
Optimal Treatment of 6-Dimethylaminopurine Enhances the In Vivo Development of Canine Embryos by Rapid Initiation of DNA Synthesis


Abstract
:1. Introduction
2. Results
2.1. Effect of Activation Treatment on DNA Synthesis of Parthenogenetic Zygotes
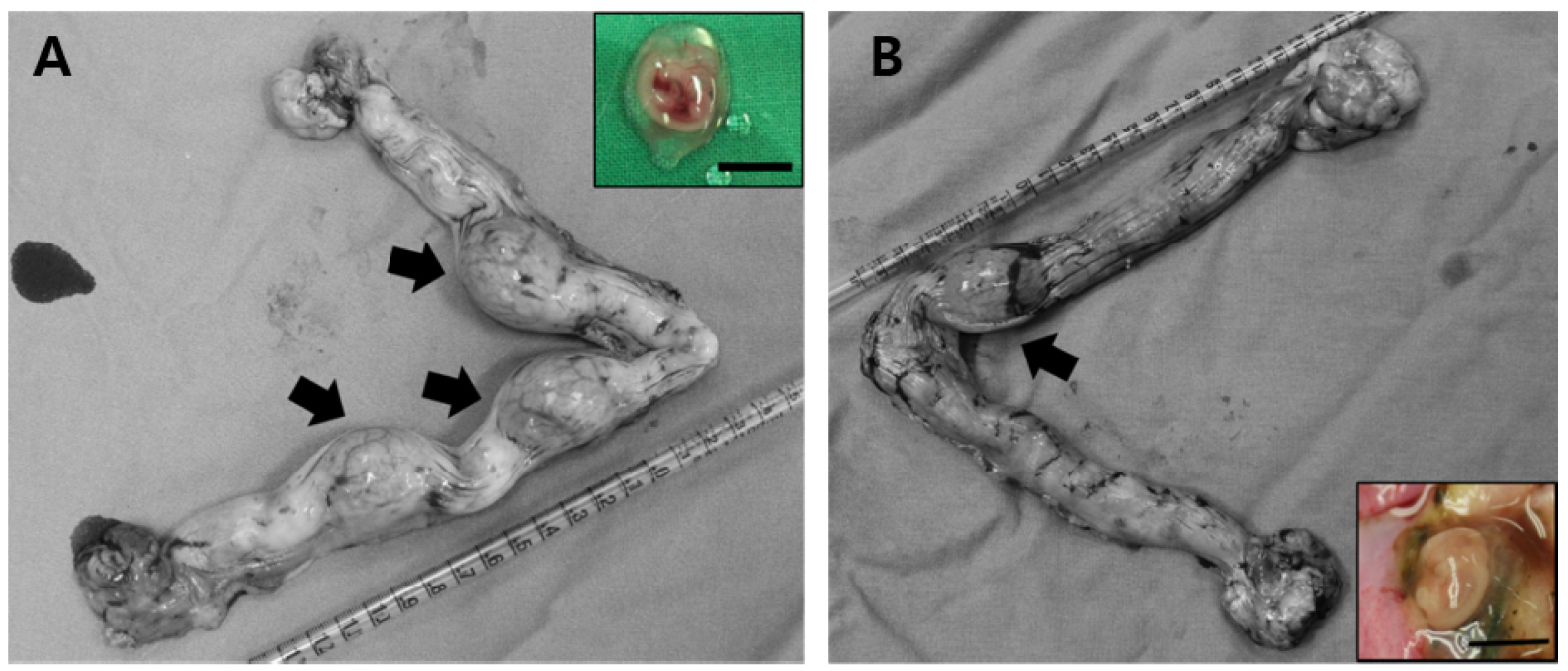
2.2. Effect of Activation Treatment on In Vivo Development of Parthenogenetic Zygotes
2.3. Effect of Activation Treatment on DNA Synthesis of Reconstructed Oocytes Produced by SCNT
2.4. Effect of Activation Treatment on In Vivo Development of Canine Reconstructed Oocytes by SCNT
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Animals
4.3. Preparation of Donor Cells
4.4. Collection of In Vivo Matured Oocytes
4.5. Parthenogenetic Activation
4.6. Somatic Cell Nuclear Transfer
4.7. Staining with 5-Bromo-20-Deoxyuridine Staining for Evaluation of DNA Synthesis in Embryos
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wilmut, I.; Schnieke, A.E.; McWhir, J.; Kind, A.J.; Campbell, K.H. Viable offspring derived from fetal and adult mammalian cells. Nature 1997, 385, 810–813. [Google Scholar] [CrossRef]
- Kishigami, S.; Wakayama, S.; Thuan, N.V.; Ohta, H.; Mizutani, E.; Hikichi, T.; Bui, H.T.; Balbach, S.; Ogura, A.; Boiani, M.; et al. Production of cloned mice by somatic cell nuclear transfer. Nat. Protoc. 2006, 1, 125–138. [Google Scholar] [CrossRef]
- Cibelli, J.B.; Stice, S.L.; Golueke, P.J.; Kane, J.J.; Jerry, J.; Blackwell, C.; Ponce de Leon, F.A.; Robl, J.M. Cloned transgenic calves produced from nonquiescent fetal fibroblasts. Science 1998, 280, 1256–1258. [Google Scholar] [CrossRef] [Green Version]
- Lee, B.C.; Kim, M.K.; Jang, G.; Oh, H.J.; Yuda, F.; Kim, H.J.; Hossein, M.S.; Kim, J.J.; Kang, S.K.; Schatten, G.; et al. Dogs cloned from adult somatic cells. Nature 2005, 436, 641. [Google Scholar] [CrossRef] [PubMed]
- Alberio, R.; Zakhartchenko, V.; Motlik, J.; Wolf, E. Mammalian oocyte activation: Lessons from the sperm and implications for nuclear transfer. Int. J. Dev. Biol. 2001, 45, 797–809. [Google Scholar]
- Fissore, R.A.; Robl, J.M. Mechanism of calcium oscillations in fertilized rabbit eggs. Dev. Biol. 1994, 166, 634–642. [Google Scholar] [CrossRef]
- Ducibella, T.; Fissore, R. The roles of Ca2+, downstream protein kinases, and oscillatory signaling in regulating fertilization and the activation of development. Dev. Biol. 2008, 315, 257–279. [Google Scholar] [CrossRef] [Green Version]
- Campbell, K.H.; Loi, P.; Otaegui, P.J.; Wilmut, I. Cell cycle co-ordination in embryo cloning by nuclear transfer. Rev. Reprod. 1996, 1, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Ju, J.C.; Yang, X. Differential inactivation of maturation-promoting factor and mitogen-activated protein kinase following parthenogenetic activation of bovine oocytes. Biol. Reprod. 1998, 59, 537–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgan, A.J.; Jacob, R. Ionomycin enhances Ca2+ influx by stimulating store-regulated cation entry and not by a direct action at the plasma membrane. Biochem. J. 1994, 300, 665–672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ledda, S.; Loi, P.; Bogliolo, L.; Moor, R.M.; Fulka, J., Jr. The effect of 6-dimethylaminopurine (6-DMAP) on DNA synthesis in activated mammalian oocytes. Zygote 1996, 4, 7–9. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.H.; Love, L.B.; Westhusin, M.E.; Hinrichs, K. Activation of equine nuclear transfer oocytes: Methods and timing of treatment in relation to nuclear remodeling. Biol. Reprod. 2004, 70, 46–53. [Google Scholar] [CrossRef]
- Dinnyes, A.; Dai, Y.; Barber, M.; Liu, L.; Xu, J.; Zhou, P.; Yang, X. Development of cloned embryos from adult rabbit fibroblasts: Effect of activation treatment and donor cell preparation. Biol. Reprod. 2001, 64, 257–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loi, P.; Ledda, S.; Fulka, J., Jr.; Cappai, P.; Moor, R.M. Development of parthenogenetic and cloned ovine embryos: Effect of activation protocols. Biol. Reprod. 1998, 58, 1177–1187. [Google Scholar] [CrossRef] [Green Version]
- Park, J.E.; Kim, M.J.; Ha, S.K.; Hong, S.G.; Oh, H.J.; Kim, G.A.; Park, E.J.; Kang, J.T.; Saadeldin, I.M.; Jang, G.; et al. Altered cell cycle gene expression and apoptosis in post-implantation dog parthenotes. PLoS ONE 2012, 7, e41256. [Google Scholar] [CrossRef] [PubMed]
- Jang, G.; Oh, H.J.; Kim, M.K.; Fibrianto, Y.H.; Hossein, M.S.; Kim, H.J.; Kim, J.J.; Hong, S.G.; Park, J.E.; Kang, S.K.; et al. Improvement of canine somatic cell nuclear transfer procedure. Theriogenology 2008, 69, 146–154. [Google Scholar] [CrossRef]
- Sanders, E.J.; Varedi, M.; French, A.S. Cell proliferation in the gastrulating chick embryo: A study using BrdU incorporation and PCNA localization. Development 1993, 118, 389–399. [Google Scholar] [CrossRef]
- Odartchenko, N.; Keneklis, T. Localization of paternal DNA in interphase nuclei of mouse eggs during early cleavage. Nature 1973, 241, 528–529. [Google Scholar] [CrossRef]
- Moussa, M.; Perreau, C.; Baril, G.; Duchamp, G.; Vidament, M.; Daels, P.; Bruyas, J.F.; Mermillod, P. Comparison of cell proliferation index in equine and caprine embryos using a modified BrdU incorporation assay. Theriogenology 2005, 64, 1823–1832. [Google Scholar] [CrossRef] [PubMed]
- Soloy, E.; Kanka, J.; Viuff, D.; Smith, S.D.; Callesen, H.; Greve, T. Time course of pronuclear deoxyribonucleic acid synthesis in parthenogenetically activated bovine oocytes. Biol. Reprod. 1997, 57, 27–35. [Google Scholar] [CrossRef] [Green Version]
- De La Fuente, R.; King, W.A. Developmental consequences of karyokinesis without cytokinesis during the first mitotic cell cycle of bovine parthenotes. Biol. Reprod. 1998, 58, 952–962. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Lee, C.; Moor, R.M. DNA synthesis, microtubule and nuclear dynamics in porcine parthenotes. Zygote 1996, 4, 139–144. [Google Scholar] [CrossRef]
- Alberio, R.; Kubelka, M.; Zakhartchenko, V.; Hajduch, M.; Wolf, E.; Motlik, J. Activation of bovine oocytes by specific inhibition of cyclin-dependent kinases. Mol. Reprod. Dev. 2000, 55, 422–432. [Google Scholar] [CrossRef]
- Szollosi, M.S.; Kubiak, J.Z.; Debey, P.; de Pennart, H.; Szollosi, D.; Maro, B. Inhibition of protein kinases by 6-dimethylaminopurine accelerates the transition to interphase in activated mouse oocytes. J. Cell Sci. 1993, 104, 861–872. [Google Scholar] [CrossRef] [PubMed]
- Susko-Parrish, J.L.; Leibfried-Rutledge, M.L.; Northey, D.L.; Schutzkus, V.; First, N.L. Inhibition of protein kinases after an induced calcium transient causes transition of bovine oocytes to embryonic cycles without meiotic completion. Dev. Biol. 1994, 166, 729–739. [Google Scholar] [CrossRef]
- Moses, R.M.; Masui, Y. Enhancement of mouse egg activation by the kinase inhibitor, 6-dimethylaminopurine (6-DMAP). J. Exp. Zool. 1994, 270, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Winger, Q.A.; De La Fuente, R.; King, W.A.; Armstrong, D.T.; Watson, A.J. Bovine parthenogenesis is characterized by abnormal chromosomal complements: Implications for maternal and paternal co-dependence during early bovine development. Dev. Genet. 1997, 21, 160–166. [Google Scholar] [CrossRef]
- Kubelka, M.; Motlik, J.; Schultz, R.M.; Pavlok, A. Butyrolactone I reversibly inhibits meiotic maturation of bovine oocytes, Without influencing chromosome condensation activity. Biol. Reprod. 2000, 62, 292–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mermillod, P.; Tomanek, M.; Marchal, R.; Meijer, L. High developmental competence of cattle oocytes maintained at the germinal vesicle stage for 24 hours in culture by specific inhibition of MPF kinase activity. Mol. Reprod. Dev. 2000, 55, 89–95. [Google Scholar] [CrossRef]
- Fancsovits, P.; Toth, L.; Takacs, Z.F.; Murber, A.; Papp, Z.; Urbancsek, J. Early pronuclear breakdown is a good indicator of embryo quality and viability. Fertil. Steril. 2005, 84, 881–887. [Google Scholar] [CrossRef] [PubMed]
- Neuber, E.; Rinaudo, P.; Trimarchi, J.R.; Sakkas, D. Sequential assessment of individually cultured human embryos as an indicator of subsequent good quality blastocyst development. Hum. Reprod. 2003, 18, 1307–1312. [Google Scholar] [CrossRef] [Green Version]
- Fenwick, J.; Platteau, P.; Murdoch, A.P.; Herbert, M. Time from insemination to first cleavage predicts developmental competence of human preimplantation embryos in vitro. Hum. Reprod. 2002, 17, 407–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hase, M.; Hori, T.; Kawakami, E.; Tsutsui, T. Plasma LH and progesterone levels before and after ovulation and observation of ovarian follicles by ultrasonographic diagnosis system in dogs. J. Vet. Med. Sci. 2000, 62, 243–248. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Park, S.W.; Hossein, M.S.; Jeong, Y.W.; Kim, J.J.; Lee, E.; Kim, Y.W.; Hyun, S.H.; Shin, T.; Hwang, W.S. Production of cloned dogs by decreasing the interval between fusion and activation during somatic cell nuclear transfer. Mol. Reprod. Dev. 2009, 76, 483–489. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.A.; Oh, H.J.; Park, J.E.; Kim, M.J.; Park, E.J.; Jo, Y.K.; Jang, G.; Kim, M.K.; Kim, H.J.; Lee, B.C. Species-specific challenges in dog cloning. Reprod. Domest. Anim. Zuchthyg. 2012, 47 (Suppl. 6), 80–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.J.; Oh, H.J.; Park, J.E.; Hong, S.G.; Kang, J.T.; Koo, O.J.; Kang, S.K.; Jang, G.; Lee, B.C. Influence of oocyte donor and embryo recipient conditions on cloning efficiency in dogs. Theriogenology 2010, 74, 473–478. [Google Scholar] [CrossRef]
- Hong, S.G.; Jang, G.; Kim, M.K.; Oh, H.J.; Park, J.E.; Kang, J.T.; Koo, O.J.; Kim, D.Y.; Lee, B.C. Dogs cloned from fetal fibroblasts by nuclear transfer. Anim. Reprod. Sci. 2009, 115, 334–339. [Google Scholar] [CrossRef] [PubMed]
- Oh, H.J.; Park, E.J.; Lee, S.Y.; Soh, J.W.; Kong, I.S.; Choi, S.W.; Ra, J.C.; Kang, S.K.; Lee, B.C. Comparison of cell proliferation and epigenetic modification of gene expression patterns in canine foetal fibroblasts and adipose tissue-derived mesenchymal stem cells. Cell Prolif. 2012, 45, 438–444. [Google Scholar] [CrossRef] [PubMed]
- Vitullo, P.; Sciamanna, I.; Baiocchi, M.; Sinibaldi-Vallebona, P.; Spadafora, C. LINE-1 retrotransposon copies are amplified during murine early embryo development. Mol. Reprod. Dev. 2012, 79, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Motlik, J.; Alberio, R.; Zakhartchenko, V.; Stojkovic, M.; Kubelka, M.; Wolf, E. The effect of activation of Mammalian oocytes on remodeling of donor nuclei after nuclear transfer. Cloning Stem Cells 2002, 4, 245–252. [Google Scholar] [CrossRef]
- Bui, H.T.; Wakayama, S.; Kishigami, S.; Park, K.K.; Kim, J.H.; Thuan, N.V.; Wakayama, T. Effect of trichostatin A on chromatin remodeling, histone modifications, DNA replication, and transcriptional activity in cloned mouse embryos. Biol. Reprod. 2010, 83, 454–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Treatment | Timing (hpa) | No. Flushed Oocytes | No. PA Zygotes | No. PA Zygotes Labeled with BrdU (%) |
|---|---|---|---|---|
| DMAP-2h | 2 | 10 | 10 | 10 (100) a |
| 4 | 12 | 12 | 12 (100) | |
| 8 | 12 | 12 | 12 (100) | |
| DMAP-4h | 2 | 10 | 10 | 9 (90) b |
| 4 | 12 | 12 | 12 (100) | |
| 8 | 11 | 11 | 11 (100) |
| Treatment | Trial No. | No. Flushed Oocytes | No. Transferred PA Zygotes | Pregnancy Rate 1 | No. Implantations (%) 2 |
|---|---|---|---|---|---|
| DMAP-2h | 1 | 11 | 11 | + | 3 |
| 2 | 10 | 10 | + | 8 | |
| 3 | 7 | 7 | + | 2 | |
| 4 | 14 | 14 | - | 0 | |
| Total | 42 | 42 | 75% | 13 (34) a | |
| DMAP-4h | 1 | 12 | 12 | + | 1 |
| 2 | 12 | 12 | - | 0 | |
| 3 | 9 | 9 | + | 1 | |
| Total | 33 | 33 | 66.7% | 2 (6.5) b |
| Treatment | Timing (hpa) | No. Flushed Oocytes | No. Reconstructed Oocytes | No. Oocytes Labeled with BrdU (%) |
|---|---|---|---|---|
| DMAP-2h | 2 | 26 | 20 | 18 (90) a |
| 4 | 16 | 12 | 12 (100) | |
| 8 | 18 | 14 | 14 (100) | |
| DMAP-4h | 2 | 23 | 18 | 14 (77.7) b |
| 4 | 15 | 12 | 12 (100) | |
| 8 | 18 | 14 | 14 (100) |
| Treatment | No. Flushed Oocytes | No. Reconstructed Oocytes (Transferred) | No. Recipients | No. Pregnant (%) 1 | No. Cloned Puppies (%) 2 |
|---|---|---|---|---|---|
| DMAP-2h | 232 | 182 | 12 | 5 (41.6) | 9 (4.9) |
| DMAP-4h | 242 | 188 | 12 | 4 (33.3) | 6 (3.2) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oh, H.J.; Lee, B.C.; Kim, M.K. Optimal Treatment of 6-Dimethylaminopurine Enhances the In Vivo Development of Canine Embryos by Rapid Initiation of DNA Synthesis. Int. J. Mol. Sci. 2021, 22, 7757. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147757
Oh HJ, Lee BC, Kim MK. Optimal Treatment of 6-Dimethylaminopurine Enhances the In Vivo Development of Canine Embryos by Rapid Initiation of DNA Synthesis. International Journal of Molecular Sciences. 2021; 22(14):7757. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147757
Chicago/Turabian StyleOh, Hyun Ju, Byeong Chun Lee, and Min Kyu Kim. 2021. "Optimal Treatment of 6-Dimethylaminopurine Enhances the In Vivo Development of Canine Embryos by Rapid Initiation of DNA Synthesis" International Journal of Molecular Sciences 22, no. 14: 7757. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147757