
Loose Morphology and High Dynamism of OSER Structures Induced by the Membrane Domain of HMG-CoA Reductase


{kind=link}
{kind=link}
{kind=link}
{kind=link}
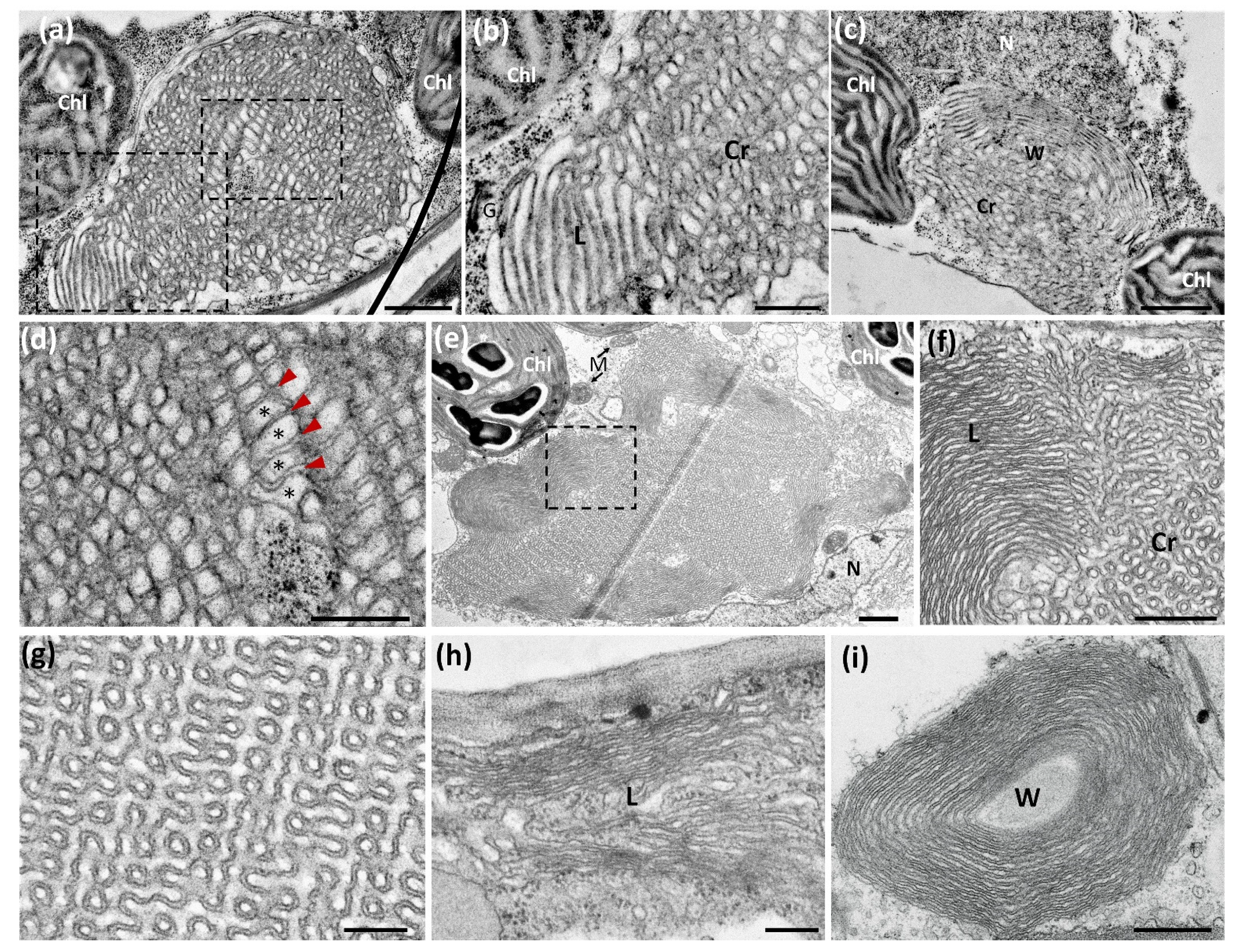{kind=link}
Abstract
:1. Introduction
2. Results
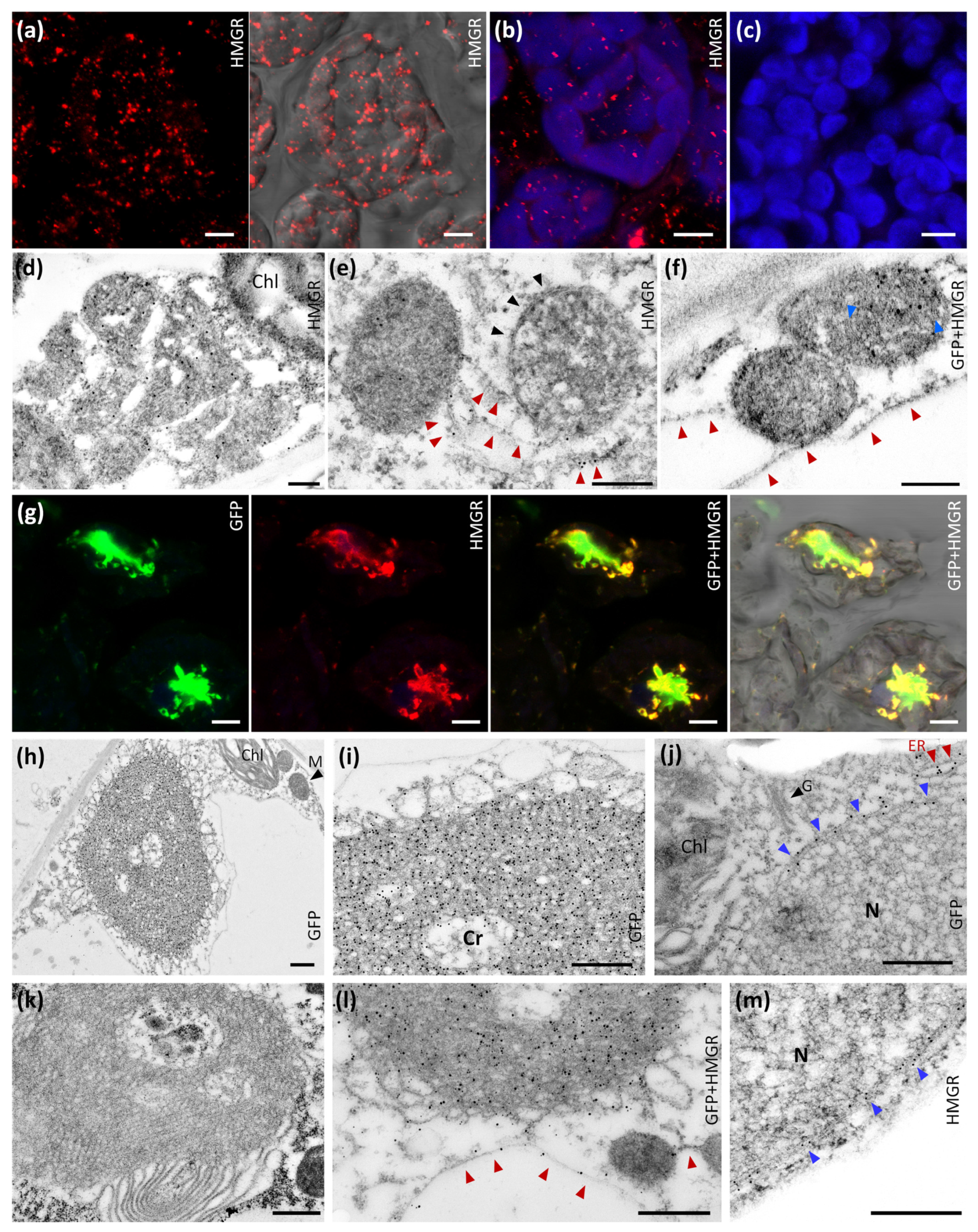
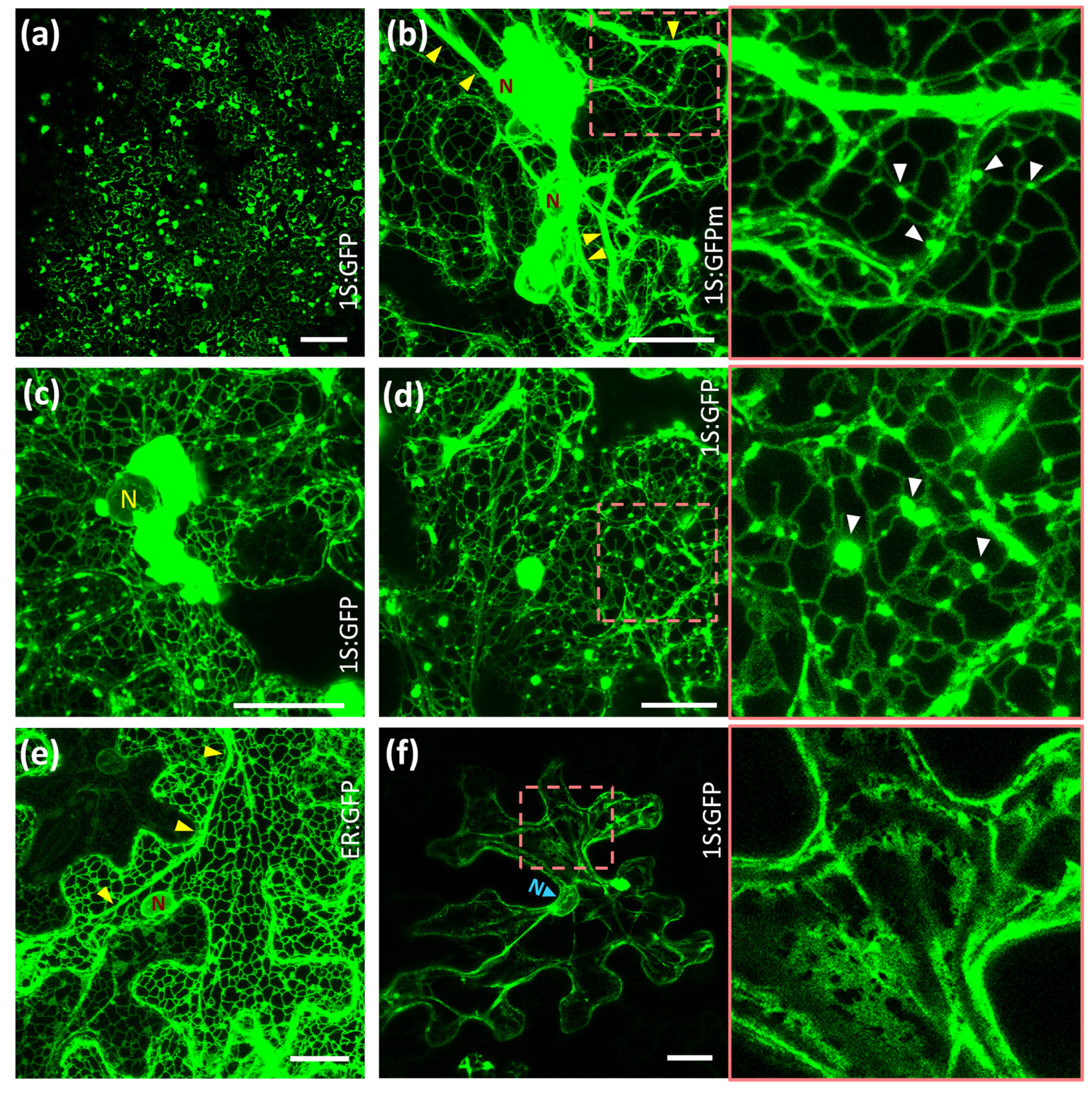
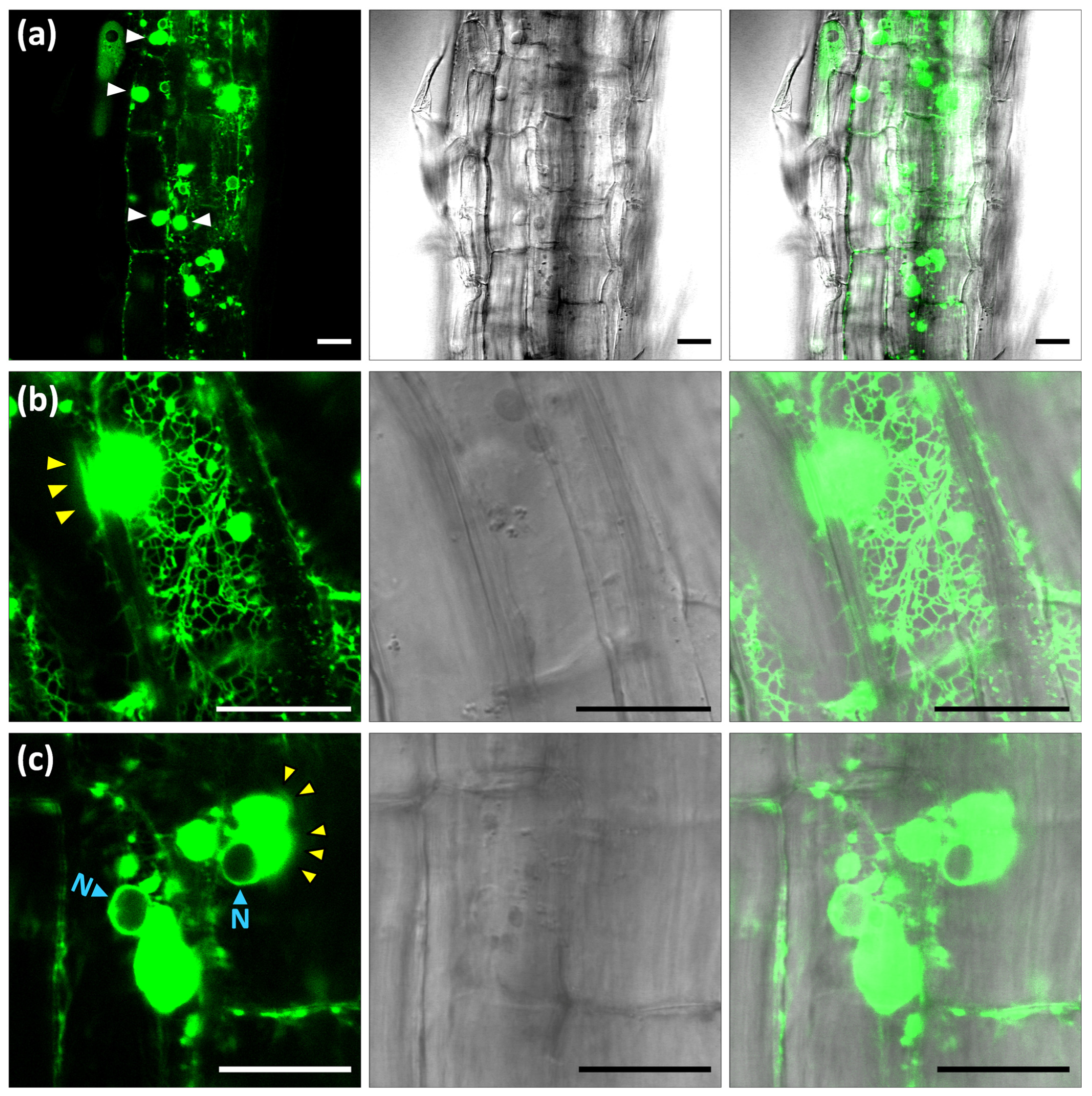
2.1. Subcellular Location of Arabidopsis HMGR in WT and 1S:GFP Transgenic Plants
2.2. Reversible Formation of ER-HMGR Domains
2.3. ER-HMGR Domains Are Highly Dynamic
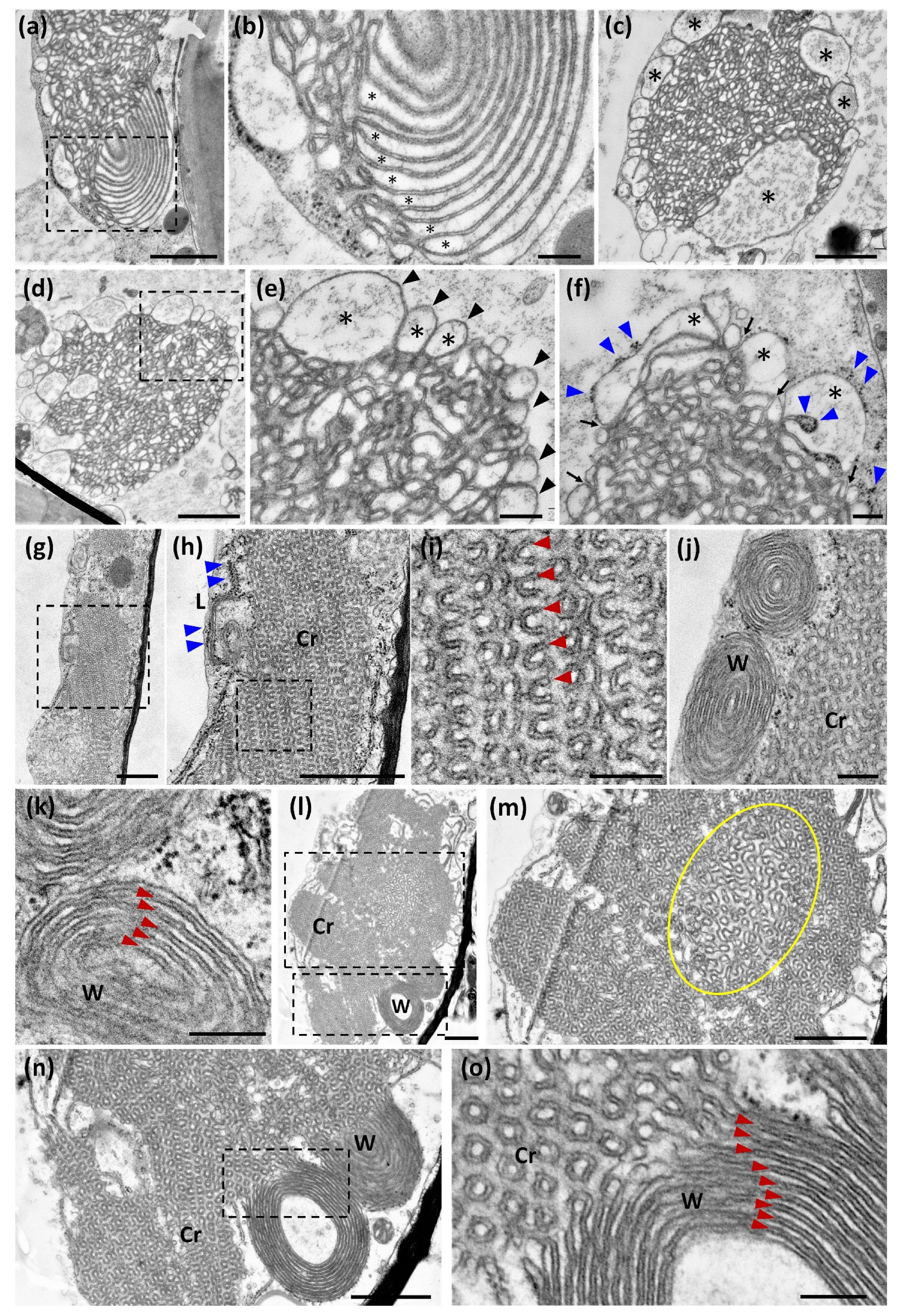
2.4. The Fixation and Dehydration Method Severely Affects OSER Ultrastructure
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Constructs
4.3. Transient Expression in Nicotiana Benthamiana Leaves
4.4. Source and Use of Antibodies
4.5. Immunolocalization in Whole Mount
4.6. Confocal Microscopy
4.7. Chemical Fixation for Ultrastructural Studies
4.8. High-Presure Freezing (HPF) for Ultrastructural Studies
4.9. Ultrastructural Analysis
4.10. Immunochemical Ultrastructural Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Burg, J.S.; Espenshade, P.J. Regulation of HMG-CoA reductase in mammals and yeast. Prog. Lipid Res. 2011, 50, 403–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutensohn, M.; Nagegowda, D.A.; Dudareva, N. Involvement of compartmentalization in monoterpene and sesquiterpene biosynthesis in plants. In Isoprenoid Synthesis in Plants and Microorganisms: New Concepts and Experimental Approaches; Springer: New York, NY, USA, 2013; pp. 155–169. ISBN 9781461440635. [Google Scholar]
- Rodríguez-Concepción, M.; Boronat, A. Breaking new ground in the regulation of the early steps of plant isoprenoid biosynthesis. Curr. Opin. Plant Biol. 2015, 25, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Kutchan, T.M.; Gershenzon, J.; Moller, B.L.; Gang, D.R. Natural products. In Biochemistry and Molecular Biology of Plants; Buchanan, B.B., Gruissem, W., Jones, R.L., Eds.; Wiley Blackwell: Chichester, UK; Hoboken, NJ, USA, 2015; pp. 1132–1206. ISBN 9780470714218. [Google Scholar]
- Lumbreras, V.; Campos, N.; Boronat, A. The use of an alternative promoter in the Arabidopsis thaliana HMG1 gene generates an mRNA that encodes a novel 3-hydroxy-3-methylglutaryl coenzyme A reductase isoform with an extended N-terminal region. Plant J. 1995, 8, 541–549. [Google Scholar] [CrossRef] [PubMed]
- NCBI Genome Browser. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/genome/browse (accessed on 9 July 2021).
- Enjuto, M.; Balcells, L.; Campos, N.; Caelles, C.; Arro, M.; Boronat, A. Arabidopsis thaliana contains two differentially expressed 3-hydroxy-3-methylglutaryl-CoA reductase genes, which encode microsomal forms of the enzyme. Proc. Natl. Acad. Sci USA 1994, 91, 927–931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chappell, J. The Biochemistry and Molecular Biology of Isoprenoid Metabolism. Plant Physiol. 1995, 107, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Mo, C.; Bard, M. A systematic study of yeast sterol biosynthetic protein–protein interactions using the split-ubiquitin system. Biochim. Biophys. Acta 2005, 1737, 152–160. [Google Scholar] [CrossRef]
- Mialoundama, A.S.; Jadid, N.; Brunel, J.; Di Pascoli, T.; Heintz, D.; Erhardt, M.; Mutterer, J.; Bergdoll, M.; Ayoub, D.; Van Dorsselaer, A.; et al. Arabidopsis ERG28 tethers the sterol C4-demethylation complex to prevent accumulation of a biosynthetic intermediate that interferes with polar auxin transport. Plant Cell 2013, 25, 4879–4893. [Google Scholar] [CrossRef] [Green Version]
- Gachotte, D.; Eckstein, J.; Barbuch, R.; Hughes, T.; Roberts, C.; Bard, M. A novel gene conserved from yeast to humans is involved in sterol biosynthesis. J. Lipid Res. 2001, 42, 150–154. [Google Scholar] [CrossRef]
- Campos, N.; Boronat, A. Targeting and topology in the membrane of plant 3-hydroxy-3-methylglutaryl coenzyme A reductase. Plant Cell 1995, 7, 2163–2174. [Google Scholar] [CrossRef] [Green Version]
- Basson, M.E.; Thorsness, M.; Finer-Moore, J.; Stroud, R.M.; Rine, J. Structural and functional conservation between yeast and human 3-hydroxy-3-methylglutaryl coenzyme A reductases, the rate-limiting enzyme of sterol biosynthesis. Mol. Cell. Biol. 1988, 8, 3797–3808. [Google Scholar] [CrossRef] [Green Version]
- Liscum, L.; Finer-Moore, J.; Stroud, R.M.; Luskey, K.L.; Brown, M.S.; Goldstein, J.L. Domain structure of 3-hydroxy-3-methylglutaryl coenzyme A reductase, a glycoprotein of the endoplasmic reticulum. J. Biol. Chem. 1985, 260, 522–530. [Google Scholar] [CrossRef]
- Olender, E.; Simon, R. The intracellular targeting and membrane topology of 3-hydroxy-3-methylglutaryl-CoA reductase. J. Biol. Chem. 1992, 267, 4223–4235. [Google Scholar] [CrossRef]
- Profant, D.A.; Roberts, C.J.; Wright, R.L. Mutational analysis of the karmellae-inducing signal in Hmg1p, a yeast HMG-CoA reductase isozyme. Yeast 2000, 16, 811–827. [Google Scholar] [CrossRef]
- Campos, N.; Palau, J.; Zwieb, C. Diversity of 7 SL RNA from the signal recognition particle of maize endosperm. Nucleic Acids Res. 1989, 17, 1573–1588. [Google Scholar] [CrossRef] [Green Version]
- Leivar, P.; González, V.M.; Castel, S.; Trelease, R.N.; López-Iglesias, C.; Arró, M.; Boronat, A.; Campos, N.; Ferrer, A.; Fernàndez-Busquets, X. Subcellular localization of Arabidopsis 3-hydroxy-3-methylglutaryl-coenzyme A reductase. Plant Physiol. 2005, 137, 57–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrero, S.; Grados-Torrez, R.E.; Leivar, P.; Antolín-Llovera, M.; López-Iglesias, C.; Cortadellas, N.; Ferrer, J.C.; Campos, N. Proliferation and morphogenesis of the endoplasmic reticulum driven by the membrane domain of 3-hydroxy-3-methylglutaryl coenzyme A reductase in plant cells. Plant Physiol. 2015, 168, 899–914. [Google Scholar] [CrossRef] [PubMed]
- Jingami, H.; Brown, M.S.; Goldstein, J.L.; Anderson, R.G.; Luskey, K.L. Partial deletion of membrane-bound domain of 3-hydroxy-3-methylglutaryl coenzyme A reductase eliminates sterol-enhanced degradation and prevents formation of crystalloid endoplasmic reticulum. J. Cell Biol. 1987, 104, 1693–1704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, R.G.; Basson, M.D.; D’Ari, L.; Rine, J. Increased amounts of HMG-CoA reductase induce "karmellae": A proliferation of stacked membrane pairs surrounding the yeast nucleus. J. Cell Biol. 1988, 107, 101–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandor, A.; Fricker, M.D.; Kriechbaumer, V.; Sweetlove, L.J. IntEResting structures: Formation and applications of organized smooth endoplasmic reticulum in plant cells. Plant Physiol. 2020, 185, 550–561. [Google Scholar] [CrossRef]
- Chin, D.J.; Luskey, K.L.; Anderson, R.G.; Faust, J.R.; Goldstein, J.L.; Brown, M.S. Appearance of crystalloid endoplasmic reticulum in compactin-resistant Chinese hamster cells with a 500-fold increase in 3-hydroxy-3-methylglutaryl-coenzyme A reductase. Proc. Natl. Acad. Sci. 1982, 79, 1185–1189. [Google Scholar] [CrossRef] [Green Version]
- Eymé, M.J. Infrastructure des cellules nectarigènes de Diplotaxis erucoides D. C., Helleborus niger L. et H. fœtidus L. Comptes Rendus des Séances l’Académie des Sci. Ser. D 1996, 262, 1629–1632. [Google Scholar]
- Eymé, M.J.; Le Blanc, M. Contribution à l’étude inframicroscopique d’inclusions cytoplasmatiques présentes dans les ovules de Ficaria et dans les nectaries d’Helleborus. Comptes Rendus des Séances l’Académie des Sci. Ser. D 1963, 256, 4958–4959. [Google Scholar]
- Eymé, J. Nouvelles observations sur l’infrastructure des tissus nectarigènes floraux. Le Bot. (serie L) 1967, 50, 169–183. [Google Scholar]
- Bassot, J.M. Une forme microtubulaire et paracristalline de reticulum endoplasmique dans les photocytes des annelides polynoinæ. J. Cell Biol. 1966, 31, 135–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wooding, F.B.P. Endoplasmic reticulum aggregates of ordered structure. Planta 1967, 76, 205–208. [Google Scholar] [CrossRef] [PubMed]
- Christensen, A.K. The fine structure of testicular interstitial cells in guinea pigs. J. Cell Biol. 1965, 26, 911–935. [Google Scholar] [CrossRef] [PubMed]
- Christensen, A.K.; Fawcett, D.W. The normal fine structure of opossum testicular interstitial cells. J. Cell Biol. 1961, 9, 653–670. [Google Scholar] [CrossRef]
- Orrenius, S.; Ericsson, J.L.E.; Ernster, L. Phenobarbital-induced synthesis of the microsomal drug-metabolizing enzyme system and its relationship to the proliferation of endoplasmic membranes. J. Cell Biol. 1965, 25, 627–639. [Google Scholar] [CrossRef]
- Jones, A.L.; Fawcett, D.W. Hypertrophy of the agranular endoplasmic reticulum in hamster liver induced by phenobarbital (with a review on the functions of this organelle in liver). J. Histochem. Cytochem. 1966, 14, 215–232. [Google Scholar] [CrossRef]
- Franke, W.W.; Scheer, U. Some structural differentiations in the HeLa cell: Heavy bodies annulate lamellae and cotte de maillet endoplasmic reticulum. Cytobiologie 1971, 4, 317–329. [Google Scholar]
- Black, V.H. The development of smooth-surfaced endoplasmic reticulum in adrenal cortical cells of fetal guinea pigs. Am. J. Anat. 1972, 135, 381–417. [Google Scholar] [CrossRef]
- Barton, B.R.; Hertig, A.T. Ultrastructure of quiescent oocytes of Cebus albifrons. J. Anat. 1975, 120, 227–238. [Google Scholar] [PubMed]
- Davidowitz, J.; Philips, G.H.; Pachter, B.R.; Breinin, G.M. Particle-free and glycogen-bearing double membrane arrays in extraocular muscle of rabbit. Am. J. Pathol. 1975, 78, 191–198. [Google Scholar]
- Grimley, P.M.; Schaff, Z. Significance of tubuloreticular inclusions in the pathobiology of human diseases. Pathobiol. Annu. 1976, 6, 221–257. [Google Scholar]
- Schaff, Z.; Lapis, K.; Grimley, P.M. Undulating membraneous structures associated with the endoplasmic reticulum in tumour cells. Int. J. Cancer 1976, 18, 697–702. [Google Scholar] [CrossRef] [PubMed]
- Linder, J.C.; Staehelin, A. The membrane lattice: A novel organelle of the trypanosomatid flagellate Leptomonas collosoma. J. Ultrastruct. Res. 1980, 72, 200–205. [Google Scholar] [CrossRef]
- Yamamoto, A.; Otsu, H.; Yoshimori, T.; Maeda, N.; Mikoshiba, K.; Tashiro, Y. Stacks of flattened smooth endoplasmic reticulum highly enriched in inositol 1,4,5-trisphosphate(InsP3)receptor in mouse cerebellar Purkinje cells. Cell Struct. Funct. 1991, 16, 419–432. [Google Scholar] [CrossRef] [Green Version]
- Karnaky, K.J.; Lau, K.R.; Garretson, L.T.; Schultz, S.G. Seasonal variations in the fine structure of the Necturus maculosus urinary bladder epithelium: Low transporters and high transporters. Am. J. Anat. 1984, 171, 227–242. [Google Scholar] [CrossRef] [PubMed]
- Kerr, J.B.; Weiss, M. Spontaneous or experimentally induced formation of a special zone in the adrenal cortex of the adult brush-tailed possum(Trichosurus vulpecula). Am. J. Anat. 1991, 190, 101–117. [Google Scholar] [CrossRef] [PubMed]
- Wolf, K.W.; Motzko, D. Paracrystalline endoplasmic reticulum is typical of gametogenesis in Hemiptera species. J. Struct. Biol. 1995, 114, 105–114. [Google Scholar] [CrossRef]
- Yamamoto, A.; Masaki, R.; Tashiro, Y. Formation of crystalloid endoplasmic reticulum in COS cells upon overexpression of microsomal aldehyde dehydrogenase by cDNA transfection. J. Cell Sci. 1996, 109, 1727–1738. [Google Scholar] [CrossRef] [PubMed]
- Snapp, E.; Hegde, R.; Francolini, M.; Lombardo, F.; Colombo, S.; Pedrazzini, E.; Borgese, N.; Lippincott-Schwartz, J. Formation of stacked ER cisternae by low affinity protein interactions. J. Cell Biol. 2003, 163, 257–269. [Google Scholar] [CrossRef] [PubMed]
- Almsherqi, Z.A.; Kohlwein, S.D.; Deng, Y. Cubic membranes: A legend beyond the Flatland* of cell membrane organization. J. Cell Biol. 2006, 173, 839–844. [Google Scholar] [CrossRef] [Green Version]
- Almsherqi, Z.A.; Landh, T.; Kohlwein, S.D.; Deng, Y. Chapter 6: Cubic membranes the missing dimension of cell membrane organization. Int. Rev. cell Mol. Biol. 2009, 274, 275–342. [Google Scholar] [CrossRef] [PubMed]
- Leivar, P.; Antolín-Llovera, M.; Ferrero, S.; Closa, M.; Arró, M.; Ferrer, A.; Boronat, A.; Campos, N. Multilevel control of Arabidopsis 3-hydroxy-3-methylglutaryl coenzyme A reductase by protein phosphatase 2A. Plant Cell 2011, 23, 1494–1511. [Google Scholar] [CrossRef] [Green Version]
- Westrate, L.; Lee, J.; Prinz, W.; Voeltz, G. Form follows function: The importance of endoplasmic reticulum shape. Annu. Rev. Biochem. 2015, 84, 791–811. [Google Scholar] [CrossRef] [PubMed]
- Hawes, C.; Kiviniemi, P.; Kriechbaumer, V. The endoplasmic reticulum: A dynamic and well-connected organelle. J. Integr. Plant Biol. 2015, 57, 50–62. [Google Scholar] [CrossRef] [Green Version]
- Nishikawa, S.; Hirata, A.; Nakano, A. Inhibition of endoplasmic reticulum (ER)-to-Golgi transport induces relocalization of binding protein (BiP) within the ER to form the BiP bodies. Mol. Biol. Cell 1994, 5, 1129–1143. [Google Scholar] [CrossRef] [Green Version]
- Bergmann, J.E.; Fusco, P.J. The G protein of vesicular stomatitis virus has free access into and egress from the smooth endoplasmic reticulum of UT-1 cells. J. Cell Biol. 1990, 110, 625–635. [Google Scholar] [CrossRef]
- Anderson, R.; Orci, L.; Brown, M.; Garcia-Segura, L.; Goldstein, J. Ultrastructural analysis of crystalloid endoplasmic reticulum in UT-1 cells and its disappearance in response to cholesterol. J. Cell Sci. 1983, 63, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Hurbain, I.; Sachse, M. The future is cold: Cryo-preparation methods for transmission electron microscopy of cells. Biol. Cell 2011, 103, 405–420. [Google Scholar] [CrossRef]
- Ebersold, H.R.; Cordier, J.-L. Bacterial mesosomes: Method dependent artifacts. Arch. Microbiol. 1981, 130, 19–22. [Google Scholar] [CrossRef]
- Higgins, M.L.; Tsien, H.C.; Daneo-Moore, L. Organization of mesosomes in fixed and unfixed cells. J. Bacteriol. 1976, 127, 1519–1523. [Google Scholar] [CrossRef] [Green Version]
- Silva, M.; Sousa, J.; Polónia, J.; Macedo, M.; Parente, A.M. Bacterial mesosomes: Real structures of artifacts? Biochim. et Biophys. Acta (BBA)-Biomembr. 1976, 443, 92–105. [Google Scholar] [CrossRef]
- Chandler, D. Comparison of quick-frozen and chemically fixed sea-urchin eggs: Structural evidence that cortical granule exocytosis is preceded by a local increase in membrane mobility. J. Cell Sci. 1984, 72, 23–36. [Google Scholar] [CrossRef] [PubMed]
- Murk, J.L.A.N.; Posthuma, G.; Koster, A.; Geuze, H.J.; Verkleij, A.J.; Kleijmeer, M.J.; Humbel, B.M. Influence of aldehyde fixation on the morphology of endosomes and lysosomes: Quantitative analysis and electron tomography. J. Microsc. 2003, 212, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Orci, L.; Brown, M.S.; Goldstein, J.L.; Garcia-Segura, L.; Anderson, R.G. Increase in membrane cholesterol: A possible trigger for degradation of HMG-CoA reductase and crystalloid endoplasmic reticulum in UT-1 cells. Cell 1984, 36, 835–845. [Google Scholar] [CrossRef]
- Kochevar, D.; Anderson, R. Purified crystalloid endoplasmic reticulum from UT-1 cells contains multiple proteins in addition to 3-hydroxy-3-methylglutaryl coenzyme A reductase. J. Biol. Chem. 1987, 262, 10321–10326. [Google Scholar] [CrossRef]
- Zacharias, D.A.; Violin, J.D.; Newton, A.C.; Tsien, R.Y. Partitioning of lipid-modified monomeric GFPs into membrane microdomains of live cells. Science 2002, 296, 913–916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Istvan, E.S.; Palnitkar, M.; Buchanan, S.K.; Deisenhofer, J. Crystal structure of the catalytic portion of human HMG-CoA reductase: Insights into regulation of activity and catalysis. EMBO J. 2000, 19, 819–830. [Google Scholar] [CrossRef] [PubMed]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio-assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Batoko, H.; Zheng, H.-Q.; Hawes, C.; Moore, I. A rab1 GTPase is required for transport between the endoplasmic reticulum and Golgi apparatus and for normal Golgi movement in plants. Plant Cell 2000, 12, 2201–2217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Der Hoorn, R.A.L.; Laurent, F.; Roth, R.; de Wit, P. Agroinfiltration is a versatile tool that facilitates comparative analyses of Avr9/Cf-9-induced and Avr4/Cf-4-induced necrosis. Mol. Plant-Microbe Interactions 2000, 13, 439–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koncz, C.; Schell, J. The promoter of TL-DNA gene 5 controls the tissue-specific expression of chimaeric genes carried by a novel type of Agrobacterium binary vector. Mol. Genet. Genom. 1986, 204, 383–396. [Google Scholar] [CrossRef]
- Sauer, M.; Paciorek, T.; Benková, E.; Friml, J. Immunocytochemical techniques for whole-mount in situ protein localization in plants. Nat. Protoc. 2006, 1, 98–103. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grados-Torrez, R.E.; López-Iglesias, C.; Ferrer, J.C.; Campos, N. Loose Morphology and High Dynamism of OSER Structures Induced by the Membrane Domain of HMG-CoA Reductase. Int. J. Mol. Sci. 2021, 22, 9132. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179132
Grados-Torrez RE, López-Iglesias C, Ferrer JC, Campos N. Loose Morphology and High Dynamism of OSER Structures Induced by the Membrane Domain of HMG-CoA Reductase. International Journal of Molecular Sciences. 2021; 22(17):9132. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179132
Chicago/Turabian StyleGrados-Torrez, Ricardo Enrique, Carmen López-Iglesias, Joan Carles Ferrer, and Narciso Campos. 2021. "Loose Morphology and High Dynamism of OSER Structures Induced by the Membrane Domain of HMG-CoA Reductase" International Journal of Molecular Sciences 22, no. 17: 9132. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179132