
Maintenance of Ligament Homeostasis of Spheroid-Colonized Embroidered and Functionalized Scaffolds after 3D Stretch

 ,
,  , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
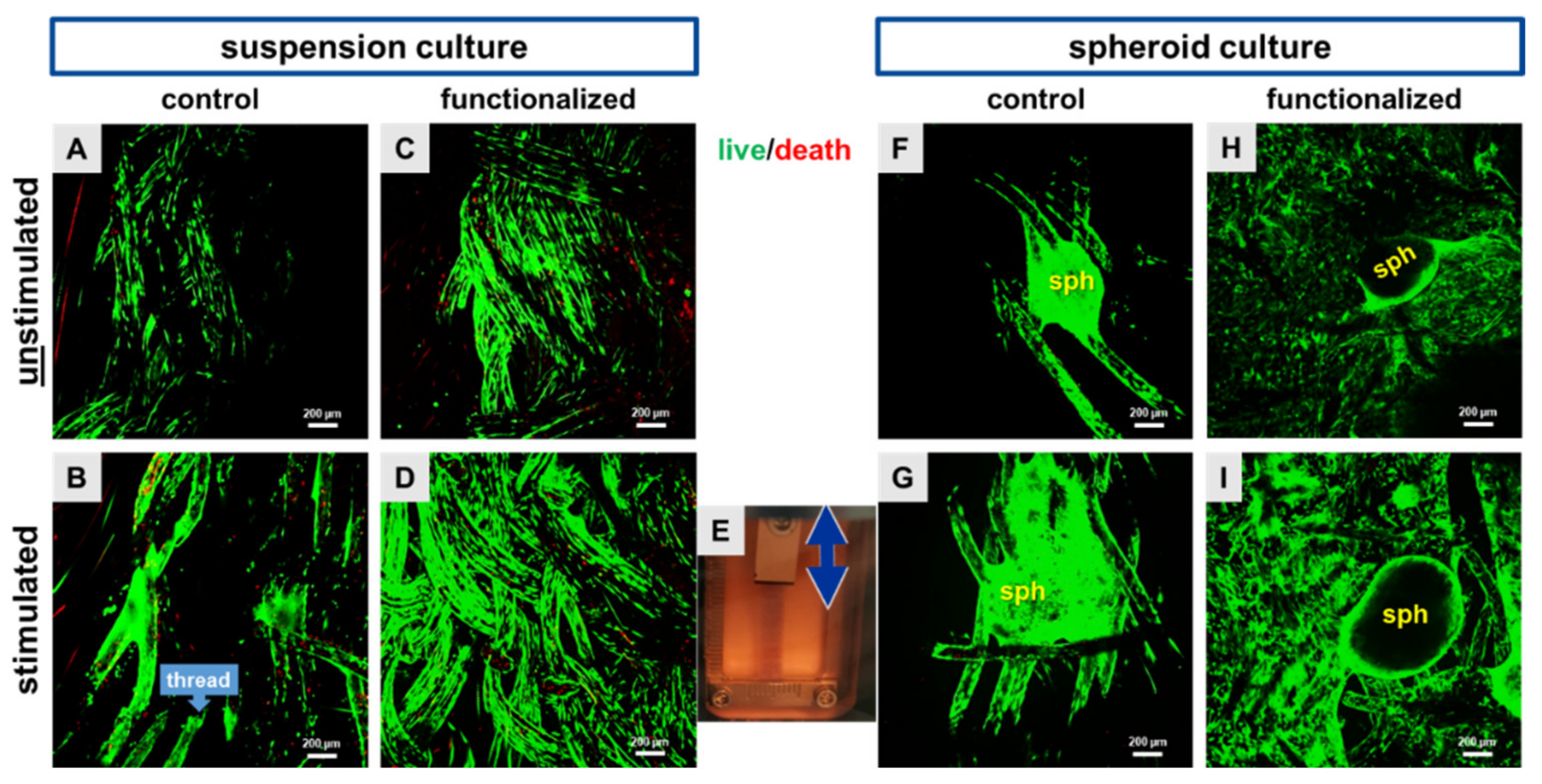
2.1. Cyclic Stretch Induced a Homogenous Cell Distribution
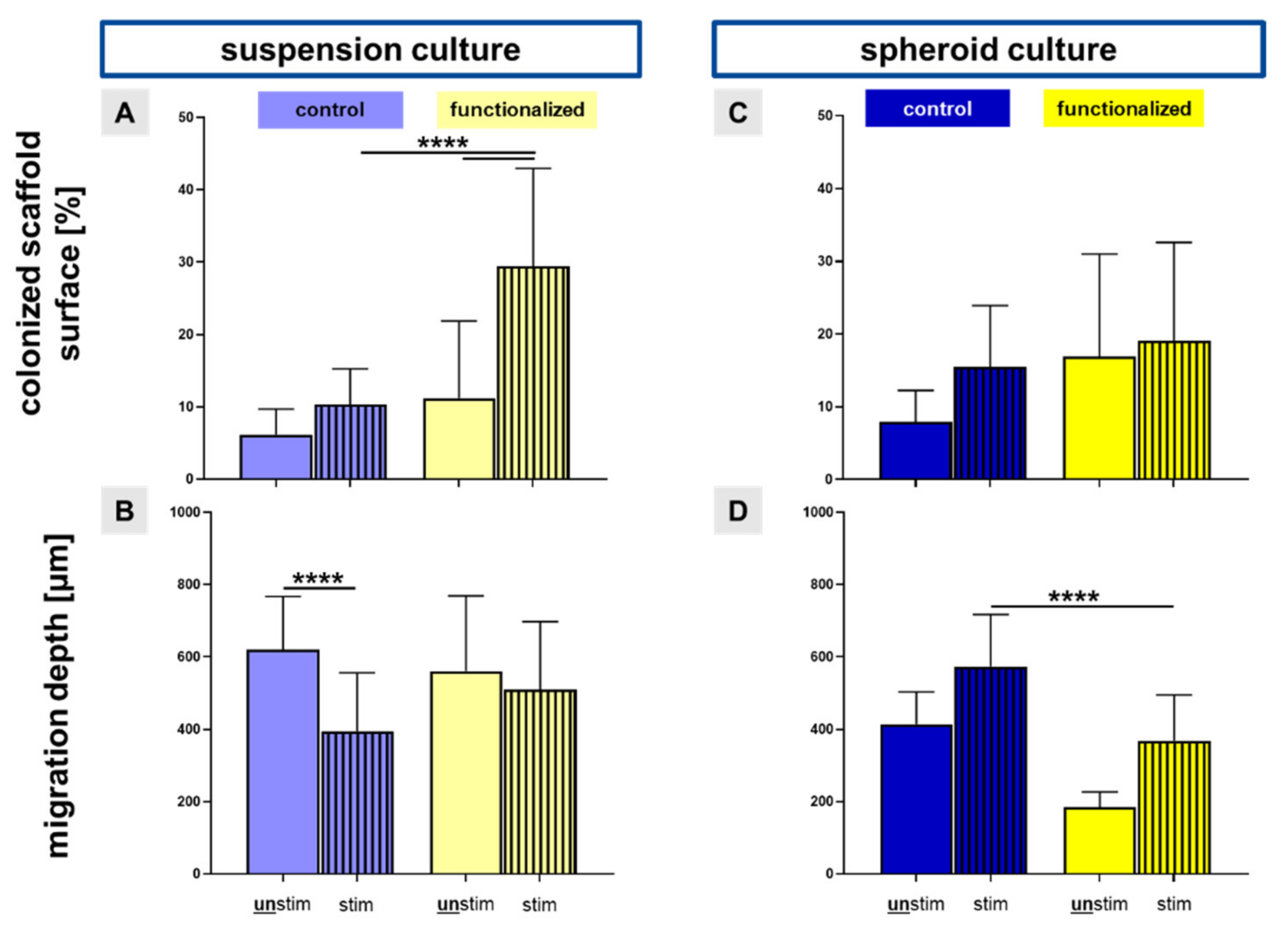
2.2. Migration into Inner Parts of the Scaffold
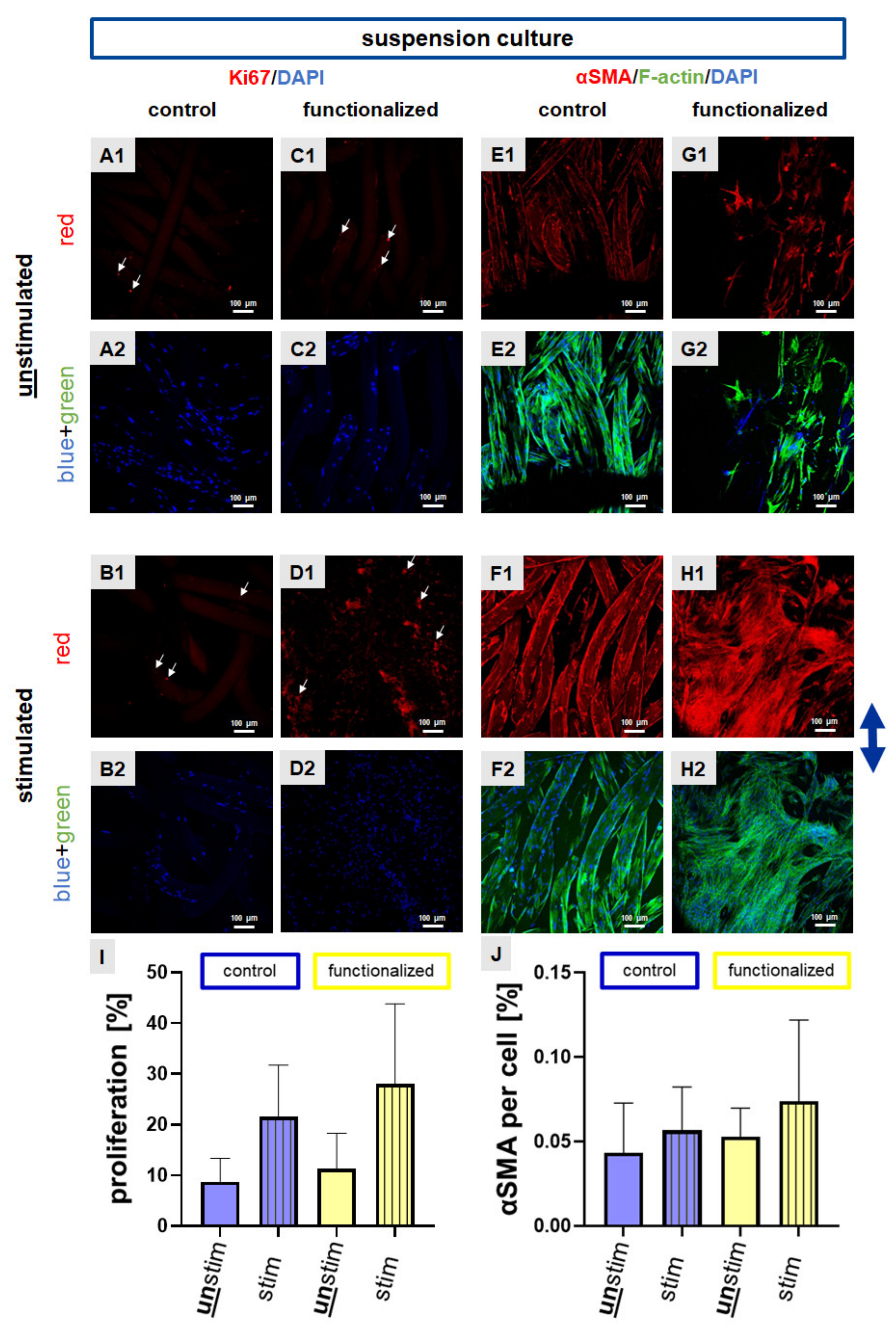
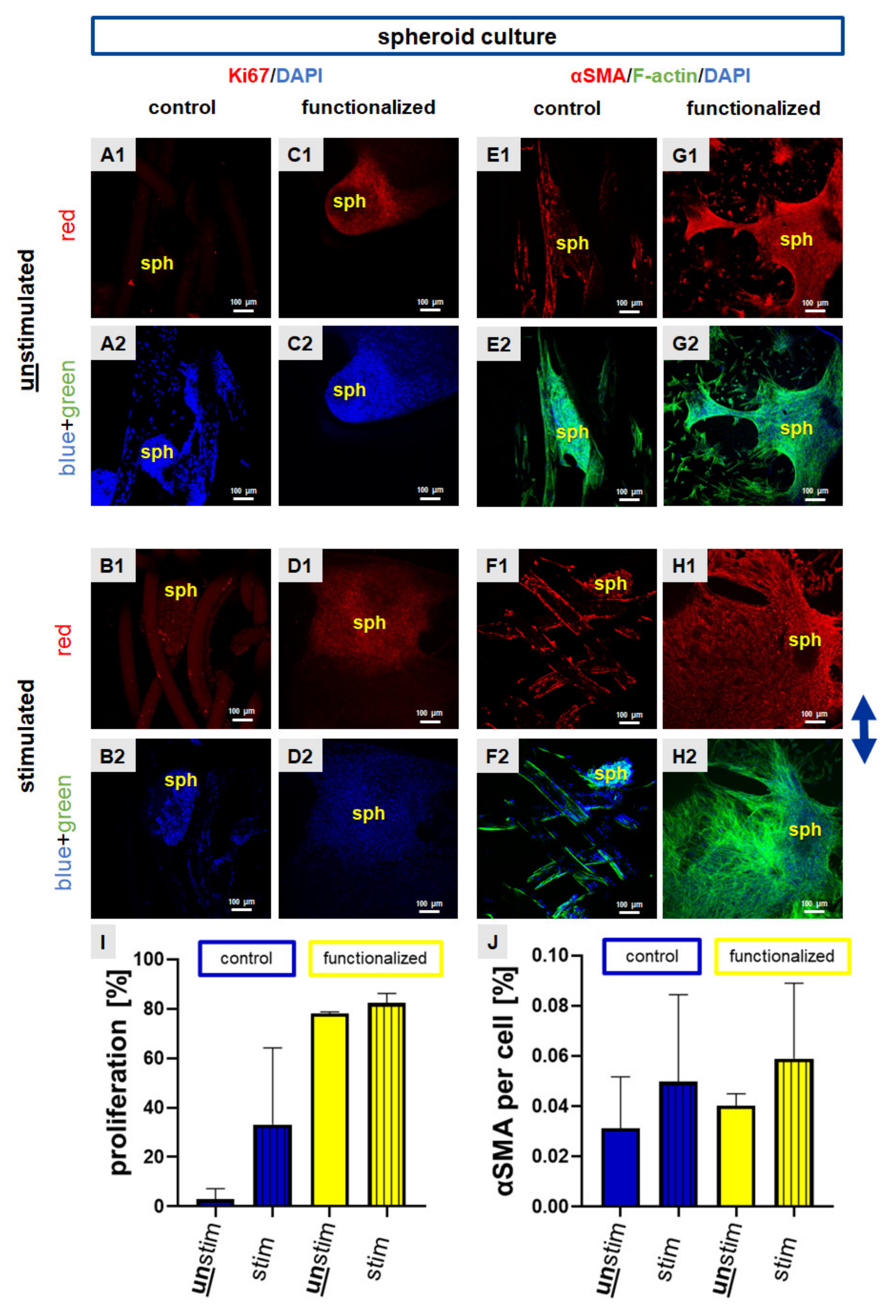
2.3. Cyclic Strain Influenced Proliferation and Cytoskeletal Protein Expression
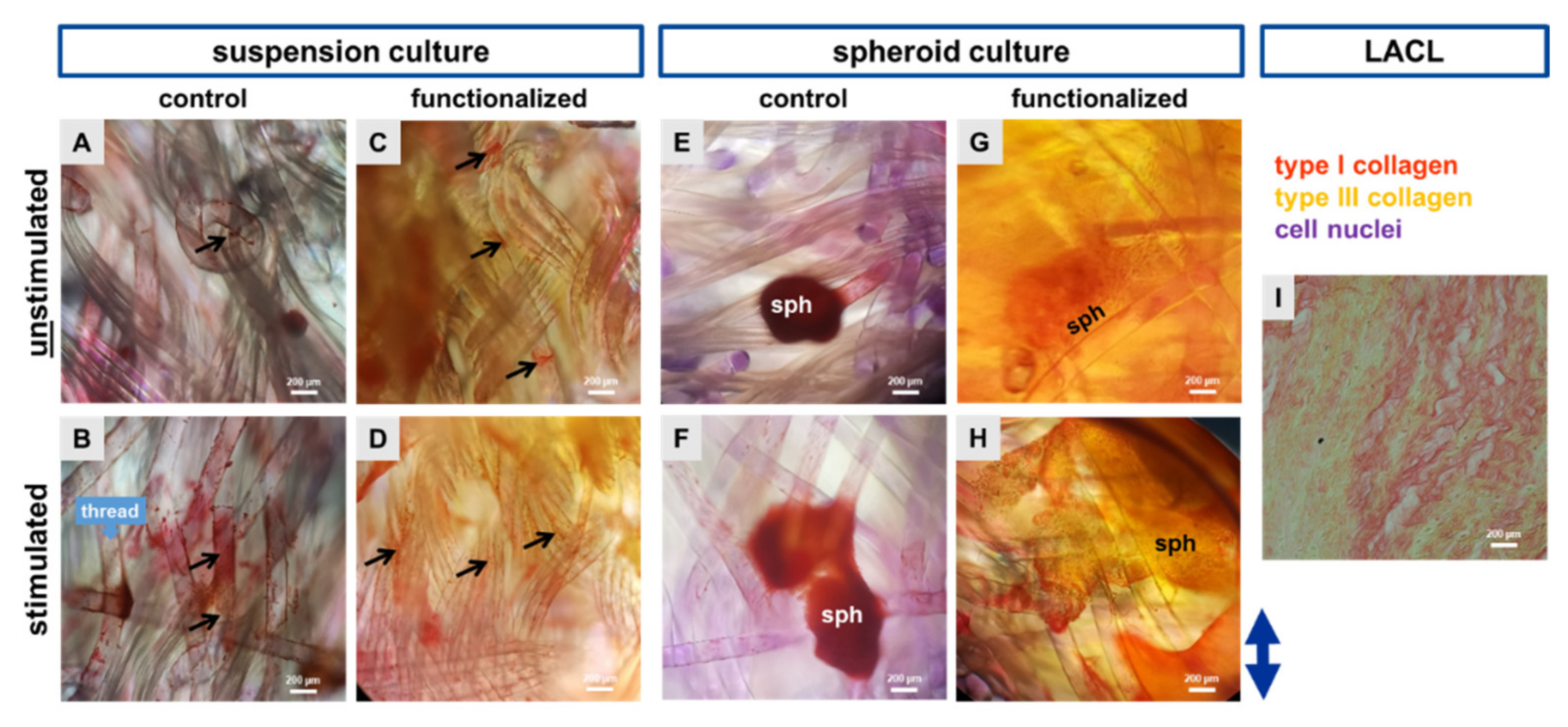
2.4. Cell Morphology Shown by Histological Staining Using Sirius Red
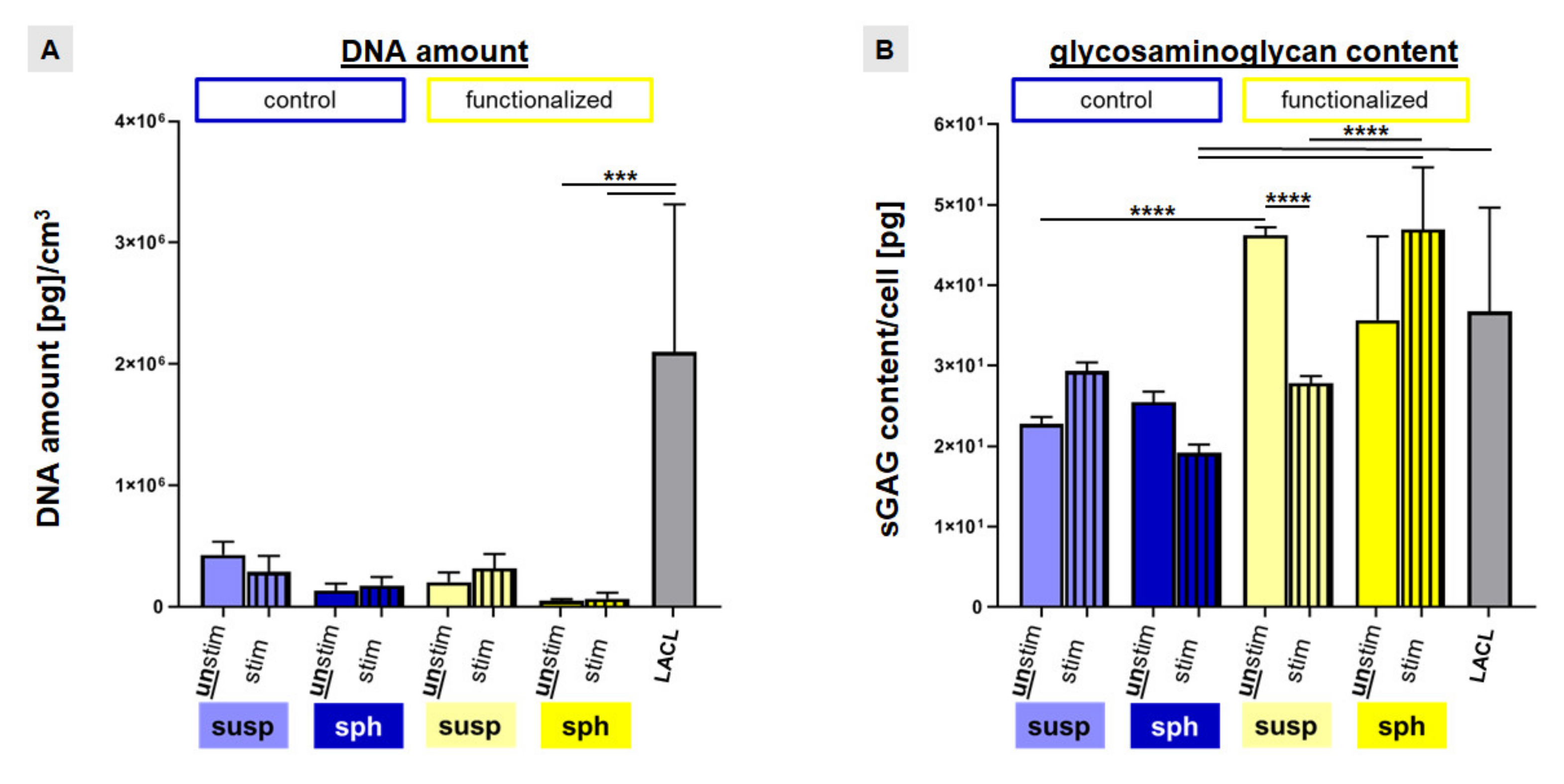
2.5. Enhanced Cell Amount and sGAG Content by Strain
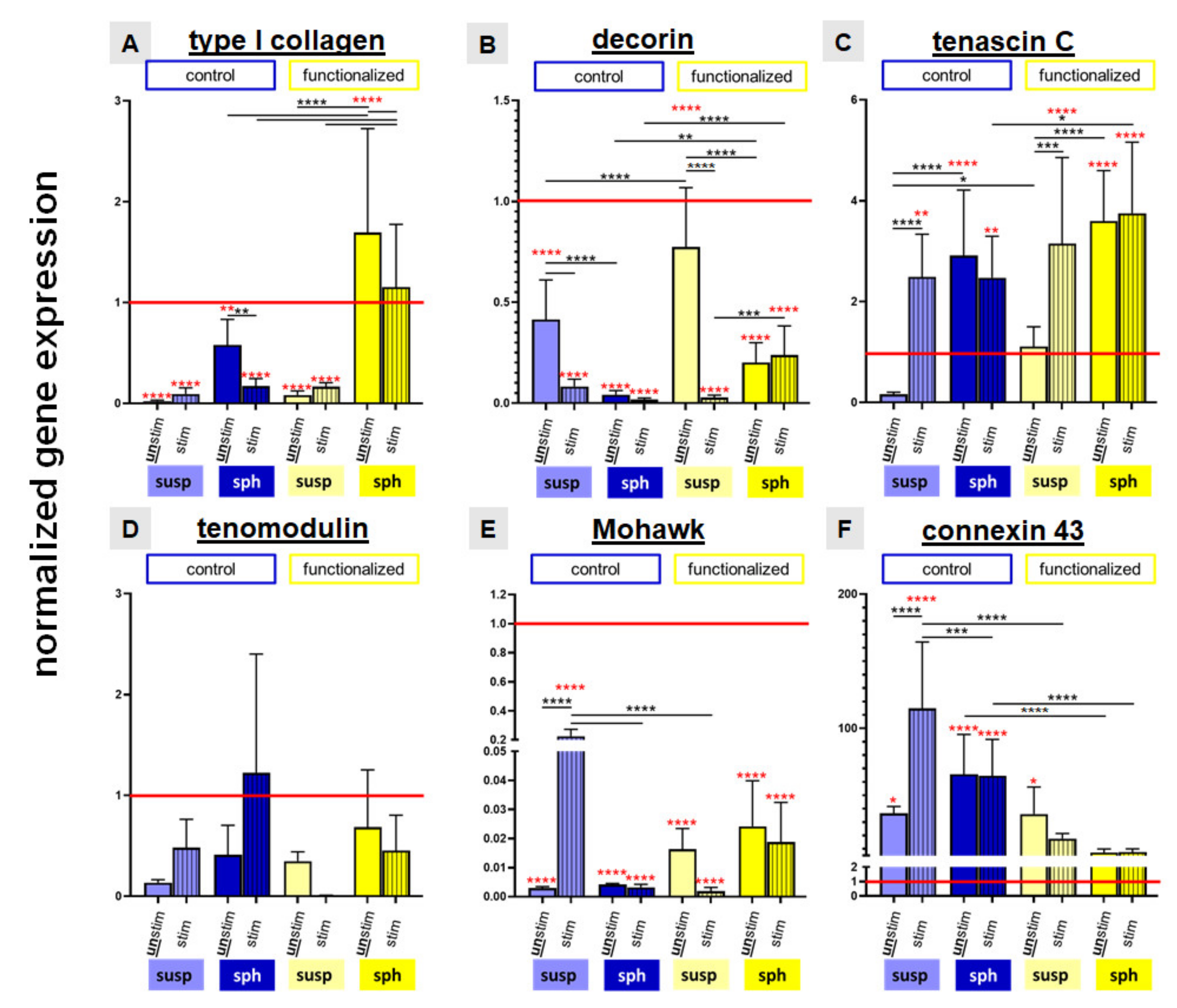
2.6. Relative Gene Expression of Ligament Associated Genes and Connexin 43
3. Discussion
4. Materials and Methods
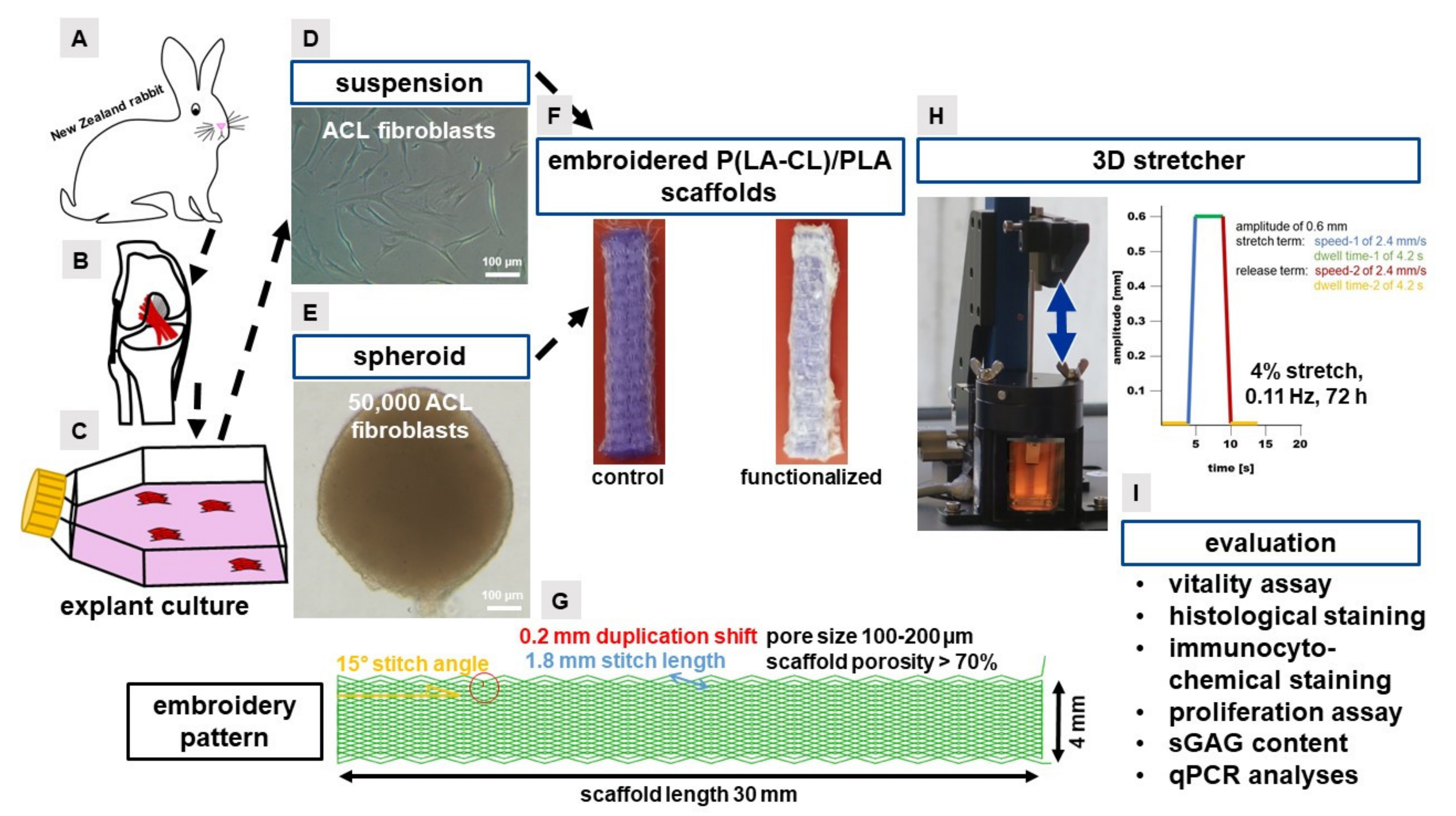
4.1. Preparation of the Embroidered P(LA-CL)/PLA Scaffolds
4.2. Functionalization of the Scaffolds
4.3. Isolation of Fibroblasts from Lapine ACLs
4.4. Scaffold Colonization
4.5. Cell Suspension-Based Scaffold Colonization
4.6. Spheroid-Based Scaffold Colonization
4.7. Three-Dimensional (3D) Mechanostimulator and Cyclic Stretching
4.8. Cell Survival
4.9. Immunocytochemical Staining and Measurement of the Migration Depth of Cells into the Scaffold
4.10. Histological Staining of Sirius Red
4.11. Quantitative Assays for DNA- and sGAG Quantification
4.12. RNA Isolation
4.13. Quantitative Real-Time PCR
4.14. Image Analyses
4.15. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Petersen, W.; Tillmann, B. Structure and vascularization of the cruciate ligaments of the human knee joint. Anat. Embryol. 1999, 200, 325–334. [Google Scholar] [CrossRef] [PubMed]
- Duthon, V.B.; Barea, C.; Abrassart, S.; Fasel, J.H.; Fritschy, D.; Menetrey, J. Anatomy of the anterior cruciate ligament. Knee Surg. Sports Traumatol. Arthrosc. 2006, 14, 204–213. [Google Scholar] [CrossRef] [Green Version]
- Amiel, D.; Nagineni, C.N.; Choi, S.H.; Lee, J. Intrinsic properties of ACL and MCL cells and their responses to growth factors. Med. Sci. Sports Exerc. 1995, 27, 844–851. [Google Scholar] [CrossRef]
- Irvine, G.B.; Glasgow, M.M. The natural history of the meniscus in anterior cruciate insufficiency. Arthroscopic analysis. J. Bone Jt. Surg. Br. 1992, 74, 403–405. [Google Scholar] [CrossRef] [PubMed]
- Beynnon, B.D.; Johnson, R.J.; Abate, J.A.; Fleming, B.C.; Nichols, C.E. Treatment of anterior cruciate ligament injuries, part I. Am. J. Sports Med. 2005, 33, 1579–1602. [Google Scholar] [CrossRef] [PubMed]
- Beynnon, B.D.; Johnson, R.J.; Abate, J.A.; Fleming, B.C.; Nichols, C.E. Treatment of anterior cruciate ligament injuries, part 2. Am. J. Sports Med. 2005, 33, 1751–1767. [Google Scholar] [CrossRef] [PubMed]
- Murawski, C.D.; van Eck, C.F.; Irrgang, J.J.; Tashman, S.; Fu, F.H. Operative treatment of primary anterior cruciate ligament rupture in adults. J. Bone Jt. Surg. Am. 2014, 96, 685–694. [Google Scholar] [CrossRef] [PubMed]
- Kartus, J.; Movin, T.; Karlsson, J. Donor-site morbidity and anterior knee problems after anterior cruciate ligament reconstruction using autografts. Arthroscopy 2001, 17, 971–980. [Google Scholar] [CrossRef]
- Ge, Z.; Yang, F.; Goh, J.C.; Ramakrishna, S.; Lee, E.H. Biomaterials and scaffolds for ligament tissue engineering. J. Biomed. Mater. Res. A. 2006, 77, 639–652. [Google Scholar] [CrossRef]
- Leong, N.L.; Petrigliano, F.A.; McAllister, D.R. Current tissue engineering strategies in anterior cruciate ligament reconstruction. J. Biomed. Mater. Res. A 2014, 102, 1614–1624. [Google Scholar] [CrossRef]
- Kuo, C.K.; Marturano, J.E.; Tuan, R.S. Novel strategies in tendon and ligament tissue engineering: Advanced biomaterials and regeneration motifs. Sports Med. Arthrosc. Rehabil. Ther. Technol. 2010, 2, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nau, T.; Teuschl, A. Regeneration of the anterior cruciate ligament: Current strategies in tissue engineering. World J. Orthop. 2015, 6, 127–136. [Google Scholar] [CrossRef] [Green Version]
- Hahn, J.; Breier, A.; Brünig, H.; Heinrich, G. Long-term hydrolytic degradation study on polymer-based embroidered scaffolds for ligament tissue engineering. J. Ind. Text. 2018, 47, 1305–1320. [Google Scholar] [CrossRef]
- Hahn, J.; Schulze-Tanzil, G.; Schroepfer, M.; Meyer, M.; Goegele, C.; Hoyer, M.; Spickenheuer, A.; Heinrich, G.; Breier, A. Viscoelastic behavior of embroidered scaffolds for acl tissue engineering made of pla and p(la-cl) after in vitro degradation. Int. J. Mol. Sci. 2019, 20, 4655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goegele, C.; Hahn, J.; Elschner, C.; Breier, A.; Schroepfer, M.; Prade, I.; Meyer, M.; Schulze-Tanzil, G. Enhanced Growth of Lapine Anterior Cruciate Ligament-Derived Fibroblasts on Scaffolds Embroidered from Poly(l-lactide-co-epsilon-caprolactone) and Polylactic Acid Threads Functionalized by Fluorination and Hexamethylene Diisocyanate Cross-Linked Collagen Foams. Int J. Mol. Sci. 2020, 21, 1132. [Google Scholar] [CrossRef] [Green Version]
- Hoyer, M.; Drechsel, N.; Meyer, M.; Meier, C.; Hinuber, C.; Breier, A.; Hahner, J.; Heinrich, G.; Rentsch, C.; Garbe, L.A.; et al. Embroidered polymer-collagen hybrid scaffold variants for ligament tissue engineering. Mater. Sci. Eng. C Mater. Biol. Appl. 2014, 43, 290–299. [Google Scholar] [CrossRef] [PubMed]
- Hoyer, M.; Meier, C.; Breier, A.; Hahner, J.; Heinrich, G.; Drechsel, N.; Meyer, M.; Rentsch, C.; Garbe, L.A.; Ertel, W.; et al. In vitro characterization of self-assembled anterior cruciate ligament cell spheroids for ligament tissue engineering. Histochem. Cell Biol. 2015, 143, 289–300. [Google Scholar] [CrossRef]
- Schulze-Tanzil, G.; Arnold, P.; Goegele, C.; Hahn, J.; Breier, A.; Meyer, M.; Kohl, B.; Schroepfer, M.; Schwarz, S. SV40 Transfected human anterior cruciate ligament derived ligamentocytes-suitable as a human in vitro model for ligament reconstruction? Int. J. Mol. Sci. 2020, 21, 593. [Google Scholar] [CrossRef] [Green Version]
- Schroepfer, M.; Junghans, F.; Voigt, D.; Meyer, M.; Breier, A.; Schulze-Tanzil, G.; Prade, I. Gas-Phase Fluorination on pla improves cell adhesion and spreading. ACS Omega 2020, 5, 5498–5507. [Google Scholar] [CrossRef]
- Varma, S.; Orgel, J.P.; Schieber, J.D. Nanomechanics of Type I Collagen. Biophys. J. 2016, 111, 50–56. [Google Scholar] [CrossRef] [Green Version]
- Kalson, N.S.; Holmes, D.F.; Kapacee, Z.; Otermin, I.; Lu, Y.; Ennos, R.A.; Canty-Laird, E.G.; Kadler, K.E. An experimental model for studying the biomechanics of embryonic tendon: Evidence that the development of mechanical properties depends on the actinomyosin machinery. Matrix Biol. 2010, 29, 678–689. [Google Scholar] [CrossRef]
- Yang, G.; Rothrauff, B.B.; Tuan, R.S. Tendon and ligament regeneration and repair: Clinical relevance and developmental paradigm. Birth Defects Res. C Embryo Today 2013, 99, 203–222. [Google Scholar] [CrossRef] [Green Version]
- Govoni, M.; Muscari, C.; Lovecchio, J.; Guarnieri, C.; Giordano, E. Mechanical Actuation systems for the phenotype commitment of stem cell-based tendon and ligament tissue substitutes. Stem Cell Rev. Rep. 2016, 12, 189–201. [Google Scholar] [CrossRef] [PubMed]
- Desrosiers, E.A.; Methot, S.; Yahia, L.; Rivard, C.H. Responses of ligamentous fibroblasts to mechanical stimulation. Ann. Chir. 1995, 49, 768–774. [Google Scholar] [PubMed]
- Young, R.G.; Butler, D.L.; Weber, W.; Caplan, A.I.; Gordon, S.L.; Fink, D.J. Use of mesenchymal stem cells in a collagen matrix for Achilles tendon repair. J. Orthop. Res. 1998, 16, 406–413. [Google Scholar] [CrossRef]
- Kim, S.G.; Akaike, T.; Sasagaw, T.; Atomi, Y.; Kurosawa, H. Gene expression of type I and type III collagen by mechanical stretch in anterior cruciate ligament cells. Cell Struct. Funct. 2002, 27, 139–144. [Google Scholar] [CrossRef] [Green Version]
- Song, F.; Jiang, D.; Wang, T.; Wang, Y.; Chen, F.; Xu, G.; Kang, Y.; Zhang, Y. Mechanical loading improves tendon-bone healing in a rabbit anterior cruciate ligament reconstruction model by promoting proliferation and matrix formation of mesenchymal stem cells and tendon cells. Cell. Physiol. Biochem. 2017, 41, 875–889. [Google Scholar] [CrossRef]
- Henshaw, D.R.; Attia, E.; Bhargava, M.; Hannafin, J.A. Canine ACL fibroblast integrin expression and cell alignment in response to cyclic tensile strain in three-dimensional collagen gels. J. Orthop. Res. 2006, 24, 481–490. [Google Scholar] [CrossRef] [PubMed]
- Lim, W.L.; Liau, L.L.; Ng, M.H.; Chowdhury, S.R.; Law, J.X. Current Progress in Tendon and Ligament Tissue Engineering. Tissue Eng. Regen. Med. 2019, 16, 549–571. [Google Scholar] [CrossRef]
- Xu, B.; Ju, Y.; Song, G. Mechanical stretch-induced f-actin reorganization and tenogenic differentiation of human mesenchymal stem cells. J. Biol. Res. 2012, 18, 218. [Google Scholar]
- Kreja, L.; Liedert, A.; Schlenker, H.; Brenner, R.E.; Fiedler, J.; Friemert, B.; Durselen, L.; Ignatius, A. Effects of mechanical strain on human mesenchymal stem cells and ligament fibroblasts in a textured poly(l-lactide) scaffold for ligament tissue engineering. J. Mater. Sci. Mater. Med. 2012, 23, 2575–2582. [Google Scholar] [CrossRef] [PubMed]
- Morita, Y.; Watanabe, S.; Ju, Y.; Xu, B. Determination of optimal cyclic uniaxial stretches for stem cell-to-tenocyte differentiation under a wide range of mechanical stretch conditions by evaluating gene expression and protein synthesis levels. Acta. Bioeng. Biomech. 2013, 15, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Nam, H.Y.; Pingguan-Murphy, B.; Amir Abbas, A.; Mahmood Merican, A.; Kamarul, T. The proliferation and tenogenic differentiation potential of bone marrow-derived mesenchymal stromal cell are influenced by specific uniaxial cyclic tensile loading conditions. Biomech. Model. Mechanobiol. 2015, 14, 649–663. [Google Scholar] [CrossRef] [PubMed]
- Beynnon, B.D.; Fleming, B.C. Anterior cruciate ligament strain in-vivo: A review of previous work. J. Biomech. 1998, 31, 519–525. [Google Scholar] [CrossRef]
- Taylor, K.A.; Cutcliffe, H.C.; Queen, R.M.; Utturkar, G.M.; Spritzer, C.E.; Garrett, W.E.; DeFrate, L.E. In vivo measurement of ACL length and relative strain during walking. J. Biomech. 2013, 46, 478–483. [Google Scholar] [CrossRef] [Green Version]
- Hahner, J.; Hoyer, M.; Hillig, S.; Schulze-Tanzil, G.; Meyer, M.; Schroepfer, M.; Lohan, A.; Garbe, L.A.; Heinrich, G.; Breier, A. Diffusion chamber system for testing of collagen-based cell migration barriers for separation of ligament enthesis zones in tissue-engineered ACL constructs. J. Biomater. Sci. Polym. Ed. 2015, 26, 1085–1099. [Google Scholar] [CrossRef]
- Yao, L.; Bestwick, C.S.; Bestwick, L.A.; Maffulli, N.; Aspden, R.M. Phenotypic drift in human tenocyte culture. Tissue Eng. 2006, 12, 1843–1849. [Google Scholar] [CrossRef]
- Stoll, C.; John, T.; Endres, M.; Rosen, C.; Kaps, C.; Kohl, B.; Sittinger, M.; Ertel, W.; Schulze-Tanzil, G. Extracellular matrix expression of human tenocytes in three-dimensional air-liquid and PLGA cultures compared with tendon tissue: Implications for tendon tissue engineering. J. Orthop. Res. 2010, 28, 1170–1177. [Google Scholar] [CrossRef]
- Theiss, F.; Mirsaidi, A.; Mhanna, R.; Kummerle, J.; Glanz, S.; Bahrenberg, G.; Tiaden, A.N.; Richards, P.J. Use of biomimetic microtissue spheroids and specific growth factor supplementation to improve tenocyte differentiation and adaptation to a collagen-based scaffold in vitro. Biomaterials 2015, 69, 99–109. [Google Scholar] [CrossRef]
- Schwarz, S.; Goegele, C.; Ondruschka, B.; Hammer, N.; Kohl, B.; Schulze-Tanzil, G. Migrating Myofibroblastic Iliotibial Band-Derived Fibroblasts Represent a Promising Cell Source for Ligament Reconstruction. Int. J. Mol. Sci. 2019, 20, 1972. [Google Scholar] [CrossRef] [Green Version]
- Freeman, J.W.; Woods, M.D.; Laurencin, C.T. Tissue engineering of the anterior cruciate ligament using a braid-twist scaffold design. J. Biomech. 2007, 40, 2029–2036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toyoda, T.; Matsumoto, H.; Fujikawa, K.; Saito, S.; Inoue, K. Tensile load and the metabolism of anterior cruciate ligament cells. Clin. Orthop. Relat. Res. 1998, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Felsenthal, N.; Zelzer, E. Mechanical regulation of musculoskeletal system development. Development 2017, 144, 4271–4283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asou, Y.; Nifuji, A.; Tsuji, K.; Shinomiya, K.; Olson, E.N.; Koopman, P.; Noda, M. Coordinated expression of scleraxis and Sox9 genes during embryonic development of tendons and cartilage. J. Orthop. Res. 2002, 20, 827–833. [Google Scholar] [CrossRef]
- Hahner, J.; Hinüber, C.; Breier, A.; Siebert, T.; Brünig, H.; Heinrich, G. Adjusting the mechanical behavior of embroidered scaffolds to lapin anterior cruciate ligaments by varying the thread materials. Text. Res. J. 2015, 85, 1431–1444. [Google Scholar] [CrossRef] [Green Version]
- Dursun, G.; Tohidnezhad, M.; Markert, B.; Stoffel, M. Effects of uniaxial stretching on tenocyte migration behaviour. Curr. Dir. Biomed. Eng. 2018, 4, 313. [Google Scholar] [CrossRef]
- Griffin, L.Y.; Albohm, M.J.; Arendt, E.A.; Bahr, R.; Beynnon, B.D.; Demaio, M.; Dick, R.W.; Engebretsen, L.; Garrett, W.E., Jr.; Hannafin, J.A.; et al. Understanding and preventing noncontact anterior cruciate ligament injuries: A review of the Hunt Valley II meeting, January 2005. Am. J. Sports Med. 2006, 34, 1512–1532. [Google Scholar] [CrossRef]
- Backman, L.J.; Fong, G.; Andersson, G.; Scott, A.; Danielson, P. Substance P is a mechanoresponsive, autocrine regulator of human tenocyte proliferation. PLoS ONE 2011, 6, e27209. [Google Scholar] [CrossRef]
- Gögele, C.; Hoffmann, C.; Konrad, J.; Merkel, R.; Schwarz, S.; Tohidnezhad, M.; Hoffmann, B.; Schulze-Tanzil, G.G. Correction to: Cyclically stretched ACL fibroblasts emigrating from spheroids adapt their cytoskeleton and ligament-related expression profile. Cell Tissue Res. 2021, 384, 789. [Google Scholar] [CrossRef]
- Scholzen, T.; Gerdes, J. The Ki-67 protein: From the known and the unknown. J. Cell. Physiol 2000, 182, 311–322. [Google Scholar] [CrossRef]
- Hinz, B.; Celetta, G.; Tomasek, J.J.; Gabbiani, G.; Chaponnier, C. Alpha-smooth muscle actin expression upregulates fibroblast contractile activity. Mol. Biol. Cell 2001, 12, 2730–2741. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Han, X.; Meng, Y.; Gao, L.; Guo, Y.; Jing, Y.; Bai, D. Favorable effect of myofibroblasts on collagen synthesis and osteocalcin production in the periodontal ligament. Am. J. Orthod. Dentofacial. Orthop. 2014, 145, 469–479. [Google Scholar] [CrossRef]
- Lee, P.Y.; Liu, Y.C.; Wang, M.X.; Hu, J.J. Fibroblast-seeded collagen gels in response to dynamic equibiaxial mechanical stimuli: A biomechanical study. J. Biomech. 2018, 78, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Tomasek, J.J.; Gabbiani, G.; Hinz, B.; Chaponnier, C.; Brown, R.A. Myofibroblasts and mechano-regulation of connective tissue remodelling. Nat. Rev. Mol. Cell Biol. 2002, 3, 349–363. [Google Scholar] [CrossRef] [PubMed]
- Kollmannsberger, P.; Bidan, C.M.; Dunlop, J.W.C.; Fratzl, P.; Vogel, V. Tensile forces drive a reversible fibroblast-to-myofibroblast transition during tissue growth in engineered clefts. Sci. Adv. 2018, 4, eaao4881. [Google Scholar] [CrossRef] [Green Version]
- Granato, G.; Ruocco, M.R.; Iaccarino, A.; Masone, S.; Cali, G.; Avagliano, A.; Russo, V.; Bellevicine, C.; Di Spigna, G.; Fiume, G.; et al. Generation and analysis of spheroids from human primary skin myofibroblasts: An experimental system to study myofibroblasts deactivation. Cell Death Discov. 2017, 3, 17038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, Q.; Gladson, C.L.; Wu, H.; Hayasaka, H.; Olman, M.A. Focal adhesion kinase (FAK)-related non-kinase inhibits myofibroblast differentiation through differential MAPK activation in a FAK-dependent manner. J. Biol. Chem. 2008, 283, 26839–26849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.J.; Shie, M.Y.; Hung, C.J.; Wu, B.C.; Liu, S.L.; Huang, T.H.; Kao, C.T. Activation of focal adhesion kinase induces extracellular signal-regulated kinase-mediated osteogenesis in tensile force-subjected periodontal ligament fibroblasts but not in osteoblasts. J. Bone Miner. Metab. 2014, 32, 671–682. [Google Scholar] [CrossRef] [PubMed]
- Gayan, S.; Teli, A.; Dey, T. Inherent aggressive character of invasive and non-invasive cells dictates the in vitro migration pattern of multicellular spheroid. Sci. Rep. 2017, 7, 11527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim-Kaneyama, J.R.; Suzuki, W.; Ichikawa, K.; Ohki, T.; Kohno, Y.; Sata, M.; Nose, K.; Shibanuma, M. Uni-axial stretching regulates intracellular localization of Hic-5 expressed in smooth-muscle cells in vivo. J. Cell Sci. 2005, 118, 937–949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulze-Tanzil, G.; Mobasheri, A.; Clegg, P.D.; Sendzik, J.; John, T.; Shakibaei, M. Cultivation of human tenocytes in high-density culture. Histochem. Cell Biol. 2004, 122, 219–228. [Google Scholar] [CrossRef]
- Liu, S.H.; Yang, R.S.; al-Shaikh, R.; Lane, J.M. Collagen in tendon, ligament, and bone healing. A current review. Clin. Orthop. Relat. Res. 1995, 318, 265–278. [Google Scholar]
- Kraus, A.; Luetzenberg, R.; Abuagela, N.; Hollenberg, S.; Infanger, M. Spheroid formation and modulation of tenocyte-specific gene expression under simulated microgravity. Muscles Ligaments Tendons J. 2017, 7, 411–417. [Google Scholar] [CrossRef] [Green Version]
- Beachley, V.; Kasyanov, V.; Nagy-Mehesz, A.; Norris, R.; Ozolanta, I.; Kalejs, M.; Stradins, P.; Baptista, L.; da Silva, K.; Grainjero, J.; et al. The fusion of tissue spheroids attached to pre-stretched electrospun polyurethane scaffolds. J. Tissue Eng. 2014, 5, 2041731414556561. [Google Scholar] [CrossRef] [PubMed]
- Sawaguchi, N.; Majima, T.; Funakoshi, T.; Shimode, K.; Harada, K.; Minami, A.; Nishimura, S. Effect of cyclic three-dimensional strain on cell proliferation and collagen synthesis of fibroblast-seeded chitosan-hyaluronan hybrid polymer fiber. J. Orthop. Sci. 2010, 15, 569–577. [Google Scholar] [CrossRef] [PubMed]
- Burk, J.; Plenge, A.; Brehm, W.; Heller, S.; Pfeiffer, B.; Kasper, C. Induction of Tenogenic Differentiation Mediated by Extracellular Tendon Matrix and Short-Term Cyclic Stretching. Stem Cells Int. 2016, 2016, 7342379. [Google Scholar] [CrossRef] [Green Version]
- Qin, T.W.; Sun, Y.L.; Thoreson, A.R.; Steinmann, S.P.; Amadio, P.C.; An, K.N.; Zhao, C. Effect of mechanical stimulation on bone marrow stromal cell-seeded tendon slice constructs: A potential engineered tendon patch for rotator cuff repair. Biomaterials 2015, 51, 43–50. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Thien, C.; Wang, C.; Ni, M.; Gao, J.; Wang, A.; Jiang, Q.; Tuan, R.S.; Zheng, Q.; Zheng, M.H. 3D uniaxial mechanical stimulation induces tenogenic differentiation of tendon-derived stem cells through a PI3K/AKT signaling pathway. FASEB J. 2018, 32, 4804–4814. [Google Scholar] [CrossRef] [Green Version]
- Qin, T.W.; Chen, Q.; Sun, Y.L.; Steinmann, S.P.; Amadio, P.C.; An, K.N.; Zhao, C. Mechanical characteristics of native tendon slices for tissue engineering scaffold. J. Biomed. Mater. Res. B Appl. Biomater. 2012, 100, 752–758. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.L.; Yin, Z.; Shen, W.L.; Chen, X.; Heng, B.C.; Zou, X.H.; Ouyang, H.W. Efficacy of hESC-MSCs in knitted silk-collagen scaffold for tendon tissue engineering and their roles. Biomaterials 2010, 31, 9438–9451. [Google Scholar] [CrossRef]
- Husse, B.; Briest, W.; Homagk, L.; Isenberg, G.; Gekle, M. Cyclical mechanical stretch modulates expression of collagen I and collagen III by PKC and tyrosine kinase in cardiac fibroblasts. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 293, R1898–R1907. [Google Scholar] [CrossRef]
- Robinson, K.A.; Sun, M.; Barnum, C.E.; Weiss, S.N.; Huegel, J.; Shetye, S.S.; Lin, L.; Saez, D.; Adams, S.M.; Iozzo, R.V.; et al. Decorin and biglycan are necessary for maintaining collagen fibril structure, fiber realignment, and mechanical properties of mature tendons. Matrix Biol. 2017, 64, 81–93. [Google Scholar] [CrossRef]
- Youngstrom, D.W.; Rajpar, I.; Kaplan, D.L.; Barrett, J.G. A bioreactor system for in vitro tendon differentiation and tendon tissue engineering. J. Orthop. Res. 2015, 33, 911–918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Govoni, M.; Berardi, A.C.; Muscari, C.; Campardelli, R.; Bonafe, F.; Guarnieri, C.; Reverchon, E.; Giordano, E.; Maffulli, N.; Della Porta, G. An Engineered multiphase three-dimensional microenvironment to ensure the controlled delivery of cyclic strain and human growth differentiation factor 5 for the tenogenic commitment of human bone marrow mesenchymal stem cells. Tissue Eng. Part A 2017, 23, 811–822. [Google Scholar] [CrossRef] [PubMed]
- Järvinen, T.A.; Jozsa, L.; Kannus, P.; Jarvinen, T.L.; Hurme, T.; Kvist, M.; Pelto-Huikko, M.; Kalimo, H.; Järvinen, M. Mechanical loading regulates the expression of tenaSci.n-C in the myotendinous junction and tendon but does not induce de novo synthesis in the skeletal muscle. J. Cell Sci. 2003, 116, 857–866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, D.; Alberton, P.; Caceres, M.D.; Volkmer, E.; Schieker, M.; Docheva, D. Tenomodulin is essential for prevention of adipocyte accumulation and fibrovascular scar formation during early tendon healing. Cell Death Dis. 2017, 8, e3116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, H.; Caceres, M.D.; Yan, Z.; Schieker, M.; Nerlich, M.; Docheva, D. Tenomodulin regulates matrix remodeling of mouse tendon stem/progenitor cells in an ex vivo collagen I gel model. Biochem. Biophys. Res. Commun. 2019, 512, 691–697. [Google Scholar] [CrossRef]
- Xu, Y.; Wang, Q.; Li, Y.; Gan, Y.; Li, P.; Li, S.; Zhou, Y.; Zhou, Q. Cyclic tensile strain induces tenogenic differentiation of tendon-derived stem cells in bioreactor culture. Biomed. Res. Int. 2015, 2015, 790804. [Google Scholar] [CrossRef] [Green Version]
- Sawadkar, P.; Player, D.; Bozec, L.; Mudera, V. The mechanobiology of tendon fibroblasts under static and uniaxial cyclic load in a 3D tissue engineered model mimicking native extracellular matrix. J. Tissue Eng. Regen. Med. 2020, 14, 135–146. [Google Scholar] [CrossRef]
- Suzuki, H.; Ito, Y.; Shinohara, M.; Yamashita, S.; Ichinose, S.; Kishida, A.; Oyaizu, T.; Kayama, T.; Nakamichi, R.; Koda, N.; et al. Gene targeting of the transcription factor Mohawk in rats causes heterotopic ossification of Achilles tendon via failed tenogenesis. Proc. Natl. Acad. Sci. USA 2016, 113, 7840–7845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kayama, T.; Mori, M.; Ito, Y.; Matsushima, T.; Nakamichi, R.; Suzuki, H.; Ichinose, S.; Saito, M.; Marumo, K.; Asahara, H. Gtf2ird1-dependent mohawk expression regulates mechanosensing properties of the tendon. Mol. Cell Biol. 2016, 36, 1297–1309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, F.; Zhang, A.; Richardson, D.W. Regulation of the tenogenic gene expression in equine tenocyte-derived induced pluripotent stem cells by mechanical loading and Mohawk. Stem Cell Res. 2019, 39, 101489. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.S.; Lu, X.H.; Chen, D.Y.; Yuan, W.; Yang, L.L.; Chen, Y.; He, H.L. Mechanical strain induces Cx43 expression in spinal ligament fibroblasts derived from patients presenting ossification of the posterior longitudinal ligament. Eur. Spine J. 2011, 20, 1459–1465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waggett, A.D.; Benjamin, M.; Ralphs, J.R. Connexin 32 and 43 gap junctions differentially modulate tenocyte response to cyclic mechanical load. Eur. J. Cell Biol. 2006, 85, 1145–1154. [Google Scholar] [CrossRef]
- Theodossiou, S.K.; Tokle, J.; Schiele, N.R. TGFbeta2-induced tenogenesis impacts cadherin and connexin cell-cell junction proteins in mesenchymal stem cells. Biochem. Biophys. Res. Commun. 2019, 508, 889–893. [Google Scholar] [CrossRef]
- Xu, C.; Fan, Z.; Shan, W.; Hao, Y.; Ma, J.; Huang, Q.; Zhang, F. Cyclic stretch influenced expression of membrane connexin 43 in human periodontal ligament cell. Arch. Oral. Biol. 2012, 57, 1602–1608. [Google Scholar] [CrossRef]
- Wall, M.E.; Otey, C.; Qi, J.; Banes, A.J. Connexin 43 is localized with actin in tenocytes. Cell Motil. Cytoskelet. 2007, 64, 121–130. [Google Scholar] [CrossRef]
- Maeda, E.; Ohashi, T. Mechano-regulation of gap junction communications between tendon cells is dependent on the magnitude of tensile strain. Biochem. Biophys. Res. Commun. 2015, 465, 281–286. [Google Scholar] [CrossRef]
- Wang, J.; Li, G.; Li, Y.; Zhao, Y.; Manthari, R.K.; Wang, J. The effects of fluoride on the gap-junctional intercellular communication of rats. Osteoblast. Biol. Trace Elem. Res. 2020, 193, 195–203. [Google Scholar] [CrossRef]
- Breier, A.C. Biomedical textiles for orthopaedic and surgical applications: Embroidery technology for hard-tissue scaffolds. In eBook; Blair, T., Ed.; Woodhead Publishing: Sawston, Cambridge, UK, 2015; pp. 1–212. [Google Scholar]
- Schefe, J.H.; Lehmann, K.E.; Buschmann, I.R.; Unger, T.; Funke-Kaiser, H. Quantitative real-time RT-PCR data analysis: Current concepts and the novel gene expression’s CT difference formula. J. Mol. Med. 2006, 84, 901–910. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Primary Antibody | Dilution | Secondary Antibody | Dilution |
|---|---|---|---|---|
| Ki67 | mouse-anti-human, Merck | 1:30 | donkey-anti-mouse Cy3, Invitrogen | 1:200 |
| αSMA | mouse-anti-human, Sigma Aldrich | 1:50 | donkey-anti-mouse Cy3, Invitrogen | 1:200 |
| Gene Symbol | Species | Gene Name | Amplicon Length | Assay ID |
|---|---|---|---|---|
| COL1A1 | O. cuniculus | type I collagen, alpha 1 | 70 | Oc03396073_g1 |
| CXN43 | O. cuniculus | connexin 43 | 68 | Oc03396056_g1 |
| DCN | Homo sapiens | decorin | 77 | Hs00370384_m1 |
| GAPDH (LOC100009074) | O. cuniculus | glyceraldehyde-3-phosphate dehydrogenase | 82 | Oc03823402_g1 |
| MKX | O. cuniculus | Mohawk | 60 | Oc06754037_m1 |
| TNC | O. cuniculus | tenascin C | 61 | Oc06726696_m1 |
| TNMD (LOC100125994) | O. cuniculus | myodulin | 146 | Oc03399505_m1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gögele, C.; Konrad, J.; Hahn, J.; Breier, A.; Schröpfer, M.; Meyer, M.; Merkel, R.; Hoffmann, B.; Schulze-Tanzil, G. Maintenance of Ligament Homeostasis of Spheroid-Colonized Embroidered and Functionalized Scaffolds after 3D Stretch. Int. J. Mol. Sci. 2021, 22, 8204. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22158204
Gögele C, Konrad J, Hahn J, Breier A, Schröpfer M, Meyer M, Merkel R, Hoffmann B, Schulze-Tanzil G. Maintenance of Ligament Homeostasis of Spheroid-Colonized Embroidered and Functionalized Scaffolds after 3D Stretch. International Journal of Molecular Sciences. 2021; 22(15):8204. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22158204
Chicago/Turabian StyleGögele, Clemens, Jens Konrad, Judith Hahn, Annette Breier, Michaela Schröpfer, Michael Meyer, Rudolf Merkel, Bernd Hoffmann, and Gundula Schulze-Tanzil. 2021. "Maintenance of Ligament Homeostasis of Spheroid-Colonized Embroidered and Functionalized Scaffolds after 3D Stretch" International Journal of Molecular Sciences 22, no. 15: 8204. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22158204