
Molecular Mechanisms of ZC3H12C/Reg-3 Biological Activity and Its Involvement in Psoriasis Pathology

 , , , and
, , , and 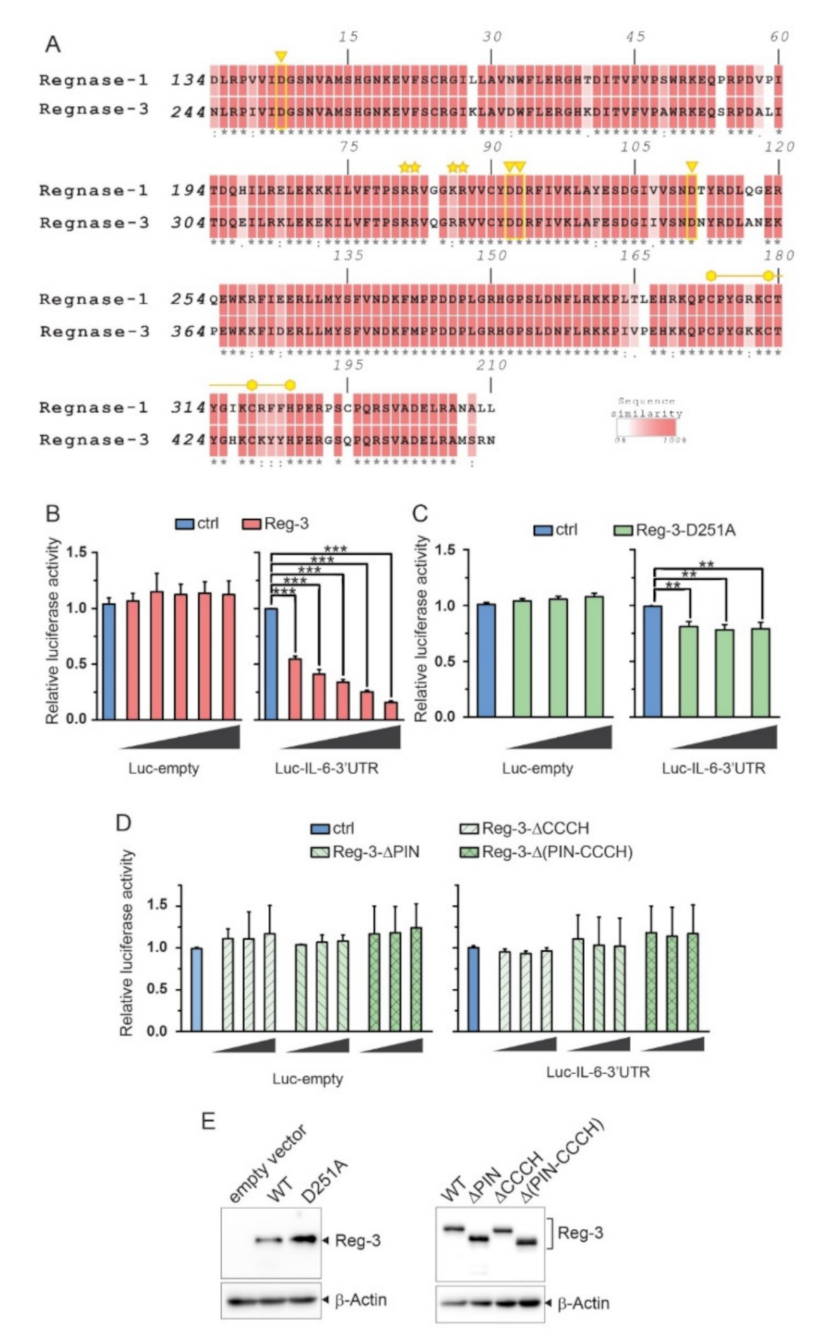{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Reg-3 Possesses Highly Conserved Domains Crucial for RNase Activity of other Members of the ZC3H12/MCPIP/Regnase Family
2.2. The Importance of the NYN/PIN and CCCH Domains in Reg-3 Nucleolytic Activity
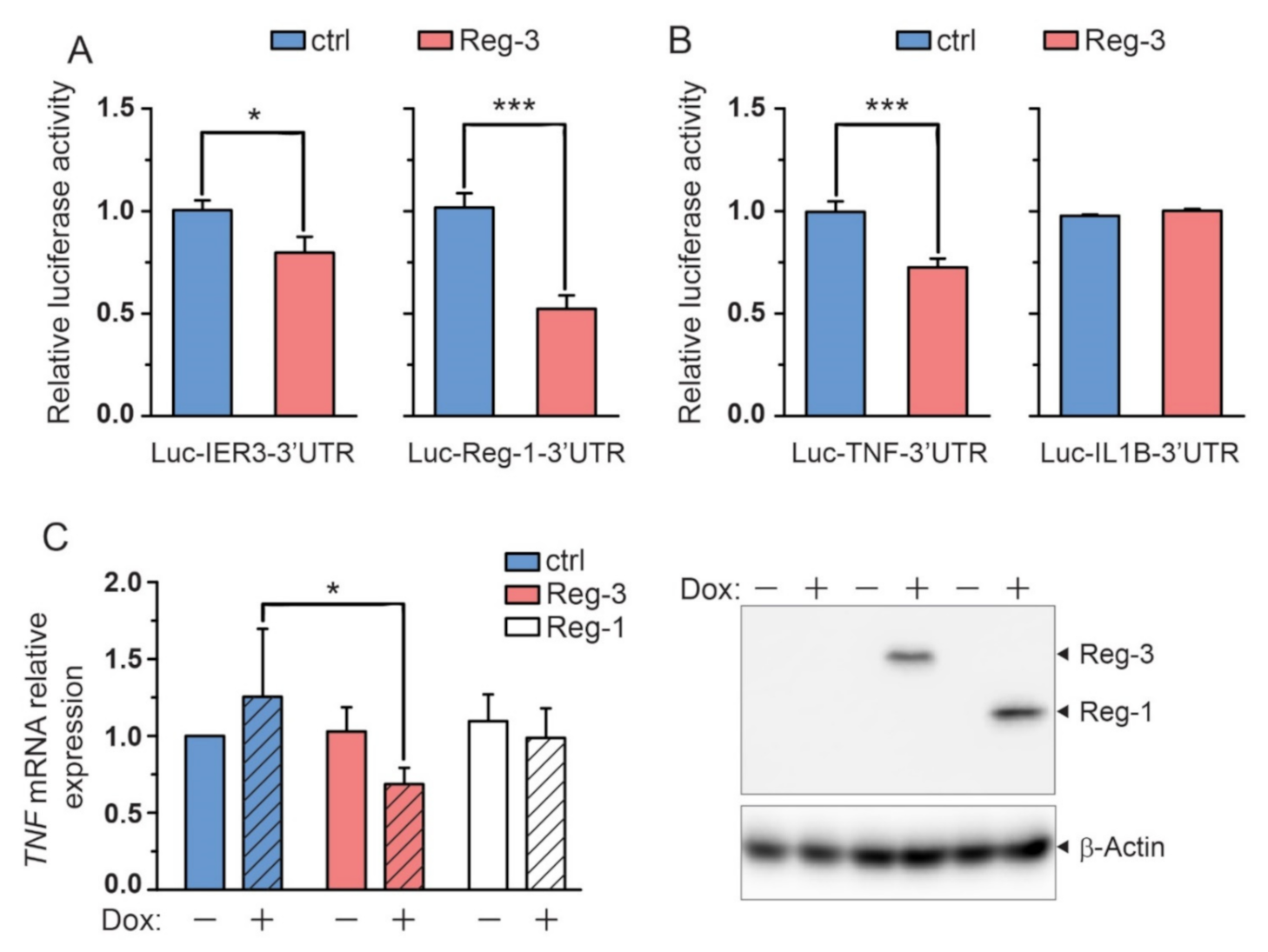
2.3. The Known Substrates of Reg-1, Reg-2 and Reg-4 Are also Substrates of Reg-3
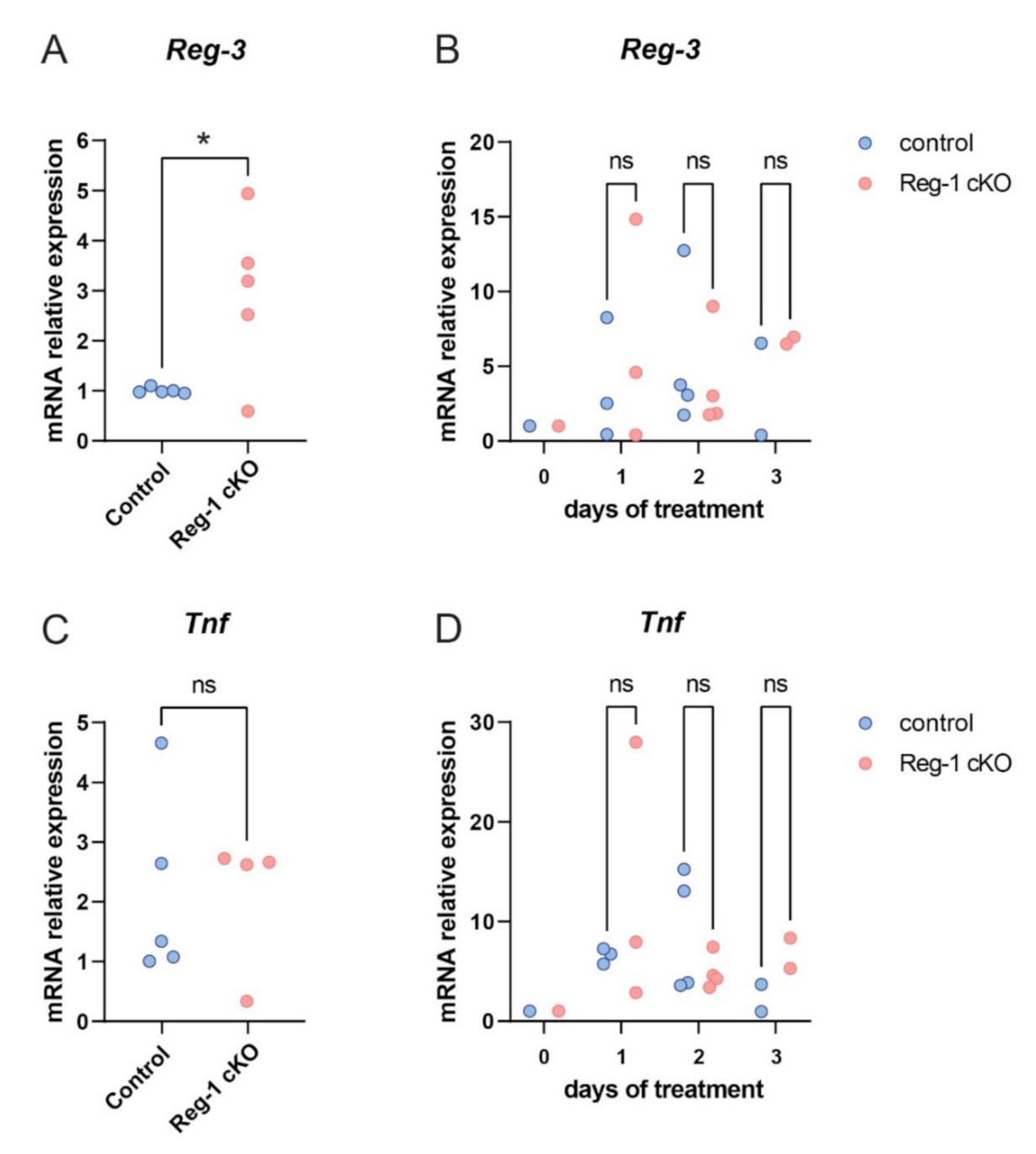
2.4. Reg-3 Regulates the Level of TNF but Not IL-1 Transcript
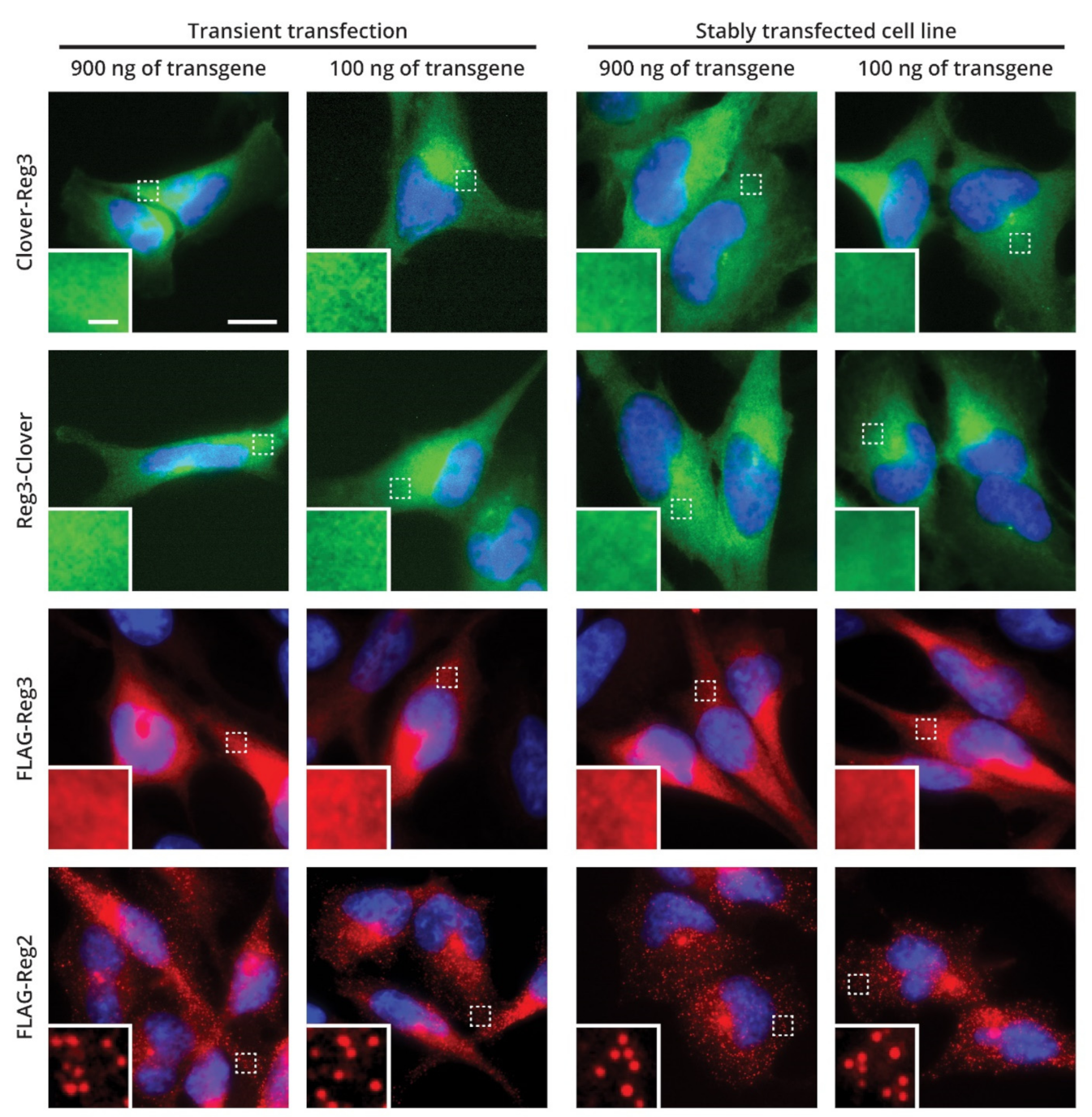
2.5. Cellular Localization of Reg-3
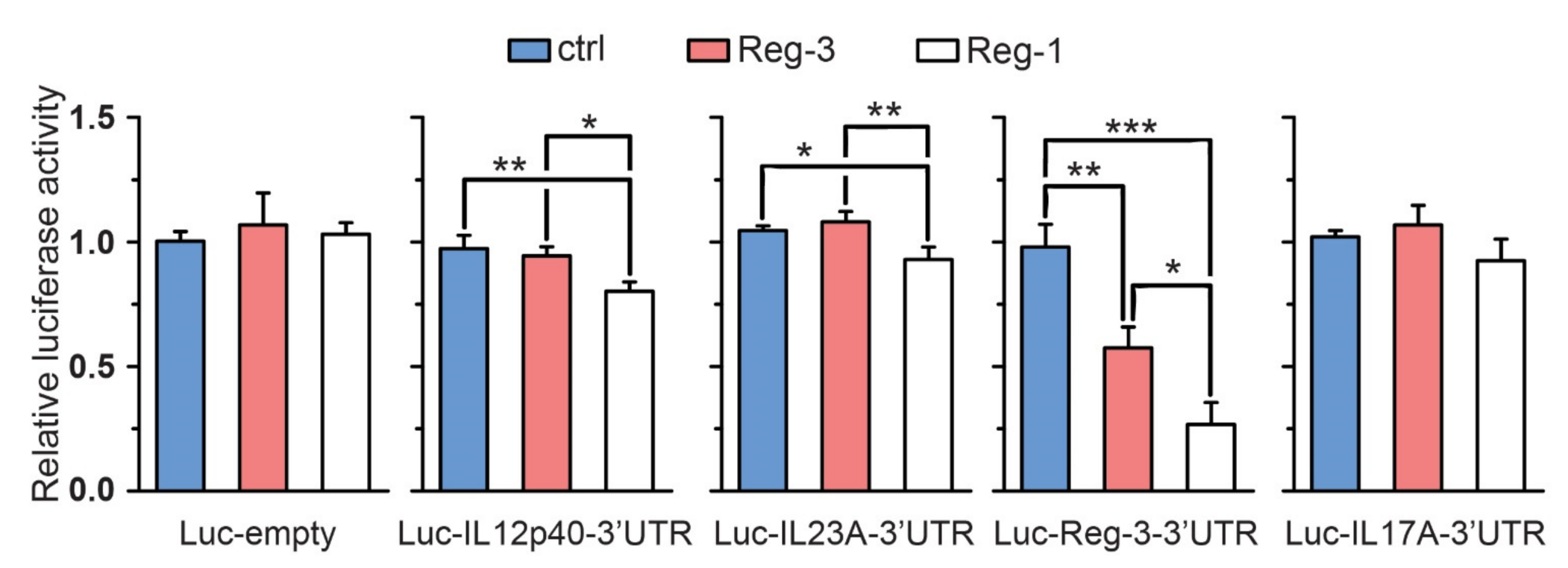
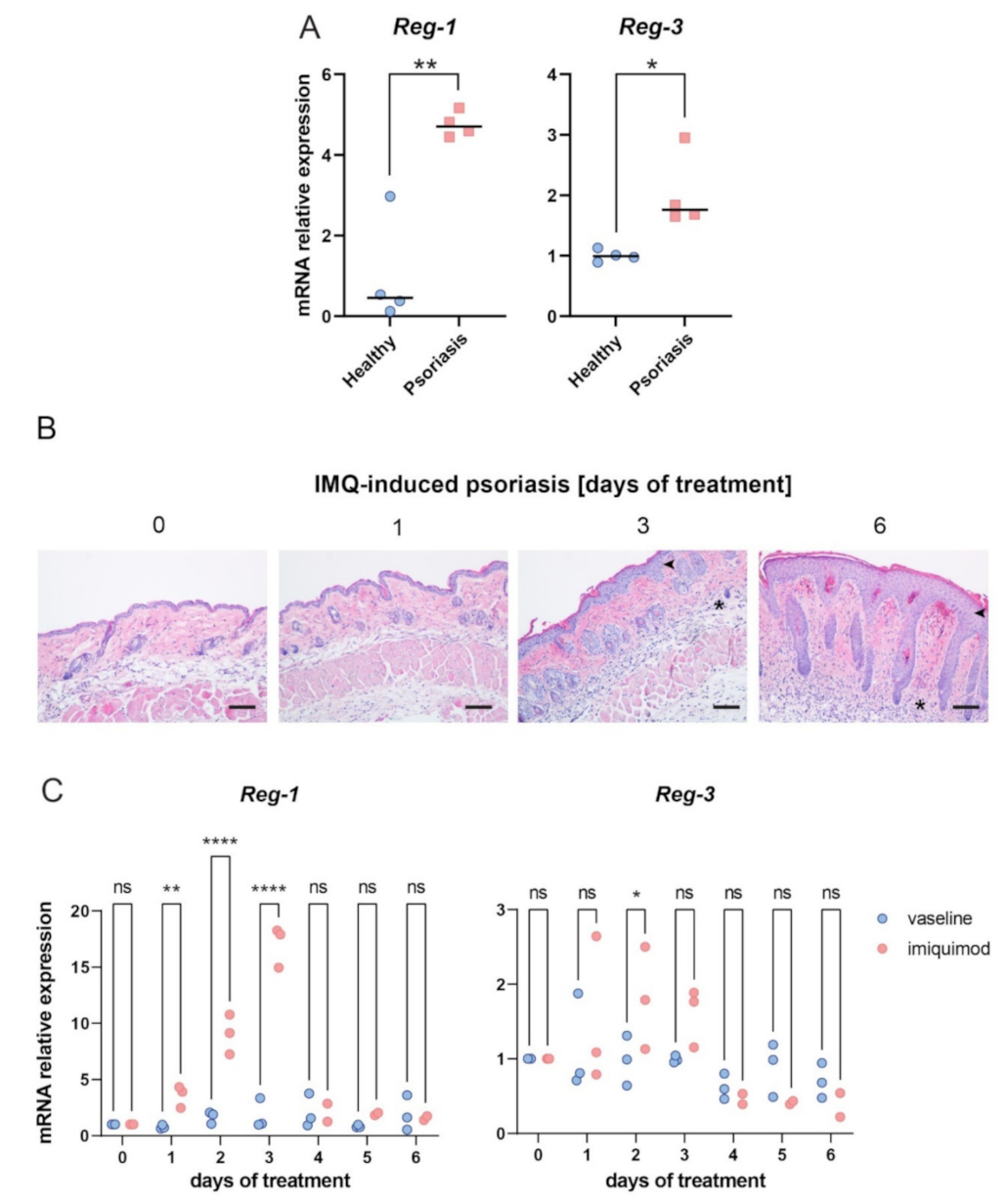
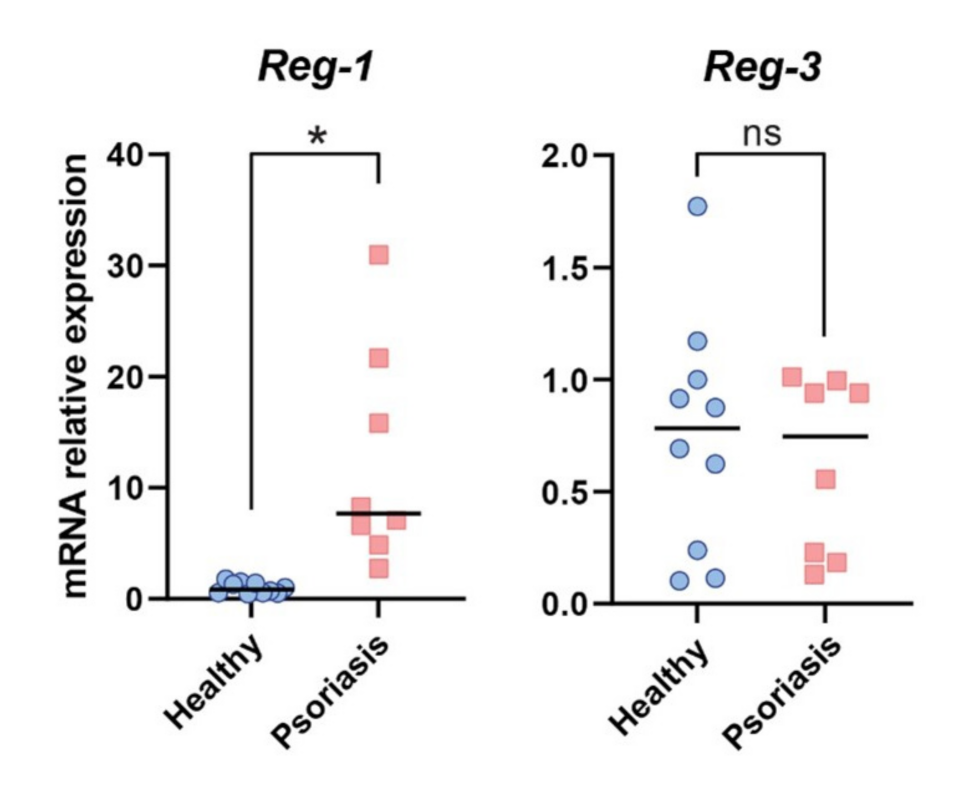
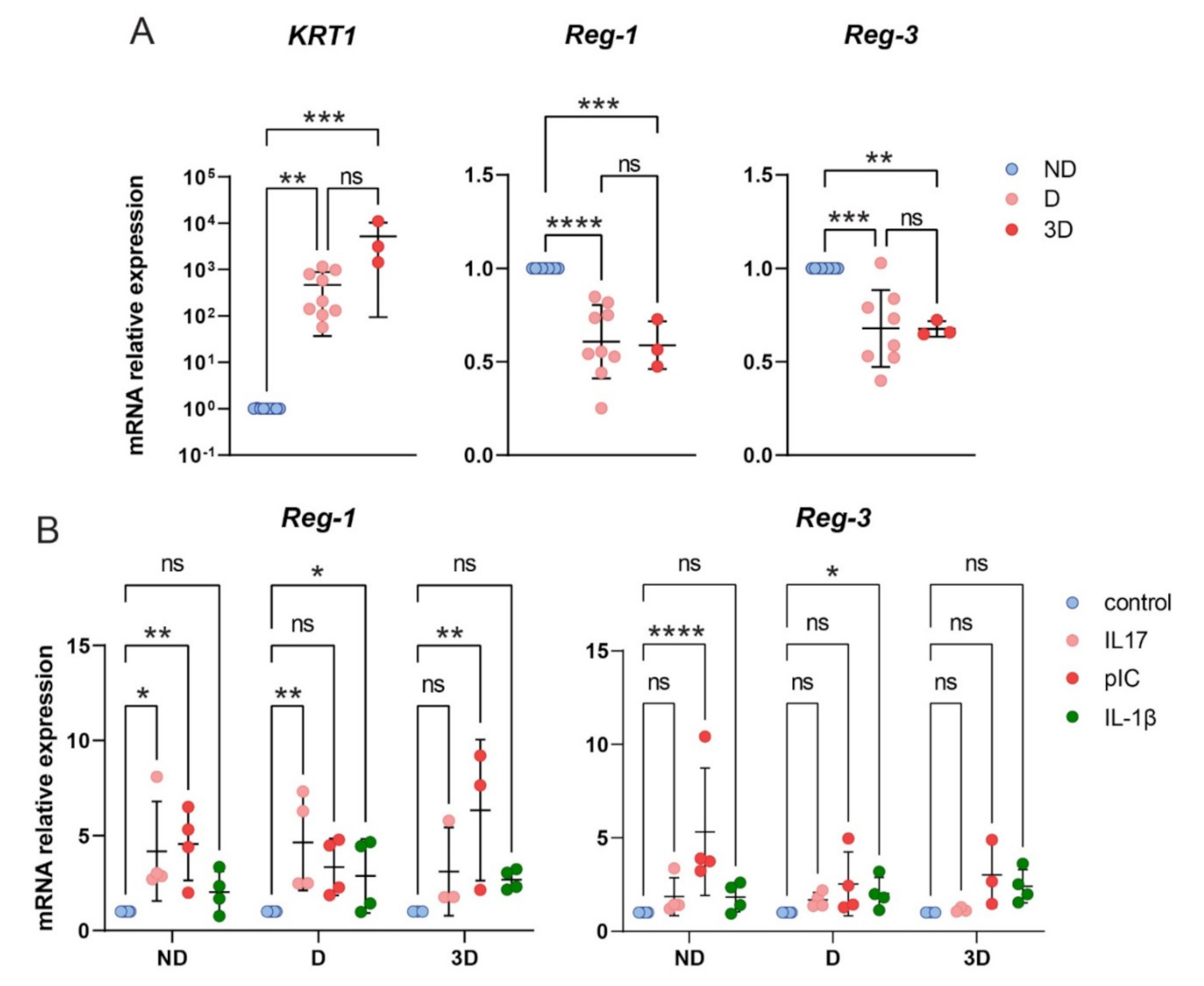
2.6. The Role of Reg-3 in Psoriasis
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Clinical Material
4.3. Isolation of Epidermis and Cell Culture
4.4. Experimental Psoriasis
4.5. Histology
4.6. Cell Line Transfections
4.7. Establishment of U251-MG Cell Lines with Doxycycline Inducible Expression of Regnase-3 and Regnase-1
4.8. Plasmid Construction
4.9. Luciferase Assays
4.10. RNA Isolation and Reverse Transcription
4.11. Quantitative PCR (RT-qPCR)
4.12. Western Blot Analysis
4.13. Immunofluorescence
4.14. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Matsushita, K.; Takeuchi, O.; Standley, D.M.; Kumagai, Y.; Kawagoe, T.; Miyake, T.; Satoh, T.; Kato, H.; Tsujimura, T.; Nakamura, H.; et al. Zc3h12a is an RNase essential for controlling immune responses by regulating mRNA decay. Nature 2009. [Google Scholar] [CrossRef]
- Mizgalska, D.; Wgrzyn, P.; Murzyn, K.; Kasza, A.; Koj, A.; Jura, J.; Jarzb, B.; Jura, J. Interleukin-1-inducible MCPIP protein has structural and functional properties of RNase and participates in degradation of IL-1β mRNA. FEBS J. 2009, 276. [Google Scholar] [CrossRef]
- Mino, T.; Murakawa, Y.; Fukao, A.; Vandenbon, A.; Wessels, H.H.; Ori, D.; Uehata, T.; Tartey, S.; Akira, S.; Suzuki, Y.; et al. Regnase-1 and roquin regulate a common element in inflammatory mRNAs by spatiotemporally distinct mechanisms. Cell 2015, 161, 1058–1073. [Google Scholar] [CrossRef] [Green Version]
- Lin, R.J.; Chien, H.L.; Lin, S.Y.; Chang, B.L.; Yu, H.P.; Tang, W.C.; Lin, Y.L. MCPIP1 ribonuclease exhibits broad-spectrum antiviral effects through viral RNA binding and degradation. Nucleic Acids Res. 2013, 41. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, H.I.; Arase, M.; Matsuyama, H.; Choi, Y.L.; Ueno, T.; Mano, H.; Sugimoto, K.; Miyazono, K. MCPIP1 ribonuclease antagonizes dicer and terminates microRNA biogenesis through precursor microRNA degradation. Mol. Cell 2011, 44. [Google Scholar] [CrossRef]
- Wawro, M.; Kochan, J.; Krzanik, S.; Jura, J.; Kasza, A. Intact NYN/PIN-Like Domain is Crucial for the Degradation of Inflammation-Related Transcripts by ZC3H12D. J. Cell. Biochem. 2017, 118. [Google Scholar] [CrossRef]
- Wawro, M.; Wawro, K.; Kochan, J.; Solecka, A.; Sowinska, W.; Lichawska-Cieslar, A.; Jura, J.; Kasza, A. ZC3H12B/MCPIP2, a new active member of the ZC3H12 family. RNA 2019, 25, 840–856. [Google Scholar] [CrossRef]
- Liang, J.; Wang, J.; Azfer, A.; Song, W.; Tromp, G.; Kolattukudy, P.E.; Fu, M. A novel CCCH-zinc finger protein family regulates proinflammatory activation of macrophages. J. Biol. Chem. 2008, 283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, J.; Saad, Y.; Lei, T.; Wang, J.; Qi, D.; Yang, Q.; Kolattukudy, P.E.; Fu, M. MCP-induced protein 1 deubiquitinates TRAF proteins and negatively regulates JNK and NF-κB signaling. J. Exp. Med. 2010, 207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, S.; Qi, D.; Liang, J.; Miao, R.; Minagawa, K.; Quinn, T.; Matsui, T.; Fan, D.; Liu, J.; Fu, M. The putative tumor suppressor Zc3h12d modulates toll-like receptor signaling in macrophages. Cell Signal. 2012, 24. [Google Scholar] [CrossRef] [Green Version]
- Minagawa, K.; Wakahashi, K.; Kawano, H.; Nishikawa, S.; Fukui, C.; Kawano, Y.; Asada, N.; Sato, M.; Sada, A.; Katayama, Y.; et al. Posttranscriptional Modulation of Cytokine Production in T Cells for the Regulation of Excessive Inflammation by TFL. J. Immunol. 2014. [Google Scholar] [CrossRef] [Green Version]
- Von Gamm, M.; Schaub, A.; Jones, A.N.; Wolf, C.; Behrens, G.; Lichti, J.; Essig, K.; Macht, A.; Pircher, J.; Ehrlich, A.; et al. Immune homeostasis and regulation of the interferon pathway require myeloid-derived Regnase-3. J. Exp. Med. 2019, 216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munir, S.; ber Rahman, S.; Rehman, S.; Saba, N.; Ahmad, W.; Nilsson, S.; Mazhar, K.; Naluai, Å.T. Association analysis of GWAS and candidate gene loci in a Pakistani population with psoriasis. Mol. Immunol. 2015, 64. [Google Scholar] [CrossRef]
- Tsoi, L.; Knight, J.; Ellinghaus, E.; Stuart, P.E.; Capon, F.; Ding, J.; Li, Y.; Tejasvi, T.; Gudjonsson, J.E.; Kang, H.M.; et al. Identification of fifteen new psoriasis susceptibility loci highlights the role of innate immunity. Nat. Genet. 2012, 44. [Google Scholar] [CrossRef] [Green Version]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; Mcgettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Peng, W.; Sun, Y.; Wang, X.; Xu, Y.; Li, X.; Gao, G.; Rao, Z. Structural study of MCPIP1 N-terminal conserved domain reveals a PIN-like RNase. Nucleic Acids Res. 2012, 40. [Google Scholar] [CrossRef] [Green Version]
- Sowińska, W.; Wawro, M.; Solecka, A.; Kasza, A. Potential limitations of the Sleeping Beauty transposon use in gene expression studies. Acta Biochim. Pol. 2019, 66, 263–268. [Google Scholar] [CrossRef]
- Di Cesare, A.; Di Meglio, P.; Nestle, F.O. The IL-23Th17 axis in the immunopathogenesis of psoriasis. J. Investig. Dermatol. 2009, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skrzeczynska-Moncznik, J.; Zabieglo, K.; Osiecka, O.; Morytko, A.; Brzoza, P.; Drozdz, L.; Kapinska-Mrowiecka, M.; Korkmaz, B.; Pastuszczak, M.; Kosalka-Wegiel, J.; et al. Differences in Staining for Neutrophil Elastase and its Controlling Inhibitor SLPI Reveal Heterogeneity among Neutrophils in Psoriasis. J. Investig. Dermatol. 2020, 140. [Google Scholar] [CrossRef]
- van der Fits, L.; Mourits, S.; Voerman, J.S.A.; Kant, M.; Boon, L.; Laman, J.D.; Cornelissen, F.; Mus, A.-M.; Florencia, E.; Prens, E.P.; et al. Imiquimod-Induced Psoriasis-Like Skin Inflammation in Mice Is Mediated via the IL-23/IL-17 Axis. J. Immunol. 2009, 182. [Google Scholar] [CrossRef] [PubMed]
- Mylonas, A.; Conrad, C. Psoriasis: Classical vs. Paradoxical: The yin-yang of TNF and Type i interferon. Front. Immunol. 2018, 9. [Google Scholar] [CrossRef]
- Konieczny, P.; Lichawska-Cieslar, A.; Kwiecinska, P.; Cichy, J.; Pietrzycka, R.; Szukala, W.; Declercq, W.; Devos, M.; Paziewska, A.; Rumienczyk, I.; et al. Keratinocyte-specific ablation of Mcpip1 impairs skin integrity and promotes local and systemic inflammation. J. Mol. Med. 2019, 97, 1669–1684. [Google Scholar] [CrossRef] [PubMed]
- Banas, M.; Zegar, A.; Kwitniewski, M.; Zabieglo, K.; Marczynska, J.; Kapinska-Mrowiecka, M.; LaJevic, M.; Zabel, B.A.; Cichy, J. The expression and regulation of chemerin in the epidermis. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [PubMed]
- Kajita, A.; Morizane, S.; Takiguchi, T.; Yamamoto, T.; Yamada, M.; Iwatsuki, K. Interferon-Gamma Enhances TLR3 Expression and Anti-Viral Activity in Keratinocytes. J. Investig. Dermatol. 2015, 135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiricozzi, A.; Guttman-Yassky, E.; Suárez-Farĩas, M.; Nograles, K.E.; Tian, S.; Cardinale, I.; Chimenti, S.; Krueger, J.G. Integrative responses to IL-17 and TNF-α in human keratinocytes account for key inflammatory pathogenic circuits in psoriasis. J. Investig. Dermatol. 2011, 131. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Liu, S.; Fu, J.J.; Wang, T.T.; Yao, X.; Kumar, A.; Liu, G.; Fu, M. Monocyte chemotactic protein-induced protein 1 and 4 form a complex but act independently in regulation of interleukin-6 mRNA degradation. J. Biol. Chem. 2015, 290. [Google Scholar] [CrossRef] [Green Version]
- Kochan, J.; Wawro, M.; Kasza, A. Simultaneous detection of mRNA and protein in single cells using immunofluorescence-combined single-molecule RNA FISH. Biotechniques 2015, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wheeler, J.R.; Matheny, T.; Jain, S.; Abrisch, R.; Parker, R. Distinct stages in stress granule assembly and disassembly. eLife 2016, 5. [Google Scholar] [CrossRef]
- Wawro, M.; Kochan, J.; Kasza, A. The perplexities of the ZC3H12A self-mRNA regulation. Acta Biochim. Pol. 2016, 63. [Google Scholar] [CrossRef]
- Lichawska-Cieslar, A.; Konieczny, P.; Szukala, W.; Declercq, W.; Fu, M.; Jura, J. Loss of keratinocyte Mcpip1 abruptly activates the IL-23/Th17 and Stat3 pathways in skin inflammation. Biochim. Biophys. Acta Mol. Cell Res. 2021, 1868. [Google Scholar] [CrossRef]
- Ruiz-Romeu, E.; Ferran, M.; Giménez-Arnau, A.; Bugara, B.; Lipert, B.; Jura, J.; Florencia, E.F.; Prens, E.P.; Celada, A.; Pujol, R.M.; et al. MCPIP1 RNase Is Aberrantly Distributed in Psoriatic Epidermis and Rapidly Induced by IL-17A. J. Investig. Dermatol. 2016, 136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seneschal, J.; Milpied, B.; Vergier, B.; Lepreux, S.; Schaeverbeke, T.; Taïeb, A. Cytokine imbalance with increased production of interferon-α in psoriasiform eruptions associated with antitumour necrosis factor-α treatments. Br. J. Dermatol. 2009, 161. [Google Scholar] [CrossRef]
- Van Kuppeveld, F.J.; van der Logt, J.T.; Angulo, A.F.; van Zoest, M.J.; Quint, W.G.; Niesters, H.G.; Galama, J.M.; Melchers, W.J. Genus- and species-specific identification of mycoplasmas by 16S rRNA amplification. Appl. Environ. Microbiol. 1993, 59, 655. [Google Scholar] [CrossRef] [Green Version]
- Skrzeczynska-Moncznik, J.; Wlodarczyk, A.; Zabieglo, K.; Kapinska-Mrowiecka, M.; Marewicz, E.; Dubin, A.; Potempa, J.; Cichy, J. Secretory Leukocyte Proteinase Inhibitor-Competent DNA Deposits Are Potent Stimulators of Plasmacytoid Dendritic Cells: Implication for Psoriasis. J. Immunol. 2012, 189. [Google Scholar] [CrossRef] [Green Version]
- Zabieglo, K.; Majewski, P.; Majchrzak-Gorecka, M.; Wlodarczyk, A.; Grygier, B.; Zegar, A.; Kapinska-Mrowiecka, M.; Naskalska, A.; Pyrc, K.; Dubin, A.; et al. The inhibitory effect of secretory leukocyte protease inhibitor (SLPI) on formation of neutrophil extracellular traps. J. Leukoc. Biol. 2015, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mátés, L.; Chuah, M.K.L.; Belay, E.; Jerchow, B.; Manoj, N.; Acosta-Sanchez, A.; Grzela, D.P.; Schmitt, A.; Becker, K.; Matrai, J.; et al. Molecular evolution of a novel hyperactive Sleeping Beauty transposase enables robust stable gene transfer in vertebrates. Nat. Genet. 2009. [Google Scholar] [CrossRef] [PubMed]
- Kowarz, E.; Löscher, D.; Marschalek, R. Optimized Sleeping Beauty transposons rapidly generate stable transgenic cell lines. Biotechnol. J. 2015, 10, 647–653. [Google Scholar] [CrossRef] [PubMed]
- Kochan, J.; Wawro, M.; Kasza, A. IF-combined smRNA FISH reveals interaction of MCPIP1 protein with IER3 mRNA. Biol. Open 2016, 5. [Google Scholar] [CrossRef] [Green Version]
- Chomczynski, P.; Sacchi, N. The single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction: Twenty-something years on. Nat. Protoc. 2006, 1, 581–585. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wawro, M.; Kochan, J.; Sowinska, W.; Solecka, A.; Wawro, K.; Morytko, A.; Kwiecinska, P.; Grygier, B.; Kwitniewski, M.; Fu, M.; et al. Molecular Mechanisms of ZC3H12C/Reg-3 Biological Activity and Its Involvement in Psoriasis Pathology. Int. J. Mol. Sci. 2021, 22, 7311. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147311
Wawro M, Kochan J, Sowinska W, Solecka A, Wawro K, Morytko A, Kwiecinska P, Grygier B, Kwitniewski M, Fu M, et al. Molecular Mechanisms of ZC3H12C/Reg-3 Biological Activity and Its Involvement in Psoriasis Pathology. International Journal of Molecular Sciences. 2021; 22(14):7311. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147311
Chicago/Turabian StyleWawro, Mateusz, Jakub Kochan, Weronika Sowinska, Aleksandra Solecka, Karolina Wawro, Agnieszka Morytko, Patrycja Kwiecinska, Beata Grygier, Mateusz Kwitniewski, Mingui Fu, and et al. 2021. "Molecular Mechanisms of ZC3H12C/Reg-3 Biological Activity and Its Involvement in Psoriasis Pathology" International Journal of Molecular Sciences 22, no. 14: 7311. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147311