
CSH RNA Interference Reduces Global Nutrient Uptake and Umbilical Blood Flow Resulting in Intrauterine Growth Restriction

 , , and
, , and
Abstract
:1. Introduction
2. Results
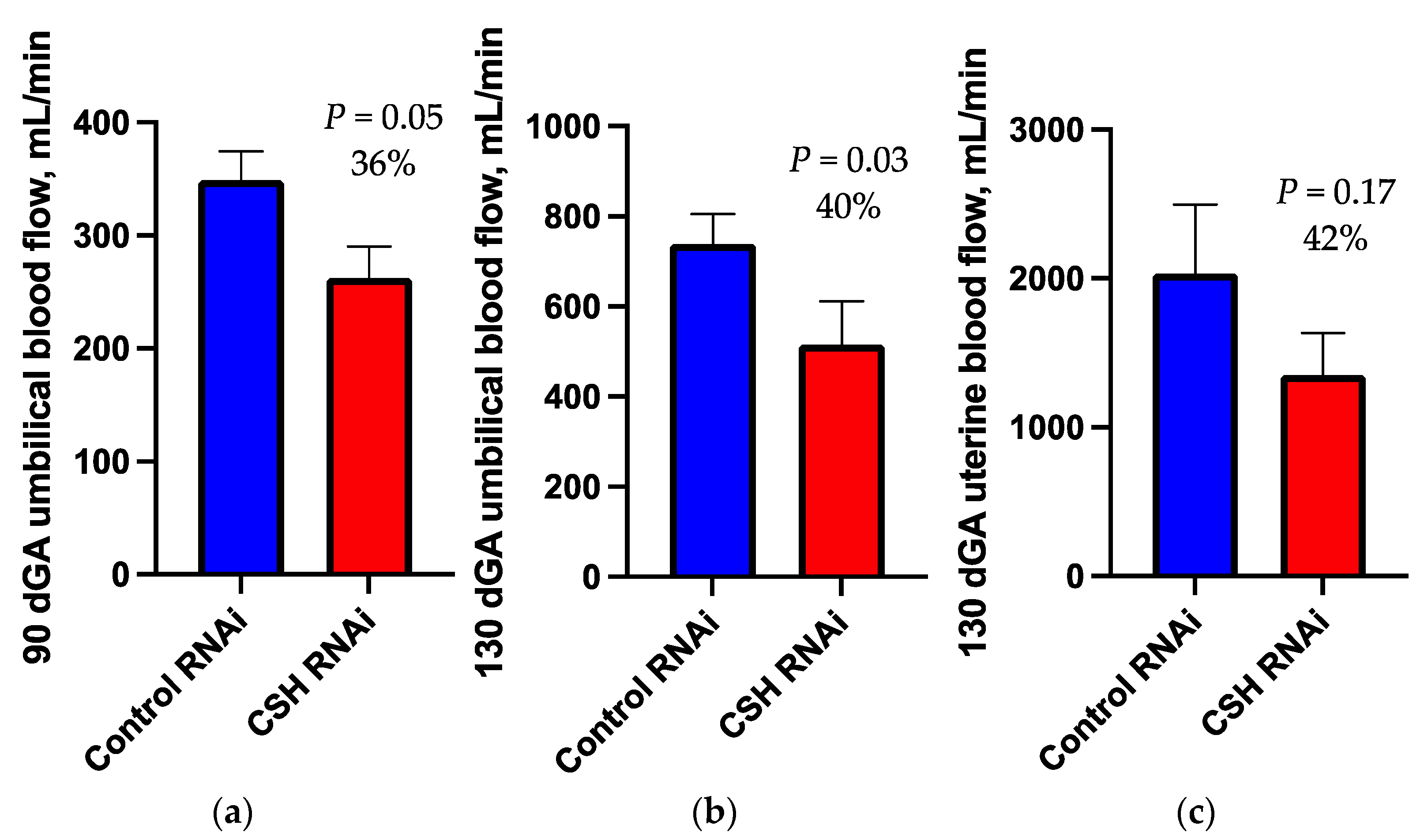
2.1. 90dGA Doppler Velocimetry
2.2. 130dGA Uterine and Umbilical Blood Flows
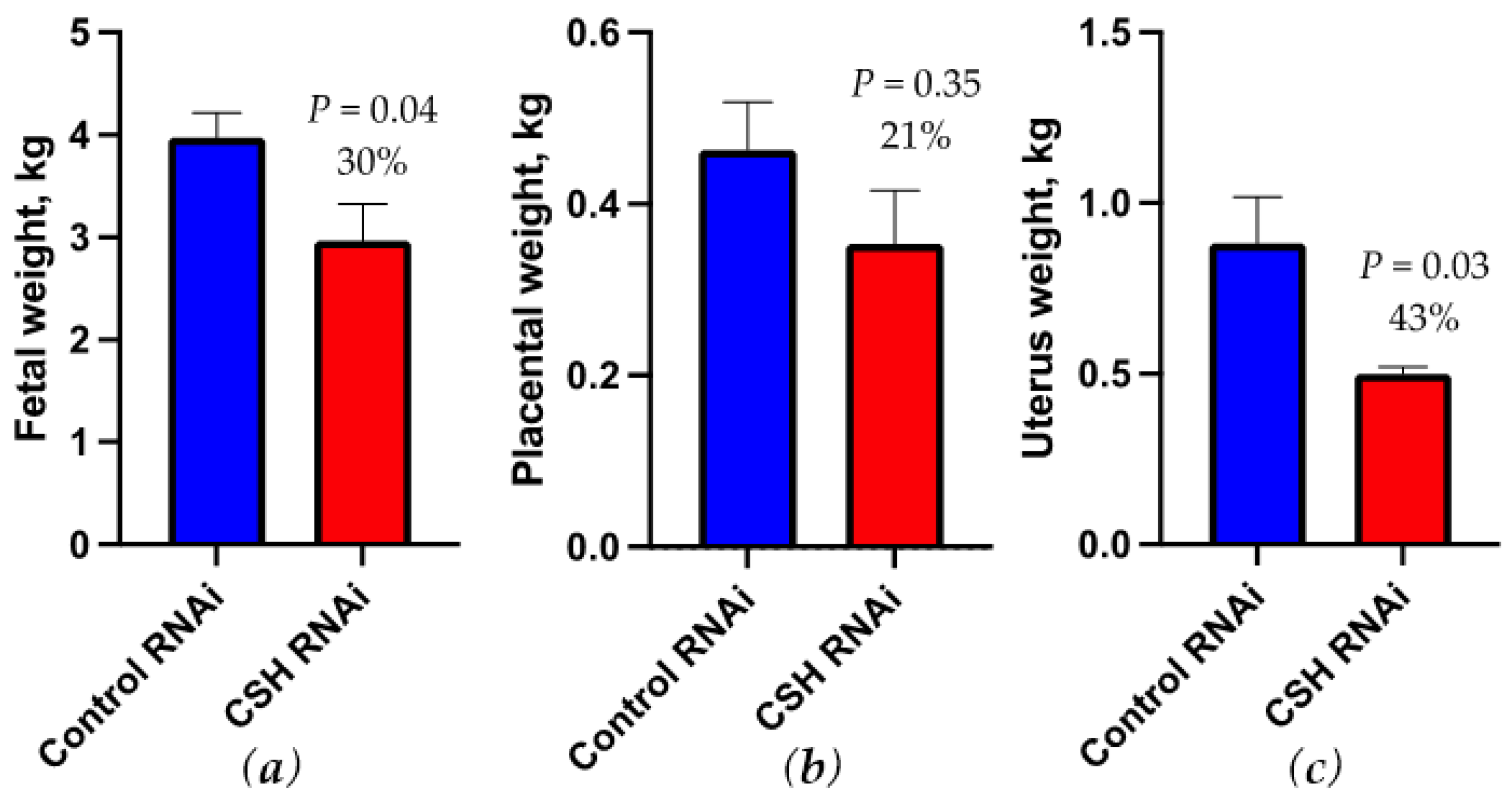
2.3. Fetal and Uteroplacental Characteristics Near-Term (130 dGA)
2.4. Blood Gas and Oxygen Uptakes
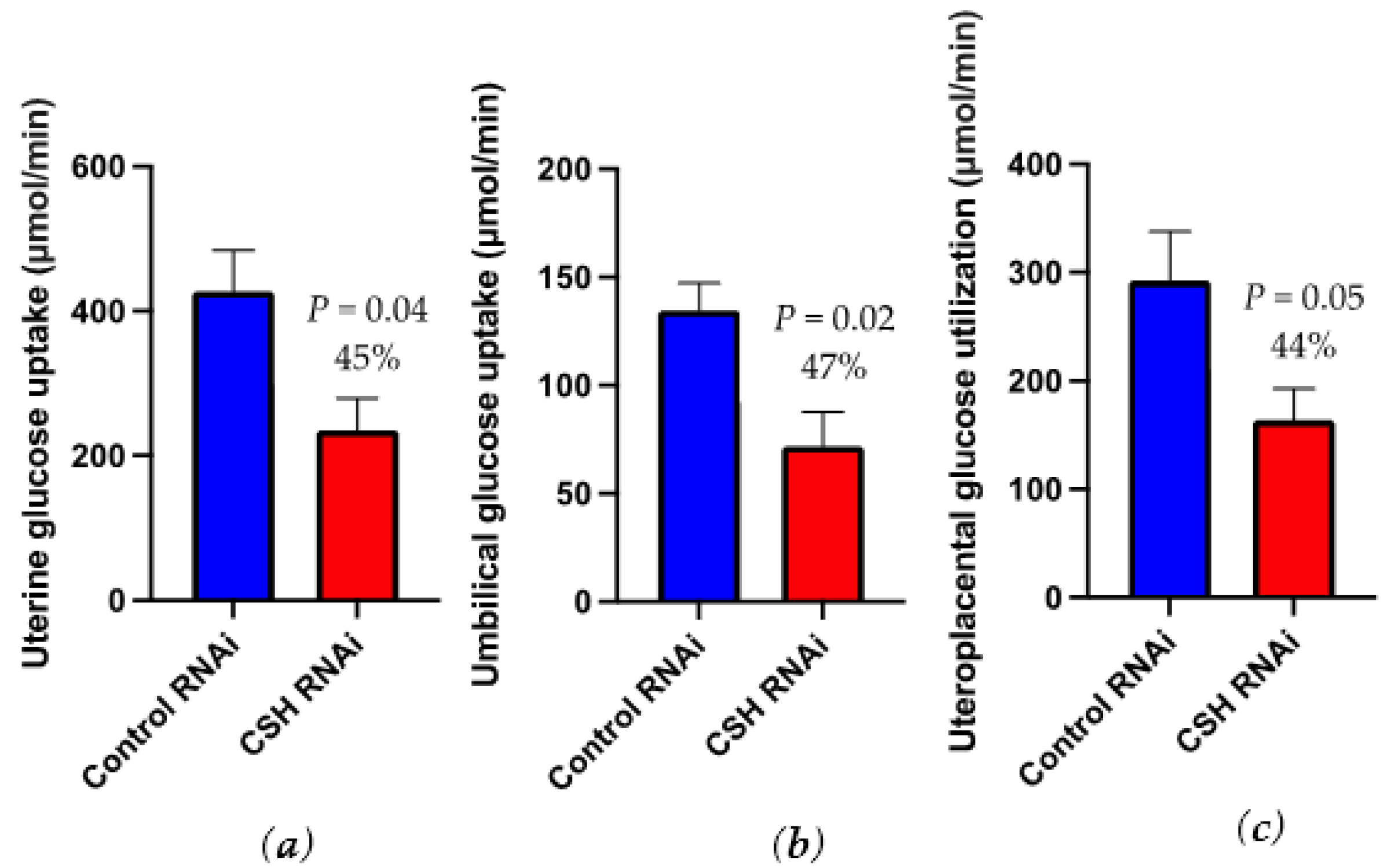
2.5. Glucose and Lactate Uptakes
2.6. Amino Acid Uptakes
2.7. Total Nutrient Uptakes
2.8. 130dGA Hormones
3. Discussion
4. Materials and Methods
4.1. Lentiviral Generation
4.2. Generation of CSH RNAi Pregnancies
4.3. Doppler Velocimetry
4.4. Surgical Instrumentation of Fetus and Ewe
4.5. Blood Flow Calculations and Tissue Collection
4.6. Biochemical Analysis of Blood Samples
4.7. Western Blot Analysis
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gagnon, R. Placental insufficiency and its consequences. Eur. J. Obs. Gynecol. Reprod. Biol. 2003, 110, S99–S107. [Google Scholar] [CrossRef]
- Barker, D.J.; Osmond, C. Low birth weight and hypertension. BMJ 1988, 297, 134–135. [Google Scholar] [CrossRef] [Green Version]
- Barker, D.J.; Bull, A.R.; Osmond, C.; Simmonds, S.J. Fetal and placental size and risk of hypertension in adult life. BMJ 1990, 301, 259–262. [Google Scholar] [CrossRef]
- Barker, D.J. The fetal and infant origins of adult disease. BMJ 1990, 301, 1111. [Google Scholar] [CrossRef] [Green Version]
- Hales, C.N.; Barker, D.J. The thrifty phenotype hypothesis. Br. Med. Bull. 2001, 60, 5–20. [Google Scholar] [CrossRef] [Green Version]
- Daikoku, N.H.; Tyson, J.E.; Graf, C.; Scott, R.; Smith, B.; Johnson, J.W.C.; King, T.M. The relative significance of human placental lactogen in the diagnosis of retarded fetal growth. Am. J. Obst. Gynecol. 1979, 135, 516–521. [Google Scholar] [CrossRef]
- Spellacy, W.N.; Buhi, W.C.; Birk, S.A. Human placental lactogen and intrauterine growth retardation. Obstet. Gynecol. 1976, 47, 446–448. [Google Scholar] [PubMed]
- Lea, R.G.; Wooding, P.; Stewart, I.; Hannah, L.T.; Morton, S.; Wallace, K.; Aitken, R.P.; Milne, J.S.; Regnault, T.R.; Anthony, R.V.; et al. The expression of ovine placental lactogen, StAR and progesterone-associated steroidogenic enzymes in placentae of overnourished growing adolescent ewes. Reproduction 2007, 133, 785–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, C.M.; Goetzmann, L.N.; Cantlon, J.D.; Jeckel, K.M.; Winger, Q.A.; Anthony, R.V. Development of ovine chorionic somatomammotropin hormone-deficient pregnancies. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2016, 310, R837–R846. [Google Scholar] [CrossRef] [Green Version]
- Jeckel, K.M.; Boyarko, A.C.; Bouma, G.J.; Winger, Q.A.; Anthony, R.V. Chorionic somatomammotropin impacts early fetal growth and placental gene expression. J. Endocrinol. 2018, 237, 301–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rygaard, K.; Revol, A.; Esquivel-Escobedo, D.; Beck, B.L.; Barrera-Saldaña, H.A. Absence of human placental lactogen and placental growth hormone (HGH-V) during pregnancy: PCR analysis of the deletion. Hum. Genet. 1998, 102, 87–92. [Google Scholar] [CrossRef]
- Ali, A.; Swanepoel, C.M.; Winger, Q.A.; Rozance, P.J.; Anthony, R.V. Chorionic somatomammotropin RNA interference alters fetal liver glucose utilization. J. Endocrinol. 2020, 247, 169–180. [Google Scholar] [CrossRef]
- Tanner, A.R.; Lynch, C.S.; Ali, A.; Winger, Q.A.; Rozance, P.J.; Anthony, R.V. Impact of chorionic somatomammotropin RNA interference on uterine blood flow and placental glucose uptake in the absence of intrauterine growth restriction. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2021, 320, R138–R148. [Google Scholar] [CrossRef]
- Galan, H.L.; Anthony, R.V.; Rigano, S.; Parker, T.A.; de Vrijer, B.; Ferrazi, E.; Wilkening, R.B.; Regnault, T.R.H. Fetal hypertension and abnormal Doppler velocimetry in an ovine model of intrauterine growth restriciton. Am. J. Obstet. Gynecol. 2005, 192, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Regnault, T.R.H.; Galan, H.L.; Parker, T.A.; Anthony, R.V. Placental development in normal and compromised pregnancies—A review. Placenta 2002, 16, S119–S129. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.; Bischof, H.; Lang, I.; Desoye, G.; Greenwood, S.L.; Johnstone, E.D.; Wareing, M.; Sibley, C.P.; Brownbill, P. Dysregulated flow-mediated vasodilatation in the human placenta in fetal growth restriction. J. Physiol. 2015, 593, 3077–3092. [Google Scholar] [CrossRef] [PubMed]
- Galan, H.L.; Hussey, M.J.; Chung, M.; Chyu, J.K.; Hobbins, J.C.; Battaglia, F.C. Doppler velocimetry of growth-restricted fetuses in an ovine model of placental insufficiency. Am. J. Obstet. Gynecol. 1998, 178, 451–456. [Google Scholar] [CrossRef]
- Arroyo, J.A.; Anthony, R.V.; Parker, T.A.; Galan, H.L. eNOS, NO, and the activation of ERK and AKT signaling at mid-gestation and near-term in an ovine model of intrauterine growth restriction. Sys. Biol. Reprod. Med. 2010, 56, 62–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Regnault, T.R.H.; de Vrijer, B.; Galan, H.L.; Davidsen, M.L.; Trembler, K.A.; Battaglia, F.C.; Wilkening, R.B.; Anthony, R.V. The relationship between transplacental O2 diffusion and placental expression of PlGF, VEGF and their receptors in a placental insufficiency model of fetal growth restriction. J. Physiol. 2003, 550, 641–656. [Google Scholar] [CrossRef]
- Vranas, S.; Heinemann, G.K.; Liu, H.; De Blasio, M.J.; Owens, J.A.; Gatford, K.L.; Black, M.J. Small size at birth predicts decreased cardiomyocyte number in the adult ovine heart. J. Dev. Orig. Health Dis. 2017, 8, 618–625. [Google Scholar] [CrossRef]
- Chang, E.I.; Rozance, P.J.; Wesolowski, S.R.; Nguyen, L.M.; Shaw, S.C.; Sclafani, R.A.; Bjorkman, K.K.; Peter, A.K.; Hay, W.W.; Brown, L.D. Rates of myogenesis and myofiber numbers are reduced in late gestation IUGR fetal sheep. J. Endocrinol. 2019, 244, 339–352. [Google Scholar] [CrossRef]
- Rozance, P.J.; Zastoupil, L.; Wesolowski, S.R.; Goldstrohm, D.A.; Strahan, B.; Cree-Green, M.; Sheffield-Moore, M.; Meschia, G.; Hay, W.W.; Wilkening, R.B.; et al. Skeletal muscle protein accretion rates and hindlimb growth are reduced in late gestation intrauterine growth-restricted fetal sheep. J. Physiol. 2018, 596, 67–82. [Google Scholar] [CrossRef] [Green Version]
- Spencer, T.E.; Gray, A.; Johnson, G.A.; Taylor, K.M.; Gerler, A.; Gootwine, E.; Ott, T.L.; Bazer, F.W. Effects of recombinant ovine interferon tau, placental lactogen, and growth hormone in the ovine uterus. Biol. Reprod. 1999, 61, 1409–1418. [Google Scholar] [CrossRef] [Green Version]
- Magness, R.R.; Shaw, C.E.; Phernetton, T.M.; Zheng, J.; Bird, I.M. Endothelial vasodilator production by uterine and systemic arteries. II. Pregnancy effects on NO synthase expression. Am. J. Physiol. 1997, 272, H1730–H1740. [Google Scholar] [CrossRef] [PubMed]
- Barry, J.S.; Anthony, R.V. The pregnant sheep as a model for human pregnancy. Theriogenology 2008, 69, 55–67. [Google Scholar] [CrossRef] [Green Version]
- Carter, A.M. Factors affecting gas transfer across the placenta and the oxygen supply to the fetus. J. Dev. Physiol. 1989, 12, 305–322. [Google Scholar] [PubMed]
- Regnault, T.R.H.; de Vrijer, B.; Galan, H.L.; Wilkening, R.B.; Battaglia, F.C.; Meschia, G. Development and mechanisms of fetal hypoxia in severe fetal growth restriction. Placenta 2007, 28, 714–723. [Google Scholar] [CrossRef] [PubMed]
- Kingdom, J.C.P.; Kaufmann, P. Current topic: Oxygen and placental villous development: Origins of fetal hypoxia. Placenta 1997, 18, 613–621. [Google Scholar] [CrossRef]
- Battaglia, F.C.; Meschia, G. An Introduction to Fetal Physiology; Academic Press: New York, NY, USA, 1986. [Google Scholar]
- Hay, W.W.; Molina, R.A.; DiGiacomo, J.E.; Meschia, G. Model of placental glucose consumption and glucose transfer. Am. J. Physiol. 1990, 258, R569–R577. [Google Scholar] [CrossRef]
- Jansson, T.; Wennergren, M.; Illsley, N.P. Glucose transporter protein expression in human placenta throughout gestation and in intrauterine growth retardation. J. Clin. Endocrinol. Metab. 1993, 77, 1554–1562. [Google Scholar] [PubMed]
- Hay, W.W. Energy and substrate requirements of the placenta and fetus. Proc. Nutr. Soc. 1991, 50, 321–336. [Google Scholar] [CrossRef] [Green Version]
- Lemons, J.A.; Adcock, E.W., III; Jones, M.D.; Naughton, M.A.; Meschia, G.; Battaglia, F.C. Umbilical uptake of amino acids in the unstressed fetal lamb. J. Clin. Investig. 1976, 58, 1428–1434. [Google Scholar] [CrossRef] [Green Version]
- Schneider, H.; Mohlen, K.H.; Dancis, J. Transfer of amino acids across the in vitro perfused human placenta. Ped. Res. 1979, 13, 236–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Battaglia, F.C. Glutamine and Glutamate Exchange between the Fetal Liver and the Placenta. J. Nutr. 2000, 130, 974S–977S. [Google Scholar] [CrossRef] [PubMed]
- Handwerger, S. Clinical counterpoint: The physiology of placental lactogen in human pregnancy. Endocr. Rev. 1991, 12, 329–336. [Google Scholar] [CrossRef] [PubMed]
- Breuer, C.B. Stimulation of DNA synthesis in cartilage of hypophysectomized rats by native and modified placental lactogen and available hormones. Endocrinology 1969, 85, 989. [Google Scholar] [CrossRef] [PubMed]
- Hurley, T.W.; D’Ercole, A.J.; Handwerger, S.; Underwood, L.E.; Fulanetto, R.W.; Fellows, R.E. Ovine placental lactogen induces somatomedin: A possible role in fetal growth. Endocrinology 1977, 101, 1635. [Google Scholar] [CrossRef]
- Daughaday, W.H.; Kapadia, M. Maintenance of serum somatomedin activity in hypophysectomized pregnant rats. Endocrinology 1978, 102, 1317. [Google Scholar] [CrossRef]
- Adams, S.O.; Nissley, S.P.; Handwerger, S.; Rechler, M.M. Developmental patterns of insulin-like growth factor I and II synthesis and regulation in rat fibroblasts. Nature 1983, 302, 150–153. [Google Scholar] [CrossRef] [PubMed]
- Kniss, D.A.; Shubert, P.J.; Zimmerman, P.D.; Landon, M.B.; Gabbe, S.G. Insulinlike growth factors. Their regulation of glucose and amino acid transport in placental trophoblasts isolated from first-trimester chorionic villi. J. Reprod. Med. 1994, 39, 249–256. [Google Scholar] [PubMed]
- Martin, J.W.; Friesen, H.G. Effect of human placental lactogen on the isolated islets of Langerhans in vitro. Endocrinology 1969, 84, 619. [Google Scholar] [CrossRef]
- Oliver, M.H.; Harding, J.E.; Breier, B.H.; Evans, P.C.; Gallaher, B.W.; Gluckman, P.D. The effects of ovine placental lactogen infusion on metabolites, insulin-like growth factors and binding proteins in the fetal sheep. J. Endocrinol. 1995, 144, 333–338. [Google Scholar] [CrossRef]
- National Research Council. Nutrient Requirements of Small Ruminants: Sheep, Goats, Cervids, and New World Camelids; The National Academies Press: Washington, DC, USA, 2007. [Google Scholar]
- Bonds, D.R.; Anderson, S.; Meschia, G. Transplacental diffusion of ethanol under steady state conditions. J. Dev. Physiol. 1980, 2, 409–416. [Google Scholar]
- Brown, L.D.; Rozance, P.J.; Bruce, J.L.; Friedman, J.E.; Hay, W.W.; Wesolowski, S.R. Limited capacity for glucose oxidation in fetal sheep with intrauterine growth restriction. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2015, 309, R920–R928. [Google Scholar] [CrossRef] [Green Version]
- Hay, W.W.; Sparks, J.W.; Quissell, B.J.; Battaglia, F.C.; Meschia, G. Simultaneous measurements of umbilical uptake, fetal utilization rate, and fetal turnover rate of glucose. Am. J. Physiol. Endocrinol. Metab. 1981, 240, E662–E668. [Google Scholar] [CrossRef]
- Jones, A.K.; Rozance, P.J.; Brown, L.D.; Goldstrohm, D.A.; Hay, W.W.; Limesand, S.W.; Wesolowski, S.R. Sustained hypoxemia in late gestation potentiates hepatic gluconeogenic gene expression but does not activate glucose production in the ovine fetus. Am. J. Physiol. Endocrinol. Metab. 2019, 317, E1–E10. [Google Scholar] [CrossRef]
- Meschia, G.; Cotter, J.R.; Breathnach, C.S.; Barron, D.H. Simultaneous measurement of uterine and umbilical blood flows and oxygen uptake. Q. J. Exp. Physiol. 1966, 52, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Cilvik, S.N.; Wesolowski, S.R.; Anthony, R.V.; Brown, L.D.; Rozance, P.J. Late gestation fetal hyperglucagonaemia impairs placental function and results in diminished fetal protein accretion and decreased fetal growth. J. Physiol. 2021, 599, 3403–3427. [Google Scholar] [CrossRef] [PubMed]
- Meschia, G.; Battaglia, F.C.; Hay, W.W.; Sparks, J.W. Utilization of substrates by the ovine placenta in vivo. Fed. Proc. 1980, 39, 245–249. [Google Scholar] [PubMed]
- Andrews, S.E.; Brown, L.D.; Thorn, S.R.; Limesand, S.W.; Davis, M.; Hay, W.W.; Rozance, P.J. Increased adrenergic signaling is responsible for decreased glucose-stimulated insulin secretion in the chronically hyperinsulinemic ovine fetus. Endocrinology 2015, 156, 367–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benjamin, J.S.; Culpepper, C.B.; Brown, L.D.; Wesolowski, S.R.; Jonker, S.S.; Davis, M.A.; Limesand, S.W.; Wilkening, R.B.; Hay, W.W.; Rozance, P.J. Chronic anemic hypoxemia attenuates glucose-stimulated insulin secretion in fetal sheep. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2017, 312, R492–R500. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Padilla, E.; Wiltbank, J.N.; Niswender, G.D. Puberty in beef heifers. The interrelationship between pituitary, hypothalamic and ovarian hormones. J. Anim. Sci. 1975, 40, 1091–1104. [Google Scholar] [CrossRef] [PubMed]
- Kappes, S.M.; Warren, W.C.; Pratt, S.L.; Liang, R.; Anthony, R.V. Quantification and cellular localization of ovine placental lactogen messenger ribonucleic acid expression during mid- and late gestation. Endocrinology 1992, 131, 2829–2838. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Blood Flow (Doppler Ultrasound) | |
|---|---|
| Pulsatility index | (PSV–EDV)/Timed-average mean velocity (TAMV) |
| Resistance index | (PSV-EDV)/PSV |
| Umbilical Blood Flow (mL/min) | TAMV × (π/4) × Artery cross-sectional area × 60 |
| Blood Flow (3H2O Tracer) | |
| Rinf 3H2O (dpm/min) | Pump rate × [infusate] |
| Racc(f) (dpm/min) | αpl slope × (0.8 × fetal weight) |
| Racc(m) (dpm/min) | Racc(f) + [αpl slope × 0.8(uterine weight)] |
| Umbilical Blood Flow (UBF; mL/min) | (Rinf − Racc(f))/([3H2O]α(WB) − [3H2O]γ(WB)) |
| Umbilical Plasma Flow (UPF; mL/min) | UBF × [1 − Hctf(avg)] |
| Uterine Blood Flow (UtBF; mL/min) | (Rinf − Racc(m))/([3H2O]V(WB) − [3H2O]A(WB)) |
| Uterine Plasma Flow (UtPF; mL/min) | UtBF × [1 − Hctm(avg)] |
| Nutrient Uptake and Utilization Rates | |
| Umbilical Oxygen Uptake (UOU; mmol/min) | UBF × ([O2]γ(WB) − [O2]α(WB)) |
| Uterine Oxygen Uptake (UtOU; mmol/min) | UtBF × ([O2]A(WB) − [O2]V(WB)) |
| Uteroplacental Oxygen Utilization (mmol/min) | UtOU − UOU |
| Plasma to WB Glucose Conversion | [G]pl × [1 − (0.24 × Hct)] − (3.3 × Hct) |
| Umbilical Glucose Uptake (UGU; µmol/min) | UBF × ([G]γ(WB) − [G]α(WB)) |
| Uterine Glucose Uptake (UtGU; µmol/min) | UtBF × ([G]A(WB) − [G]V(WB)) |
| Uteroplacental Glucose Utilization (µmol/min) | UtGU − UGU |
| Umbilical Lactate Uptake (µmol/min; ULU) | UBF × ([L]γ(pl) − [L]α(pl)) |
| Uterine Lactate Secretion (µmol/min; UtLS) | UtBF × ([L]V(pl) − [L]A(pl)) |
| Uteroplacental Lactate Production (µmol/min) | ULU + UtLS |
| Umbilical AA Uptake (µmol/min; UAAU) | UPF × ([AA]γ(pl) − [AA]α(pl)) |
| Uterine AA Uptake (µmol/min; UtAAU) | UtPF × ([AA]A(pl) − [AA]V(pl)) |
| Umbilical AA Carbon Uptake (µmol/min; UCU) | (#AA carbons) × UAAU |
| Uterine AA Carbon Uptake (µmol/min; UtCU) | (#AA carbons) × UtAAU |
| Umbilical AA Nitrogen Uptake (µmol/min; UNU) | (#AA nitrogens) × UAAU |
| Uterine AA Nitrogen Uptake (µmol/min; (UtNU) | (#AA nitrogens) × UtAAU |
| Fetal Nutrient: Oxygen Quotients | |
| Glucose:Oxygen (G:O) quotient | 6 × ([G]γ(WB) − [G]α(WB))/([O2]γ(WB) − [O2]α(WB)) |
| Lactate:Oxygen (L:O) quotient | 3 × ([L]γ(pl) − [L]α(pl))/([O2]γ(WB) − [O2]α(WB)) |
| Amino Acid:Oxygen (AA:O) quotient | Q × ([AA]γ(pl) − [AA]α(pl))/([O2]γ(WB) − [O2]α(WB)) |
| Total Nutrient:Oxygen quotient | G:O quotient + L:O quotient + Total AA:O quotient |
| CON RNAi | CSH RNAi | % Change | p-Value | |
|---|---|---|---|---|
| 90 dGA Doppler Ultrasound Measurements | (n = 6) | (n = 6) | ||
| Binocular distance, cm | 4.91 ± 0.25 | 4.63 ± 0.19 | 5.70 | 0.48 |
| Biparietal circumference, cm | 16.56 ± 0.63 | 15.11 ± 0.54 | 8.72 | 0.19 |
| Abdominal circumference, cm | 22.03 ± 0.95 | 20.30 ± 1.27 | 7.82 | 0.40 |
| Femur length, cm | 4.24 ± 0.08 | 4.17 ± 0.09 | 1.81 | 0.62 |
| Tibia length, cm | 3.05 ± 0.11 | 3.03 ± 0.04 | 0.75 | 0.87 |
| Pulsatility Index | 1.98 ± 0.16 | 2.04 ± 0.14 | 2.88 | 0.84 |
| Resistance Index | 0.69 ± 0.04 | 0.70 ± 0.02 | 1.12 | 0.89 |
| Systolic:Diastolic | 3.63 ± 0.34 | 3.59 ± 0.28 | 0.97 | 0.95 |
| Fetal heart rate, bpm | 190.48 ± 3.38 | 199.81 ± 8.91 | 4.90 | 0.48 |
| Umbilical artery cross-sectional area, cm2 | 0.24 ± 0.02 | 0.18 ± 0.02 | 25.23 | 0.09 |
| Umbilical artery cross-sectional diameter, cm | 0.55 ± 0.02 | 0.47 ± 0.03 | 15.31 | 0.08 |
| 130 dGA Transplacental Diffusion Blood Flow Measurements | (n = 4) | (n = 4) | ||
| Uterine plasma flow (mL/min) | 1417.74 ± 328.48 | 807.99 ± 208.40 | 43.01 | 0.17 |
| Relative uterine blood flow (mL/min/kg fetus) | 500.07 ± 86.44 | 400.98 ± 59.35 | 19.81 | 0.38 |
| Relative uterine plasma flow (mL/min/kg fetus) | 348.20 ± 61.93 | 274.78 ± 41.11 | 21.09 | 0.36 |
| Uterine blood flow/100 g placenta | 429.05 ± 54.03 | 320.80 ± 59.32 | 25.23 | 0.23 |
| Umbilical plasma flow (mL/min) | 490.84 ± 52.03 | 293.18 ± 63.87 | 40.27 | 0.05 |
| Relative umbilical blood flow (mL/min/kg fetus) | 185.35 ± 8.81 | 155.61 ± 11.26 | 16.04 | 0.08 |
| Relative umbilical plasma flow (mL/min/kg fetus) | 123.17 ± 8.27 | 101.08 ± 12.74 | 17.93 | 0.20 |
| Umbilical blood flow/100 g placenta | 163.27 ± 14.51 | 123.84 ± 13.63 | 24.15 | 0.09 |
| Uterine: umbilical blood flow | 2.68 ± 0.37 | 2.58 ± 0.37 | 3.74 | 0.85 |
| Average umbilical arterial hematocrit | 0.34 ± 0.02 | 0.36 ± 0.04 | 6.25 | 0.65 |
| Average uterine arterial hematocrit | 0.30 ± 0.01 | 0.31 ± 0.02 | 2.94 | 0.64 |
| CON RNAi | CSH RNAi | % Change | p-Value | |
|---|---|---|---|---|
| (n = 4) | (n = 4) | |||
| Crown-rump length, cm | 49.98 ± 1.67 | 45.73 ± 2.10 | 8.50 | 0.16 |
| Ponderal index | 3.19 ± 0.15 | 2.86 ± 0.24 | 10.48 | 0.28 |
| Lower leg length, cm | 37.25 ± 1.16 | 32.38 ± 1.15 | 13.09 | 0.02 |
| Brain, g | 47.71 ± 1.21 | 46.05 ± 3.72 | 3.48 | 0.69 |
| Brain: fetal weight | 0.0121 ± 0.000 | 0.0172 ± 0.002 | 42.63 | 0.03 |
| Liver, g | 100.33 ± 9.13 | 65.50 ± 15.80 | 34.71 | 0.10 |
| Brain: liver | 0.45 ± 0.03 | 0.79 ± 0.21 | 73.65 | 0.17 |
| Liver: fetal weight | 0.0267 ± 0.001 | 0.0247 ± 0.004 | 7.64 | 0.62 |
| Left liver lobe, g | 27.19 ± 4.53 | 17.74 ± 4.07 | 34.76 | 0.17 |
| Right liver lobe, g | 75.46 ± 3.24 | 47.88 ± 11.71 | 36.54 | 0.06 |
| Heart, g | 22.48 ± 1.79 | 17.28 ± 2.14 | 23.15 | 0.11 |
| Heart: fetal weight | 0.0056 ± 0.000 | 0.0063 ± 0.001 | 12.29 | 0.23 |
| Left ventricle, g | 9.02 ± 0.81 | 6.68 ± 0.48 | 25.91 | 0.05 |
| Right ventricle, g | 4.99 ± 0.32 | 3.81 ± 0.70 | 23.56 | 0.18 |
| Lungs, g | 123.18 ± 8.76 | 101.61 ± 15.44 | 17.51 | 0.27 |
| Lungs: fetal weight | 0.031 ± 0.001 | 0.0364 ± 0.001 | 17.31 | 0.01 |
| Pancreas, g | 3.25 ± 0.33 | 2.40 ± 0.42 | 26.21 | 0.16 |
| Kidneys, g | 20.05 ± 1.21 | 15.61 ± 2.33 | 22.17 | 0.14 |
| Perirenal adipose tissue (PRAT), g | 13.36 ± 1.96 | 10.58 ± 1.23 | 20.80 | 0.27 |
| Spleen, g | 6.48 ± 0.58 | 5.24 ± 1.24 | 19.24 | 0.40 |
| Adrenal glands, g | 0.32 ± 0.02 | 0.34 ± 0.06 | 4.38 | 0.86 |
| Biceps femoris (BF), g | 28.69 ± 1.22 | 17.97 ± 2.19 | 37.35 | 0.01 |
| Soleus, g | 0.35 ± 0.06 | 0.14 ± 0.04 | 60.28 | 0.02 |
| Flexor digitorum superficialis (FDS), g | 3.08 ± 0.27 | 1.93 ± 0.31 | 37.45 | 0.03 |
| Tibialis anterior (TA), g | 3.8 ± 0.35 | 2.37 ± 0.32 | 37.59 | 0.02 |
| Extensor digitorum longus (EDL), g | 1.01 ± 0.10 | 0.59 ± 0.10 | 42.22 | 0.03 |
| Uteroplacental weight, g | 1829.40 ± 136.67 | 1333.65 ± 207.93 | 27.10 | 0.09 |
| Membrane weight, g | 483.55 ± 32.18 | 470.28 ± 112.73 | 2.75 | 0.91 |
| Total placentome, # | 67.25 ± 4.09 | 71.75 ± 10.09 | 6.69 | 0.69 |
| 130 dGA Nutrient Uptakes | CON RNAi | CSH RNAi | % Change | p-Value |
|---|---|---|---|---|
| (n = 4) | (n = 4) | |||
| Relative umbilical oxygen uptake (mmol/min/kg fetus) | 0.33 ± 0.01 | 0.29 ± 0.02 | 12.25 | 0.07 |
| Relative uterine oxygen uptake (mmol/min/kg uterus) | 3.18 ± 0.54 | 2.96 ± 0.45 | 6.93 | 0.76 |
| Relative uteroplacental oxygen utilization (mmol/min/kg placenta) | 2.78 ± 0.42 | 1.90 ± 0.25 | 31.61 | 0.12 |
| Relative umbilical glucose uptake (μmol/min/kg fetus) | 34.07 ± 2.98 | 24.75 ± 2.70 | 27.36 | 0.06 |
| Glucose transferred per placental weight (μmol/kg/min) | 301.82 ± 41.64 | 199.24 ± 32.59 | 33.99 | 0.10 |
| Relative uterine glucose uptake (μmol/min/kg uterus) | 515.33 ± 85.74 | 461.76 ± 70.98 | 10.39 | 0.65 |
| Relative uteroplacental glucose utilization (μmol/min/kg placenta) | 634.00 ± 59.81 | 472.72 ± 76.49 | 25.44 | 0.15 |
| Umbilical lactate uptake (μmol/min) | 123.16 ± 5.97 | 75.52 ± 16.11 | 38.68 | 0.03 |
| Relative umbilical lactate uptake (μmol/min/kg fetus) | 31.23 ± 1.78 | 26.26 ± 2.37 | 15.91 | 0.14 |
| Uterine lactate secretion (μmol/min) | 134.17 ± 1.78 | −80.41 ± 154.00 | 159.93 | 0.22 |
| Relative uterine lactate secretion (μmol/min/kg uterus) | 301.02 ± 49.60 | −518.48 ± 692.82 | 272.24 | 0.28 |
| Uteroplacental lactate production (μmol/min) | 257.33 ± 16.19 | −4.89 ± 168.14 | 101.90 | 0.17 |
| Relative uteroplacental lactate production (μmol/min/kg placenta) | 576.83 ± 68.83 | −311.58 ± 705.10 | 154.02 | 0.26 |
| 130 dGA Nutrient Uptakes | CON RNAi | CSH RNAi | % Change | p-Value |
|---|---|---|---|---|
| Maternal (Uterine) | (n = 4) | (n = 4) | ||
| Uterine artery insulin, ng/mL | 0.92 ± 0.17 | 0.77 ± 0.10 | 15.92 | 0.49 |
| Uterine artery IGF1, ng/mL | 297.91 ± 45.18 | 260.24 ± 25.74 | 12.64 | 0.50 |
| Uterine artery cortisol, ng/mL | 126.53 ± 12.95 | 96.27 ± 29.07 | 23.91 | 0.38 |
| Uterine vein estradiol, pg/mL | 7.32 ± 2.34 | 4.42 ± 1.81 | 39.71 | 0.36 |
| Fetal (Umbilical) | (n = 4) | (n = 4) | ||
| Umbilical artery insulin, ng/mL | 1.12 ± 0.15 | 0.69 ± 0.23 | 38.63 | 0.16 |
| Umbilical artery IGF1, ng/mL | 140.89 ± 9.30 | 72.85 ± 14.94 | 48.29 | 0.01 |
| Umbilical artery cortisol, ng/mL | 11.98 ± 4.14 | 58.62 ± 29.83 | 389.25 | 0.17 |
| Umbilical vein estradiol, pg/mL | 5.59 ± 1.99 | 6.59 ± 0.75 | 17.99 | 0.65 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tanner, A.R.; Lynch, C.S.; Kennedy, V.C.; Ali, A.; Winger, Q.A.; Rozance, P.J.; Anthony, R.V. CSH RNA Interference Reduces Global Nutrient Uptake and Umbilical Blood Flow Resulting in Intrauterine Growth Restriction. Int. J. Mol. Sci. 2021, 22, 8150. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22158150
Tanner AR, Lynch CS, Kennedy VC, Ali A, Winger QA, Rozance PJ, Anthony RV. CSH RNA Interference Reduces Global Nutrient Uptake and Umbilical Blood Flow Resulting in Intrauterine Growth Restriction. International Journal of Molecular Sciences. 2021; 22(15):8150. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22158150
Chicago/Turabian StyleTanner, Amelia R., Cameron S. Lynch, Victoria C. Kennedy, Asghar Ali, Quinton A. Winger, Paul J. Rozance, and Russell V. Anthony. 2021. "CSH RNA Interference Reduces Global Nutrient Uptake and Umbilical Blood Flow Resulting in Intrauterine Growth Restriction" International Journal of Molecular Sciences 22, no. 15: 8150. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22158150