
Complex Conformational Dynamics of the Heart Failure-Associated Pre-miRNA-377 Hairpin Revealed by Single-Molecule Optical Tweezers

 , , and
, , and
Abstract
:1. Introduction
2. Results
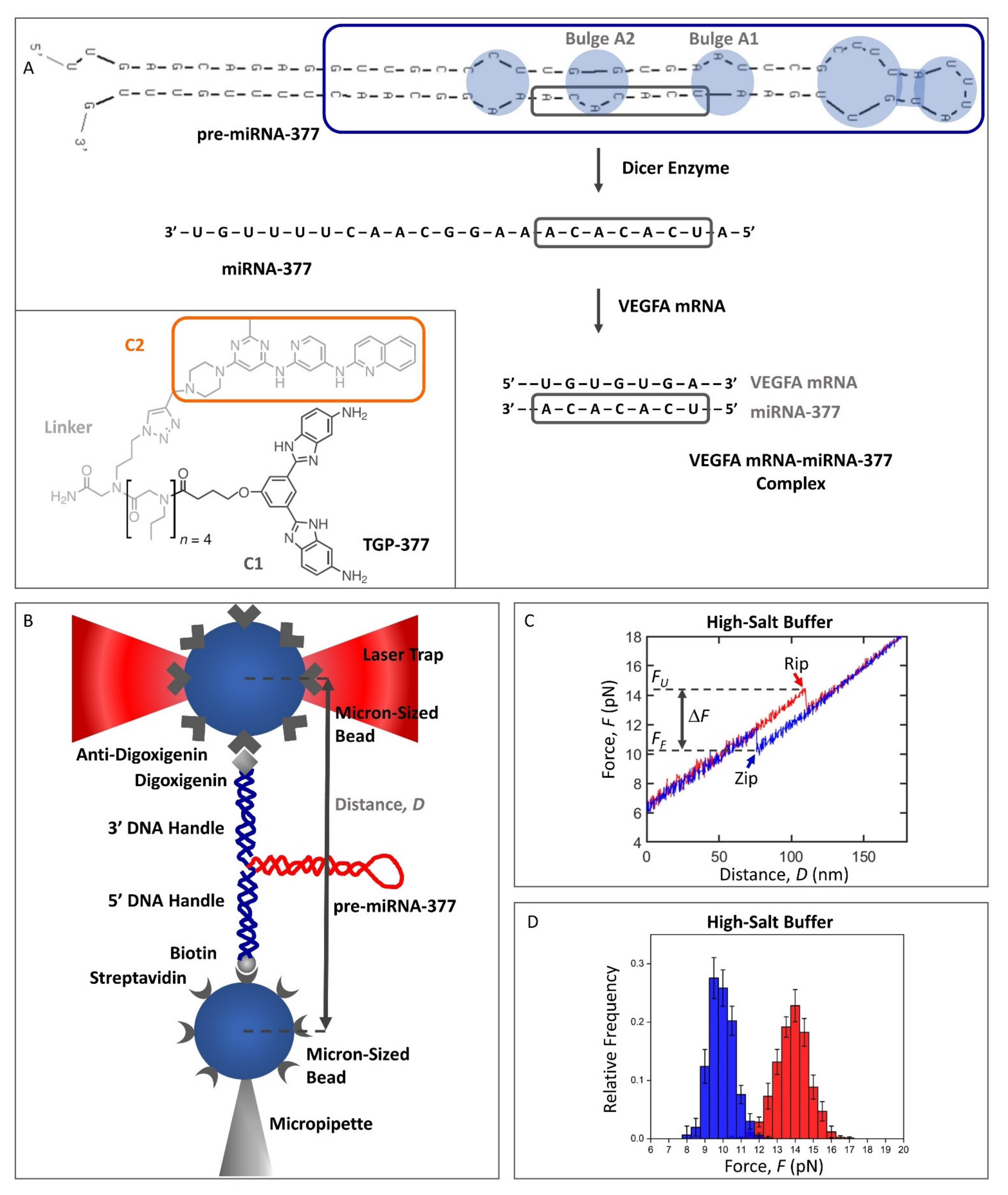
2.1. Design of the Pre-miRNA-377 Construct for Optical Tweezers Studies
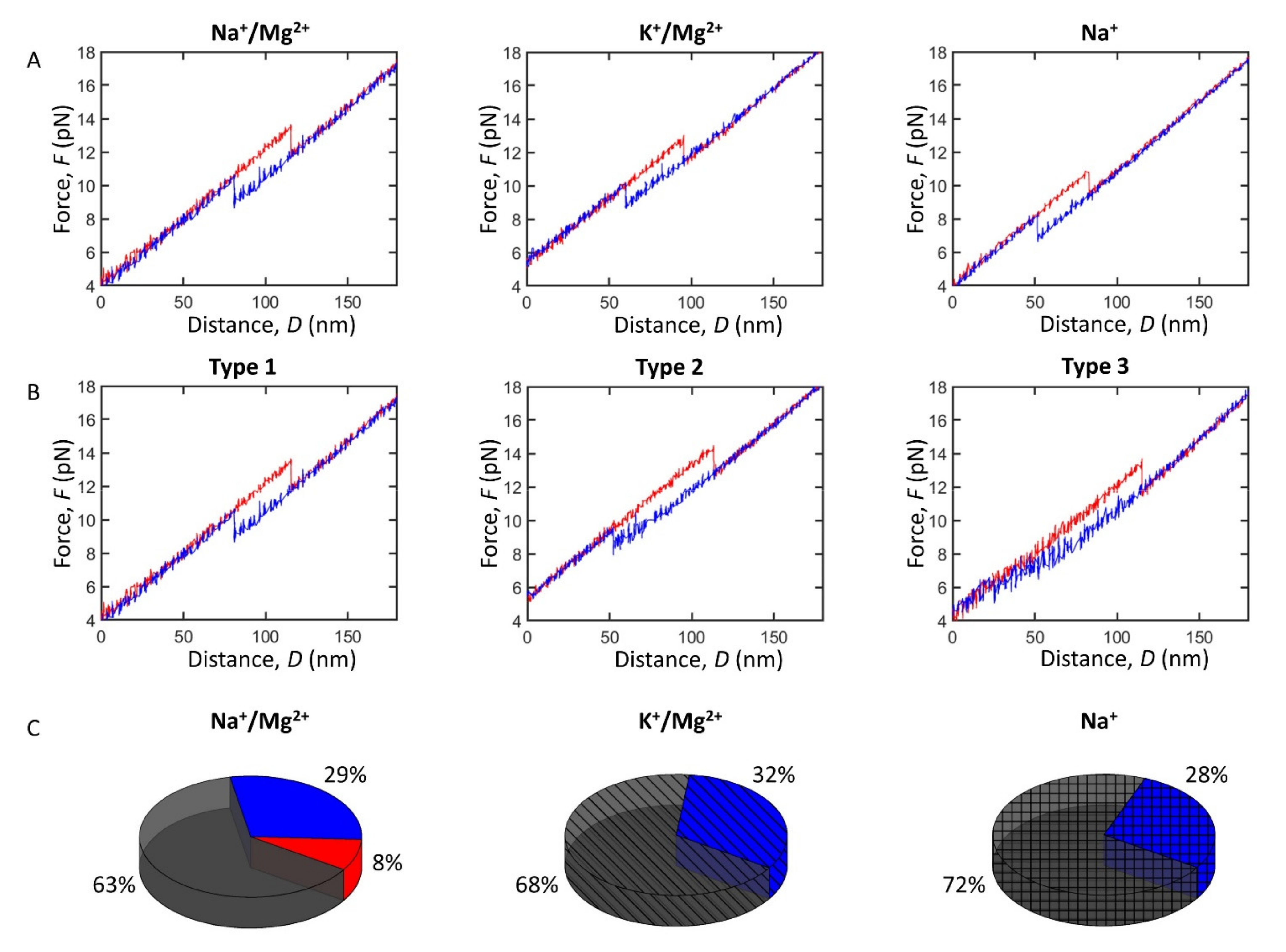
2.2. Impact of Monovalent and Divalent Ions on Pre-miRNA-377 Stability
2.3. Impact of Monovalent and Divalent Ions on Pre-miRNA-377 Dynamics
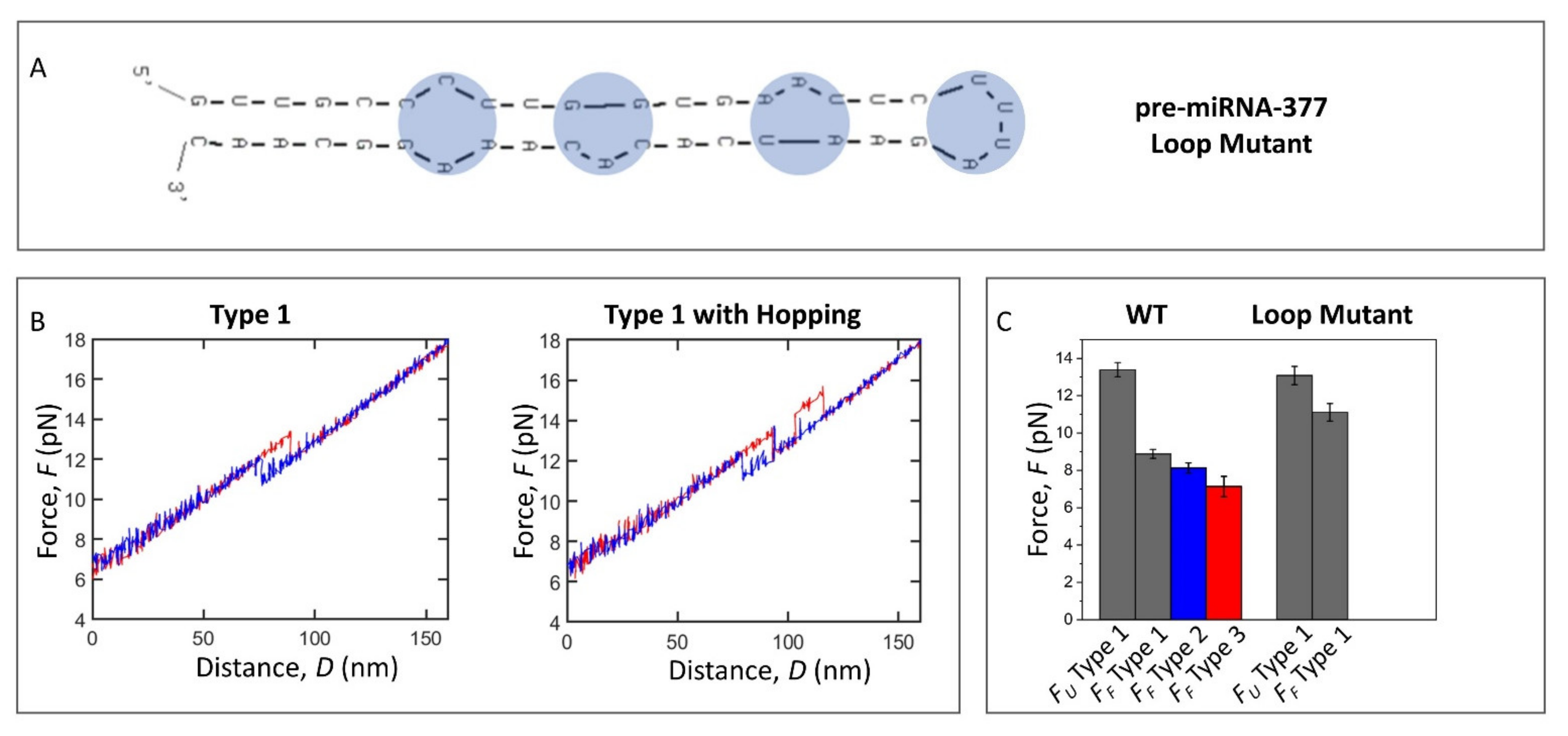
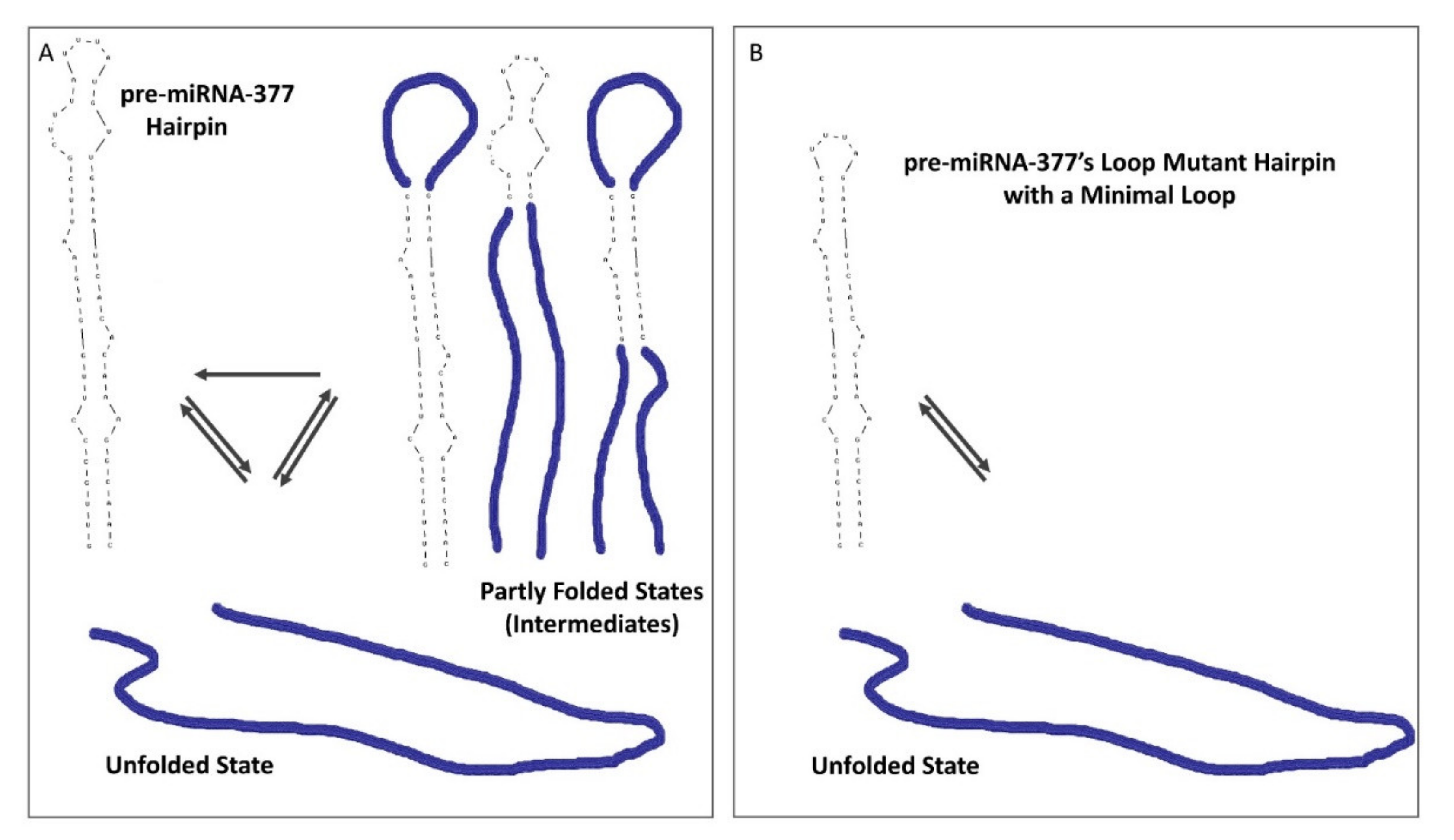
2.4. Loop Mutant of the Pre-miRNA-377
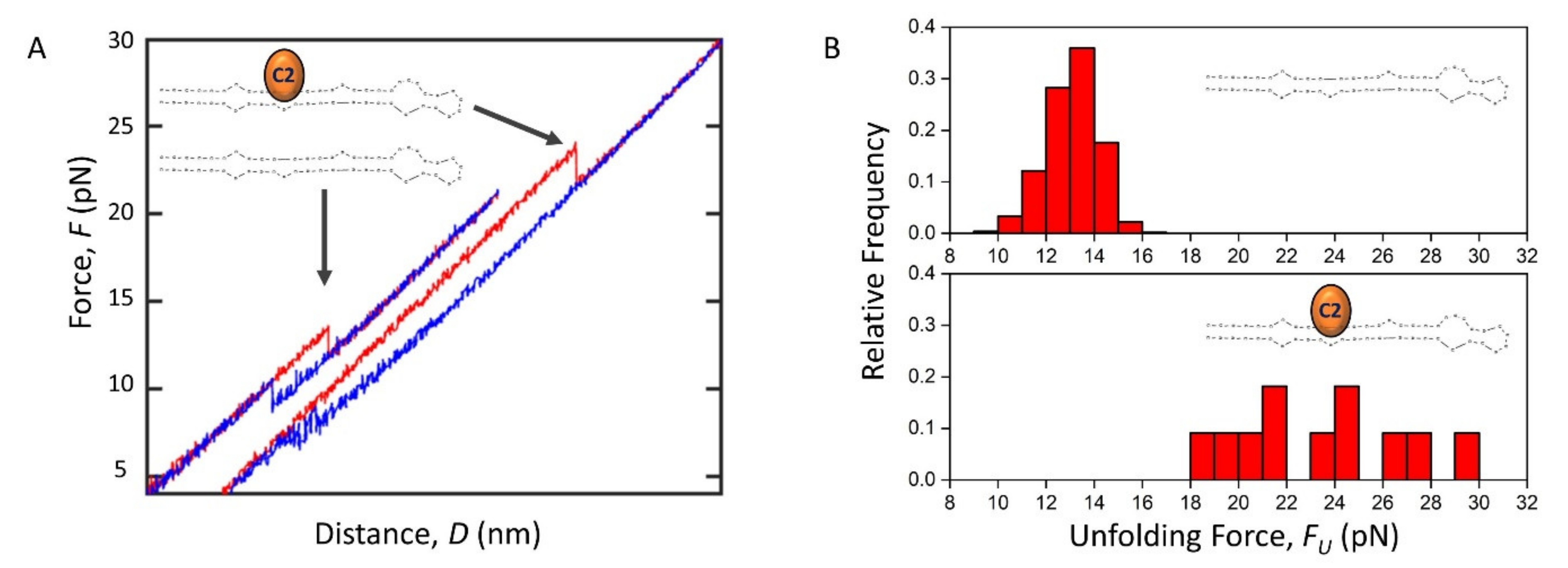
2.5. The Effect of C2 Ligand Binding on Pre-miRNA-377 Unfolding and Folding
3. Discussion
4. Materials and Methods
4.1. Pre-miRNA-377 Constructs
4.2. Buffers
4.3. Optical Tweezers Measurements of Pre-miRNA-377
4.4. Optical Tweezers Measurements of the C2 Ligand Binding to the Pre-miRNA-377
4.5. Analysis of Optical Tweezers Data
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 5′ UTR | 5′ untranslated region |
| ΔF | force difference |
| ΔG | free energy |
| dsDNA | double-stranded DNA |
| E | extension |
| EF | folding extension |
| EU | unfolding extension |
| F | force |
| FDC | force–distance curve |
| FF | folding force |
| FU | unfolding force |
| IRE | iron-responsive element |
| KD | dissociation constant |
| miRNA | microRNA |
| mRNA | messenger RNA |
| pre-miRNA | pre-microRNA |
| PRF | programmed ribosomal frameshifting |
| RNAi | RNA interference |
| RT | room temperature |
| ssDNA | single-stranded DNA |
| Tm | melting temperature |
| TRAIL | tumor necrosis factor (TNF)-related apoptosis-inducing ligand |
| VEGFA | vascular endothelial growth factor A |
| WT | wild type |
References
- Ha, T.Y. MicroRNAs in Human Diseases: From Autoimmune Diseases to Skin, Psychiatric and Neurodegenerative Diseases. Immune Netw. 2011, 11, 227–244. [Google Scholar] [CrossRef]
- Kowalska, J.; Wypijewska del Nogal, A.; Darzynkiewicz, Z.M.; Buck, J.; Nicola, C.; Kuhn, A.N.; Lukaszewicz, M.; Zuberek, J.; Strenkowska, M.; Ziemniak, M.; et al. Synthesis, properties, and biological activity of boranophosphate analogs of the mRNA cap: Versatile tools for manipulation of therapeutically relevant cap-dependent processes. Nucleic Acids Res. 2014, 42, 10245–10264. [Google Scholar] [CrossRef] [Green Version]
- Wypijewska del Nogal, A.; Surleac, M.; Kowalska, J.; Lukaszewicz, M.; Jemielity, J.; Bisaillon, M.; Darzynkiewicz, E.; Milac, A.; Bojarska, E. Analysis of Decapping Scavenger (DcpS)-cap complex using modified cap analogs reveals molecular determinants for efficient cap binding. FEBS J. 2013, 280, 6508. [Google Scholar] [CrossRef]
- Joladarashi, D.; Garikipati, V.N.S.; Thandavarayan, R.A.; Verma, S.K.; Mackie, A.R.; Khan, M.; Gumpert, A.M.; Bhimaraj, A.; Youker, K.A.; Uribe, C.; et al. Enhanced Cardiac Regenerative Ability of Stem Cells After Ischemia-Reperfusion Injury: Role of Human CD34+ Cells Deficient in MicroRNA-377. J. Am. Coll. Cardiol. 2015, 66, 2214–2226. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.D.; Chi, J.Y.; Liang, H.H.; Huangfu, L.T.; Guo, Z.D.; Zou, H.; Yin, X.H. MicroRNA-377 Mediates Cardiomyocyte Apoptosis Induced by Cyclosporin A. Can. J. Cardiol. 2016, 32, 1249–1259. [Google Scholar] [CrossRef]
- Haniff, H.S.; Knerr, L.; Liu, X.; Crynen, G.; Boström, J.; Abegg, D.; Adibekian, A.; Lekah, E.; Wang, K.W.; Cameron, M.D.; et al. Design of a small molecule that stimulates vascular endothelial growth factor A enabled by screening RNA fold–small molecule interactions. Nat. Chem. 2020, 12, 952–961. [Google Scholar] [CrossRef]
- Wen, Z.; Huang, W.; Feng, Y.; Cai, W.; Wang, Y.; Wang, X.; Liang, J.; Wani, M.; Chen, J.; Zhu, P.; et al. MicroRNA-377 regulates mesenchymal stem cell-induced angiogenesis in ischemic hearts by targeting VEGF. PLoS ONE 2014, 9, e104666. [Google Scholar] [CrossRef] [PubMed]
- Cartland, S.P.; Lin, R.C.Y.; Genner, S.; Patil, M.S.; Martinez, G.J.; Barraclough, J.Y.; Gloss, B.; Misra, A.; Patel, S.; Kavurma, M.M. Vascular transcriptome landscape of Trail(−/−) mice: Implications and therapeutic strategies for diabetic vascular disease. FASEB J. 2020, 34, 9547–9562. [Google Scholar] [CrossRef] [PubMed]
- Metra, M.; Teerlink, J.R. Heart failure. Lancet 2017, 390, 1981–1995. [Google Scholar] [CrossRef]
- Creemers, E.E.; Wilde, A.A.; Pinto, Y.M. Heart failure: Advances through genomics. Nat. Rev. Genet. 2011, 12, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Wronska, A.; Kurkowska-Jastrzebska, I.; Santulli, G. Application of microRNAs in diagnosis and treatment of cardiovascular disease. Acta Physiol. 2015, 213, 60–83. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, M.F. Drug target miRNAs: Chances and challenges. Trends Biotechnol. 2014, 32, 578–585. [Google Scholar] [CrossRef] [PubMed]
- Fan, R.; Xiao, C.; Wan, X.; Cha, W.; Miao, Y.; Zhou, Y.; Qin, C.; Cui, T.; Su, F.; Shan, X. Small molecules with big roles in microRNA chemical biology and microRNA-targeted therapeutics. RNA Biol. 2019, 16, 707–718. [Google Scholar] [CrossRef] [PubMed]
- Velagapudi, S.P.; Gallo, S.M.; Disney, M.D. Sequence-based design of bioactive small molecules that target precursor microRNAs. Nat. Chem. Biol. 2014, 10, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, D.B.; Woodside, M.T. Probing the structural dynamics of proteins and nucleic acids with optical tweezers. Curr. Opin. Struct. Biol. 2015, 34, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Taylor, K.; Sobczak, K. Intrinsic Regulatory Role of RNA Structural Arrangement in Alternative Splicing Control. Int. J. Mol. Sci. 2020, 21, 5161. [Google Scholar] [CrossRef]
- Dethoff, E.A.; Chugh, J.; Mustoe, A.M.; Al-Hashimi, H.M. Functional complexity and regulation through RNA dynamics. Nature 2012, 482, 322–330. [Google Scholar] [CrossRef] [Green Version]
- Liphardt, J.; Onoa, B.; Smith, S.B.; Tinoco, I., Jr.; Bustamante, C. Reversible unfolding of single RNA molecules by mechanical force. Science 2001, 292, 733–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ritchie, D.B.; Cappellano, T.R.; Tittle, C.; Rezajooei, N.; Rouleau, L.; Sikkema, W.K.A.; Woodside, M.T. Conformational dynamics of the frameshift stimulatory structure in HIV-1. RNA 2017, 23, 1376–1384. [Google Scholar] [CrossRef]
- Neupane, K.; Yu, H.; Foster, D.A.; Wang, F.; Woodside, M.T. Single-molecule force spectroscopy of the add adenine riboswitch relates folding to regulatory mechanism. Nucleic Acids Res. 2011, 39, 7677–7687. [Google Scholar] [CrossRef]
- Ritchie, D.B.; Foster, D.A.; Woodside, M.T. Programmed -1 frameshifting efficiency correlates with RNA pseudoknot conformational plasticity, not resistance to mechanical unfolding. Proc. Natl. Acad. Sci. USA 2012, 109, 16167–16172. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.; Zhang, X.; Graves, P.; Zeng, Y. A comprehensive analysis of precursor microRNA cleavage by human Dicer. RNA 2012, 18, 2083–2092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Q.J.; Zhang, J.; Li, P.; Wang, Q.; Zhang, Y.; Roy-Chaudhuri, B.; Xu, J.; Kay, M.A.; Zhang, Q.C. RNA structure probing reveals the structural basis of Dicer binding and cleavage. Nat. Commun. 2021, 12, 3397. [Google Scholar] [CrossRef] [PubMed]
- Bood, M.; Wypijewska del Nogal, A.; Nilsson, J.R.; Edfeldt, F.; Dahlen, A.; Lemurell, M.; Wilhelmsson, L.M.; Grotli, M. Interbase-FRET binding assay for pre-microRNAs. Sci. Rep. 2021, 11, 9396. [Google Scholar] [CrossRef]
- Forns, N.; de Lorenzo, S.; Manosas, M.; Hayashi, K.; Huguet, J.M.; Ritort, F. Improving signal/noise resolution in single-molecule experiments using molecular constructs with short handles. Biophys. J. 2011, 100, 1765–1774. [Google Scholar] [CrossRef] [Green Version]
- Alemany, A.; Ritort, F. Determination of the elastic properties of short ssDNA molecules by mechanically folding and unfolding DNA hairpins. Biopolymers 2014, 101, 1193–1199. [Google Scholar] [CrossRef] [PubMed]
- Landuzzi, F.; Viader-Godoy, X.; Cleri, F.; Pastor, I.; Ritort, F. Detection of single DNA mismatches by force spectroscopy in short DNA hairpins. J. Chem. Phys. 2020, 152, 074204. [Google Scholar] [CrossRef] [PubMed]
- Sundar Rajan, V.; Viader-Godoy, X.; Lin, Y.L.; Dutta, U.; Ritort, F.; Westerlund, F.; Wilhelmsson, L.M. Mechanical characterization of base analogue modified nucleic acids by force spectroscopy. Phys. Chem. Chem. Phys. 2021, 23, 14151–14155. [Google Scholar] [CrossRef]
- Bizarro, C.V.; Alemany, A.; Ritort, F. Non-specific binding of Na+ and Mg2+ to RNA determined by force spectroscopy methods. Nucleic Acids Res. 2012, 40, 6922–6935. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.; Wen, J.D.; Tinoco, I., Jr. Single-molecule mechanical unfolding and folding of a pseudoknot in human telomerase RNA. RNA 2007, 13, 2175–2188. [Google Scholar] [CrossRef] [Green Version]
- Li, P.T.; Collin, D.; Smith, S.B.; Bustamante, C.; Tinoco, I., Jr. Probing the mechanical folding kinetics of TAR RNA by hopping, force-jump, and force-ramp methods. Biophys. J. 2006, 90, 250–260. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.; Chang, K.Y.; Chou, M.Y.; Bustamante, C.; Tinoco, I., Jr. Triplex structures in an RNA pseudoknot enhance mechanical stability and increase efficiency of -1 ribosomal frameshifting. Proc. Natl. Acad. Sci. USA 2009, 106, 12706–12711. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Zhong, Z.; Tong, C.; Jia, H.; Liu, Y.; Chen, G. Single-Molecule Mechanical Folding and Unfolding of RNA Hairpins: Effects of Single A-U to A.C Pair Substitutions and Single Proton Binding and Implications for mRNA Structure-Induced −1 Ribosomal Frameshifting. J. Am. Chem. Soc. 2018, 140, 8172–8184. [Google Scholar] [CrossRef] [PubMed]
- Li, P.T.; Bustamante, C.; Tinoco, I., Jr. Unusual mechanical stability of a minimal RNA kissing complex. Proc. Natl. Acad. Sci. USA 2006, 103, 15847–15852. [Google Scholar] [CrossRef] [Green Version]
- Manosas, M.; Collin, D.; Ritort, F. Force-dependent fragility in RNA hairpins. Phys. Rev. Lett. 2006, 96, 218301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, X.; Li, H. Force spectroscopy studies on protein-ligand interactions: A single protein mechanics perspective. FEBS Lett. 2014, 588, 3613–3620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yangyuoru, P.M.; Di Antonio, M.; Ghimire, C.; Biffi, G.; Balasubramanian, S.; Mao, H. Dual binding of an antibody and a small molecule increases the stability of TERRA G-quadruplex. Angew. Chem. Int. Ed. Engl. 2015, 54, 910–913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, P.T.X.; Vieregg, J.; Ignacio Tinoco, J. How RNA Unfolds and Refolds. Annu. Rev. Biochem. 2008, 77, 77–100. [Google Scholar] [CrossRef] [Green Version]
- Sharp, K.A.; Honig, B. Salt effects on nucleic acids. Curr. Opin. Struct. Biol. 1995, 5, 323–328. [Google Scholar] [CrossRef]
- Buck, J.; Noeske, J.; Wohnert, J.; Schwalbe, H. Dissecting the influence of Mg2+ on 3D architecture and ligand-binding of the guanine-sensing riboswitch aptamer domain. Nucleic Acids Res. 2010, 38, 4143–4153. [Google Scholar] [CrossRef]
- Mirihana Arachchilage, G.; Dassanayake, A.C.; Basu, S. A potassium ion-dependent RNA structural switch regulates human pre-miRNA 92b maturation. Chem. Biol. 2015, 22, 262–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woodson, S.A. Metal ions and RNA folding: A highly charged topic with a dynamic future. Curr. Opin. Chem. Biol. 2005, 9, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Valiyaveetil, F.I.; Leonetti, M.; Muir, T.W.; Mackinnon, R. Ion selectivity in a semisynthetic K+ channel locked in the conductive conformation. Science 2006, 314, 1004–1007. [Google Scholar] [CrossRef]
- Foloppe, N.; Matassova, N.; Aboul-Ela, F. Towards the discovery of drug-like RNA ligands? Drug Discov. Today 2006, 11, 1019–1027. [Google Scholar] [CrossRef]
- Woodside, M.T.; Behnke-Parks, W.M.; Larizadeh, K.; Travers, K.; Herschlag, D.; Block, S.M. Nanomechanical measurements of the sequence-dependent folding landscapes of single nucleic acid hairpins. Proc. Natl. Acad. Sci. USA 2006, 103, 6190–6195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woodside, M.T.; Anthony, P.C.; Behnke-Parks, W.M.; Larizadeh, K.; Herschlag, D.; Block, S.M. Direct measurement of the full, sequence-dependent folding landscape of a nucleic acid. Science 2006, 314, 1001–1004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neupane, K.; Manuel, A.P.; Lambert, J.; Woodside, M.T. Transition-Path Probability as a Test of Reaction-Coordinate Quality Reveals DNA Hairpin Folding Is a One-Dimensional Diffusive Process. J. Phys. Chem. Lett. 2015, 6, 1005–1010. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Park, H.J.; Zhang, J.; Junn, E.; Andrews, R.J.; Velagapudi, S.P.; Abegg, D.; Vishnu, K.; Costales, M.G.; Childs-Disney, J.L.; et al. Translation of the intrinsically disordered protein alpha-synuclein is inhibited by a small molecule targeting its structured mRNA. Proc. Natl. Acad. Sci. USA 2020, 117, 1457–1467. [Google Scholar] [CrossRef] [Green Version]
- Wypijewska del Nogal, A.; Fuchtbauer, A.F.; Bood, M.; Nilsson, J.R.; Wranne, M.S.; Sarangamath, S.; Pfeiffer, P.; Rajan, V.S.; El-Sagheer, A.H.; Dahlen, A.; et al. Getting DNA and RNA out of the dark with 2CNqA: A bright adenine analogue and interbase FRET donor. Nucleic Acids Res. 2020, 48, 7640–7652. [Google Scholar] [CrossRef]
- Laurent, V.M.; Duperray, A.; Sundar Rajan, V.; Verdier, C. Atomic force microscopy reveals a role for endothelial cell ICAM-1 expression in bladder cancer cell adherence. PLoS ONE 2014, 9, e98034. [Google Scholar] [CrossRef]
- Sundar Rajan, V.; Laurent, V.M.; Verdier, C.; Duperray, A. Unraveling the Receptor-Ligand Interactions between Bladder Cancer Cells and the Endothelium Using AFM. Biophys. J. 2017, 112, 1246–1257. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Buffer | FDC | Nb | nc | Unfolding | Folding | ΔF (pN) d | ||
|---|---|---|---|---|---|---|---|---|
| FU (pN) | EU (nm) | FF (pN) | EF (nm) | |||||
| High-Salt | Type 1 | 11/11 | 765 | 14.1 ± 0.3 | 20.8 ± 0.3 | 10.1 ± 0.3 | 21.0 ± 0.5 | 3.9 ± 0.4 |
| Type 2 | 8/11 | 42 | - | - | 9.2 ± 0.3 | 18.2 ± 1.3 | 4.8 ± 0.3 | |
| Na+/Mg2+ | Type 1 | 13/14 | 545 | 13.4 ± 0.4 | 20.9 ± 0.6 | 8.9 ± 0.2 | 20.5 ± 0.7 | 4.2 ± 0.4 |
| Type 2 | 13/14 | 246 | - | - | 8.1 ± 0.3 | 16.3 ± 1.0 | 5.5 ± 0.5 | |
| Type 3 | 3/14 | 71 | - | - | 7.1 ± 0.5 | - | 5.7 ± 0.9 | |
| K+/Mg2+ | Type 1 | 8/8 | 453 | 12.7 ± 0.4 | 20.2 ± 0.5 | 9.0 ± 0.3 | 20.1 ± 0.7 | 3.5 ± 0.4 |
| Type 2 | 8/8 | 216 | - | - | 8.1 ± 0.4 | 16.9 ± 1.1 | 5.0 ± 0.4 | |
| Na+ | Type 1 | 5/5 | 238 | 11.0 ± 0.6 | 20.0 ± 0.6 | 7.4 ± 0.5 | 19.6 ± 0.9 | 3.3 ± 0.6 |
| Type 2 | 5/5 | 91 | - | - | 7.2 ± 0.4 | 16.7 ± 1.5 | 4.3 ± 0.4 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wypijewska del Nogal, A.; Sundar Rajan, V.; Westerlund, F.; Wilhelmsson, L.M. Complex Conformational Dynamics of the Heart Failure-Associated Pre-miRNA-377 Hairpin Revealed by Single-Molecule Optical Tweezers. Int. J. Mol. Sci. 2021, 22, 9008. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22169008
Wypijewska del Nogal A, Sundar Rajan V, Westerlund F, Wilhelmsson LM. Complex Conformational Dynamics of the Heart Failure-Associated Pre-miRNA-377 Hairpin Revealed by Single-Molecule Optical Tweezers. International Journal of Molecular Sciences. 2021; 22(16):9008. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22169008
Chicago/Turabian StyleWypijewska del Nogal, Anna, Vinoth Sundar Rajan, Fredrik Westerlund, and L. Marcus Wilhelmsson. 2021. "Complex Conformational Dynamics of the Heart Failure-Associated Pre-miRNA-377 Hairpin Revealed by Single-Molecule Optical Tweezers" International Journal of Molecular Sciences 22, no. 16: 9008. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22169008