
Heme Oxygenase-1 Induction by Cobalt Protoporphyrin Ameliorates Cholestatic Liver Disease in a Xenobiotic-Induced Murine Model


Abstract
:1. Introduction
2. Results
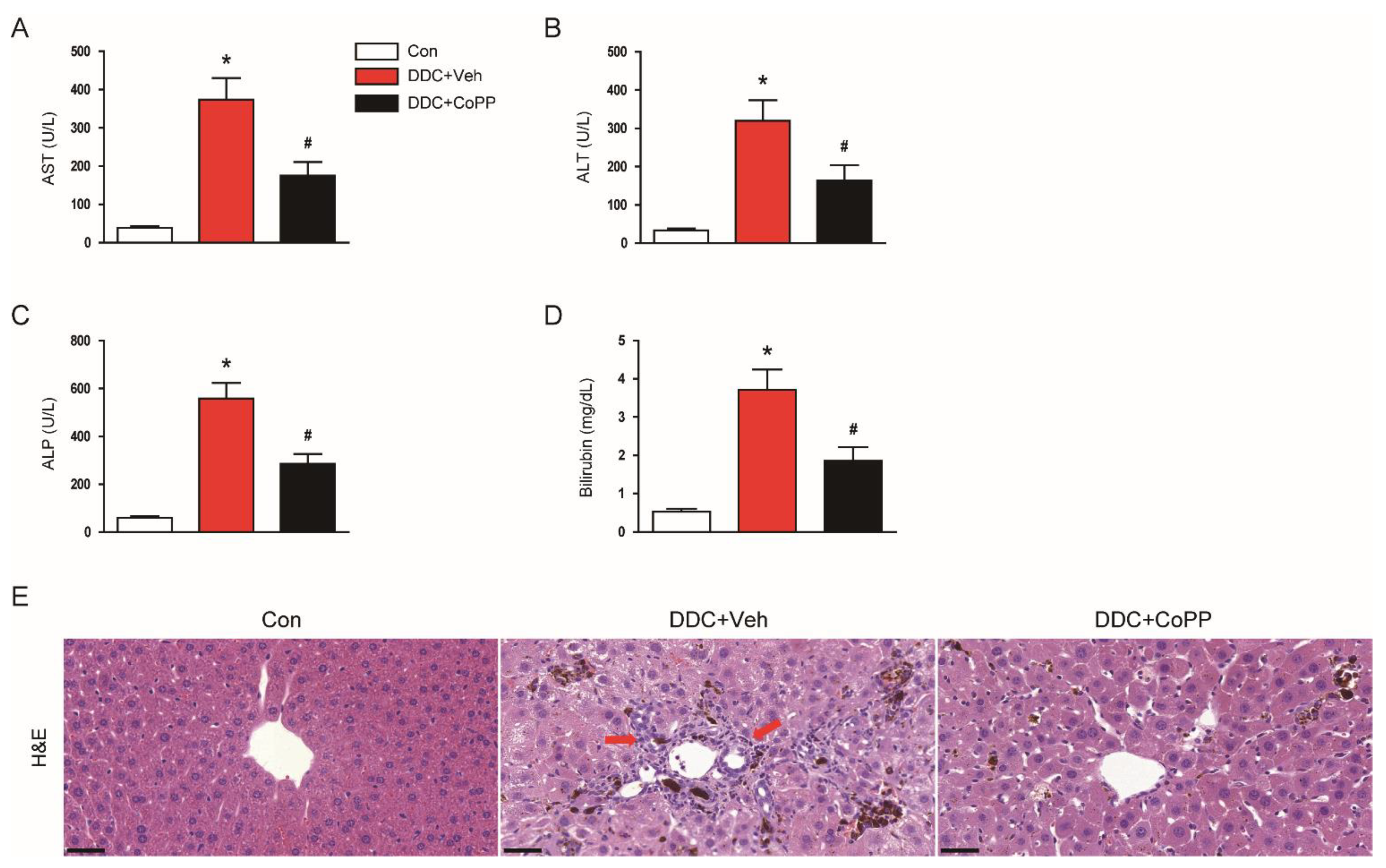
2.1. Administration of CoPP Ameliorated Liver Damage and Cholestasis in DDC-Fed Mice
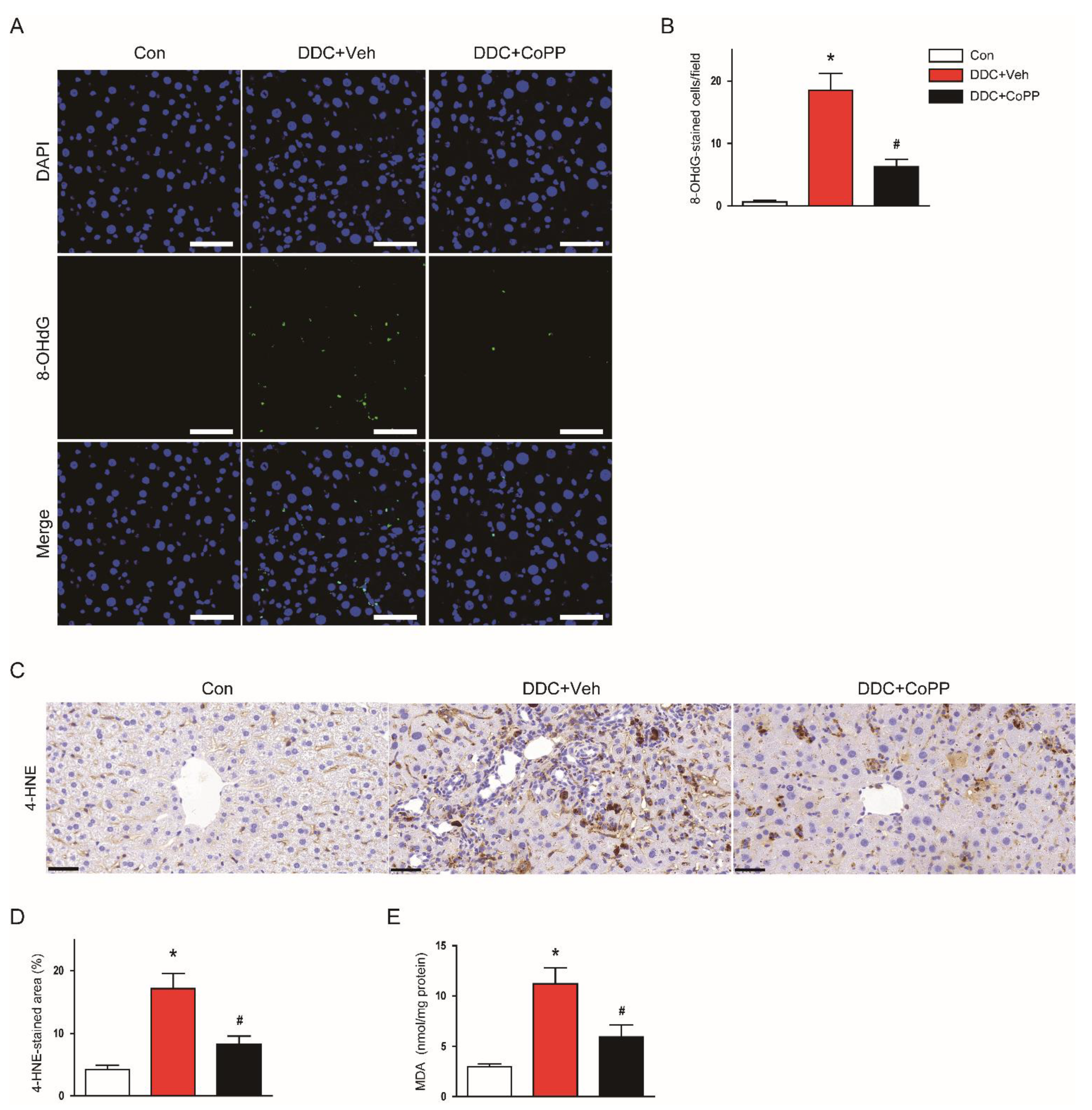
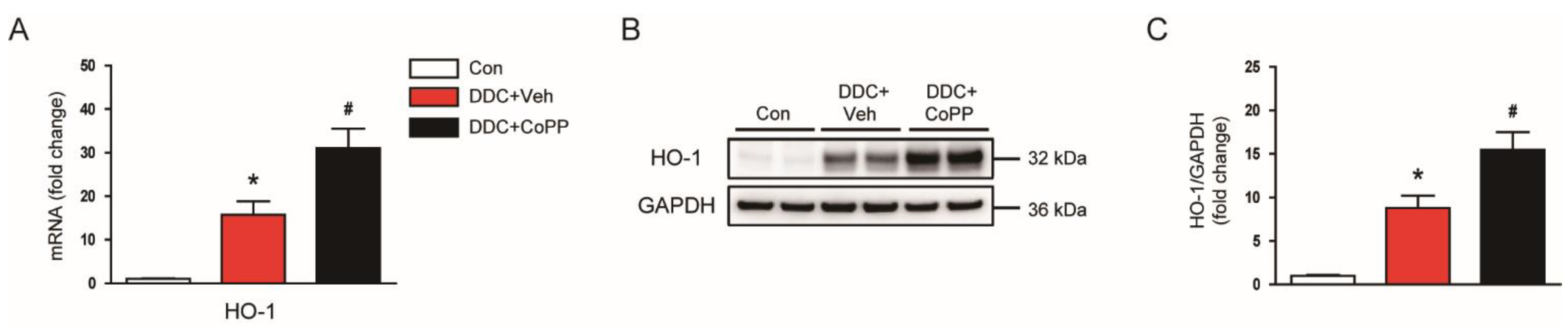
2.2. Induction of HO-1 by CoPP Attenuated Oxidative Stress in DDC-Fed Mice
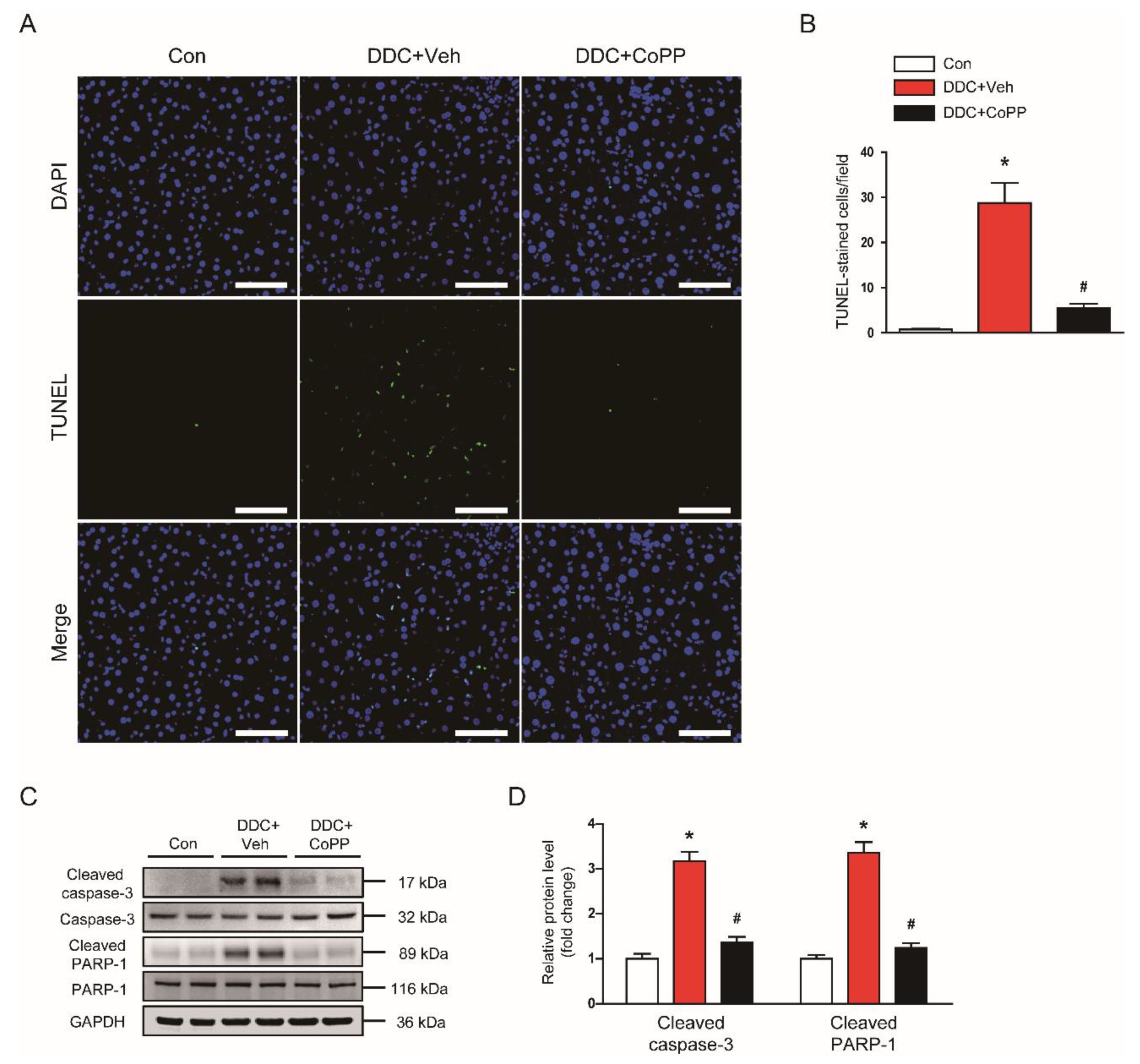
2.3. Induction of HO-1 by CoPP Inhibited Apoptotic Cell Death in DDC-Fed Mice
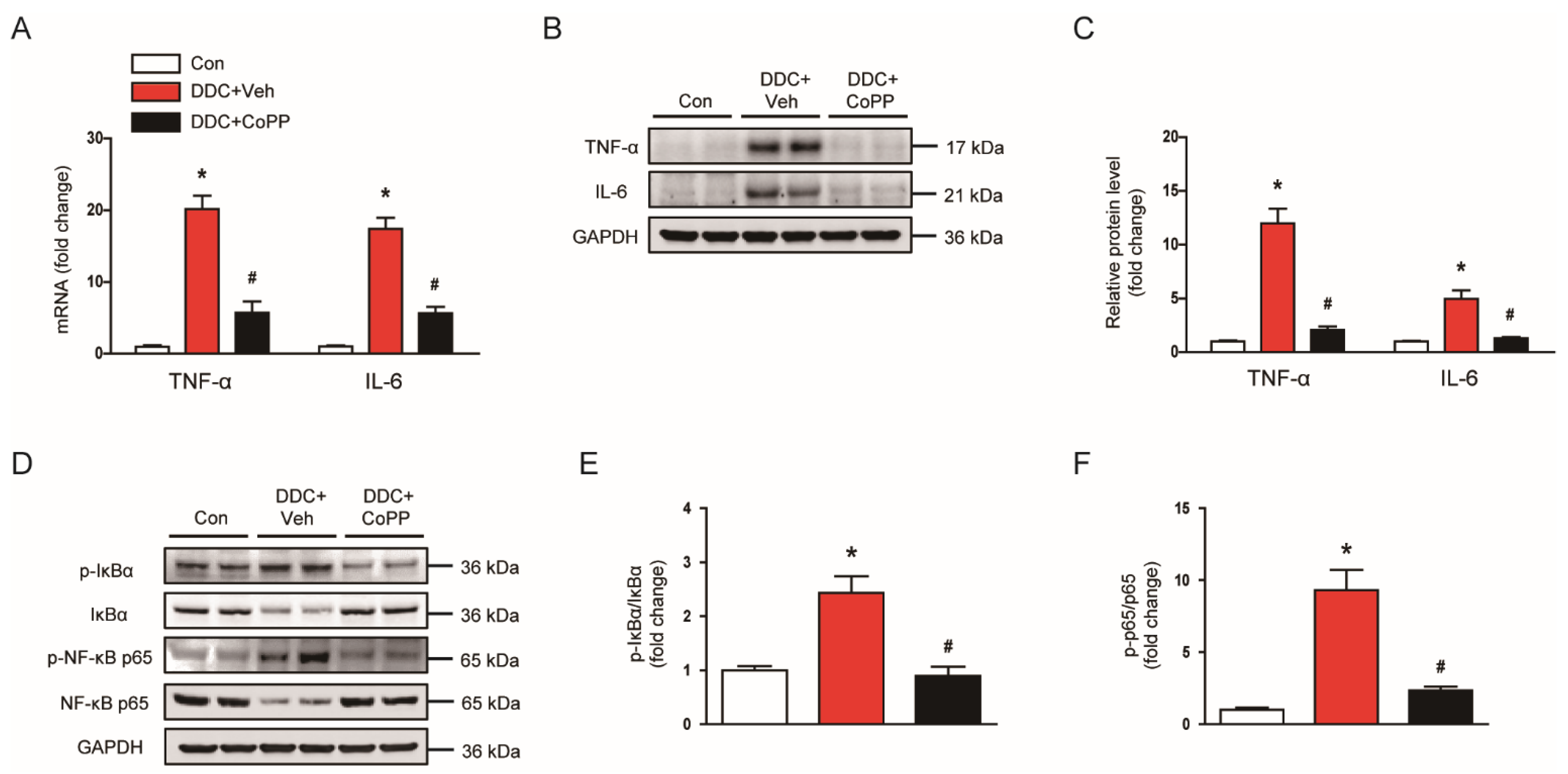
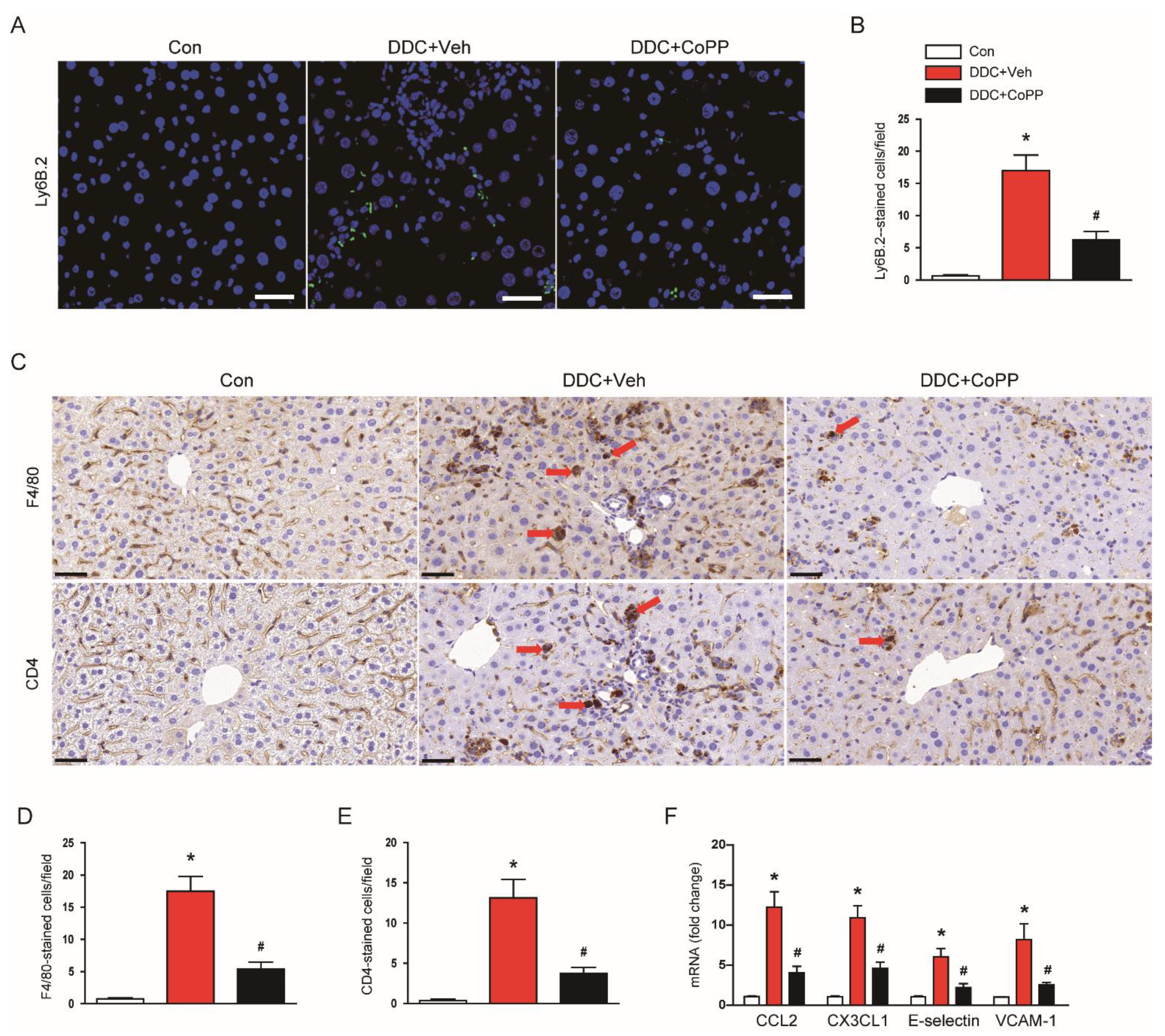
2.4. Induction of HO-1 by CoPP Suppressed Inflammatory Responses in DDC-Fed Mice
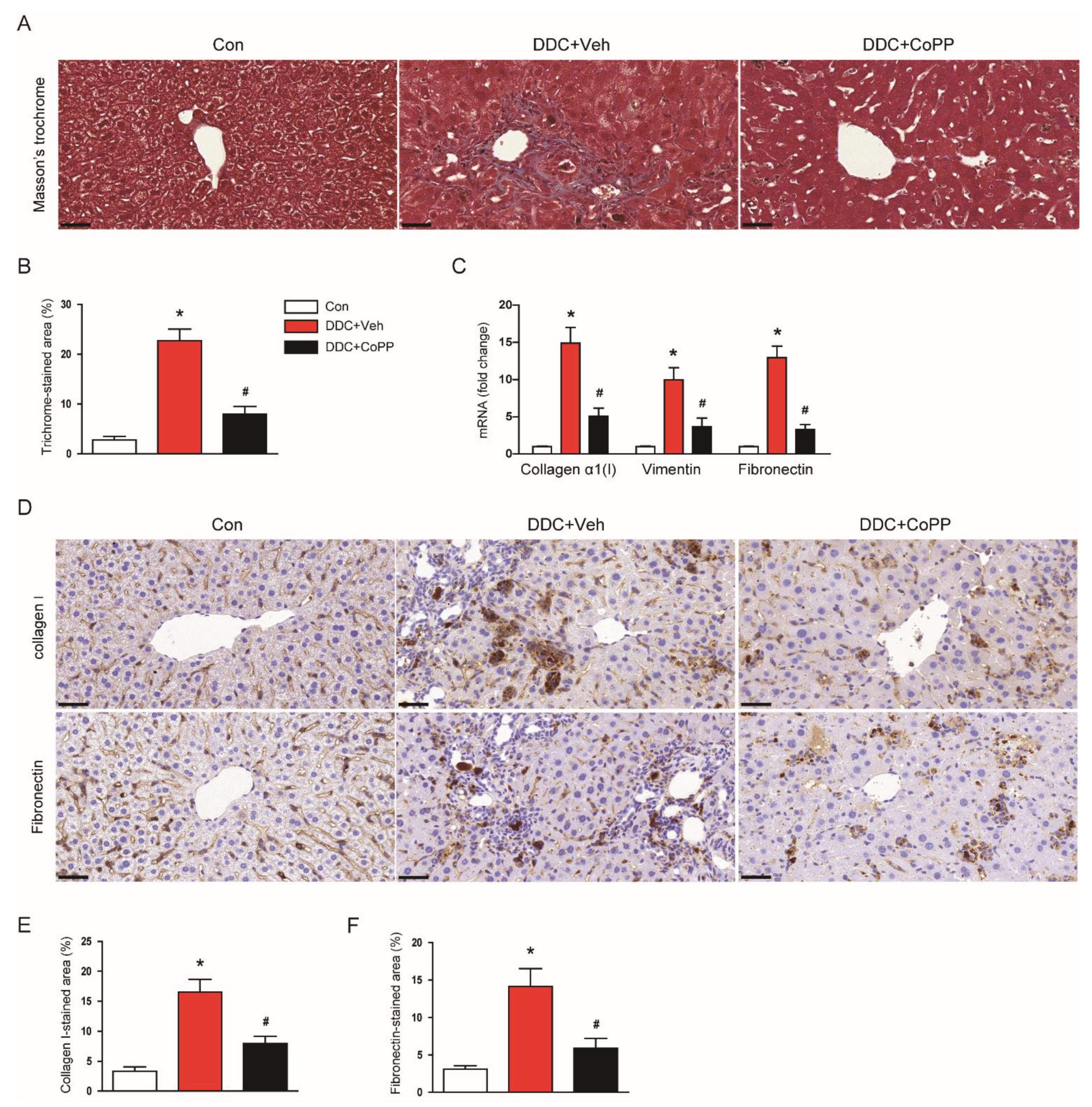
2.5. Induction of HO-1 by CoPP Inhibited the Fibrotic Process in DDC-Fed Mice
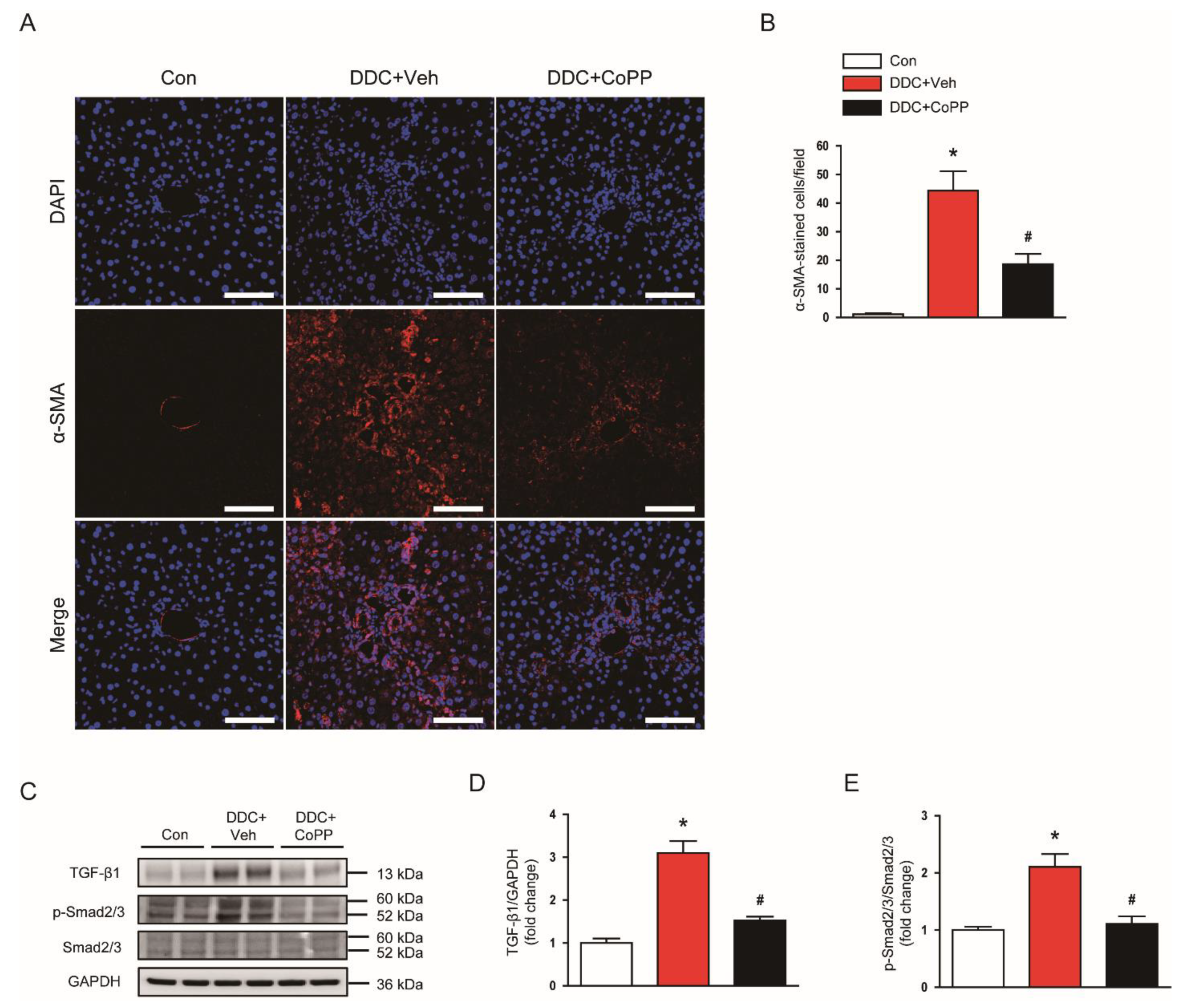
2.6. Induction of HO-1 by CoPP Suppressed Myofibroblasts and Phosphorylation of Smad2/3 in DDC-Fed Mice
3. Discussion
4. Materials and Methods
4.1. Animals Procedures
4.2. Biochemical Analysis
4.3. Histological Analysis and Immunohistochemistry
4.4. Immunofluorescent Staining
4.5. TUNEL Assay
4.6. Western Blot Analysis
4.7. Real-Time Reverse Transcription Polymerase Chain Reaction (RT-PCR)
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kriegermeier, A.; Green, R. Pediatric Cholestatic Liver Disease: Review of Bile Acid Metabolism and Discussion of Current and Emerging Therapies. Front. Med. (Lausanne) 2020, 7, 149. [Google Scholar] [CrossRef]
- Lazaridis, K.M.; LaRusso, N.F. The Cholangiopathies. Mayo Clin. Proc. 2015, 90, 791–800. [Google Scholar] [CrossRef] [Green Version]
- Roma, M.G.; Sanchez Pozzi, E.J. Oxidative stress: A radical way to stop making bile. Ann. Hepatol. 2008, 7, 16–33. [Google Scholar] [CrossRef]
- Galicia-Moreno, M.; Lucano-Landeros, S.; Monroy-Ramirez, H.C.; Silva-Gomez, J.; Gutierrez-Cuevas, J.; Santos, A.; Armendariz-Borunda, J. Roles of Nrf2 in Liver Diseases: Molecular, Pharmacological, and Epigenetic Aspects. Antioxidants 2020, 9, 980. [Google Scholar] [CrossRef]
- Aboutwerat, A.; Pemberton, P.W.; Smith, A.; Burrows, P.C.; McMahon, R.F.T.; Jain, S.K.; Warnes, T.W. Oxidant stress is a significant feature of primary biliary cirrhosis. Biochim. Biophys. Acta 2003, 1637, 142–150. [Google Scholar] [CrossRef]
- Kaffe, E.T.; Rigopoulou, E.I.; Koukoulis, G.K.; Dalekos, G.N.; Moulas, A.N. Oxidative stress and antioxidant status in patients with autoimmune liver diseases. Redox Rep. 2015, 20, 33–41. [Google Scholar] [CrossRef]
- Sommer, J.; Dorn, C.; Gäbele, E.; Bataille, F.; Freese, K.; Seitz, T.; Thasler, W.E.; Büttner, R.; Weiskirchen, R.; Bosserhoff, A.; et al. Four-And-A-Half LIM-Domain Protein 2 (FHL2) Deficiency Aggravates Cholestatic Liver Injury. Cells 2020, 9, 248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shearn, C.T.; Fennimore, B.; Orlicky, D.J.; Gao, Y.R.; Saba, L.M.; Battista, K.D.; Aivazidis, S.; Assiri, M.; Harris, P.S.; Michel, C.; et al. Cholestatic liver disease results increased production of reactive aldehydes and an atypical periportal hepatic antioxidant response. Free Radic. Biol. Med. 2019, 143, 101–114. [Google Scholar] [CrossRef]
- Hua, W.; Zhang, S.; Lu, Q.; Sun, Y.; Tan, S.; Chen, F.; Tang, L. Protective effects of n-Butanol extract and iridoid glycosides of Veronica ciliata Fisch. Against ANIT-induced cholestatic liver injury in mice. J. Ethnopharmacol. 2021, 266, 113432. [Google Scholar] [CrossRef] [PubMed]
- Salas-Silva, S.; Simoni-Nieves, A.; Razori, M.V.; López-Ramirez, J.; Barrera-Chimal, J.; Lazzarini, R.; Bello, O.; Souza, V.; Miranda-Labra, R.U.; Gutiérrez-Ruiz, M.C.; et al. HGF induces protective effects in α-naphthylisothiocyanate-induced intrahepatic cholestasis by counteracting oxidative stress. Biochem. Pharmacol. 2020, 174, 113812. [Google Scholar] [CrossRef] [PubMed]
- El-Agamy, D.S.; Almaramhy, H.H.; Ahmed, N.; Bojan, B.; Alrohily, W.D.; Elkablawy, M.A. Anti-Inflammatory Effects of Vardenafil Against Cholestatic Liver Damage in Mice: A Mechanistic Study. Cell. Physiol. Biochem. 2018, 47, 523–534. [Google Scholar] [CrossRef]
- Puentes-Pardo, J.D.; Moreno-SanJuan, S.; Carazo, Á.; León, J. Heme Oxygenase-1 in Gastrointestinal Tract Health and Disease. Antioxidants 2020, 9, 1214. [Google Scholar] [CrossRef]
- Kishimoto, Y.; Kondo, K.; Momiyama, Y. The Protective Role of Heme Oxygenase-1 in Atherosclerotic Diseases. Int. J. Mol. Sci. 2019, 20, 3628. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Ding, M.; Zhu, P.; Huang, H.; Zhuang, Q.; Shen, J.; Cai, Y.; Zhao, M.; He, Q. New Insights into the Nrf-2/HO-1 Signaling Axis and Its Application in Pediatric Respiratory Diseases. Oxid. Med. Cell. Longev. 2019, 2019, 3214196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pose, E.; Sancho-Bru, P.; Coll, M. 3,5-Diethoxycarbonyl-1,4-Dihydrocollidine Diet: A Rodent Model in Cholestasis Research. Methods Mol. Biol. 2019, 1981, 249–257. [Google Scholar] [PubMed]
- Kim, K.-H.; Sung, H.-J.; Lee, W.-R.; An, H.-J.; Kim, J.-Y.; Pak, S.C.; Han, S.-M.; Park, K.-K. Effects of Melittin Treatment in Cholangitis and Biliary Fibrosis in a Model of Xenobiotic-Induced Cholestasis in Mice. Toxins 2015, 7, 3372–3387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.H.; Zhang, B.L.; Zhang, X.M.; Tong, J.D.; Gu, Y.H.; Guo, L.L.; Jin, H.M. EGCG Attenuates Renal Damage via Reversing Klotho Hypermethylation in Diabetic db/db Mice and HK-2 Cells. Oxid. Med. Cell. Longev. 2020, 2020, 6092715. [Google Scholar] [CrossRef]
- Kim, J.-Y.; Jo, J.; Kim, K.; An, H.-J.; Gwon, M.-G.; Gu, H.; Kim, H.-J.; Yang, A.Y.; Kim, S.-W.; Jeon, E.J.; et al. Pharmacological Activation of Sirt1 Ameliorates Cisplatin-Induced Acute Kidney Injury by Suppressing Apoptosis, Oxidative Stress, and Inflammation in Mice. Antioxidants 2019, 8, 322. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.-Y.; Jo, J.; Leem, J.; Park, K.-K. Inhibition of p300 by Garcinol Protects against Cisplatin-Induced Acute Kidney Injury through Suppression of Oxidative Stress, Inflammation, and Tubular Cell Death in Mice. Antioxidants 2020, 9, 1271. [Google Scholar] [CrossRef] [PubMed]
- Shojaie, L.; Iorga, A.; Dara, L. Cell Death in Liver Diseases: A Review. Int. J. Mol. Sci. 2020, 21, 9682. [Google Scholar] [CrossRef]
- Luangmonkong, T.; Suriguga, S.; Mutsaers, H.A.M.; Groothuis, G.M.M.; Olinga, P.; Boersema, M. Targeting Oxidative Stress for the Treatment of Liver Fibrosis. Rev. Physiol. Biochem. Pharmacol. 2018, 175, 71–102. [Google Scholar] [PubMed]
- Chen, Q.; Lu, X.; Zhang, X. Noncanonical NF-κB Signaling Pathway in Liver Diseases. J. Clin. Transl. Hepatol. 2021, 9, 81–89. [Google Scholar] [PubMed]
- Jiang, A.; Okabe, H.; Popovic, B.; Preziosi, M.E.; Pradhan-Sundd, T.; Poddar, M.; Singh, S.; Bell, A.; England, S.G.; Nagarajan, S.; et al. Loss of Wnt Secretion by Macrophages Promotes Hepatobiliary Injury after Administration of 3,5-Diethoxycarbonyl-1, 4-Dihydrocollidine Diet. Am. J. Pathol. 2019, 189, 590–603. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Luo, M.; Sun, X.; Qin, J.; Yu, C.; Wen, Y.; Zhang, Q.; Gu, J.; Xia, Q.; Kong, X. DJ-1 deficiency attenuates expansion of liver progenitor cells through modulating the inflammatory and fibrogenic niches. Cell Death Dis. 2016, 7, e2257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oo, Y.H.; Shetty, S.; Adams, D.H. The role of chemokines in the recruitment of lymphocytes to the liver. Dig. Dis. 2010, 28, 31–44. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.-Y.; Kubes, P. Leukocyte adhesion in the liver: Distinct adhesion paradigm from other organs. J. Hepatol. 2008, 48, 504–512. [Google Scholar] [CrossRef]
- Kim, J.-Y.; Park, J.-H.; Kim, K.; Leem, J.; Park, K.-K. Melatonin Inhibits Transforming Growth Factor-β1-Induced Epithelial–Mesenchymal Transition in AML12 Hepatocytes. Biology 2019, 8, 84. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.-Y.; Park, J.-H.; Jeon, E.J.; Leem, J.; Park, K.-K. Melatonin Prevents Transforming Growth Factor-β1-Stimulated Transdifferentiation of Renal Interstitial Fibroblasts to Myofibroblasts by Suppressing Reactive Oxygen Species-Dependent Mechanisms. Antioxidants 2020, 9, 39. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.-Y.; Leem, J.; Jeon, E.J. Protective Effects of Melatonin Against Aristolochic Acid-Induced Nephropathy in Mice. Biomolecules 2020, 10, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gwon, M.-G.; An, H.-J.; Kim, J.-Y.; Kim, W.-H.; Gu, H.; Kim, H.-J.; Leem, J.; Jung., H.J.; Park, K.-K. Anti-fibrotic effects of synthetic TGF-β1 and Smad oligodeoxynucleotide on kidney fibrosis in vivo and in vitro through inhibition of both epithelial dedifferentiation and endothelial-mesenchymal transitions. FASEB J. 2020, 34, 333–349. [Google Scholar] [CrossRef] [Green Version]
- Banales, J.M.; Huebert, R.C.; Karlsen, T.; Strazzabosco, M.; LaRusso, N.F.; Gores, G.M. Cholangiocyte pathobiology. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 269–281. [Google Scholar] [CrossRef] [PubMed]
- Kie, J.-H.; Kapturczak, M.H.; Traylor, A.; Agarwal, A.; Hill-Kapturczak, N. Heme oxygenase-1 deficiency promotes epithelial-mesenchymal transition and renal fibrosis. J. Am. Soc. Nephrol. 2008, 19, 1681–1691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mark, A.; Hock, T.; Kapturczak, M.H.; Agarwal, A.; Hill-Kapturczak, N. Induction of heme oxygenase-1 modulates the profibrotic effects of transforming growth factor-beta in human renal tubular epithelial cells. Cell. Mol. Biol. (Noisy-le-grand) 2005, 51, 357–362. [Google Scholar]
- Barikbin, R.; Neureiter, D.; Wirth, J.; Erhardt, A.; Schwinge, D.; Kluwe, J.; Schramm, C.; Tiegs, G.; Sass, G. Induction of heme oxygenase 1 prevents progression of liver fibrosis in Mdr2 knockout mice. Hepatology 2012, 55, 553–562. [Google Scholar] [CrossRef]
- Du, J.; Ren, W.; Zhang, Q.; Fu, N.; Han, F.; Cui, P.; Li, W.; Kong, L.; Zhao, S.; Wang, R.; et al. Heme Oxygenase-1 Suppresses Wnt Signaling Pathway in Nonalcoholic Steatohepatitis-Related Liver Fibrosis. Biomed. Res. Int. 2020, 2020, 4910601. [Google Scholar] [CrossRef]
- Li, D.; Zhao, D.; Du, J.; Dong, S.; Aldhamin, Z.; Yuan, X.; Li, W.; Du, H.; Zhao, W.; Cui, L.; et al. Heme oxygenase-1 alleviated non-alcoholic fatty liver disease via suppressing ROS-dependent endoplasmic reticulum stress. Life Sci. 2020, 253, 117678. [Google Scholar] [CrossRef]
- Yang, H.; Zhao, L.-F.; Zhao, Z.-F.; Wang, Y.; Zhao, J.-J.; Zhang, L. Heme oxygenase-1 prevents liver fibrosis in rats by regulating the expression of PPARγ and NF-κB. World J. Gastroenterol. 2012, 18, 1680–1688. [Google Scholar] [CrossRef] [PubMed]
- Tsui, T.-Y.; Lau, C.-K.; Ma, J.; Glockzin, G.; Obed, A.; Schlitt, H.-J.; Fan, S.-T. Adeno-associated virus-mediated heme oxygenase-1 gene transfer suppresses the progression of micronodular cirrhosis in rats. World J. Gastroenterol. 2006, 12, 2016–2023. [Google Scholar] [CrossRef] [Green Version]
- Perez, M.-J.; Briz, O. Bile-acid-induced cell injury and protection. World J. Gastroenterol. 2009, 14, 1677–1689. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yang, C.; Elsheikh, N.A.H.; Li, C.; Yang, F.; Wang, G.; Li, L. HO-1 reduces heat stress-induced apoptosis in bovine granulosa cells by suppressing oxidative stress. Aging (Albany N. Y.) 2019, 11, 5535–5547. [Google Scholar] [CrossRef]
- Yamamoto, H.; Saito, M.; Goto, T.; Ueshima, K.; Ishida, M.; Hayashi, S.; Ikoma, K.; Mazda, O.; Kubo, T. Heme oxygenase-1 prevents glucocorticoid and hypoxia-induced apoptosis and necrosis of osteocyte-like cells. Med. Mol. Morphol. 2019, 52, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wu, B.; Teng, D.; Sun, X.; Li, J.; Li, J.; Zhang, G.; Cai, J. Cobalt-protoporphyrin enhances heme oxygenase 1 expression and attenuates liver ischemia/reperfusion injury by inhibiting apoptosis. Mol. Med. Rep. 2018, 17, 4567–4572. [Google Scholar] [CrossRef] [Green Version]
- Vitali, S.H.; Fernandez-Gonzalez, A.; Nadkarni, J.; Kwong, A.; Rose, C.; Mitsialis, S.A.; Kourembanas, S. Heme oxygenase-1 dampens the macrophage sterile inflammasome response and regulates its components in the hypoxic lung. Am. J. Physiol. Lung Cell. Mol. Physiol. 2020, 318, L125–L134. [Google Scholar] [CrossRef]
- Rossi, M.; Delbauve, S.; Roumeguère, T.; Wespes, E.; Leo, O.; Flamand, V.; Moine, A.L.; Hougardy, J.-M. HO-1 mitigates acute kidney injury and subsequent kidney-lung cross-talk. Free Radic. Res. 2019, 53, 1035–1043. [Google Scholar] [CrossRef] [PubMed]
- Lakhani, H.V.; Zehra, M.; Pillai, S.S.; Puri, N.; Shapiro, J.I.; Abraham, N.G.; Sodhi, K. Beneficial Role of HO-1-SIRT1 Axis in Attenuating Angiotensin II-Induced Adipocyte Dysfunction. Int. J. Mol. Sci. 2019, 20, 3205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roehlen, N.; Crouchet, E.; Baumert, T.F. Liver Fibrosis: Mechanistic Concepts and Therapeutic Perspectives. Cells 2020, 9, 875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weng, H.L.; Feng, D.C.; Radaeva, S.; Kong, X.N.; Wang, L.; Liu, Y.; Li, Q.; Shen, H.; Gao, Y.P.; Müllenbach, R.; et al. IFN-γ inhibits liver progenitor cell proliferation in HBV-infected patients and in 3,5-diethoxycarbonyl-1,4-dihydrocollidine diet-fed mice. J. Hepatol. 2013, 59, 738–745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barikbin, R.; Berkhout, L.; Bolik, J.; Schmidt-Arras, D.; Ernst, T.; Ittrich, H.; Adam, G.; Parplys, A.; Casar, C.; Krech, T.; et al. Early heme oxygenase 1 induction delays tumour initiation and enhances DNA damage repair in liver macrophages of Mdr2 -/- mice. Sci. Rep. 2018, 8, 16238. [Google Scholar] [CrossRef] [PubMed]
- Gu, Q.; Wu, Q.; Jin, M.; Xiao, Y.; Xu, J.; Mao, C.; Zhao, F.; Zhang, Y.; Zhang, Y. Heme oxygenase-1 alleviates mouse hepatic failure through suppression of adaptive immune responses. J. Pharmacol. Exp. Ther. 2012, 340, 2–10. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′→3′) |
|---|---|
| HO-1 1 | Sense: TCAAGGCCTCAGACAAATCC Antisense: ACAACCAGTGAGTGGAGCCT |
| TNF-α 2 | Sense: GACGTGGAACTGGCAGAAGAG Antisense: CCGCCTGGAGTTCTGGAA |
| IL-6 3 | Sense: CCAGAGATACAAAGAAATGATGG Antisense: ACTCCAGAAGACCAGAGGAAAT |
| CCL2 4 | Sense: GGGCCTGCTGTTCACAGTT Antisense: CCAGCCTACTCATTGGGAT |
| CX3CL1 5 | Sense: ATGACCTCACGAATCCCAGTG Antisense: CCGCCTCAAAACTTCCAATGC |
| E-selectin | Sense: AGCTACCCATGGAACACGAC Antisense: ACGCAAGTTCTCCAGCTGTT |
| VCAM-1 6 | Sense: CCCAGGTGGAGGTCTACTCA Antisense: CAGGATTTTGGGAGCTGGTA |
| collagen α1(I) | Sense: GAGTGAGGCCACGCATGA Antisense: AGCCGGAGGTCCACAAAG |
| vimentin | Sense: GATCGATGTGGACGTTTCCAA Antisense: GTTGGCAGCCTCAGAGAGGT |
| fibronectin | Sense: CGAGGTGACAGAGACCACAA Antisense: CTGGAGTCAAGCCAGACACA |
| GAPDH 7 | Sense: ACTCCACTCACGGCAAATTC Antisense: TCTCCATGGTGGTGAAGACA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.-Y.; Choi, Y.; Leem, J.; Song, J.E. Heme Oxygenase-1 Induction by Cobalt Protoporphyrin Ameliorates Cholestatic Liver Disease in a Xenobiotic-Induced Murine Model. Int. J. Mol. Sci. 2021, 22, 8253. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22158253
Kim J-Y, Choi Y, Leem J, Song JE. Heme Oxygenase-1 Induction by Cobalt Protoporphyrin Ameliorates Cholestatic Liver Disease in a Xenobiotic-Induced Murine Model. International Journal of Molecular Sciences. 2021; 22(15):8253. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22158253
Chicago/Turabian StyleKim, Jung-Yeon, Yongmin Choi, Jaechan Leem, and Jeong Eun Song. 2021. "Heme Oxygenase-1 Induction by Cobalt Protoporphyrin Ameliorates Cholestatic Liver Disease in a Xenobiotic-Induced Murine Model" International Journal of Molecular Sciences 22, no. 15: 8253. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22158253