
Visfatin Promotes the Metastatic Potential of Chondrosarcoma Cells by Stimulating AP-1-Dependent MMP-2 Production in the MAPK Pathway

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
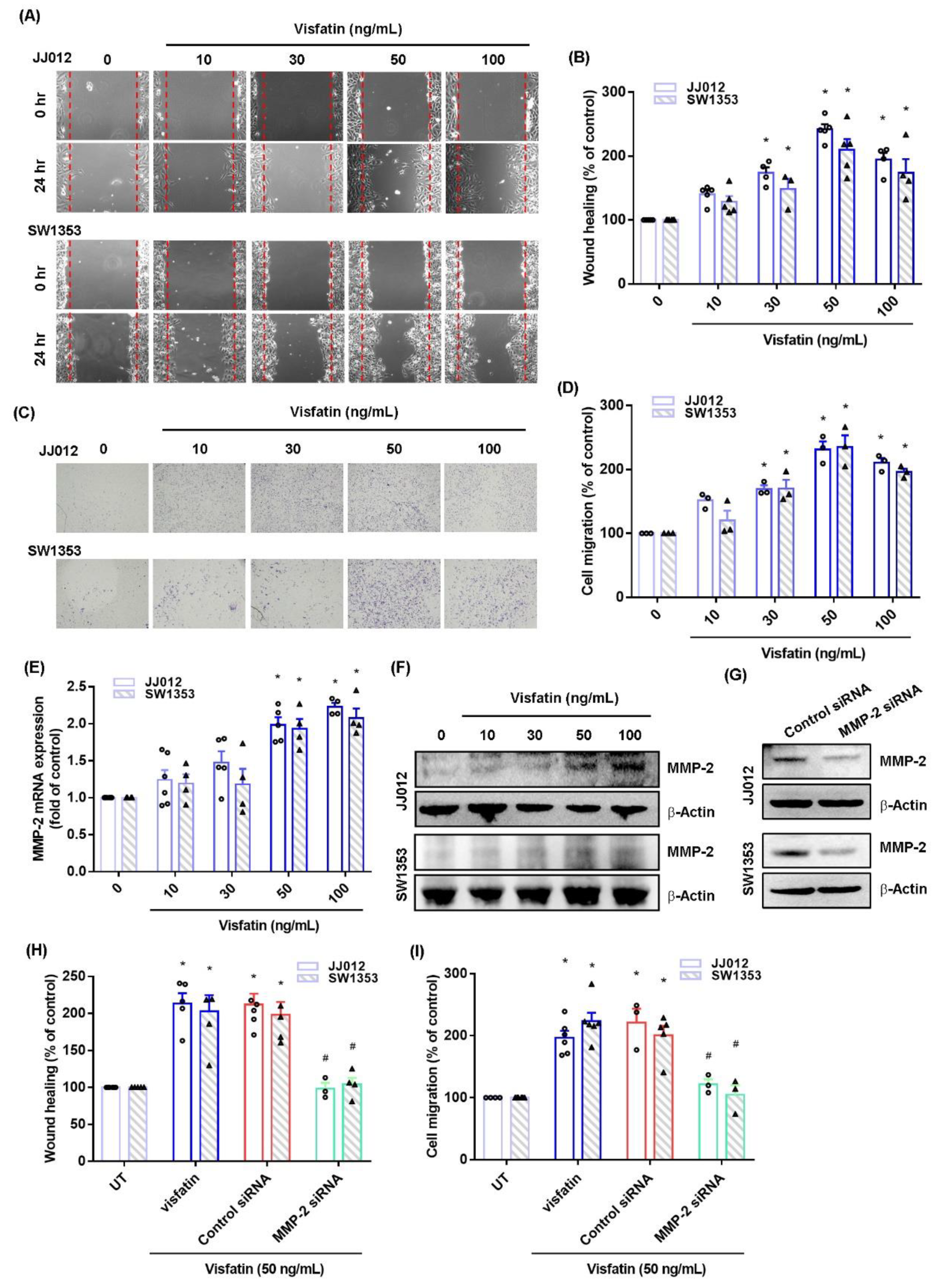
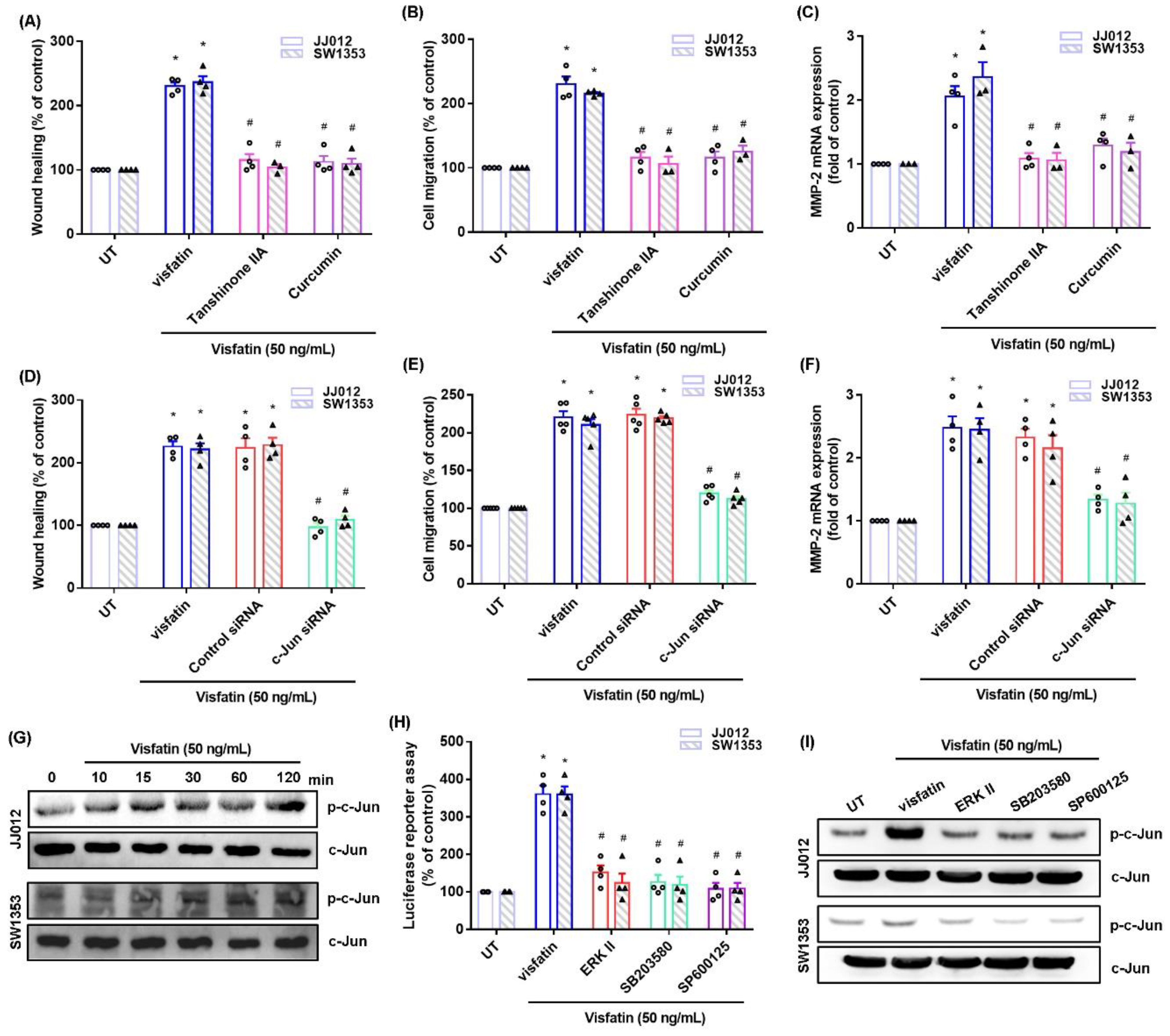
2.1. Visfatin Facilitates MMP-2-Dependent Migration in Chondrosarcoma Cells
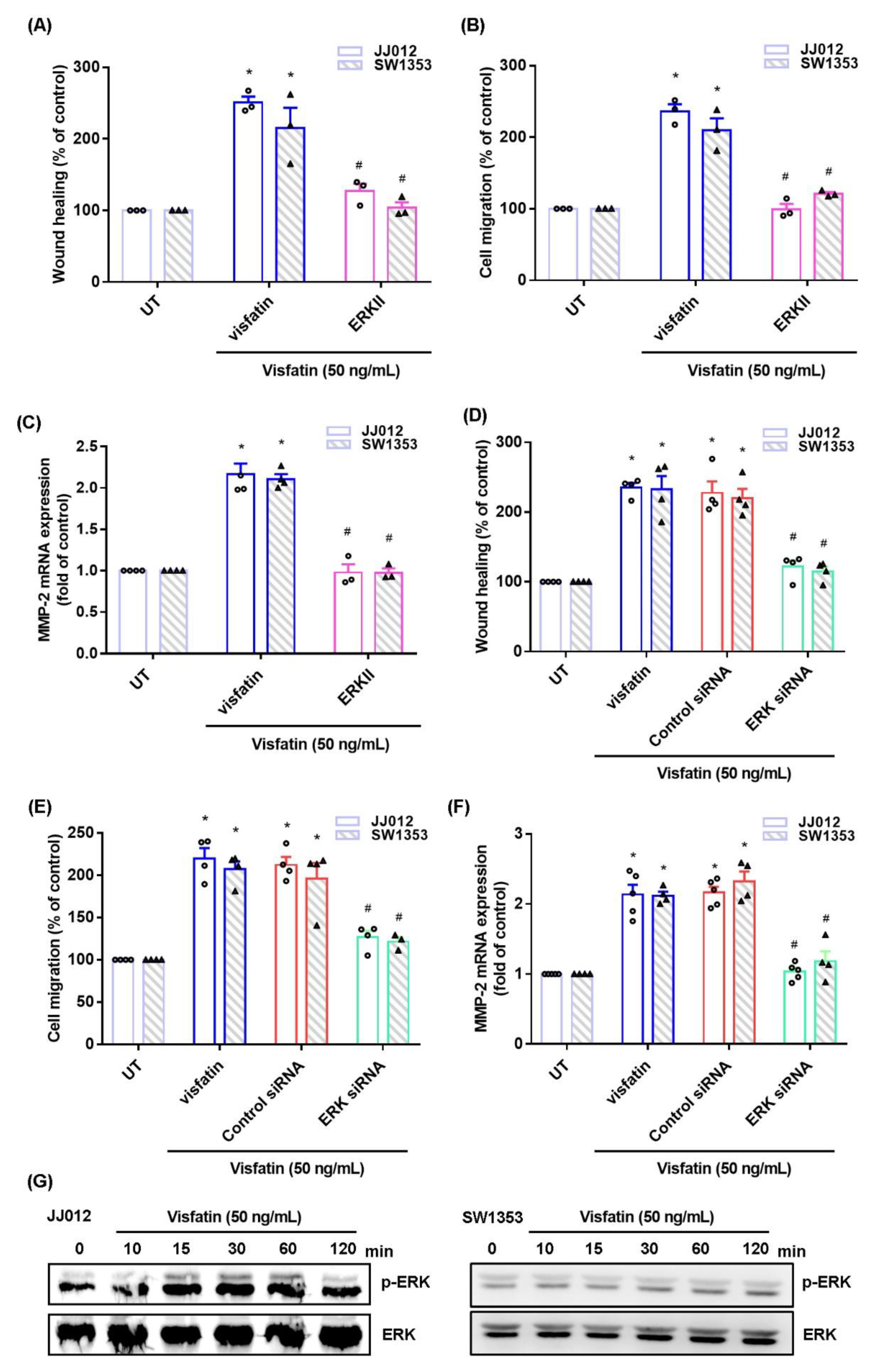
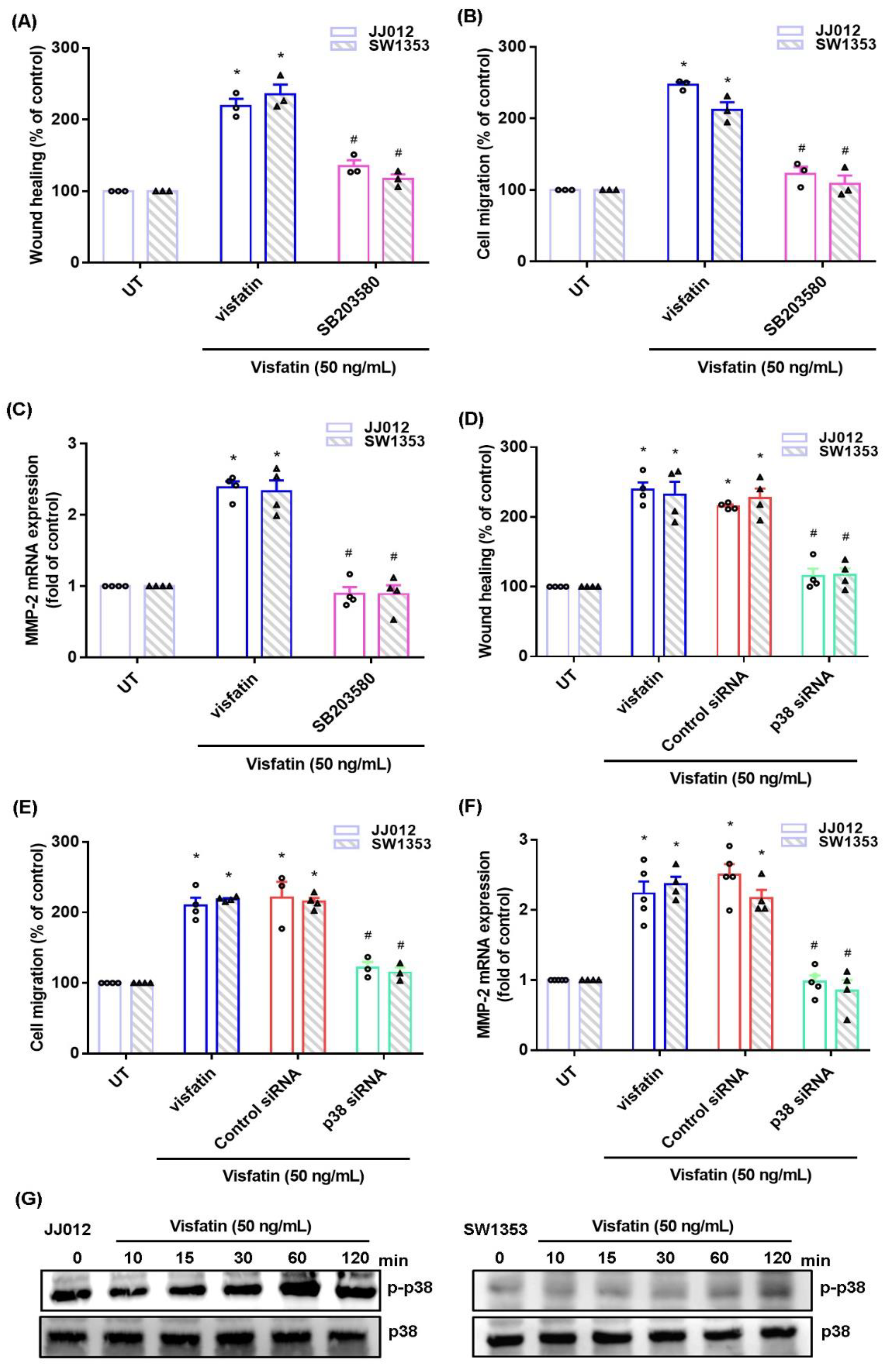
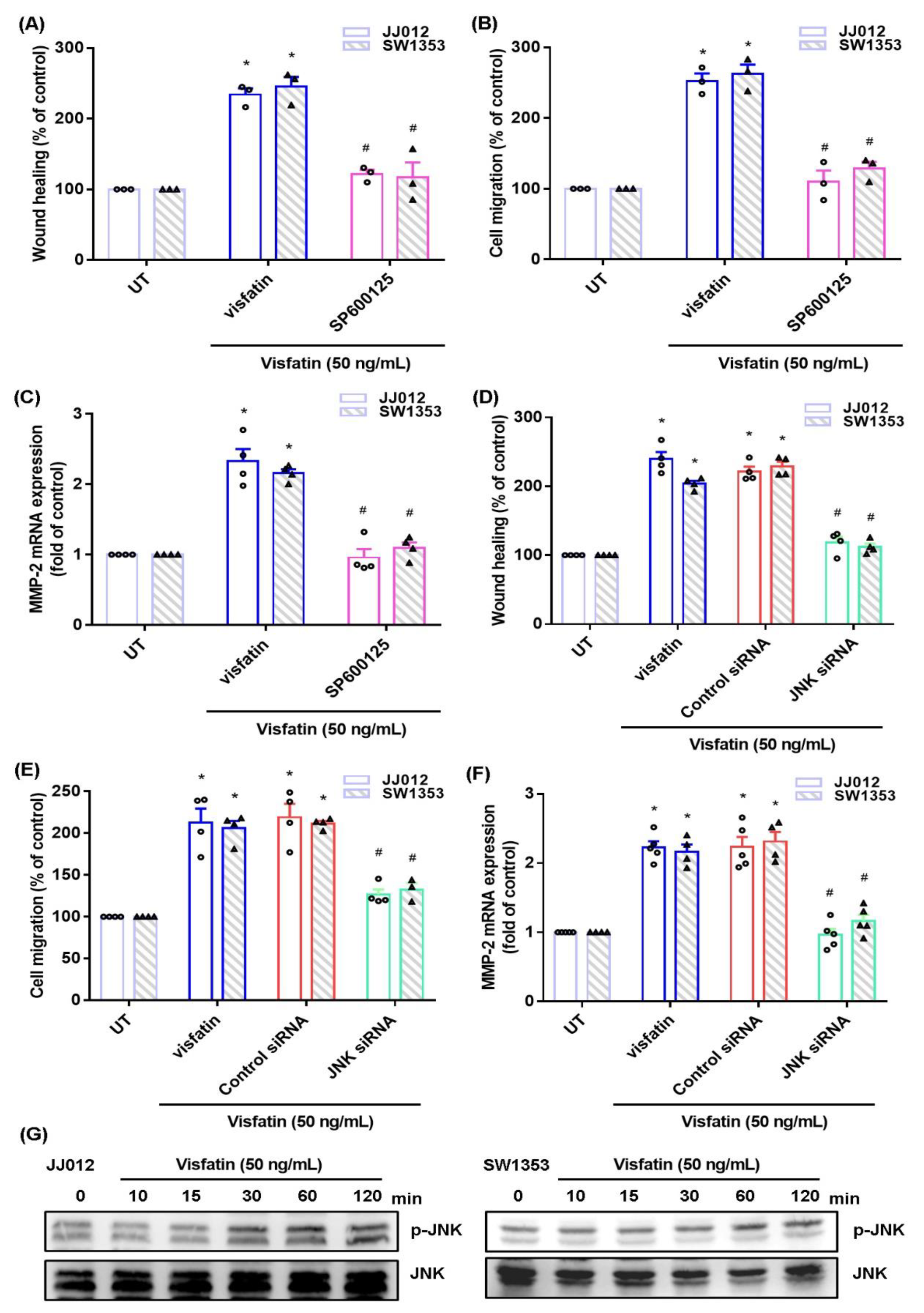
2.2. The MAPK Signaling Pathway Mediates the Effects of Visfatin upon MMP-2 Synthesis and Migration of Human Chondrosarcoma Cells
2.3. AP-1-Mediated MMP-2 Expression Regulates Visfatin-Induced Stimulation of Chondrosarcoma Cell Migration
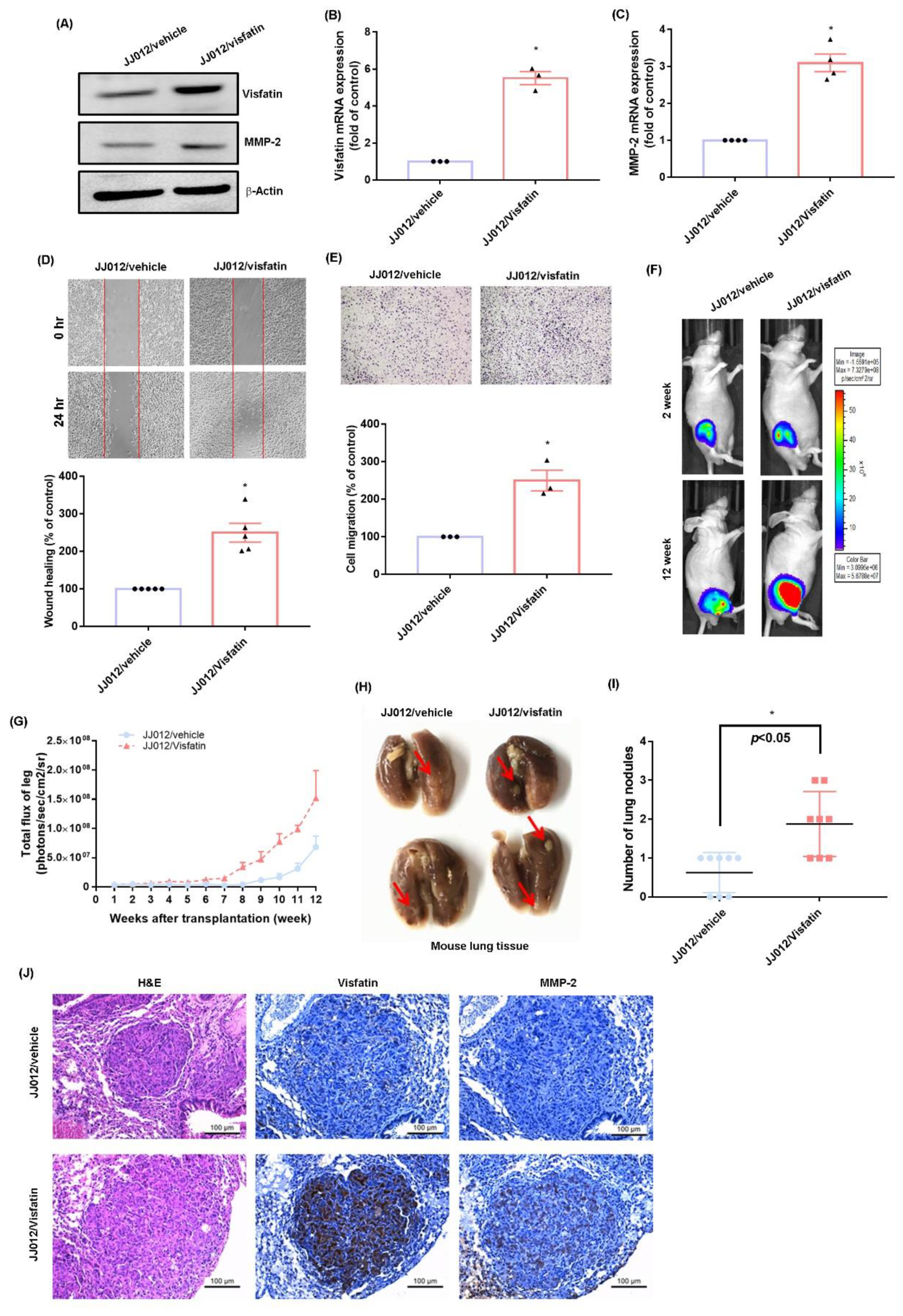
2.4. Overexpression of Visfatin Facilitates the Metastasis of Chondrosarcoma Cells in the Mouse Lung
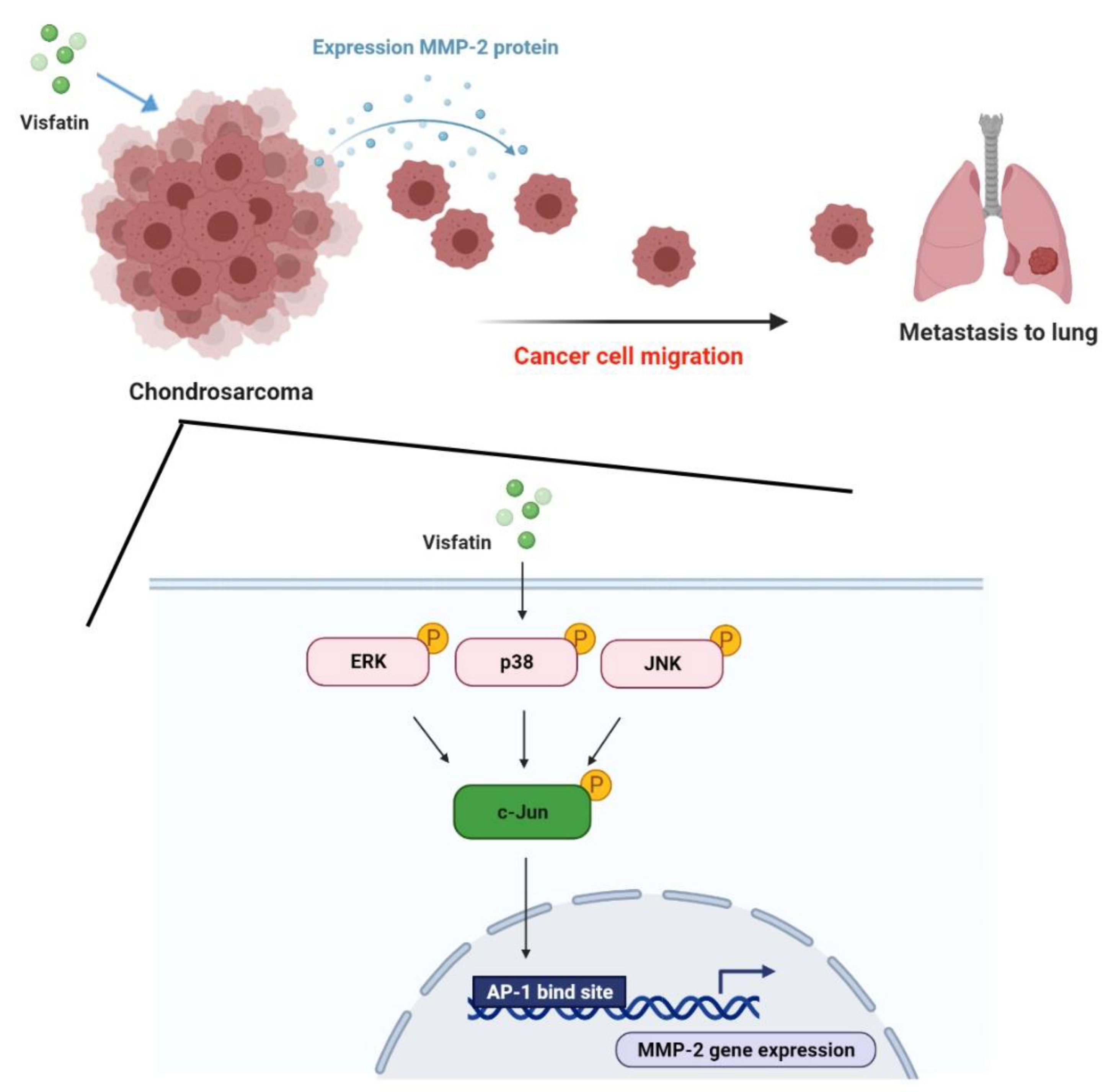
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture
4.3. Cell Migration Assay
4.4. Wound Healing Assay
4.5. Western Blot
4.6. mRNA and miRNA Quantification
4.7. Luciferase Assay
4.8. Small Interfering RNA (siRNA) Transfection
4.9. Establishment of Stably Transfected Cells
4.10. Tumor Xenograft Study
4.11. Immunohistochemistry (IHC) Staining
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Macdonald, I.J.; Lin, C.-Y.; Kuo, S.-J.; Su, C.-M.; Tang, C.-H. An update on current and future treatment options for chondrosarcoma. Expert Rev. Anticancer Ther. 2019, 19, 773–786. [Google Scholar] [CrossRef]
- Chen, P.-C.; Cheng, H.-C.; Yang, S.-F.; Lin, C.-W.; Tang, C.-H. The CCN Family Proteins: Modulators of Bone Development and Novel Targets in Bone-Associated Tumors. BioMed Res. Int. 2014, 2014, 1–11. [Google Scholar] [CrossRef]
- Chen, J.-C.; Fong, Y.-C.; Tang, C.-H. Novel Strategies for the Treatment of Chondrosarcomas: Targeting Integrins. BioMed Res. Int. 2013, 2013, 1–11. [Google Scholar] [CrossRef]
- Ferguson, J.L.; Turner, S.P. Bone Cancer: Diagnosis and Treatment Principles. Am. Fam. Physician 2018, 98, 205–213. [Google Scholar]
- Fan, T.M.; Roberts, R.D.; Lizardo, M.M. Understanding and Modeling Metastasis Biology to Improve Therapeutic Strategies for Combating Osteosarcoma Progression. Front. Oncol. 2020, 10, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steeg, P.S. Targeting metastasis. Nature reviews. Cancer 2016, 16, 201–218. [Google Scholar]
- Hsu, C.-J.; Wu, M.-H.; Chen, C.-Y.; Tsai, C.-H.; Hsu, H.-C.; Tang, C.-H. AMP-activated protein kinase activation mediates CCL3-induced cell migration and matrix metalloproteinase-2 expression in human chondrosarcoma. Cell Commun. Signal. 2013, 11, 68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsou, H.-K.; Chen, H.-T.; Hung, Y.-H.; Chang, C.-H.; Li, T.-M.; Fong, Y.-C.; Tang, C.-H. HGF and c-Met Interaction Promotes Migration in Human Chondrosarcoma Cells. PLoS ONE 2013, 8, e53974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khaidizar, F.; Bessho, Y.; Nakahata, Y. Nicotinamide Phosphoribosyltransferase as a Key Molecule of the Aging/Senescence Process. Int. J. Mol. Sci. 2021, 22, 3709. [Google Scholar] [CrossRef]
- Dakroub, A.; Nasser, S.A.; Younis, N.; Bhagani, H.; Al-Dhaheri, Y.; Pintus, G.; Eid, A.A.; El-Yazbi, A.F.; Eid, A.H. Visfatin: A Possible Role in Cardiovasculo-Metabolic Disorders. Cells 2020, 9, 2444. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, M.; Mianabadi, F.; Mehrad-Majd, H. Circulating visfatin levels and cancers risk: A systematic review and meta-analysis. J. Cell. Physiol. 2018, 234, 5011–5022. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.-C. The role of visfatin in cancer proliferation, angiogenesis, metastasis, drug resistance and clinical prognosis. Cancer Manag. Res. 2019, 11, 3481–3491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Liu, N.; Zhao, Y.; Zhu, X.; Wang, C.; Liu, Q.; Gao, C.; Zhao, X.; Li, J. Oncogenic USP22 supports gastric cancer growth and metastasis by activating c-Myc/NAMPT/SIRT1-dependent FOXO1 and YAP signaling. Aging 2019, 11, 9643–9660. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhang, N.; Liu, Y.; Su, P.; Liang, Y.; Li, Y.; Wang, X.; Chen, T.; Song, X.; Sang, Y.; et al. Epigenetic Regulation of NAMPT by NAMPT-AS Drives Metastatic Progression in Triple-Negative Breast Cancer. Cancer Res. 2019, 79, 3347–3359. [Google Scholar] [CrossRef] [Green Version]
- Ezzati-Mobaser, S.; Malekpour-Dehkordi, Z.; Nourbakhsh, M.; Tavakoli-Yaraki, M.; Ahmadpour, F.; Golpour, P.; Nourbakhsh, M. The up-regulation of markers of adipose tissue fibrosis by visfatin in pre-adipocytes as well as obese children and adolescents. Cytokine 2020, 134, 155193. [Google Scholar] [CrossRef]
- Roomi, M.W.; Kalinovsky, T.; Monterrey, J.; Rath, M.; Niedzwiecki, A. In vitro modulation of MMP-2 and MMP-9 in adult human sarcoma cell lines by cytokines, inducers and inhibitors. Int. J. Oncol. 2013, 43, 1787–1798. [Google Scholar] [CrossRef] [Green Version]
- Wu, M.H.; Chen, L.-M.; Hsu, H.-H.; Lin, J.A.; Lin, Y.-M.; Tsai, F.-J.; Tsai, C.-H.; Huang, C.-Y.; Tang, C.-H. Endothelin-1 enhances cell migration through COX-2 up-regulation in human chondrosarcoma. Biochim. Biophys. Acta Gen. Subj. 2013, 1830, 3355–3364. [Google Scholar] [CrossRef]
- Hou, C.-H.; Chiang, Y.-C.; Fong, Y.-C.; Tang, C.-H. WISP-1 increases MMP-2 expression and cell motility in human chondrosarcoma cells. Biochem. Pharmacol. 2011, 81, 1286–1295. [Google Scholar] [CrossRef]
- Zheng, P.; Chen, X.; Xie, J.; Chen, X.; Lin, S.; Ye, L.; Chen, L.; Lin, J.; Yu, X.; Zheng, M. Capn4 is induced by and required for Epstein-Barr virus latent membrane protein 1 promotion of nasopharyngeal carcinoma metastasis through ERK/AP-1 signaling. Cancer Sci. 2019, 111, 72–83. [Google Scholar] [CrossRef] [Green Version]
- Tzeng, H.E.; Tang, C.H.; Wu, S.H.; Chen, H.T.; Fong, Y.C.; Lu, Y.C.; Chen, W.C.; Huang, H.D.; Lin, C.Y.; Wang, S.W. CCN6-mediated MMP-9 activation enhances metastatic potential of human chondrosarcoma. Cell Death Dis. 2018, 9, 955. [Google Scholar] [CrossRef] [Green Version]
- Leddy, L.R.; Holmes, R.E. Chondrosarcoma of bone. Cancer Treat. Res. 2014, 162, 117–130. [Google Scholar] [PubMed]
- Gelderblom, H.; Hogendoorn, P.C.; Dijkstra, S.D.; van Rijswijk, C.S.; Krol, A.D.; Taminiau, A.H.; Bovee, J.V. The clinical approach towards chondrosarcoma. Oncologist 2008, 13, 320–329. [Google Scholar] [CrossRef] [PubMed]
- Ye, C.; Qi, L.; Li, X.; Wang, J.; Yu, J.; Zhou, B.; Guo, C.; Chen, J.; Zheng, S. Targeting the NAD(+) salvage pathway suppresses APC mutation-driven colorectal cancer growth and Wnt/beta-catenin signaling via increasing Axin level. Cell Commun. Signal. CCS 2020, 18, 16. [Google Scholar] [CrossRef] [Green Version]
- Cao, D.; Chu, L.; Xu, Z.; Gong, J.; Deng, R.; Wang, B.; Zhou, S. Visfatin facilitates gastric cancer malignancy by targeting snai1 via the NF-kappaB signaling. Hum. Exp. Toxicol. 2021, 09603271211006168. [Google Scholar] [CrossRef]
- Li, Y.; Yu, C.; Deng, W. Roles and mechanisms of adipokines in drug resistance of tumor cells. Eur. J. Pharmacol. 2021, 899, 174019. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.-S.; Lin, C.-W.; Su, S.-C.; Yang, S.-F. Pharmacodynamic considerations in the use of matrix metalloproteinase inhibitors in cancer treatment. Expert Opin. Drug Metab. Toxicol. 2016, 12, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Lu, Y.-T.; Sun, Y.; Shi, Z.-H.; Li, N.-G.; Tang, Y.-P.; Duan, J.-A. Recent opportunities in matrix metalloproteinase inhibitor drug design for cancer. Expert Opin. Drug Discov. 2017, 13, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-H.; Kuo, S.-J.; Liu, S.-C.; Wang, S.-W.; Tsai, C.-H.; Fong, Y.-C.; Tang, C.-H. Apelin Affects the Progression of Osteoarthritis by Regulating VEGF-Dependent Angiogenesis and miR-150-5p Expression in Human Synovial Fibroblasts. Cells 2020, 9, 594. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.-Y.; Su, C.-M.; Huang, Y.-L.; Tsai, C.-H.; Fuh, L.-J.; Tang, C.-H. CCN1 Induces Oncostatin M Production in Osteoblasts via Integrin-Dependent Signal Pathways. PLoS ONE 2014, 9, e106632. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Wang, K.; Chen, Y. Ophiopogonin D suppresses TGF-beta1-mediated metastatic behavior of MDA-MB-231 breast carcinoma cells via regulating ITGB1/FAK/Src/AKT/beta-catenin/MMP-9 signaling axis. Toxicol. Vitr. 2020, 69, 104973. [Google Scholar] [CrossRef]
- Chuang, C.-Y.; Ho, Y.-C.; Lin, C.-W.; Yang, W.-E.; Yu, Y.-L.; Tsai, M.-C.; Yang, S.-F.; Su, S.-C. Salvianolic acid A suppresses MMP-2 expression and restrains cancer cell invasion through ERK signaling in human nasopharyngeal carcinoma. J. Ethnopharmacol. 2020, 252, 112601. [Google Scholar] [CrossRef] [PubMed]
- Egeblad, M.; Werb, Z. New functions for the matrix metalloproteinases in cancer progression. Nat. Rev. Cancer 2002, 2, 161–174. [Google Scholar] [CrossRef] [PubMed]
- Jie, Z.; Xie, Z.; Zhao, X.; Sun, X.; Yu, H.; Pan, X.; Shen, S.; Qin, A.; Fang, X.; Fan, S. Glabridin inhibits osteosarcoma migration and invasion via blocking the p38- and JNK-mediated CREB-AP1 complexes formation. J. Cell. Physiol. 2018, 234, 4167–4178. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-P.; Wang, S.-W.; Wu, Y.-C.; Lin, L.-W.; Tsai, F.-J.; Yang, J.-S.; Li, T.-M.; Tang, C.-H. Soya-cerebroside inhibits VEGF-facilitated angiogenesis in endothelial progenitor cells. Food Agric. Immunol. 2020, 31, 193–204. [Google Scholar] [CrossRef] [Green Version]
- Hou, C.-H.; Lin, F.-L.; Hou, S.-M.; Liu, J.-F. Cyr61 promotes epithelial-mesenchymal transition and tumor metastasis of osteosarcoma by Raf-1/MEK/ERK/Elk-1/TWIST-1 signaling pathway. Mol. Cancer 2014, 13, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.-F.; Lee, C.-W.; Tsai, M.-H.; Tang, C.-H.; Chen, P.-C.; Lin, L.-W.; Lin, C.-Y.; Lu, C.-H.; Lin, Y.-F.; Yang, S.-H.; et al. Thrombospondin 2 promotes tumor metastasis by inducing matrix metalloproteinase-13 production in lung cancer cells. Biochem. Pharmacol. 2018, 155, 537–546. [Google Scholar] [CrossRef]
- Lee, H.-P.; Chen, P.-C.; Wang, S.-W.; Fong, Y.-C.; Tsai, C.-H.; Tsai, F.-J.; Chung, J.-G.; Huang, C.-Y.; Yang, J.-S.; Hsu, Y.-M.; et al. Plumbagin suppresses endothelial progenitor cell-related angiogenesis in vitro and in vivo. J. Funct. Foods 2018, 52, 537–544. [Google Scholar] [CrossRef]
- Lee, H.-P.; Wang, S.-W.; Wu, Y.-C.; Tsai, C.-H.; Tsai, F.-J.; Chung, J.-G.; Huang, C.-Y.; Yang, J.-S.; Hsu, Y.-M.; Yin, M.-C.; et al. Glucocerebroside reduces endothelial progenitor cell-induced angiogenesis. Food Agric. Immunol. 2019, 30, 1033–1045. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.C.; Tsai, C.H.; Wu, T.Y.; Tsai, C.H.; Tsai, F.J.; Chung, J.G.; Huang, C.Y.; Yang, J.S.; Hsu, Y.M.; Yin, M.C.; et al. Soya-cerebroside reduces IL-1 beta-induced MMP-1 production in chondrocytes and inhibits cartilage degradation: Implications for the treatment of osteoarthritis. Food Agric. Immunol. 2019, 30, 620–632. [Google Scholar] [CrossRef]
- Yang, Y.-C.; Chiou, P.-C.; Chen, P.-C.; Liu, P.-Y.; Huang, W.-C.; Chao, C.-C.; Tang, C.-H. Melatonin reduces lung cancer stemness through inhibiting of PLC, ERK, p38, β-catenin, and Twist pathways. Environ. Toxicol. 2018, 34, 203–209. [Google Scholar] [CrossRef]
- Wang, M.; Chao, C.-C.; Chen, P.-C.; Liu, P.-I.; Yang, Y.-C.; Su, C.-M.; Huang, W.-C.; Tang, C.-H. Thrombospondin enhances RANKL-dependent osteoclastogenesis and facilitates lung cancer bone metastasis. Biochem. Pharmacol. 2019, 166, 23–32. [Google Scholar] [CrossRef]
- Su, C.-M.; Tang, C.-H.; Chi, M.-J.; Lin, C.-Y.; Fong, Y.-C.; Liu, Y.-C.; Chen, W.-C.; Wang, S.-W. Resistin facilitates VEGF-C-associated lymphangiogenesis by inhibiting miR-186 in human chondrosarcoma cells. Biochem. Pharmacol. 2018, 154, 234–242. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.J.; Lin, C.Y.; Tsai, C.H.; Huang, Y.L.; Tang, C.H. Glucose suppresses IL-1beta-induced MMP-1 expression through the FAK, MEK, ERK, and AP-1 signaling pathways. Environ. Toxicol. 2018, 33, 1061–1068. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.-H.; Yang, D.-Y.; Lin, C.-Y.; Chen, T.-M.; Tang, C.-H.; Huang, Y.-L. Sphingosine-1-phosphate suppresses chondrosarcoma metastasis by upregulation of tissue inhibitor of metalloproteinase 3 through suppressing miR-101 expression. Mol. Oncol. 2017, 11, 1380–1398. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hung, S.-Y.; Lin, C.-Y.; Yu, C.-C.; Chen, H.-T.; Lien, M.-Y.; Huang, Y.-W.; Fong, Y.-C.; Liu, J.-F.; Wang, S.-W.; Chen, W.-C.; et al. Visfatin Promotes the Metastatic Potential of Chondrosarcoma Cells by Stimulating AP-1-Dependent MMP-2 Production in the MAPK Pathway. Int. J. Mol. Sci. 2021, 22, 8642. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168642
Hung S-Y, Lin C-Y, Yu C-C, Chen H-T, Lien M-Y, Huang Y-W, Fong Y-C, Liu J-F, Wang S-W, Chen W-C, et al. Visfatin Promotes the Metastatic Potential of Chondrosarcoma Cells by Stimulating AP-1-Dependent MMP-2 Production in the MAPK Pathway. International Journal of Molecular Sciences. 2021; 22(16):8642. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168642
Chicago/Turabian StyleHung, Shih-Ya, Chih-Yang Lin, Cheng-Chieh Yu, Hsien-Te Chen, Ming-Yu Lien, Yu-Wen Huang, Yi-Chin Fong, Ju-Fang Liu, Shih-Wei Wang, Wei-Cheng Chen, and et al. 2021. "Visfatin Promotes the Metastatic Potential of Chondrosarcoma Cells by Stimulating AP-1-Dependent MMP-2 Production in the MAPK Pathway" International Journal of Molecular Sciences 22, no. 16: 8642. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168642