
Cappable-Seq Reveals Specific Patterns of Metabolism and Virulence for Salmonella Typhimurium Intracellular Survival within Acanthamoeba castellanii

 , ,
, ,  and
and
Abstract
:1. Introduction
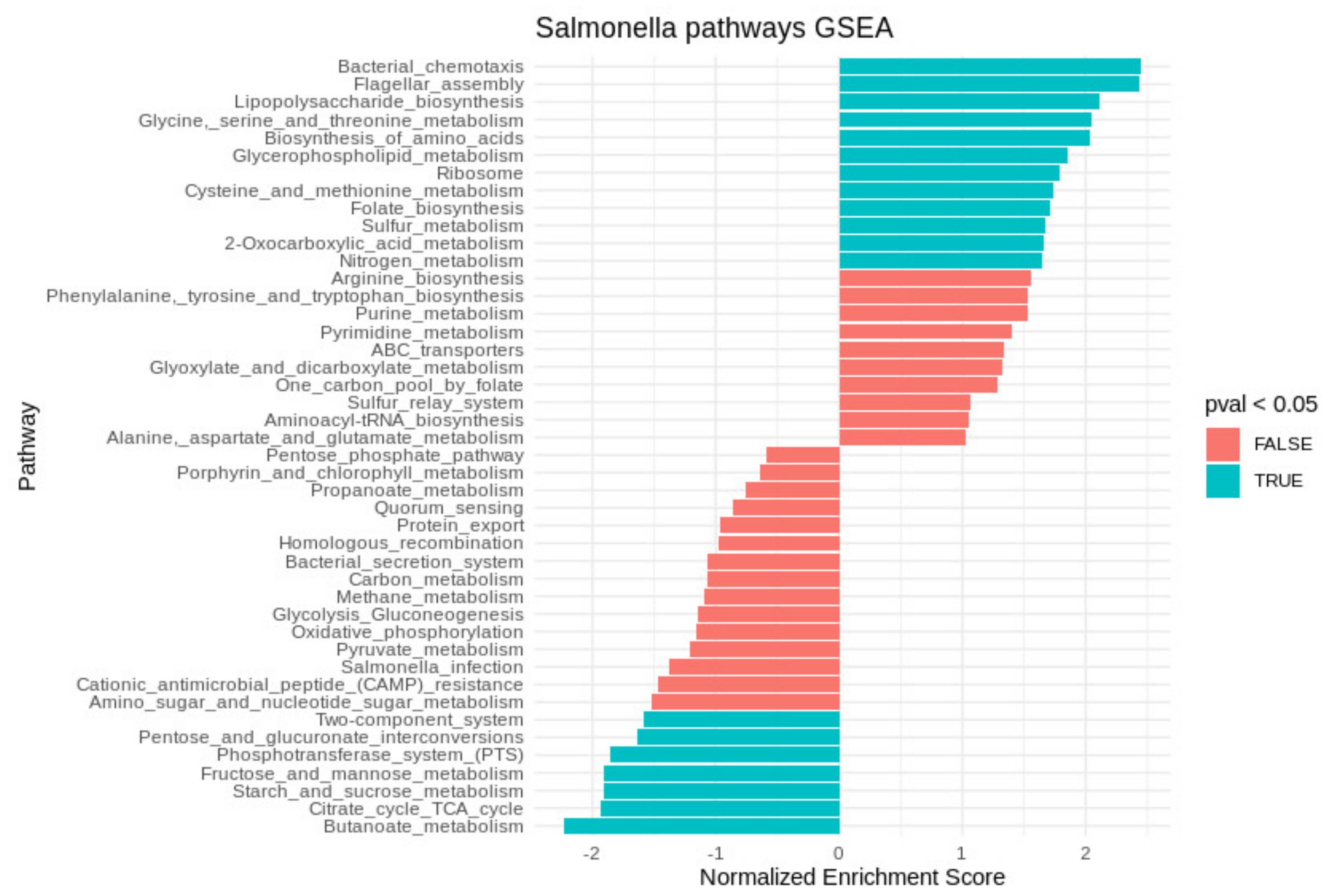
2. Results
2.1. Sequencing
2.2. Transcriptional Profile of Salmonella within the Acanthamoebae Phagosomes
2.3. Expression of Genes Associated with Iron Uptake and Stress Response
2.4. Expression of Genes Associated with Virulence
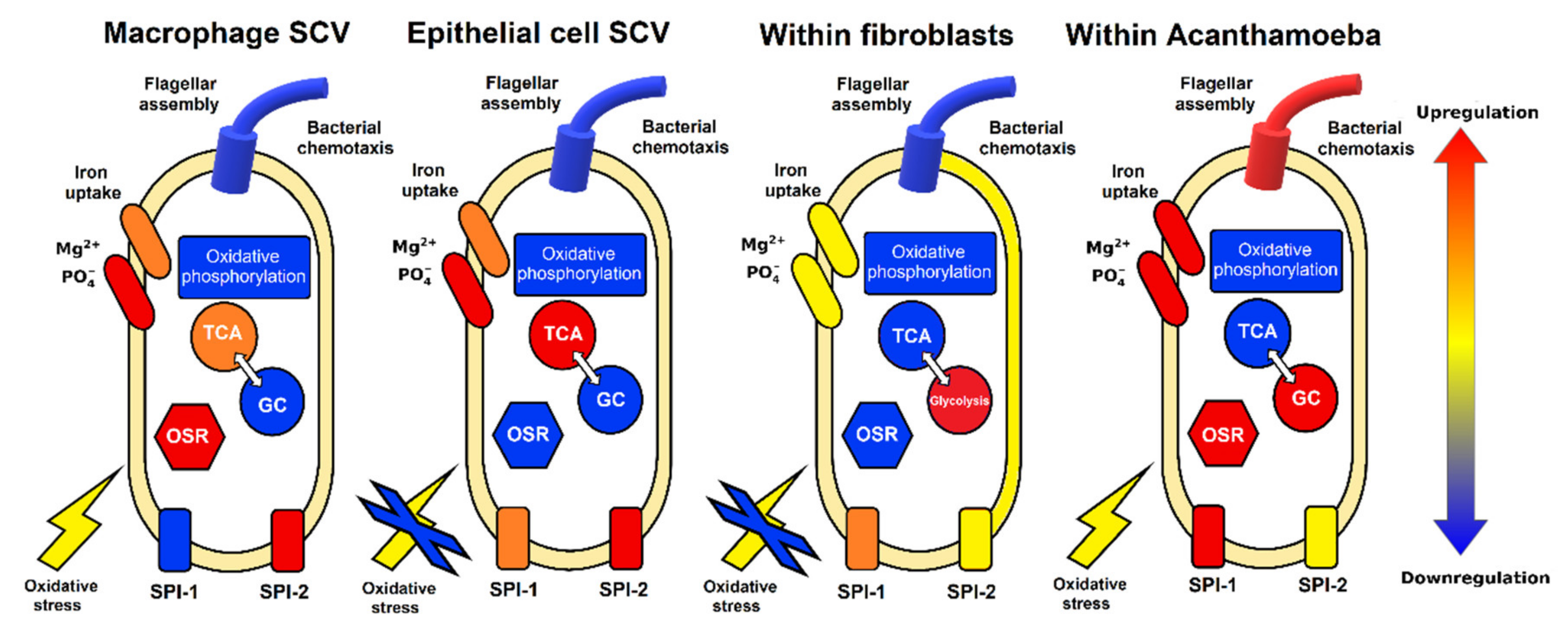
3. Discussion
3.1. Metabolic Pathways of Salmonella within Acanthamoeba SCV
3.2. Iron Uptake and Oxidative Stress
3.3. Virulence and Motility
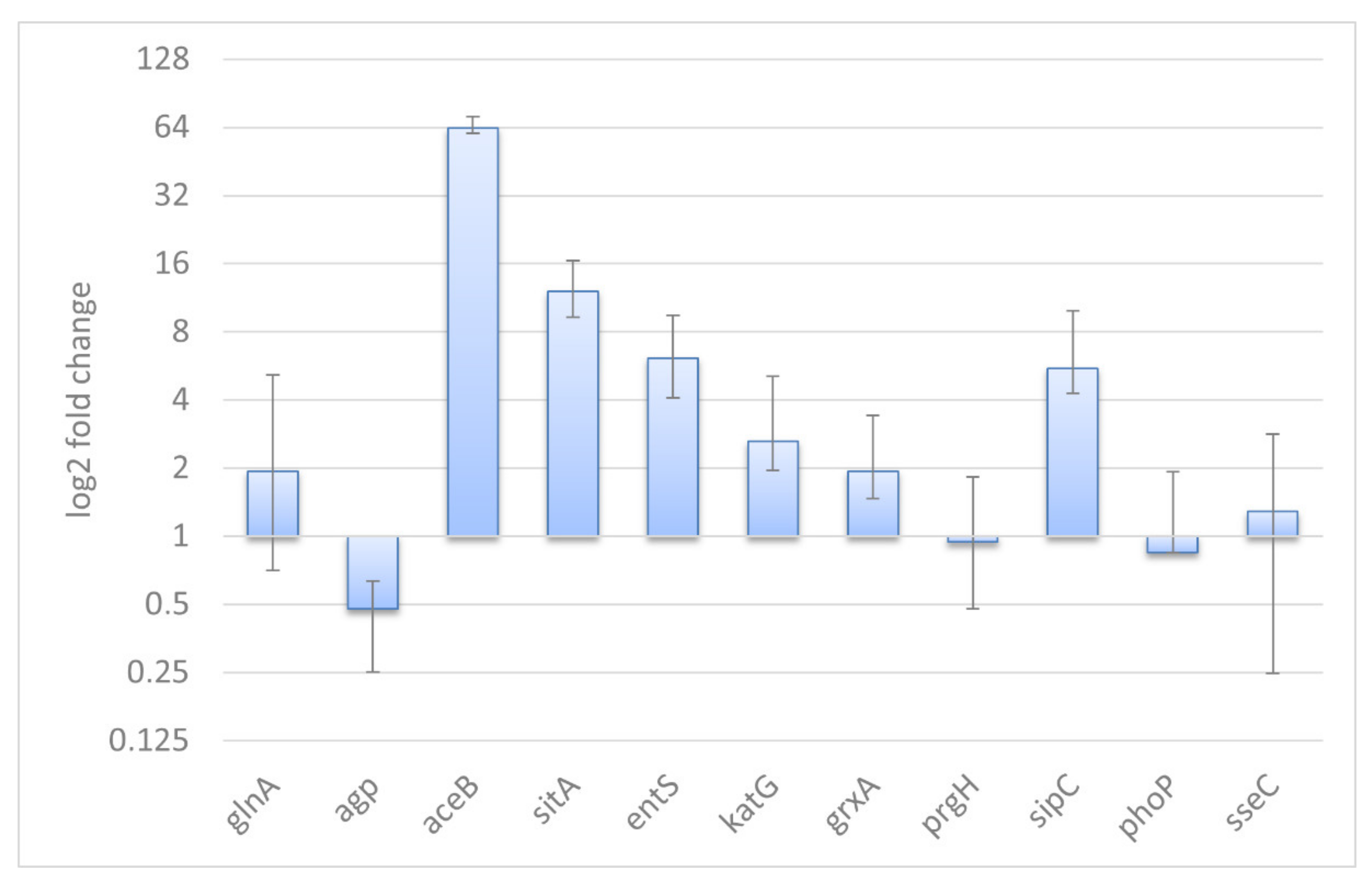
3.4. Validation of the RNA-Seq Results and General Remarks
4. Materials and Methods
4.1. Strains
4.2. Co-Culture Conditions
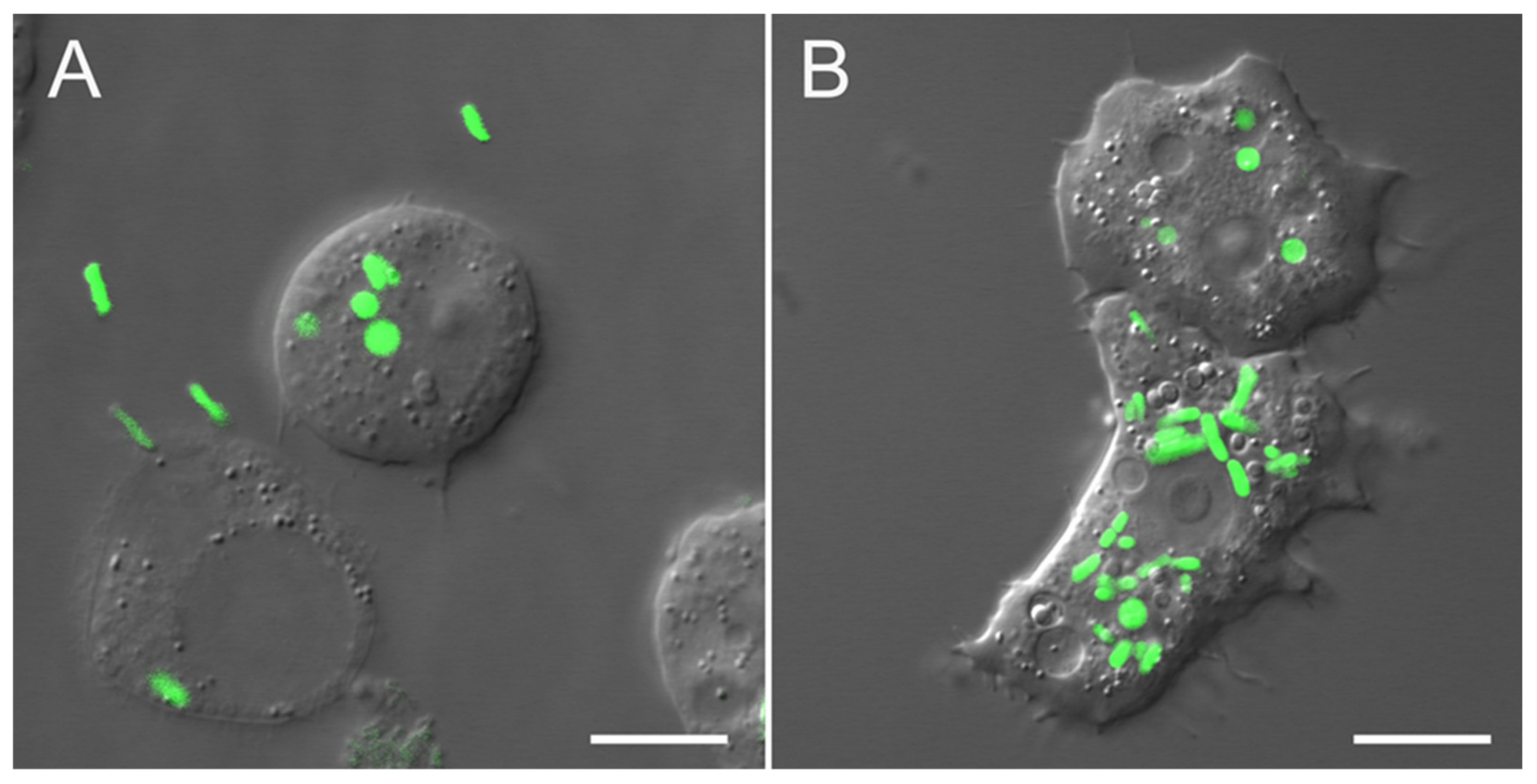
4.3. Microscopy
4.4. RNA Extraction and cDNA Library Preparation
4.5. qRT-PCR Analysis
4.6. Bioinformatic Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- European Food Safety Authority (EFSA); European Centre for Disease Prevention and Control (ECDC). The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2017. EFSA J. 2018, 16, e05500. [Google Scholar] [CrossRef]
- Vaerewijck, M.J.M.; Baré, J.; Lambrecht, E.; Sabbe, K.; Houf, K. Interactions of Foodborne Pathogens with Free-living Protozoa: Potential Consequences for Food Safety. Compr. Rev. Food Sci. Food Saf. 2014, 13, 924–944. [Google Scholar] [CrossRef]
- Balczun, C.; Scheid, P.L. Free-living amoebae as hosts for and vectors of intracellular microorganisms with public health significance. Viruses 2017, 9, 65. [Google Scholar] [CrossRef] [PubMed]
- Pace, M.L. Bacterial mortality and the fate of bacterial production. Hydrobiologia 1988, 159, 41–49. [Google Scholar] [CrossRef]
- Hahn, M.W.; Höfle, M.G. Grazing of protozoa and its effect on populations of aquatic bacteria. FEMS Microbiol. Ecol. 2001, 35, 113–121. [Google Scholar] [CrossRef]
- Denoncourt, A.M.; Paquet, V.E.; Charette, S.J. Potential role of bacteria packaging by protozoa in the persistence and transmission of pathogenic bacteria. Front. Microbiol. 2014, 5, 240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffmann, C.; Harrison, C.F.; Hilbi, H. The natural alternative: Protozoa as cellular models for Legionella infection. Cell. Microbiol. 2014, 16, 15–26. [Google Scholar] [CrossRef]
- Gaze, W.H.; Burroughs, N.; Gallagher, M.P.; Wellington, E.M.H. Interactions between Salmonella typhimurium and Acanthamoeba polyphaga, and Observation of a New Mode of Intracellular Growth within Contractile Vacuoles. Microb. Ecol. 2003, 46, 358–369. [Google Scholar] [CrossRef] [PubMed]
- Douesnard-Malo, F.; Daigle, F. Increased persistence of Salmonella enterica Serovar typhi in the presence of Acanthamoeba castellanii. Appl. Environ. Microbiol. 2011, 77, 7640–7646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno-Mesonero, L.; Ferrús, M.A.; Moreno, Y. Determination of the bacterial microbiome of free-living amoebae isolated from wastewater by 16S rRNA amplicon-based sequencing. Environ. Res. 2020, 190, e109987. [Google Scholar] [CrossRef]
- Tezcan-Merdol, D.; Ljungström, M.; Winiecka-Krusnell, J.; Linder, E.; Engstrand, L.; Rhen, M. Uptake and replication of Salmonella enterica in Acanthamoeba rhysodes. Appl. Environ. Microbiol. 2004, 70, 3706–3714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riquelme, S.; Varas, M.; Valenzuela, C.; Velozo, P.; Chahin, N.; Aguilera, P.; Sabag, A.; Labra, B.; Álvarez, S.A.; Chávez, F.P.; et al. Relevant genes linked to virulence are required for Salmonella typhimurium to survive intracellularly in the social amoeba Dictyostelium discoideum. Front. Microbiol. 2016, 7, 1305. [Google Scholar] [CrossRef] [PubMed]
- Gourabathini, P.; Brandi, M.T.; Redding, K.S.; Gunderson, J.H.; Berk, S.G. Interactions between food-borne pathogens and protozoa isolated from lettuce and spinach. Appl. Environ. Microbiol. 2008, 74, 2518–2525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rehfuss, M.Y.M.; Parker, C.T.; Brandl, M.T. Salmonella transcriptional signature in Tetrahymena phagosomes and role of acid tolerance in passage through the protist. ISME J. 2011, 5, 262–273. [Google Scholar] [CrossRef] [PubMed]
- Hautefort, I.; Thompson, A.; Eriksson-Ygberg, S.; Parker, M.L.; Lucchini, S.; Danino, V.; Bongaerts, R.J.M.; Ahmad, N.; Rhen, M.; Hinton, J.C.D. During infection of epithelial cells Salmonella enterica serovar Typhimurium undergoes a time-dependent transcriptional adaptation that results in simultaneous expression of three type 3 secretion systems. Cell. Microbiol. 2008, 10, 958–984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westermann, A.J.; Förstner, K.U.; Amman, F.; Barquist, L.; Chao, Y.; Schulte, L.N.; Müller, L.; Reinhardt, R.; Stadler, P.F.; Vogel, J. Dual RNA-seq unveils noncoding RNA functions in host-pathogen interactions. Nature 2016, 529, 496–501. [Google Scholar] [CrossRef]
- Srikumar, S.; Kröger, C.; Hébrard, M.; Colgan, A.; Owen, S.V.; Sivasankaran, S.K.; Cameron, A.D.S.; Hokamp, K.; Hinton, J.C.D. RNA-seq Brings New Insights to the Intra-Macrophage Transcriptome of Salmonella Typhimurium. PLoS Pathog. 2015, 11, e1005262. [Google Scholar] [CrossRef] [Green Version]
- Noster, J.; Chao, T.C.; Sander, N.; Schulte, M.; Reuter, T.; Hansmeier, N.; Hensel, M. Proteomics of intracellular Salmonella enterica reveals roles of Salmonella pathogenicity island 2 in metabolism and antioxidant defense. PLoS Pathog. 2019, 15, e1007741. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Liu, Y.; Fu, J.; Zhang, B.; Cheng, S.; Wu, M.; Wang, Z.; Jiang, J.; Chang, C.; Liu, X. Salmonella Proteomic Profiling during Infection Distinguishes the Intracellular Environment of Host Cells. mSystems 2019, 4, e00314-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Núñez-Hernández, C.; Tierrez, A.; Ortega, A.D.; Pucciarelli, M.G.; Godoy, M.; Eisman, B.; Casadesús, J.; García-del Portillo, F. Genome expression analysis of nonproliferating intracellular Salmonella enterica serovar Typhimurium unravels an acid pH-dependent PhoP-PhoQ response essential for dormancy. Infect. Immun. 2013, 81, 154–165. [Google Scholar] [CrossRef] [Green Version]
- Bleasdale, B.; Lott, P.J.; Jagannathan, A.; Stevens, M.P.; Birtles, R.J.; Wigley, P. The Salmonella pathogenicity island 2-encoded type III secretion system is essential for the survival of Salmonella enterica serovar typhimurium in free-living amoebae. Appl. Environ. Microbiol. 2009, 75, 1793–1795. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.; Hsiao, Y.H.; Chen, H.L.; Chu, C.; Tang, P.; Chiu, C.H. Apoptosis-like cell death induced by Salmonella in Acanthamoeba rhysodes. Genomics 2009, 94, 132–137. [Google Scholar] [CrossRef] [Green Version]
- Sharma, C.M.; Hoffmann, S.; Darfeuille, F.; Reignier, J.; Findeiß, S.; Sittka, A.; Chabas, S.; Reiche, K.; Hackermüller, J.; Reinhardt, R.; et al. The primary transcriptome of the major human pathogen Helicobacter pylori. Nature 2010, 464, 250–255. [Google Scholar] [CrossRef] [PubMed]
- Ettwiller, L.; Buswell, J.; Yigit, E.; Schildkraut, I. A novel enrichment strategy reveals unprecedented number of novel transcription start sites at single base resolution in a model prokaryote and the gut microbiome. BMC Genom. 2016, 17, 199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bjarnason, J.; Southward, C.M.; Surette, M.G. Genomic profiling of iron-responsive genes in Salmonella enterica serovar Typhimurium by high-throughput screening of a random promoter library. J. Bacteriol. 2003, 185, 4973–4982. [Google Scholar] [CrossRef] [Green Version]
- Johnson, R.; Byrne, A.; Berger, C.N.; Klemm, E.; Crepin, V.F.; Dougan, G.; Frankel, G. The type III secretion system effector SptP of Salmonella enterica serovar Typhi. J. Bacteriol. 2017, 199, e00647-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirold, S.; Ehrbar, K.; Weissmüller, A.; Prager, R.; Tschäpe, H.; Rüssmann, H.; Hardt, W.D. Salmonella host cell invasion emerged by acquisition of a mosaic of separate genetic elements, including Salmonella pathogenicity island 1 (SPI1), SPI5, and sopE2. J. Bacteriol. 2001, 183, 2348–2358. [Google Scholar] [CrossRef] [Green Version]
- Eriksson, S.; Lucchini, S.; Thompson, A.; Rhen, M.; Hinton, J.C.D. Unravelling the biology of macrophage infection by gene expression profiling of intracellular Salmonella enterica. Mol. Microbiol. 2003, 47, 103–118. [Google Scholar] [CrossRef] [PubMed]
- Steeb, B.; Claudi, B.; Burton, N.A.; Tienz, P.; Schmidt, A.; Farhan, H.; Mazé, A.; Bumann, D. Parallel Exploitation of Diverse Host Nutrients Enhances Salmonella Virulence. PLoS Pathog. 2013, 9, e1003301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diacovich, L.; Lorenzi, L.; Tomassetti, M.; Méresse, S.; Gramajo, H. The infectious intracellular lifestyle of salmonella enterica relies on the adaptation to nutritional conditions within the salmonella-containing vacuole. Virulence 2017, 8, 975–992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, F.C.; Libby, S.J.; Castor, M.E.; Fung, A.M. Isocitrate lyase (AceA) is required for Salmonella persistence but not for acute lethal infection in mice. Infect. Immun. 2005, 73, 2547–2549. [Google Scholar] [CrossRef] [Green Version]
- Ahn, S.; Jung, J.; Jang, I.A.; Madsen, E.L.; Park, W. Role of glyoxylate shunt in oxidative stress response. J. Biol. Chem. 2016, 291, 11928–11938. [Google Scholar] [CrossRef] [Green Version]
- Fitzsimmons, L.; Liu, L.; Porwollik, S.; Chakraborty, S.; Desai, P.; Tapscott, T.; Henard, C.; McClelland, M.; Vazquez-Torres, A. Zinc-dependent substrate-level phosphorylation powers Salmonella growth under nitrosative stress of the innate host response. PLoS Pathog. 2018, 14, e1007388. [Google Scholar] [CrossRef]
- Frawley, E.R.; Crouch, M.L.V.; Bingham-Ramos, L.K.; Robbins, H.F.; Wang, W.; Wright, G.D.; Fang, F.C. Iron and citrate export by a major facilitator superfamily pump regulates metabolism and stress resistance in Salmonella Typhimurium. Proc. Natl. Acad. Sci. USA 2013, 110, 12054–12059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dandekar, T.; Fieselmann, A.; Fischer, E.; Popp, J.; Hensel, M.; Noster, J. Salmonella-how a metabolic generalist adopts an intracellular lifestyle during infection. Front. Cell. Infect. Microbiol. 2014, 4. [Google Scholar] [CrossRef] [Green Version]
- Turnbull, A.L.; Surette, M.G. Cysteine biosynthesis, oxidative stress and antibiotic resistance in Salmonella typhimurium. Res. Microbiol. 2010, 161, 643–650. [Google Scholar] [CrossRef]
- Slonczewski, J. Stress Responses: pH. In Encyclopedia of Microbiology, 3rd ed.; Konopka, A., Ed.; Academic Press: Cambridge, MA, USA, 2009; Volume 3, pp. 477–484. [Google Scholar]
- Dalebroux, Z.D.; Matamouros, S.; Whittington, D.; Bishop, R.E.; Miller, S.I. PhoPQ regulates acidic glycerophospholipid content of the Salmonella Typhimurium outer membrane. Proc. Natl. Acad. Sci. USA 2014, 111, 1963–1968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karlinsey, J.E.; Stepien, T.A.; Mayho, M.; Singletary, L.A.; Bingham-Ramos, L.K.; Brehm, M.A.; Greiner, D.L.; Shultz, L.D.; Gallagher, L.A.; Bawn, M.; et al. Genome-wide Analysis of Salmonella enterica serovar Typhi in Humanized Mice Reveals Key Virulence Features. Cell Host Microbe 2019, 26, 426–434.e6. [Google Scholar] [CrossRef]
- Schaible, U.E.; Kaufmann, S.H.E. Iron and microbial infection. Nat. Rev. Microbiol. 2004, 2, 946–953. [Google Scholar] [CrossRef] [PubMed]
- Litwin, C.M.; Calderwood, S.B. Role of iron in regulation of virulence genes. Clin. Microbiol. Rev. 1993, 6, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Khajanchi, B.K.; Xu, J.; Grim, C.J.; Ottesen, A.R.; Ramachandran, P.; Foley, S.L. Global transcriptomic analyses of Salmonella enterica in Iron-depleted and iron-rich growth conditions. BMC Genom. 2019, 20, 490. [Google Scholar] [CrossRef] [Green Version]
- Crouch, M.L.V.; Castor, M.; Karlinsey, J.E.; Kalhorn, T.; Fang, F.C. Biosynthesis and IroC-dependent export of the siderophore salmochelin are essential for virulence of Salmonella enterica serovar Typhimurium. Mol. Microbiol. 2008, 67, 971–983. [Google Scholar] [CrossRef] [PubMed]
- Nairz, M.; Ferring-Appel, D.; Casarrubea, D.; Sonnweber, T.; Viatte, L.; Schroll, A.; Haschka, D.; Fang, F.C.; Hentze, M.W.; Weiss, G.; et al. Iron regulatory proteins mediate host resistance to Salmonella infection. Cell Host Microbe 2015, 18, 254–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ray, P.D.; Huang, B.W.; Tsuji, Y. Reactive oxygen species (ROS) homeostasis and redox regulation in cellular signaling. Cell. Signal. 2012, 24, 981–990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunn, J.D.; Bosmani, C.; Barisch, C.; Raykov, L.; Lefrançois, L.H.; Cardenal-Muñoz, E.; López-Jiménez, A.T.; Soldati, T. Eat prey, live: Dictyostelium discoideum as a model for cell-autonomous defenses. Front. Immunol. 2018, 8, 1906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ezraty, B.; Gennaris, A.; Barras, F.; Collet, J.F. Oxidative stress, protein damage and repair in bacteria. Nat. Rev. Microbiol. 2017, 15, 385–396. [Google Scholar] [CrossRef]
- Zheng, J.; Cui, S.; Meng, J. Effect of transcriptional activators RamA and SoxS on expression of multidrug efflux pumps AcrAB and AcrEF in fluoroquinolone-resistant Salmonella Typhimurium. J. Antimicrob. Chemother. 2009, 63, 95–102. [Google Scholar] [CrossRef] [Green Version]
- McLean, S.; Bowman, L.A.H.; Poole, R.K. KatG from Salmonella Typhimurium is a peroxynitritase. FEBS Lett. 2010, 584, 1628–1632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Festa, R.A.; Thiele, D.J. Copper at the Front Line of the Host-Pathogen Battle. PLoS Pathog. 2012, 8, e1002887. [Google Scholar] [CrossRef] [Green Version]
- Kapetanovic, R.; Bokil, N.J.; Achard, M.E.S.; Ong, C.L.Y.; Peters, K.M.; Stocks, C.J.; Phan, M.D.; Monteleone, M.; Schroder, K.; Irvine, K.M.; et al. Salmonella employs multiple mechanisms to subvert the TLR-inducible zinc-mediated antimicrobial response of human macrophages. FASEB J. 2016, 30, 1901–1912. [Google Scholar] [CrossRef] [Green Version]
- Hao, X.; Lüthje, F.L.; Qin, Y.; McDevitt, S.F.; Lutay, N.; Hobman, J.L.; Asiani, K.; Soncini, F.C.; German, N.; Zhang, S.; et al. Survival in amoeba—A major selection pressure on the presence of bacterial copper and zinc resistance determinants? Identification of a “copper pathogenicity island”. Appl. Microbiol. Biotechnol. 2015, 99, 5817–5824. [Google Scholar] [CrossRef] [PubMed]
- Rome, K.; Borde, C.; Taher, R.; Cayron, J.; Lesterlin, C.; Gueguen, E.; De Rosny, E.; Rodrigue, A. The Two-Component System ZraPSR Is a Novel ESR that Contributes to Intrinsic Antibiotic Tolerance in Escherichia coli. J. Mol. Biol. 2018, 430, 4971–4985. [Google Scholar] [CrossRef] [PubMed]
- Velayudhan, J.; Karlinsey, J.E.; Frawley, E.R.; Becker, L.A.; Nartea, M.; Fang, F.C. Distinct roles of the Salmonella enterica serovar typhimurium CyaY and YggX proteins in the biosynthesis and repair of iron-sulfur clusters. Infect. Immun. 2014, 82, 1390–1401. [Google Scholar] [CrossRef] [Green Version]
- Outten, F.W.; Djaman, O.; Storz, G. A suf operon requirement for Fe-S cluster assembly during iron starvation in Escherichia coli. Mol. Microbiol. 2004, 52, 861–872. [Google Scholar] [CrossRef] [PubMed]
- De la Cruz, M.A.; Pérez-Morales, D.; Palacios, I.J.; Fernández-Mora, M.; Calva, E.; Bustamante, V.H. The two-component system CpxR/A represses the expression of Salmonella virulence genes by affecting the stability of the transcriptional regulator HilD. Front. Microbiol. 2015, 6, 807. [Google Scholar] [CrossRef]
- Ong, S.Y.; Ng, F.L.; Badai, S.S.; Yuryev, A.; Alam, M. Analysis and construction of pathogenicity island regulatory pathways in Salmonella enterica serovar Typhi. J. Integr. Bioinform. 2010, 7, 145. [Google Scholar] [CrossRef] [Green Version]
- Lou, L.; Zhang, P.; Piao, R.; Wang, Y. Salmonella Pathogenicity Island 1 (SPI-1) and Its Complex Regulatory Network. Front. Cell. Infect. Microbiol. 2019, 9, 270. [Google Scholar] [CrossRef] [Green Version]
- Mangan, M.W.; Lucchini, S.; Cróinín, T.Ó.; Fitzgerald, S.; Hinton, J.C.D.; Dorman, C.J. Nucleoid-associated protein HU controls three regulons that coordinate virulence, response to stress and general physiology in Salmonella enterica serovar Typhimurium. Microbiology 2011, 157, 1075–1087. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Yi, L.; Zhang, J.; Sun, L.; Wen, W.; Zhang, C.; Wang, S. Functional analysis of superoxide dismutase of Salmonella typhimurium in serum resistance and biofilm formation. J. Appl. Microbiol. 2018, 125, 1526–1533. [Google Scholar] [CrossRef]
- Lee, Y.H.; Kim, S.; Helmann, J.D.; Kim, B.H.; Park, Y.K. RaoN, a small RNA encoded within Salmonella pathogenicity island-11, confers resistance to macrophage-induced stress. Microbiology 2013, 159, 1366–1378. [Google Scholar] [CrossRef] [Green Version]
- Löber, S.; Jäckel, D.; Kaiser, N.; Hensel, M. Regulation of Salmonella pathogenicity island 2 genes by independent environmental signals. Int. J. Med Microbiol. 2006, 296, 435–447. [Google Scholar] [CrossRef]
- Lim, S.; Kim, M.; Choi, J.; Ryu, S. A mutation in tdcA attenuates the virulence of Salmonella enterica serovar Typhimurium. Mol. Cells 2010, 29, 509–517. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Lim, S.; Kim, D.; Choy, H.E.; Ryu, S. A tdcA mutation reduces the invasive ability of Salmonella enterica serovar typhimurium. Mol. Cells 2009, 28, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Ibarra, J.A.; Knodler, L.A.; Sturdevant, D.E.; Virtaneva, K.; Carmody, A.B.; Fischer, E.R.; Porcella, S.F.; Steele-Mortimer, O. Induction of Salmonella pathogenicity island 1 under different growth conditions can affect Salmonella-host cell interactions in vitro. Microbiology 2010, 156, 1120–1133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maloy, S.R.; Nunn, W.D. Genetic regulation of the glyoxylate shunt in Escherichia coli K-12. J. Bacteriol. 1982, 149, 173–180. [Google Scholar] [CrossRef] [Green Version]
- Luck, A.N.; Slatko, B.E.; Foster, J.M. Removing the needle from the haystack: Enrichment of Wolbachia endosymbiont transcripts from host nematode RNA by Cappable-seqTM. PLoS ONE 2017, 12, e0173186. [Google Scholar] [CrossRef]
- Kopylova, E.; Noé, L.; Touzet, H. SortMeRNA: Fast and accurate filtering of ribosomal RNAs in metatranscriptomic data. Bioinformatics 2012, 28, 3211–3217. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Jarvik, T.; Smillie, C.; Groisman, E.A.; Ochman, H. Short-term signatures of evolutionary change in the Salmonella enterica serovar Typhimurium 14028 genome. J. Bacteriol. 2010, 192, 560–567. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- Boutard, M.; Ettwiller, L.; Cerisy, T.; Alberti, A.; Labadie, K.; Salanoubat, M.; Schildkraut, I.; Tolonen, A.C. Global repositioning of transcription start sites in a plant-fermenting bacterium. Nat. Commun. 2016, 7, 13783. [Google Scholar] [CrossRef]
- Quinlan, A.R.; Hall, I.M. BEDTools: A flexible suite of utilities for comparing genomic features. Bioinformatics 2010, 26, 841–842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, Y.; Smyth, G.K.; Shi, W. FeatureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [Green Version]
- Mao, X.; Ma, Q.; Zhou, C.; Chen, X.; Zhang, H.; Yang, J.; Mao, F.; Lai, W.; Xu, Y. DOOR 2.0: Presenting operons and their functions through dynamic and integrated views. Nucleic Acids Res. 2014, 42, D654–D659. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korotkevich, G.; Sukhov, V.; Budin, N.; Shpak, B.; Artyomov, M.; Sergushichev, A. Fast gene set enrichment analysis. bioRxiv 2016. [Google Scholar] [CrossRef] [Green Version]
- Huerta-Cepas, J.; Szklarczyk, D.; Forslund, K.; Cook, H.; Heller, D.; Walter, M.C.; Rattei, T.; Mende, D.R.; Sunagawa, S.; Kuhn, M.; et al. EGGNOG 4.5: A hierarchical orthology framework with improved functional annotations for eukaryotic, prokaryotic and viral sequences. Nucleic Acids Res. 2016, 44, D286–D293. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Operon | Log2 Fold Change | Description |
|---|---|---|
| Metal uptake | ||
| sufABCDSE | 2.76 | iron-sulfur cluster assembly |
| fhu | 2.65 | uptake of ferric hydroxamate |
| ent | 3.55 | enterobactin exporter |
| fep | 1.23 | ferric enterobactin transport protein |
| fes | 2.73 | ferric enterobactin esterase |
| corA | 1.65 | magnesium/nickel/cobalt transporter |
| Oxidative stress response | ||
| ahpCF | 3.67 | alkyl hydroperoxide reductase |
| grxA | 2.40 | glutaredoxin A |
| katG | 1.89 | hydroperoxidase |
| sodA | 2.18 | superoxide dismutase (manganese) |
| sodB | −3.20 | superoxide dismutase (iron) |
| SPI-1 | ||
| sitABCD | 1.13 | manganese/iron transport |
| prgKJIH-orgABC | 1.27 | needle complex |
| hilA-iagB | 1.56 | invasion protein |
| spaPOL-invJI | 2.17 | needle complex |
| invABEFG | 1.20 | invasion complex |
| invH | 2.46 | involved in the synthesis of the type III secretion system |
| SPI-2 | ||
| ssaBCDE | −2.36 | type III secretion system apparatus |
| sseABCDE | −2.47 | translocation machinery components |
| ssaGHIJ | −1.88 | type III secretion system |
| ssaMVN | −2.66 | type III secretion system |
| ssaRSTU | −1.88 | type III secretion system |
| ydhC | 1.68 | inner membrane transport |
| Motility and chemotaxis | ||
| flh | 1.70 | flagellar biosynthesis |
| fli | 1.48 | flagellar biosynthesis |
| mot | 1.70 | flagellar motor protein |
| che | 1.70 | chemotaxis |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balkin, A.S.; Plotnikov, A.O.; Gogoleva, N.E.; Gogolev, Y.V.; Demchenko, K.N.; Cherkasov, S.V. Cappable-Seq Reveals Specific Patterns of Metabolism and Virulence for Salmonella Typhimurium Intracellular Survival within Acanthamoeba castellanii. Int. J. Mol. Sci. 2021, 22, 9077. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22169077
Balkin AS, Plotnikov AO, Gogoleva NE, Gogolev YV, Demchenko KN, Cherkasov SV. Cappable-Seq Reveals Specific Patterns of Metabolism and Virulence for Salmonella Typhimurium Intracellular Survival within Acanthamoeba castellanii. International Journal of Molecular Sciences. 2021; 22(16):9077. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22169077
Chicago/Turabian StyleBalkin, Alexander S., Andrey O. Plotnikov, Natalia E. Gogoleva, Yuri V. Gogolev, Kirill N. Demchenko, and Sergey V. Cherkasov. 2021. "Cappable-Seq Reveals Specific Patterns of Metabolism and Virulence for Salmonella Typhimurium Intracellular Survival within Acanthamoeba castellanii" International Journal of Molecular Sciences 22, no. 16: 9077. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22169077