
Arabidopsis thaliana Myb59 Gene Is Involved in the Response to Heterodera schachtii Infestation, and Its Overexpression Disturbs Regular Development of Nematode-Induced Syncytia

 ,
,
Abstract
:1. Introduction
2. Results
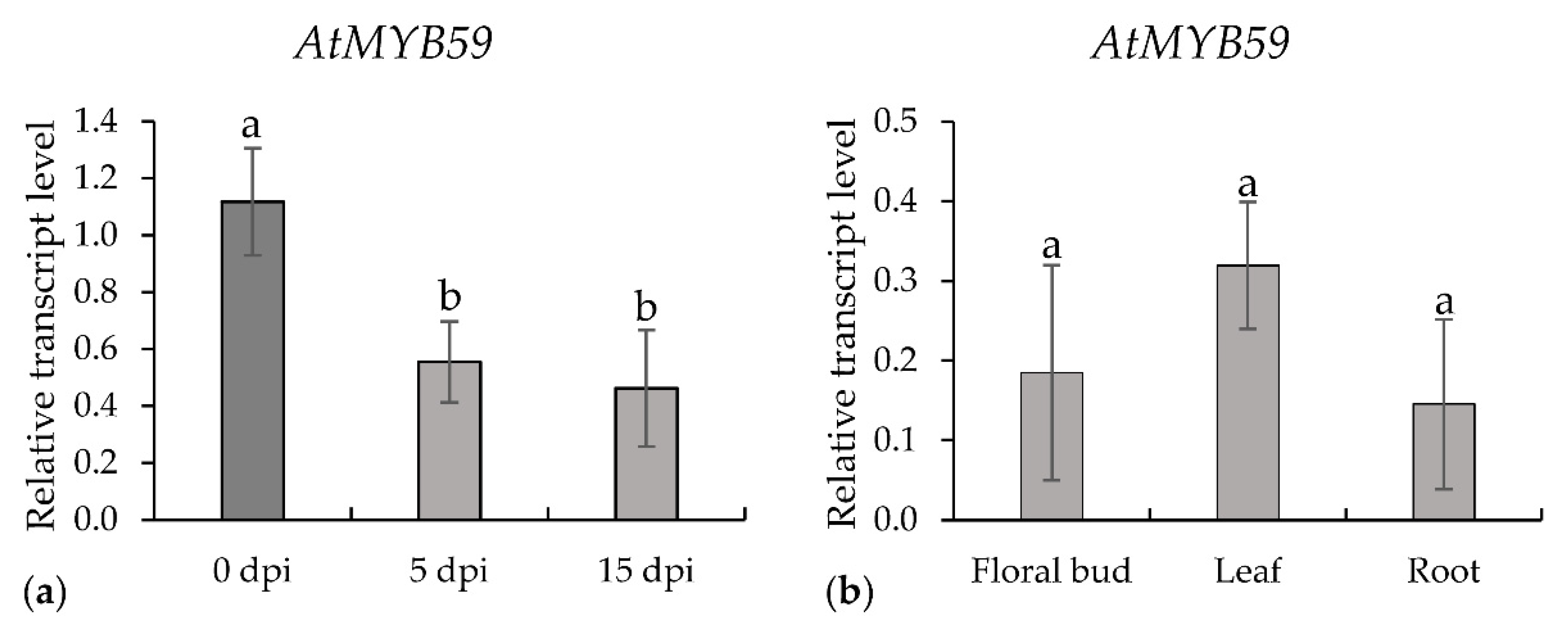
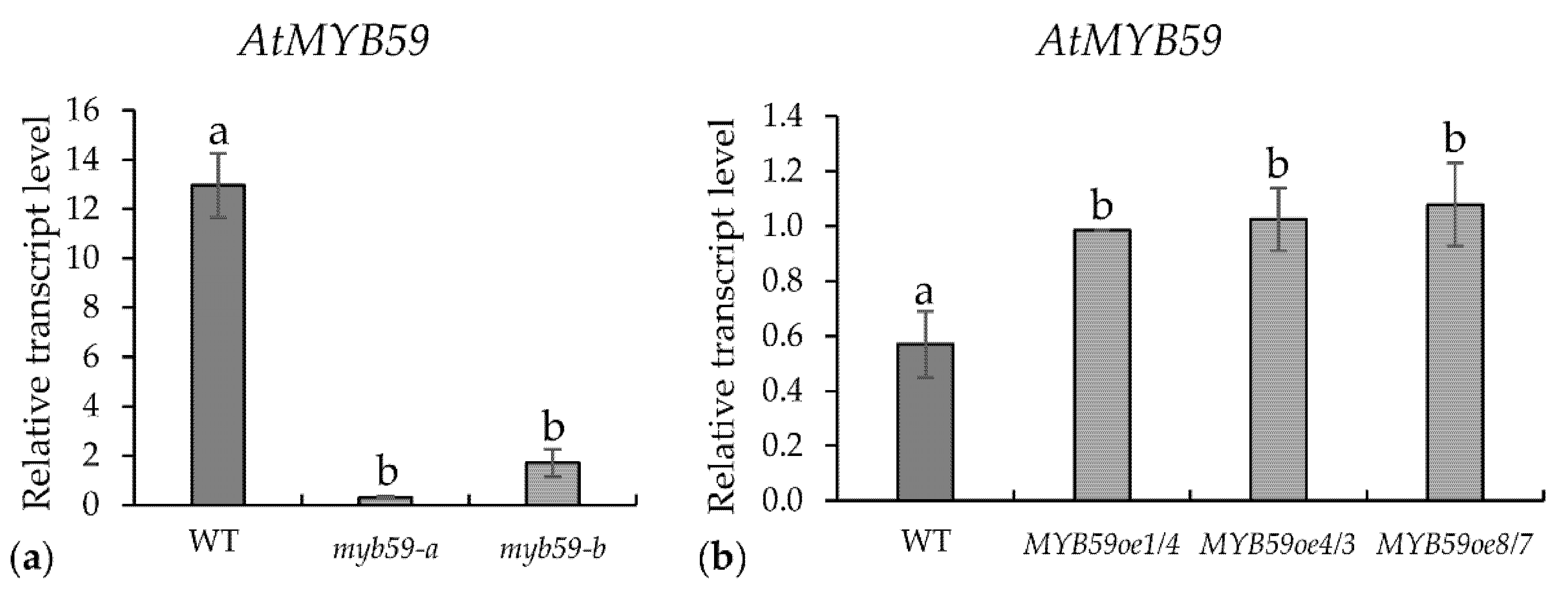
2.1. Expression of AtMYB59 Gene
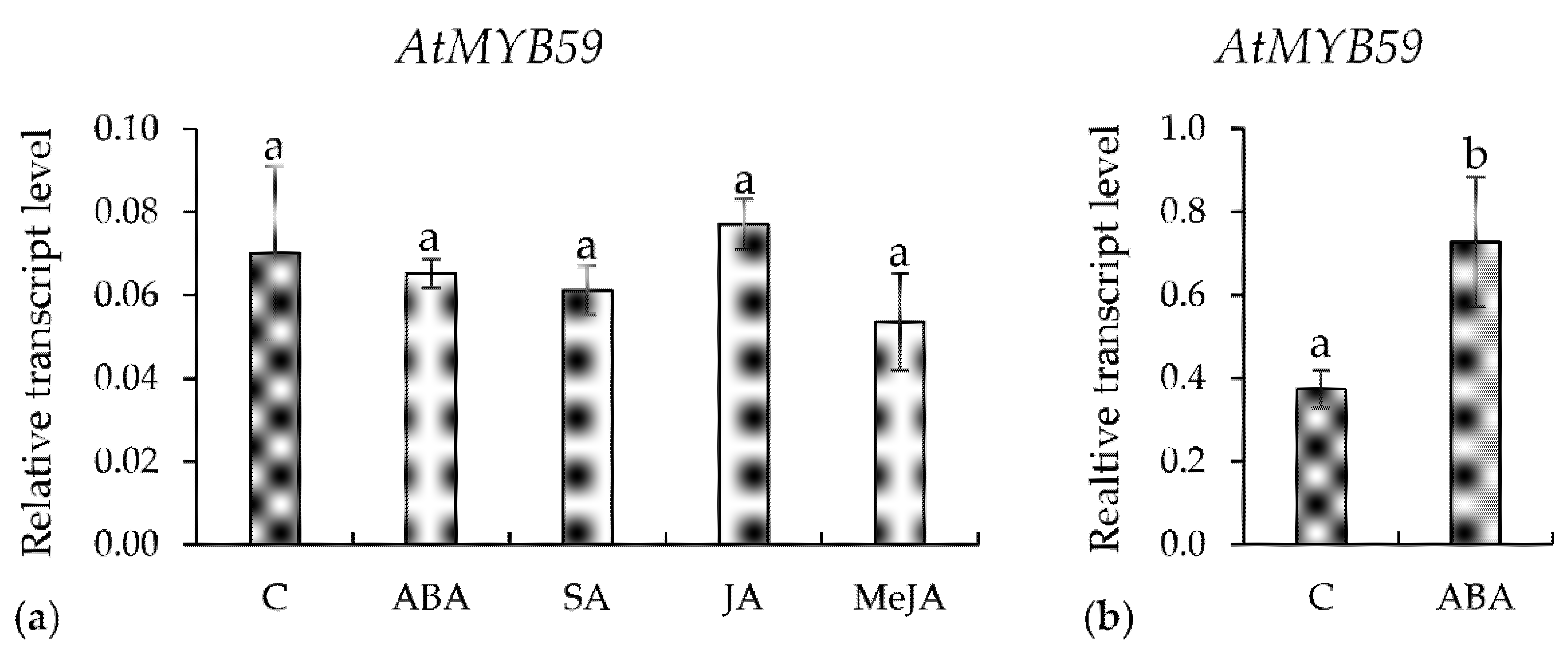
2.2. Expression of AtMYB59 Gene in Roots or Leaves Treated with Phytohormones
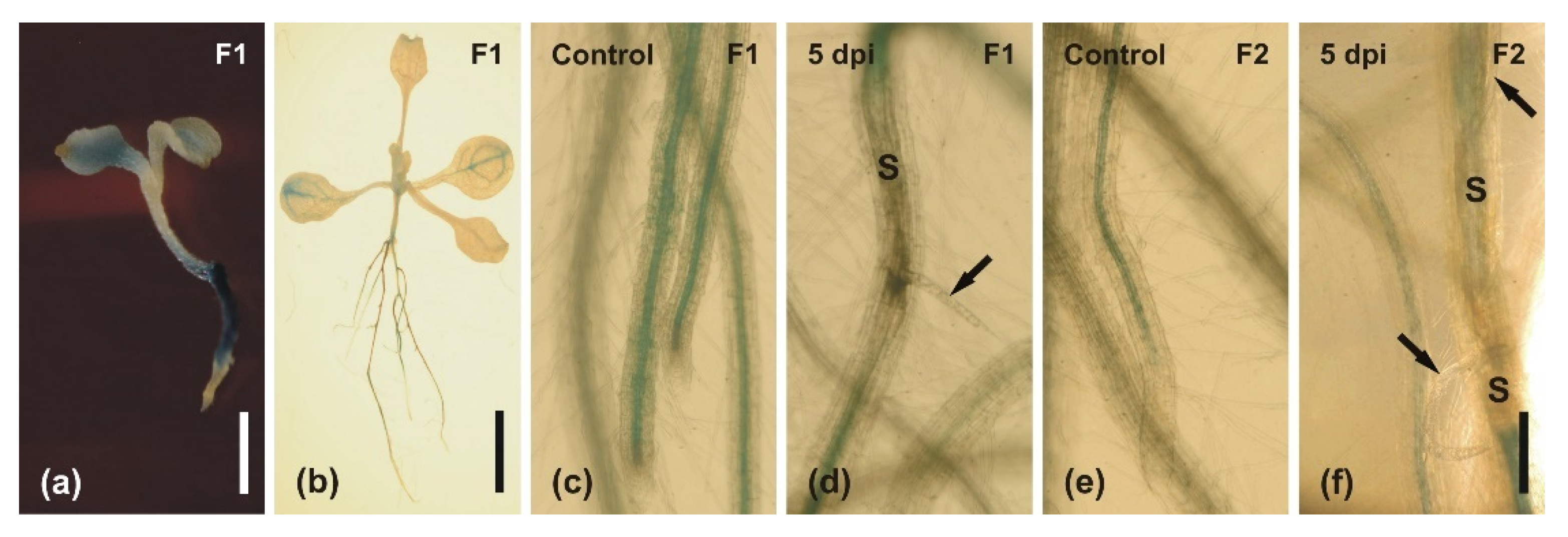
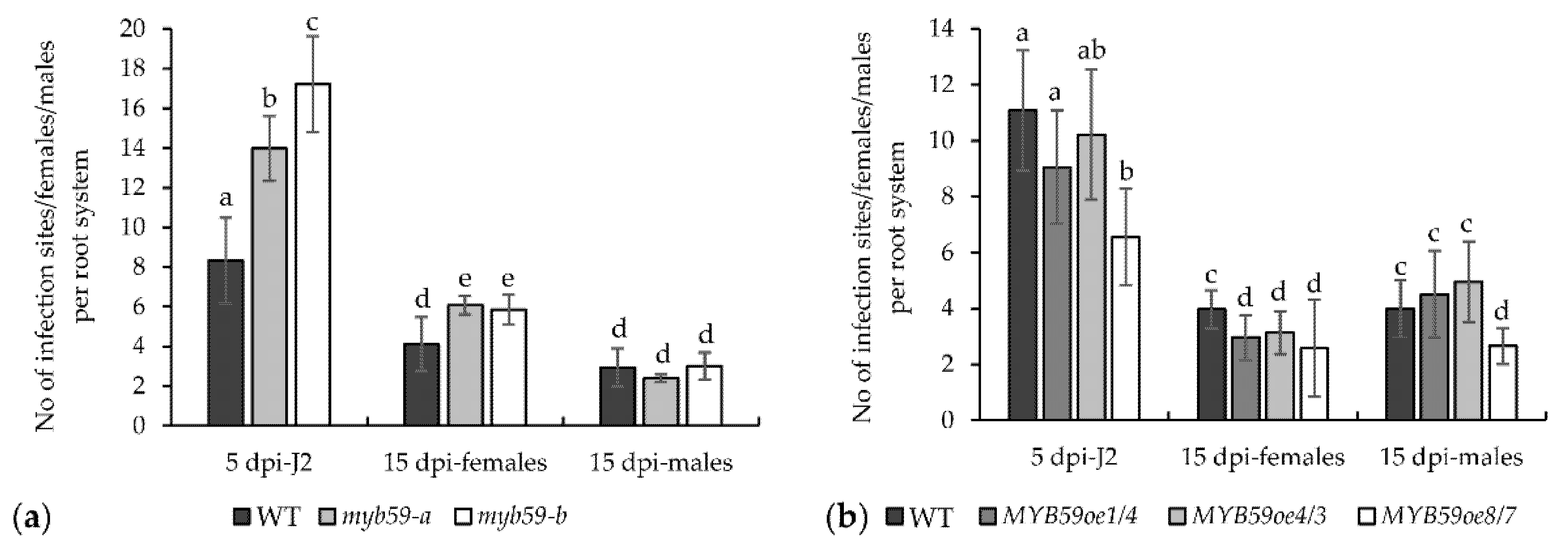
2.3. Infection of Roots with H. schachtii
2.4. Cellular and Ultrastructural Differences of Syncytia
2.5. cis Regulatory Elements Analysis of AtMYB59 Gene Promoter
3. Discussion
4. Materials and Methods
4.1. Plant Material and Culture Conditions
4.2. Gene Construct Preparation
4.3. A. thaliana Transformation
4.4. Mutants Genotyping
4.5. RNA Extraction and RT-PCR
4.6. Promoter cis Element Analysis
4.7. GUS Activity Assay
4.8. Nematode Infection Assay
4.9. Anatomic and Ultrastructural Analysis
4.10. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jones, J.T.; Haegeman, A.; Danchin, E.G.J.; Gaur, H.S.; Helder, J.; Jones, M.G.K.; Kikuchi, T.; Manzanilla-López, R.; Palomares-Rius, J.E.; Wesemael, W.M.L.; et al. Top 10 plant-parasitic nematodes in molecular plant pathology. Mol. Plant Pathol. 2013, 14, 946–961. [Google Scholar] [CrossRef] [PubMed]
- Wyss, U. Observations on the feeding behaviour of Heterodera schachtii throughout development, including events during moulting. Fundam. Appl. Nematol. 1992, 15, 75–89. [Google Scholar]
- Golinowski, W.; Grundler, F.M.W.; Sobczak, M. Changes in the structure of Arabidopsis thaliana during female development of the plant-parasitic nematode Heterodera schachtii. Protoplasma 1996, 194, 103–116. [Google Scholar] [CrossRef]
- Siddique, S.; Radakovic, Z.S.; Carola, M.; Chronis, D.; Novák, O.; Ramireddy, E.; Holbein, J.; Matera, C.; Hütten, M.; Gutbrod, P.; et al. A parasitic nematode releases cytokinin that controls cell division and orchestrates feeding site formation in host plants. Proc. Natl. Acad. Sci. USA 2015, 112, 12669–12674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feller, A.; MacHemer, K.; Braun, E.L.; Grotewold, E. Evolutionary and comparative analysis of MYB and bHLH plant transcription factors. Plant J. 2011, 66, 94–116. [Google Scholar] [CrossRef]
- Kranz, H.D.; Denekamp, M.; Greco, R.; Jin, H.; Leyva, A.; Meissner, R.C.; Petroni, K.; Urzainqui, A.; Bevan, M.; Martin, C.; et al. Towards functional characterisation of the members of the R2R3-MYB gene family from Arabidopsis thaliana. Plant J. 1998, 16, 263–276. [Google Scholar] [CrossRef]
- Cheng, C.-Y.; Krishnakumar, V.; Chan, A.P.; Thibaud-Nissen, F.; Schobel, S.; Town, C.D. Araport11: A complete reannotation of the Arabidopsis thaliana reference genome. Plant J. 2017, 89, 789–804. [Google Scholar] [CrossRef] [Green Version]
- Prouse, M.B.; Campbell, M.M. The interaction between MYB proteins and their target DNA binding sites. Biochim. Biophys. Acta Gene Regul. Mech. 2012, 1819, 67–77. [Google Scholar] [CrossRef]
- Glazebrook, J. Contrasting mechanisms of defense against biotrophic and necrotrophic pathogens. Annu. Rev. Phytopathol. 2005, 43, 205–227. [Google Scholar] [CrossRef]
- Spoel, S.H.; Dong, X. Making sense of hormone crosstalk during plant immune responses. Cell Host Microbe 2008, 3, 348–351. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Q.; Putker, V.; Goverse, A. Molecular and cellular mechanisms involved in host-specific resistance to cyst nematodes in crops. Front. Plant Sci. 2021, 12, 641582. [Google Scholar] [CrossRef]
- Szakasits, D.; Heinen, P.; Wieczorek, K.; Hofmann, J.; Wagner, F.; Kreil, D.P.; Sykacek, P.; Grundler, F.M.W.; Bohlmann, H. The transcriptome of syncytia induced by the cyst nematode Heterodera schachtii in Arabidopsis roots. Plant J. 2009, 57, 771–784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barcala, M.; García, A.; Cabrera, J.; Casson, S.; Lindsey, K.; Favery, B.; García-Casado, G.; Solano, R.; Fenoll, C.; Escobar, C. Early transcriptomic events in microdissected Arabidopsis nematode-induced giant cells. Plant J. 2010, 61, 698–712. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, X.; Guo, L.; Lu, F.; Feng, X.; He, K.; Wei, L.; Chen, Z.; Qu, L.-J.; Gu, H. A subgroup of MYB transcription factor genes undergoes highly conserved alternative splicing in Arabidopsis and rice. J. Exp. Bot. 2006, 57, 1263–1273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L. The roles of MYB transcription factors on plant defense responses and its molecular mechanism. Heredity 2008, 30, 1265–1271. [Google Scholar] [CrossRef] [PubMed]
- Mu, R.L.; Cao, Y.R.; Liu, Y.F.; Lei, G.; Zou, H.F.; Liao, Y.; Wang, H.W.; Zhang, W.K.; Ma, B.; Du, J.Z.; et al. An R2R3-type transcription factor gene AtMYB59 regulates root growth and cell cycle progression in Arabidopsis. Cell Res. 2009, 19, 1291–1304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fasani, E.; DalCorso, G.; Costa, A.; Zenoni, S.; Furini, A. The Arabidopsis thaliana transcription factor MYB59 regulates calcium signalling during plant growth and stress response. Plant Mol. Biol. 2019, 99, 517–534. [Google Scholar] [CrossRef]
- Chen, K.; Li, G.; Bressan, R.A.; Song, C.; Zhu, J.; Zhao, Y. Abscisic acid dynamics, signaling, and functions in plants. J. Integr. Plant Biol. 2020, 62, 25–54. [Google Scholar] [CrossRef] [Green Version]
- Kyndt, T.; Nahar, K.; Haeck, A.; Verbeek, R.; Demeestere, K.; Gheysen, G. Interplay between carotenoids, abscisic acid and jasmonate guides the compatible rice-Meloidogyne graminicola interaction. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Moosavi, M.R. The effect of gibberellin and abscisic acid on plant defense responses and on disease severity caused by Meloidogyne javanica on tomato plants. J. Gen. Plant Pathol. 2017, 83, 173–184. [Google Scholar] [CrossRef]
- Du, X.Q.; Wang, F.L.; Li, H.; Jing, S.; Yu, M.; Li, J.; Wu, W.H.; Kudla, J.; Wang, Y. The transcription factor MYB59 regulates K+/NO3¯ translocation in the Arabidopsis response to low K+ stress. Plant Cell 2019, 31, 699–714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riechmann, J.L. Transcriptional regulation: A genomic overview. Arab. B. 2002, 1, e0085. [Google Scholar] [CrossRef] [Green Version]
- Dubos, C.; Stracke, R.; Grotewold, E.; Weisshaar, B.; Martin, C.; Lepiniec, L. MYB transcription factors in Arabidopsis. Trends Plant Sci. 2010, 15, 573–581. [Google Scholar] [CrossRef] [PubMed]
- Rushton, P.J.; Somssich, I.E.; Ringler, P.; Shen, Q.J. WRKY transcription factors. Trends Plant Sci. 2010, 15, 247–258. [Google Scholar] [CrossRef]
- Geri, C.; Cecchini, E.; Giannakou, M.E.; Covey, S.N.; Milner, J.J. Altered patterns of gene expression in Arabidopsis elicited by cauliflower mosaic virus (CaMV) infection and by a CaMV gene VI transgene. Mol. Plant Microbe Interact. 1999, 12, 377–384. [Google Scholar] [CrossRef] [Green Version]
- Seo, P.J.; Park, C.-M. MYB96-mediated abscisic acid signals induce pathogen resistance response by promoting salicylic acid biosynthesis in Arabidopsis. New Phytol. 2010, 186, 471–483. [Google Scholar] [CrossRef] [PubMed]
- Vailleau, F.; Daniel, X.; Tronchet, M.; Montillet, J.-L.; Triantaphylides, C.; Roby, D. A R2R3-MYB gene, AtMYB30, acts as a positive regulator of the hypersensitive cell death program in plants in response to pathogen attack. Proc. Natl. Acad. Sci. USA 2002, 99, 10179–10184. [Google Scholar] [CrossRef] [Green Version]
- Ramírez, V.; Agorio, A.; Coego, A.; García-Andrade, J.; Hernández, M.J.; Balaguer, B.; Ouwerkerk, P.B.F.; Zarra, I.; Vera, P. MYB46 modulates disease susceptibility to Botrytis cinerea in Arabidopsis. Plant Physiol. 2011, 155, 1920–1935. [Google Scholar] [CrossRef] [Green Version]
- Grunewald, W.; Karimi, M.; Wieczorek, K.; Van de Cappelle, E.; Wischnitzki, E.; Grundler, F.; Inzé, D.; Beeckman, T.; Gheysen, G. A role for AtWRKY23 in feeding site establishment of plant-parasitic nematodes. Plant Physiol. 2008, 148, 358–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattarai, K.K.; Atamian, H.S.; Kaloshian, I.; Eulgem, T. WRKY72-type transcription factors contribute to basal immunity in tomato and Arabidopsis as well as gene-for-gene resistance mediated by the tomato R gene Mi-1. Plant J. 2010, 63, 229–240. [Google Scholar] [CrossRef]
- Ali, M.A.; Wieczorek, K.; Kreil, D.P.; Bohlmann, H. The beet cyst nematode Heterodera schachtii modulates the expression of WRKY transcription factors in syncytia to favour its development in Arabidopsis roots. PLoS ONE 2014, 9, e102360. [Google Scholar] [CrossRef] [Green Version]
- Shim, J.S.; Choi, Y.D. Direct regulation of WRKY70 by AtMYB44 in plant defense responses. Plant Signal. Behav. 2013, 8, e24509. [Google Scholar] [CrossRef] [Green Version]
- Shim, J.S.; Jung, C.; Lee, S.; Min, K.; Lee, Y.-W.; Choi, Y.; Lee, J.S.; Song, J.T.; Kim, J.-K.; Choi, Y.D. AtMYB44 regulates WRKY70 expression and modulates antagonistic interaction between salicylic acid and jasmonic acid signaling. Plant J. 2013, 73, 483–495. [Google Scholar] [CrossRef]
- Hamamouch, N.; Winkel, B.S.J.; Li, C.; Davis, E.L. Modulation of Arabidopsis flavonol biosynthesis genes by cyst and root-knot nematodes. Plants 2020, 9, 253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piya, S.; Kihm, C.; Rice, J.H.; Baum, T.J.; Hewezi, T. Cooperative regulatory functions of miR858 and MYB83 during cyst nematode parasitism. Plant Physiol. 2017, 174, 1897–1912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sijmons, P.C.; Grundler, F.M.W.; Mende, N.; Burrows, P.R.; Wyss, U. Arabidopsis thaliana as a new model host for plant-parasitic nematodes. Plant J. 1991, 1, 245–254. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [Green Version]
- Chomczynski, P.; Sacchi, N. The single-step method of RNA isolation by acid guanidinium thiocyanate–phenol–chloroform extraction: Twenty-something years on. Nat. Protoc. 2006, 1, 581–585. [Google Scholar] [CrossRef] [PubMed]
- Grundler, F.; Hofmann, J. Identification of reference genes for qRT-PCR studies of gene expression in giant cells and syncytia induced in Arabidopsis thaliana by Meloidogyne incognita and Heterodera schachtii. Nematology 2007, 9, 317–323. [Google Scholar] [CrossRef]
- Higo, K.; Ugawa, Y.; Iwamoto, M.; Korenaga, T. Plant cis-acting regulatory DNA elements (PLACE) database: 1999. Nucleic Acids Res. 1999, 27, 297–300. [Google Scholar] [CrossRef] [Green Version]
- Wiśniewska, A.; Dąbrowska-Bronk, J.; Szafrański, K.; Fudali, S.; Święcicka, M.; Czarny, M.; Wilkowska, A.; Morgiewicz, K.; Matusiak, J.; Sobczak, M.; et al. Analysis of tomato gene promoters activated in syncytia induced in tomato and potato hairy roots by Globodera rostochiensis. Transgenic Res. 2013, 22, 557–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siddique, S.; Matera, C.; Radakovic, Z.S.; Shamim Hasan, M.; Gutbrod, P.; Rozanska, E.; Sobczak, M.; Torres, M.A.; Grundler, F.M.W. Parasitic worms stimulate host NADPH oxidases to produce reactive oxygen species that limit plant cell death and promote infection. Sci. Signal. 2014, 7, ra33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Różańska, E.; Czarnocka, W.; Baranowski, Ł.; Mielecki, J.; de Almeida Engler, J.; Sobczak, M. Expression of both Arabidopsis γ-tubulin genes is essential for development of a functional syncytium induced by Heterodera schachtii. Plant Cell Rep. 2018, 37, 1279–1292. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Transcription Factor or cis Element Name | Signal Sequence | New Place Ref. | Number | Description |
|---|---|---|---|---|
| ABRELATERD1 | ACGTG | S000414 | 1 | ABA- and dehydration responsive |
| ABRERATCAL | MACGYGB | S000507 | 1 | ABA- and dehydration responsive |
| ACGTATERD1 | ACGT | S000415 | 2 | dehydration responsive |
| ANAERO1CONSENSUS | AAACAAA | S000477 | 2 | anaerobically induced |
| BIHD1OS | TGTCA | S000498 | 1 | resistance responsive* |
| CATATGGMSAUR | CATATG | S000370 | 2 | auxin responsive |
| CCAATBOX1 | CCAAT | S000030 | 2 | heat shock responsive |
| CIACADIANLELHC | CAANNNNATC | S000252 | 1 | circadian related |
| CMSRE1IBSPOA | TGGACGG | S000511 | 1 | carbohydrate metabolite signal responsive element 1 |
| DPBFCOREDCDC3 | ACACNNG | S000292 | 2 | ABA responsive |
| EBOXBNNAPA | CANNTG | S000144 | 4 | light responsive, phenylpropanoid biosynthesis related |
| EECCRCAH1 | GANTTNC | S000494 | 1 | CO2 responsive |
| GT1CONSENSUS | GRWAAW | S000198 | 6 | light regulated |
| MYB1AT | WAACCA | S000408 | 1 | Dehydration responsive |
| MYBCORE | CNGTTR | S000176 | 2 | dehydration responsive, flavonoid biosynthesis related |
| MYCCONSENSUSAT | CANNTG | S000407 | 4 | dehydration and cold responsive |
| NODCON1GM | AAAGAT | S000461 | 1 | nodule specific |
| OSE1ROOTNODULE | AAAGAT | S000467 | 3 | nodule specific |
| PALBOXAPC | CCGTCC | S000137 | 1 | fungal elicitor, wounding and light responsive |
| POLASIG3 | AATAAT | S000088 | 2 | polyA signal |
| QARBNEXTA | AACGTGT | S000244 | 1 | wounding responsive |
| RAV1AAT | CAACA | S000314 | 4 | ABA, drought and cold responsive |
| T/GBOXATPIN2 | AACGTG | S000458 | 1 | jasmonate-responsive |
| TATABOX2 | TATAAAT | S000109 | 1 | TATA box |
| TATABOX4 | TATATAA | S000111 | 2 | TATA box |
| TATAPVTRNALEU | TTTATATA | S000340 | 1 | TATA box-like |
| WUSATAg | TTAATGG | S000433 | 1 | root meristem specific |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wiśniewska, A.; Wojszko, K.; Różańska, E.; Lenarczyk, K.; Kuczerski, K.; Sobczak, M. Arabidopsis thaliana Myb59 Gene Is Involved in the Response to Heterodera schachtii Infestation, and Its Overexpression Disturbs Regular Development of Nematode-Induced Syncytia. Int. J. Mol. Sci. 2021, 22, 6450. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22126450
Wiśniewska A, Wojszko K, Różańska E, Lenarczyk K, Kuczerski K, Sobczak M. Arabidopsis thaliana Myb59 Gene Is Involved in the Response to Heterodera schachtii Infestation, and Its Overexpression Disturbs Regular Development of Nematode-Induced Syncytia. International Journal of Molecular Sciences. 2021; 22(12):6450. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22126450
Chicago/Turabian StyleWiśniewska, Anita, Kamila Wojszko, Elżbieta Różańska, Klaudia Lenarczyk, Karol Kuczerski, and Mirosław Sobczak. 2021. "Arabidopsis thaliana Myb59 Gene Is Involved in the Response to Heterodera schachtii Infestation, and Its Overexpression Disturbs Regular Development of Nematode-Induced Syncytia" International Journal of Molecular Sciences 22, no. 12: 6450. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22126450