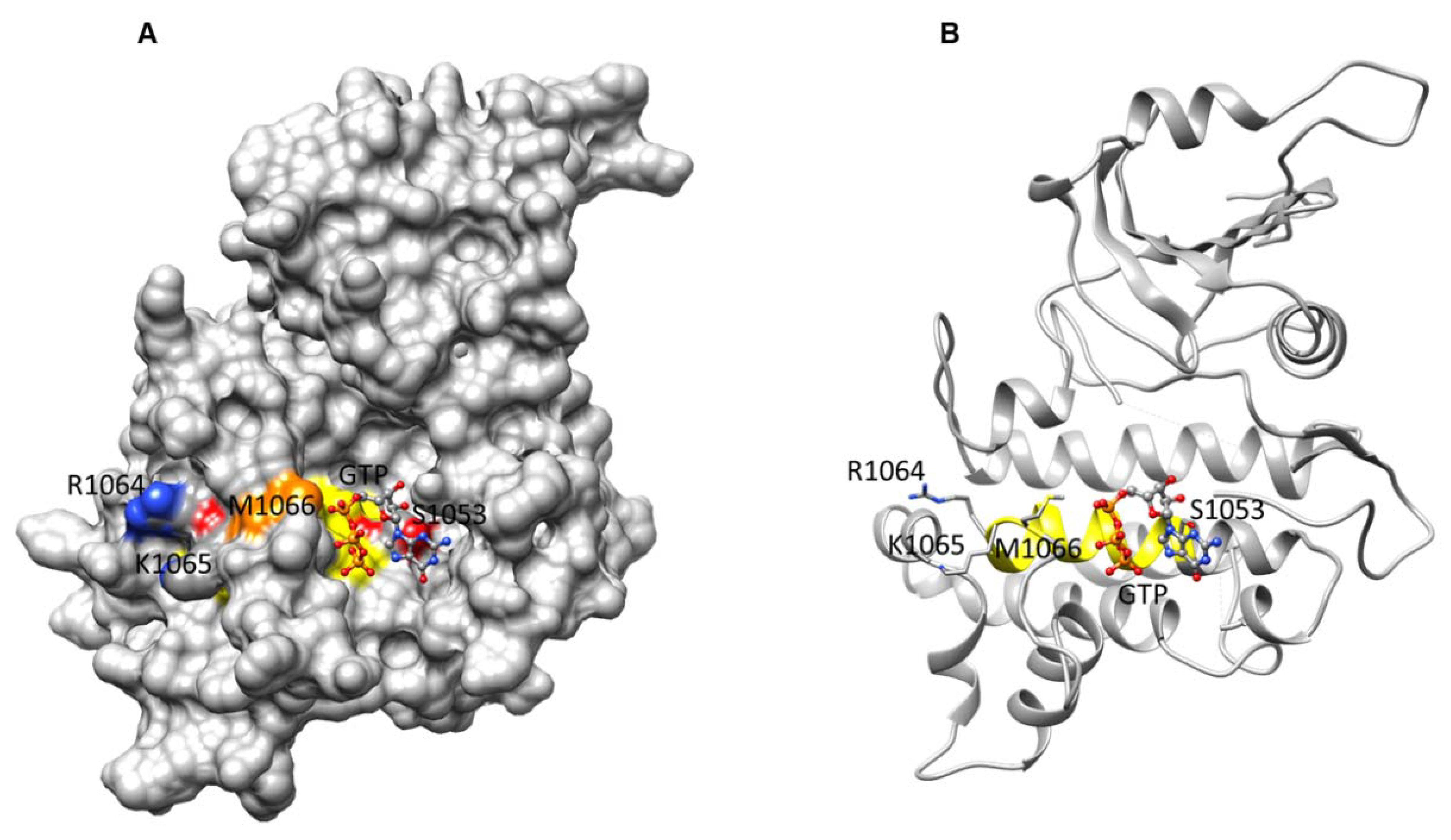
2.1. Sequence Analysis of BdPepR2
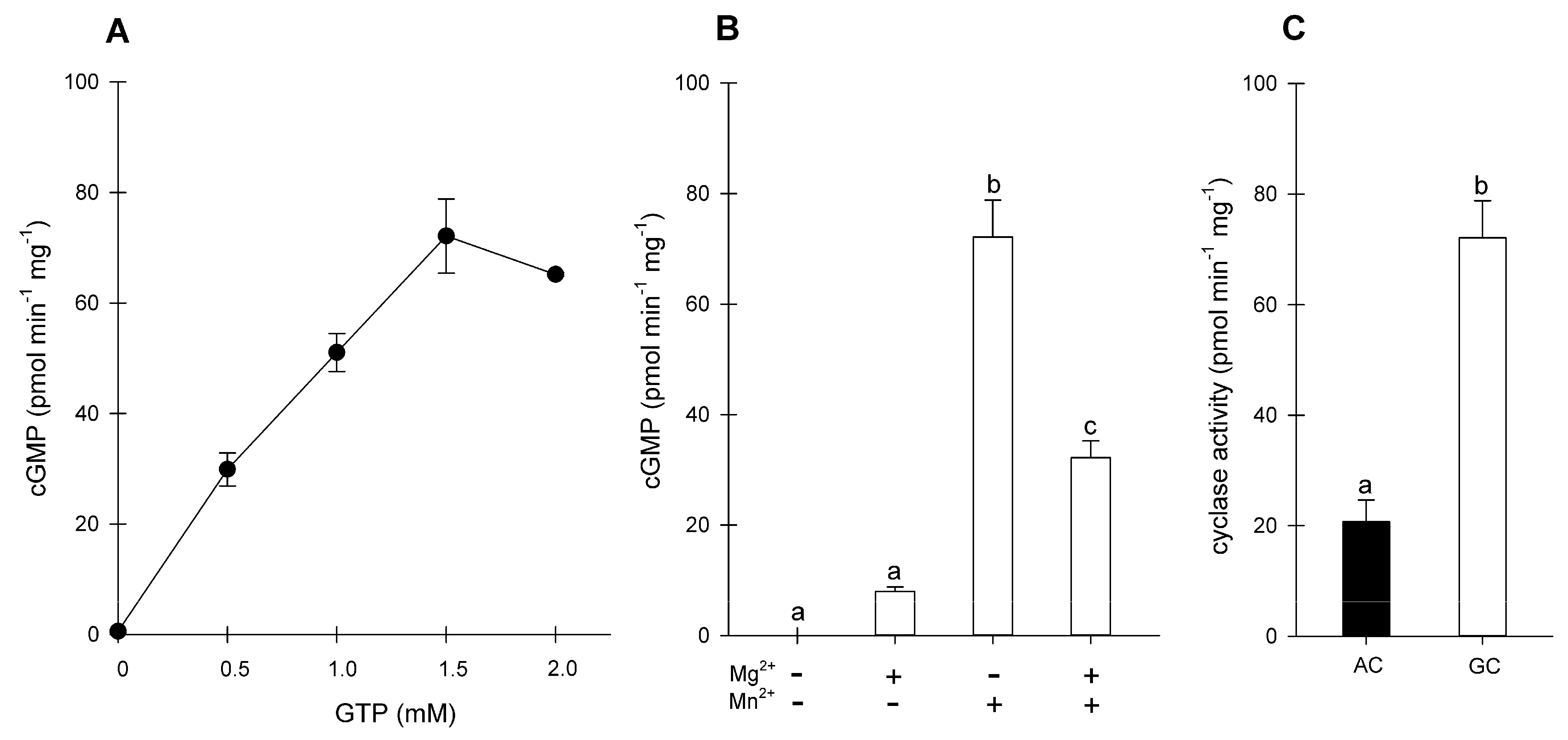
2.2. Purification of BdPepR2 Recombinant Protein and In Vitro GC Activity
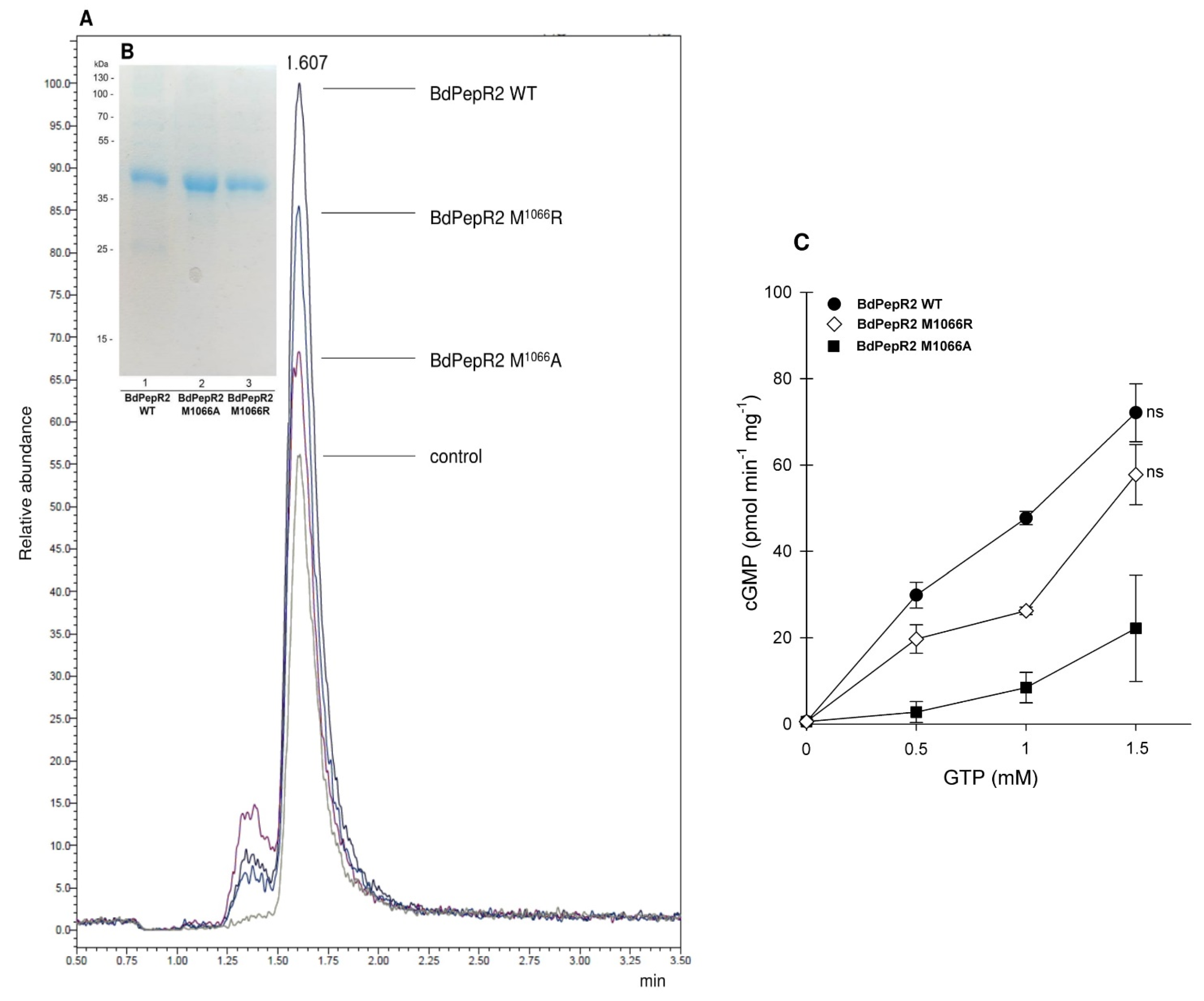
2.3. Mutational Analysis
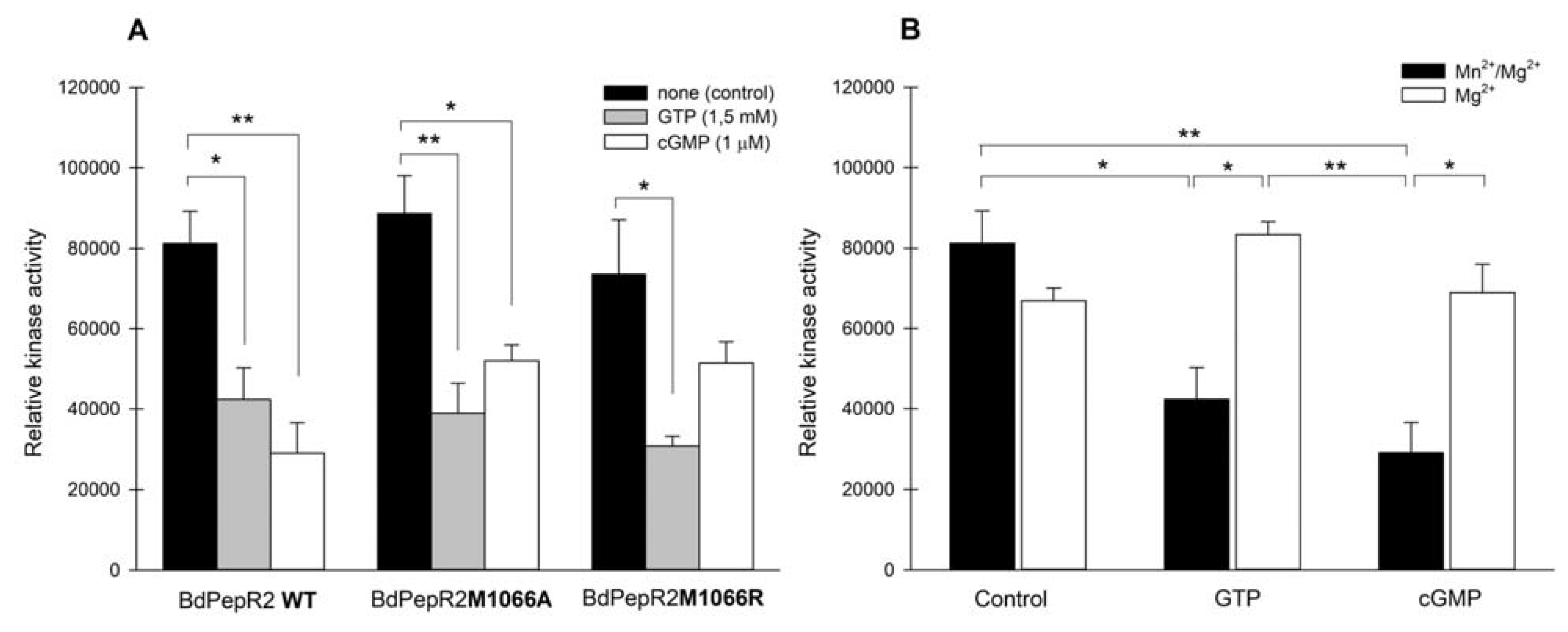
2.4. BdPepR2 Kinase Activity
3. Materials and Methods
3.1. Construction of Expression Vectors
3.2. Expression and Purification of the Recombinant Proteins
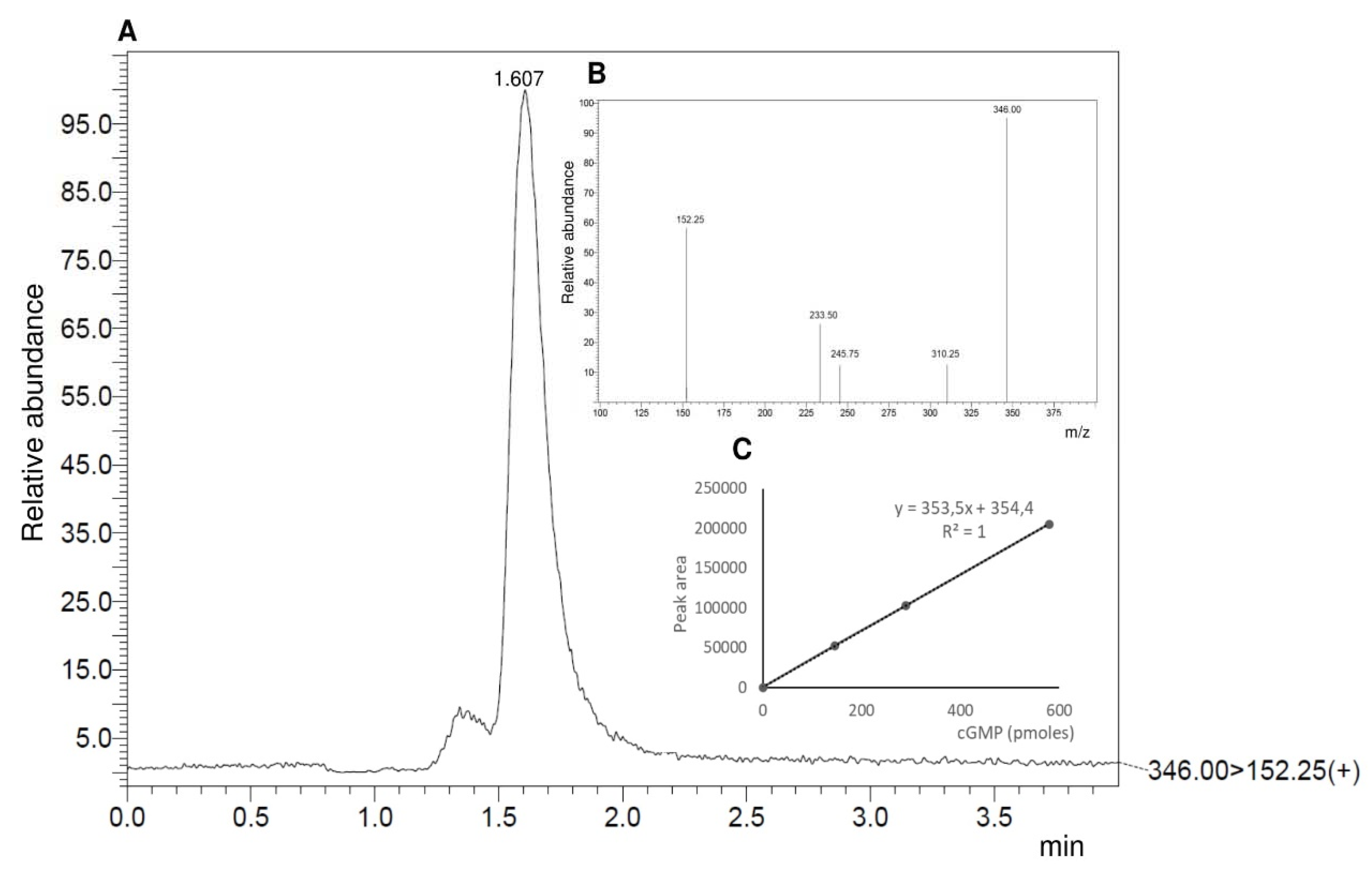
3.3. Determination of Guanylyl Cyclase Activity
3.4. Determination of Proteins Kinase Activity
3.5. Computational Modeling
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ashton, A.R. Guanylyl cyclase activity in plants? Proc. Natl. Acad. Sci. USA 2011, 108, E97–E98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bojar, D.; Martinez, J.; Santiago, J.; Rybin, V.; Bayliss, R.; Hothorn, M. Crystal structures of the phosphorylated BRI1 kinase domain and implications for brassinosteroid signal initiation. Plant J. 2014, 78, 31–43. [Google Scholar] [CrossRef] [PubMed]
- Gehring, C.; Turek, I.S. Cyclic nucleotide monophosphates and their cyclases in plant signaling. Front. Plant. Sci. 2017, 8, 1704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turek, I.; Irving, H. Moonlighting Proteins Shine New Light on Molecular Signaling Niches. Int. J. Mol. Sci. 2021, 22, 1367. [Google Scholar] [CrossRef]
- Hu, X.; Neill, S.J.; Tang, Z.; Cai, W. Nitric oxide mediates gravitropic bending in soybean roots. Plant Physiol. 2005, 137, 663–670. [Google Scholar] [CrossRef] [Green Version]
- Szmidt-Jaworska, A.; Jaworski, K.; Kopcewicz, J. Involvement of cyclic GMP in phytochrome-controlled flowering of Pharbitis nil. J. Plant Physiol. 2008, 165, 858–867. [Google Scholar] [CrossRef]
- Teng, Y.; Xu, W.; Ma, M. cGMP is required for seed germination in Arabidopsis thaliana. J. Plant Physiol. 2010, 167, 885–889. [Google Scholar] [CrossRef]
- Kwezi, L.; Meier, S.; Mungur, L.; Ruzvidzo, O.; Irving, H.; Gehring, C. The Arabidopsis thaliana Brassinosteroid Receptor (AtBRI1) Contains a Domain that Functions as a Guanylyl Cyclase In Vitro. PLoS ONE 2007, 2, e449. [Google Scholar] [CrossRef] [Green Version]
- Kwezi, L.; Ruzvidzo, O.; Wheeler, J.I.; Govender, K.; Iacuone, S.; Thompson, P.E.; Gehring, C.; Irving, H.R. The phytosulfokine (PSK) receptor is capable of guanylate cyclase activity and enabling cyclic GMP-dependent signaling in plants. J. Biol. Chem. 2011, 286, 22580–22588. [Google Scholar] [CrossRef] [Green Version]
- Muleya, V.; Wheeler, J.I.; Ruzvidzo, O.; Freihat, L.; Manallack, D.T.; Gehring, C.; Irving, H.R. Calcium is the switch in the moonlighting dual function of the ligand-activated receptor kinase phytosulfokine receptor 1. Cell Commun. Signal. 2014, 12, 60. [Google Scholar] [CrossRef]
- Wong, A.; Donaldson, L.; Portes, M.T.; Eppinger, J.; Feijó, J.A.; Gehring, C. Arabidopsis DIACYLGLYCEROL KINASE4 is involved in nitric oxide-dependent pollen tube guidance and fertilization. Development 2020, 147, dev183715. [Google Scholar] [CrossRef]
- Qi, Z.; Verma, R.; Gehring, C.; Yamaguchi, Y.; Zhao, Y.; Ryan, C.A.; Berkowitz, G.A. Ca2+ signaling by plant Arabidopsis thaliana Pep peptides depends on AtPepR1, a receptor with guanylyl cyclase activity, and cGMP-activated Ca2+ channels. Proc. Natl. Acad. Sci. USA 2010, 107, 21193–21198. [Google Scholar] [CrossRef] [Green Version]
- Świeżawska, B.; Jaworski, K.; Duszyn, M.; Pawełek, A.; Szmidt-Jaworska, A. The Hippeastrum hybridum PepR1 gene (HpPepR1) encodes a functional guanylyl cyclase and is involved in early response to fungal infection. J. Plant Physiol. 2017, 216, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Malukani, K.K.; Ranjan, A.; Hota, S.J.; Patel, H.K.; Sonti, R.V. Dual Activities of Receptor-Like Kinase OsWAKL21.2 Induce Immune Responses. Plant Physiol. 2020, 183, 1345–1363. [Google Scholar] [CrossRef] [PubMed]
- Rahman, H.; Wang, X.Y.; Xu, Y.P.; He, Y.H.; Cai, X.Z. Characterization of tomato protein kinases embedding guanylate cyclase catalytic center motif. Sci. Rep. 2020, 10, 4078. [Google Scholar] [CrossRef] [PubMed]
- Ludidi, N.; Gehring, C. Identification of a novel protein with guanylyl cyclase activity in Arabidopsis thaliana. J. Biol. Chem. 2003, 278, 6490–6494. [Google Scholar] [CrossRef] [Green Version]
- Wong, A.; Gehring, C.; Irving, H.R. Conserved functional motifs and homology modeling to predict hidden moonlighting functional sites. Front. Bioeng. Biotechnol. 2015, 3, 82. [Google Scholar] [CrossRef] [Green Version]
- Wong, A.; Tian, X.; Gehring, C.; Marondedze, C. Discovery of Novel Functional Centers with Rationally Designed Amino Acid Motifs. Comput. Struct. Biotechnol. J. 2018, 16, 70–76. [Google Scholar] [CrossRef]
- Chakraborty, S.; Nguyen, B.; Wasti, S.D.; Xu, G. Plant Leucine-Rich Repeat Receptor Kinase (LRR-RK): Structure, Ligand Perception, and Activation Mechanism. Molecules 2019, 24, 3081. [Google Scholar] [CrossRef] [Green Version]
- Meier, S.; Ruzvidzo, O.; Morse, M.; Donaldson, L.; Kwezi, L.; Gehring, C. The Arabidopsis Wall Associated Kinase-Like 10 Gene Encodes a Functional Guanylyl Cyclase and Is Co-Expressed with Pathogen Defense Related Genes. PLoS ONE 2010, 5, e8904. [Google Scholar] [CrossRef] [Green Version]
- Mulaudzi, T.; Ludidi, N.; Ruzvidzo, O.; Morse, M.; Hendricks, N.; Iwuoha, E.; Gehring, C. Identification of a novel Arabidopsis thaliana nitric oxide-binding molecule with guanylate cyclase activity in vitro. FEBS Lett. 2011, 585, 2693–2697. [Google Scholar] [CrossRef] [Green Version]
- Turek, I.; Gehring, C. The plant natriuretic peptide receptor is a guanylyl cyclase and enables cGMP-dependent signaling. Plant Mol. Biol. 2016, 91, 275–286. [Google Scholar] [CrossRef]
- Szmidt-Jaworska, A.; Jaworski, K.; Pawełek, A.; Kopcewicz, J. Molecular cloning and characterization of a guanylyl cyclase, PnGC-1: Involved in light signaling in Pharbitis nil. J. Plant Growth Regul. 2009, 28, 367–380. [Google Scholar] [CrossRef]
- Świeżawska, B.; Jaworski, K.; Szewczuk, P.; Pawełek, A.; Szmidt-Jaworska, A. Identification of a Hippeastrum hybridum guanylyl cyclase responsive to wounding and pathogen infection. J. Plant Physiol. 2015, 189, 77–86. [Google Scholar] [CrossRef]
- Rogers, N.M.; Seeger, F.; Garcin, E.D.; Roberts, D.D.; Isenberg, J.S. Regulation of soluble guanylate cyclase by matricellular thrombospondins: Implications for blood flow. Front. Physiol. 2014, 5, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wheeler, J.I.; Wong, A.; Marondedze, C.; Groen, A.J.; Kwezi, L.; Freihat, L.; Vyas, J.; Raji, M.A.; Irving, H.R.; Gehring, C. The brassinosteroid receptor BRI1 can generate cGMP enabling cGMP-dependent downstream signaling. Plant J. 2017, 91, 590–600. [Google Scholar] [CrossRef] [Green Version]
- Wong, A.; Gehring, C. The Arabidopsis thaliana proteome harbors undiscovered multi-domain molecules with functional guanylyl cyclase catalytic centers. Cell Commun. Signal. 2013, 11, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, N.; Fu, D.; Li, S.; Wang, Y.; Wong, A. GCPred: A web tool for guanylyl cyclase functional centre prediction from amino acid sequence. Bioinformatics 2018, 34, 2134–2135. [Google Scholar] [CrossRef]
- Al-Younis, I.; Wong, A.; Gehring, C. The Arabidopsis thaliana K+-uptake permease 7 (AtKUP7) contains a functional cytosolic adenylate cyclase catalytic centre. FEBS Lett. 2015, 589, 3848–3852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Younis, I.; Wong, A.; Lemtiri-Chlieh, F.; Schmöckel, S.; Tester, M.; Gehring, C.; Donaldson, L. The Arabidopsis thaliana K+—Uptake permease 5 (AtKUP5) contains a functional cytosolic adenylate cyclase essential for K+ transport. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Šali, A.; Blundell, T.L. Comparative protein modelling by satisfaction of spatial restraints. J. Mol. Biol. 1993, 234, 779–815. [Google Scholar] [CrossRef] [PubMed]
- Troot, O.; Olson, A.J. Software news and update AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duszyn, M.; Świeżawska-Boniecka, B.; Wong, A.; Jaworski, K.; Szmidt-Jaworska, A. In Vitro Characterization of Guanylyl Cyclase BdPepR2 from Brachypodium distachyon Identified through a Motif-Based Approach. Int. J. Mol. Sci. 2021, 22, 6243. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22126243
Duszyn M, Świeżawska-Boniecka B, Wong A, Jaworski K, Szmidt-Jaworska A. In Vitro Characterization of Guanylyl Cyclase BdPepR2 from Brachypodium distachyon Identified through a Motif-Based Approach. International Journal of Molecular Sciences. 2021; 22(12):6243. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22126243
Chicago/Turabian StyleDuszyn, Maria, Brygida Świeżawska-Boniecka, Aloysius Wong, Krzysztof Jaworski, and Adriana Szmidt-Jaworska. 2021. "In Vitro Characterization of Guanylyl Cyclase BdPepR2 from Brachypodium distachyon Identified through a Motif-Based Approach" International Journal of Molecular Sciences 22, no. 12: 6243. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22126243