
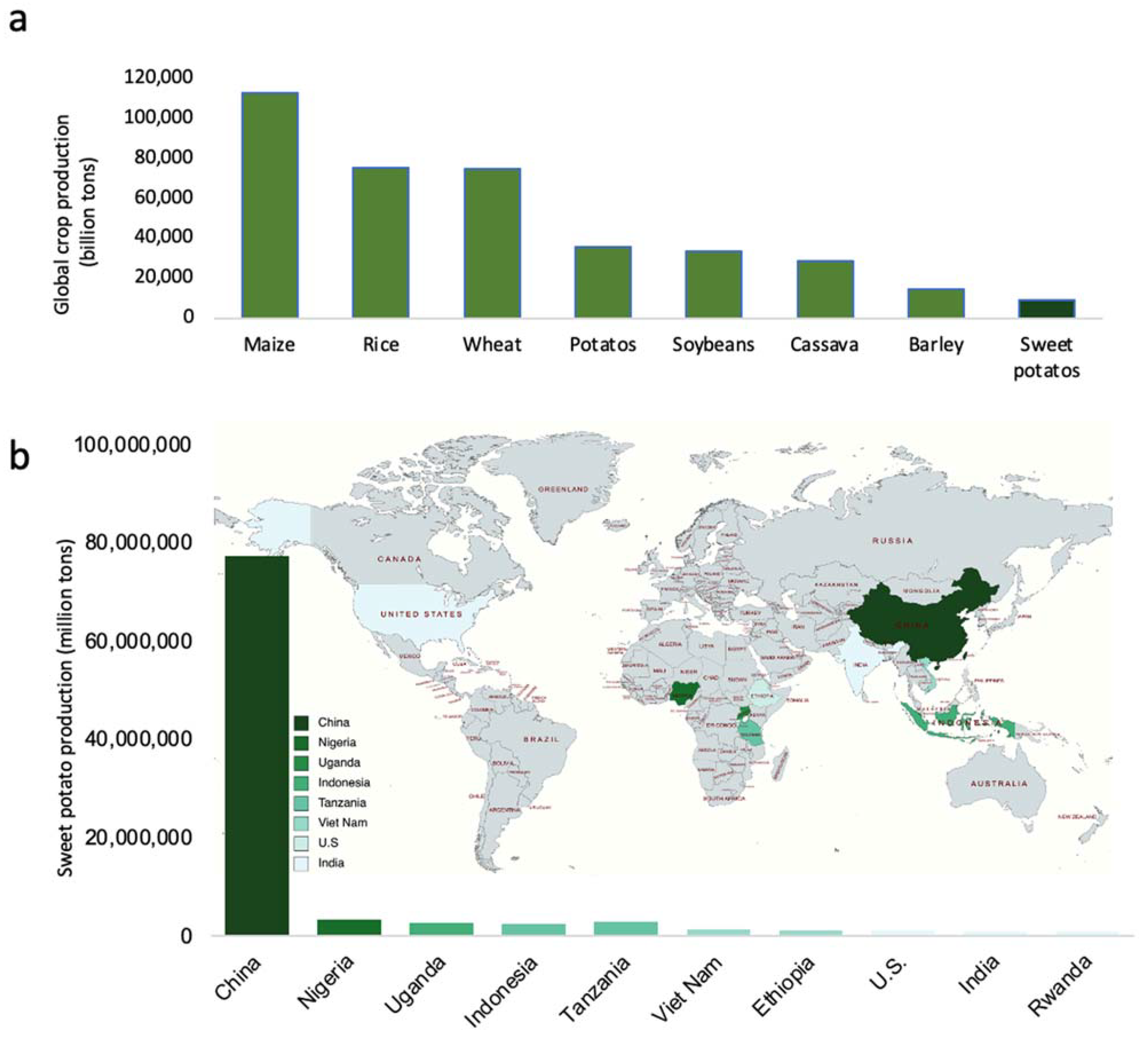
Engineering Properties of Sweet Potato Starch for Industrial Applications by Biotechnological Techniques including Genome Editing

 ,
,  , , , , ,
, , , , ,
Abstract
:1. Introduction
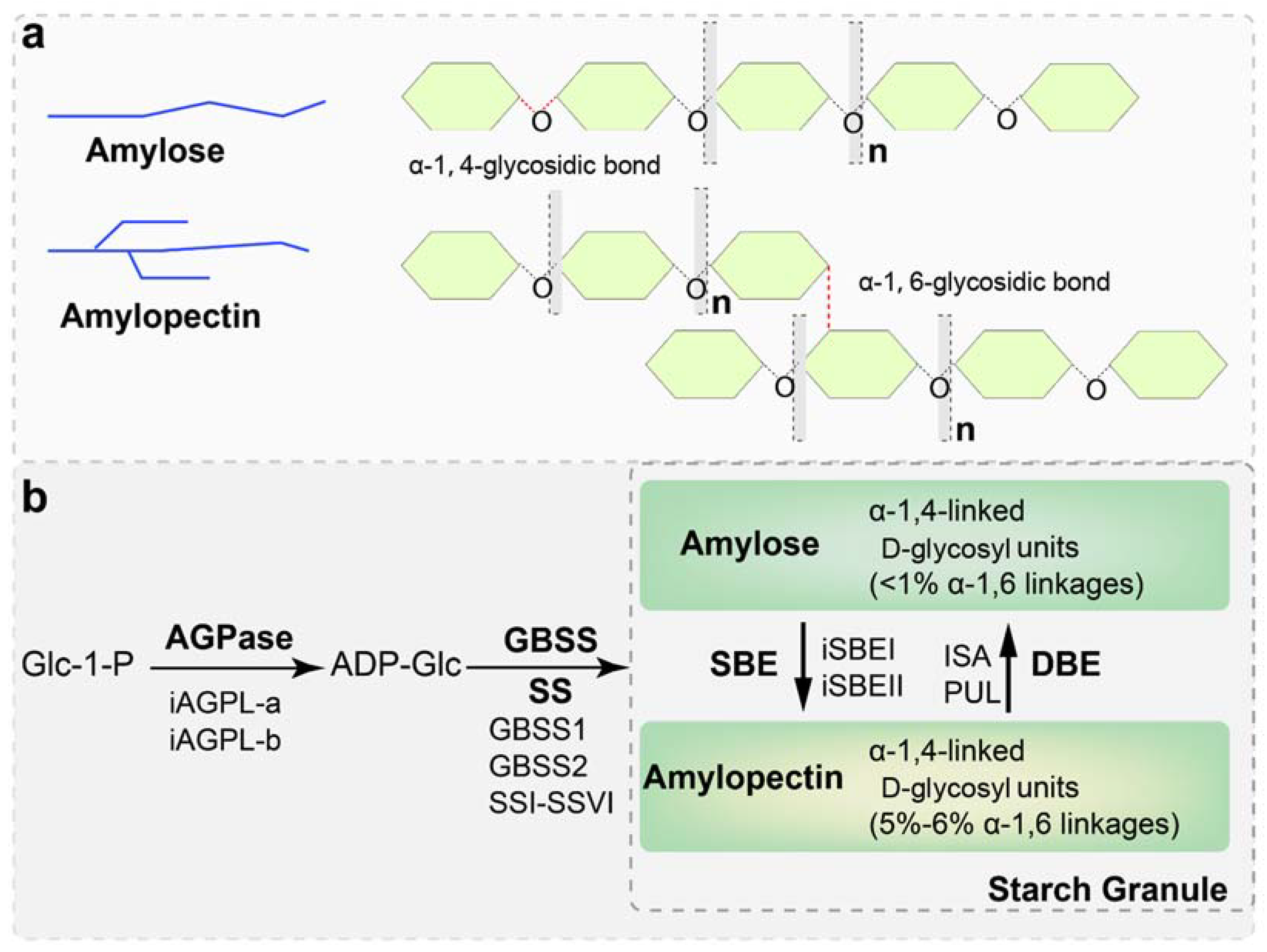
2. Starch Biosynthesis in Sweet Potato and Other Plants
3. Functionality and Regulation of Biosynthesis of Starch
3.1. Pasting and Gelatinization Properties
3.2. Amylose Content and Amylose/Amylopectin Ratio
3.3. Starch Granule Size
3.4. Chain Length Distributions (CLDs)
3.5. Starch Phosphorylation
4. Value-Added Products from Sweet Potato Starch
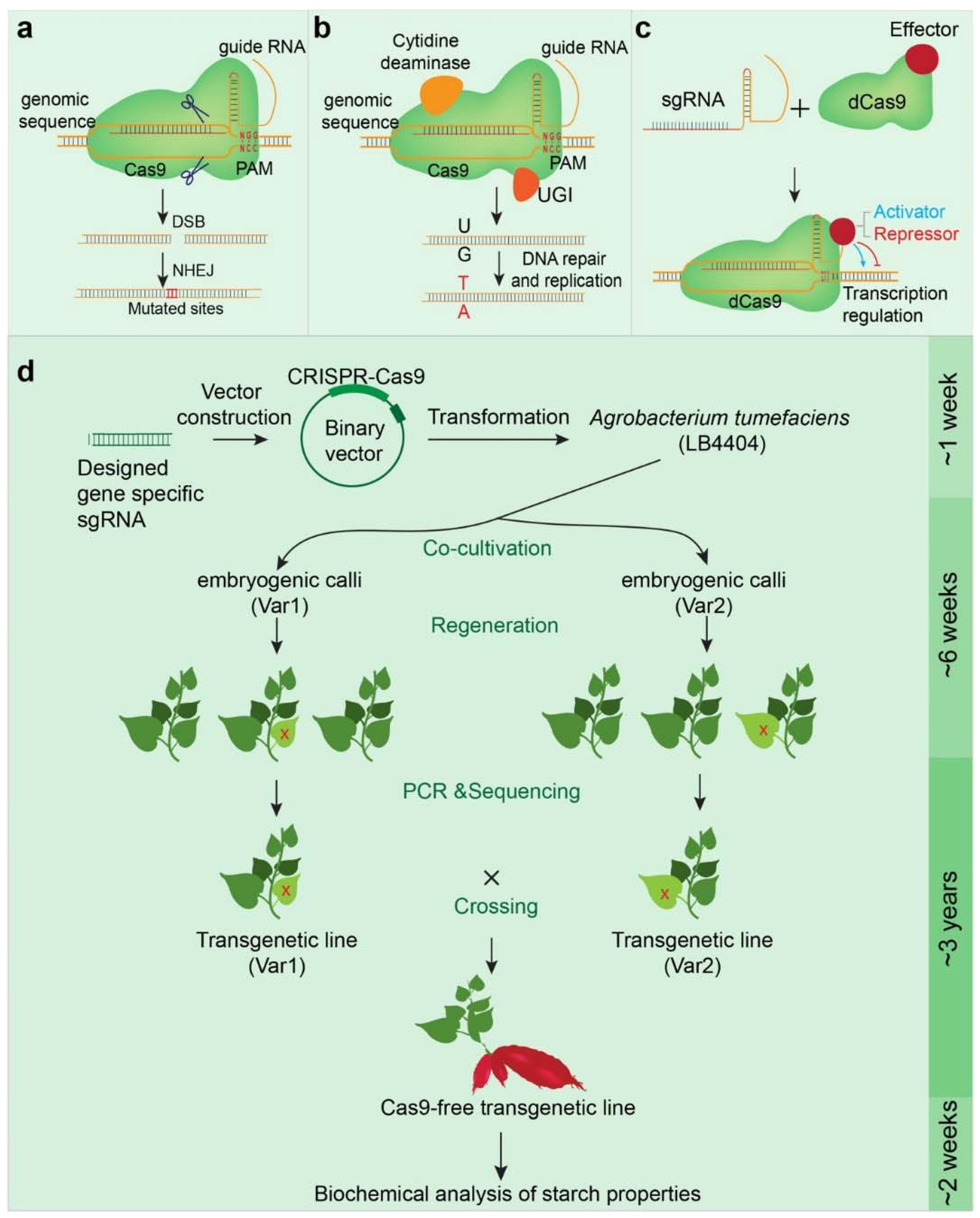
5. The Promise and Challenge of Genome-Editing Sweet Potato
6. Discussion
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- O’Brien, P.J. The Sweet Potato: Its Origin and Dispersal. Am. Anthropol. 1972, 74, 342–365. [Google Scholar] [CrossRef]
- Loebenstein, G.; Fuentes, S.; Cohen, J.; Salazar, L.F. Sweet Potato. In Virus and Virus-like Diseases of Major Crops in Developing Countries; Loebenstein, G., Thottappilly, G., Eds.; Springer: Dordrecht, The Netherlands, 2003; pp. 223–248. [Google Scholar] [CrossRef]
- Abegunde, O.K.; Mu, T.-H.; Chen, J.-W.; Deng, F.-M. Physicochemical characterization of sweet potato starches popularly used in Chinese starch industry. Food Hydrocoll. 2013, 33, 169–177. [Google Scholar] [CrossRef]
- James, M.G.; Denyer, K.; Myers, A.M. Starch synthesis in the cereal endosperm. Curr. Opin. Plant Biol. 2003, 6, 215–222. [Google Scholar] [CrossRef]
- Jeon, J.-S.; Ryoo, N.; Hahn, T.-R.; Walia, H.; Nakamura, Y. Starch biosynthesis in cereal endosperm. Plant Physiol. Biochem. 2010, 48, 383–392. [Google Scholar] [CrossRef]
- Ballicora, M.A.; Iglesias, A.A.; Preiss, J. ADP-Glucose Pyrophosphorylase: A Regulatory Enzyme for Plant Starch Synthesis. Photosynth. Res. 2004, 79, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, Y.; Zhang, H.; Zhai, H.; Liu, Q.; He, S. A soluble starch synthase I gene, IbSSI, alters the content, composition, granule size and structure of starch in transgenic sweet potato. Sci. Rep. 2017, 7, 2315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tetlow, I.J.; Emes, M.J. A review of starch-branching enzymes and their role in amylopectin biosynthesis. IUBMB Life 2014, 66, 546–558. [Google Scholar] [CrossRef]
- Kubo, A.; Fujita, N.; Harada, K.; Matsuda, T.; Satoh, H.; Nakamura, Y. The starch-debranching enzymes isoamylase and pullulanase are both involved in amylopectin biosynthesis in rice endosperm. Plant Physiol. 1999, 121, 399–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khlestkin, V.K.; Peltek, S.E.; Kolchanov, N.A. Review of direct chemical and biochemical transformations of starch. Carbohydr. Polym. 2018, 181, 460–476. [Google Scholar] [CrossRef] [PubMed]
- Crofts, N.; Iizuka, Y.; Abe, N.; Miura, S.; Kikuchi, K.; Matsushima, R.; Fujita, N. Rice Mutants Lacking Starch Synthase I or Branching Enzyme IIb Activity Altered Starch Biosynthetic Protein Complexes. Front. Plant Sci 2018, 9, 1817. [Google Scholar] [CrossRef] [Green Version]
- Yoo, S.-H.; Lee, B.-H.; Li, L.; Perris, S.D.N.; Spalding, M.H.; Han, S.Y.; Jane, J.-L. Biocatalytic role of potato starch synthase III for α-glucan biosynthesis in Synechocystis sp. PCC6803 mutants. Int. J. Biol. Macromol. 2015, 81, 710–717. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Liu, L.; Qu, J.; Li, S.; Blennow, A.; Seytahmetovna, S.A.; Liu, X.; Guo, D. The relationship between the expression pattern of starch biosynthesis enzymes and molecular structure of high amylose maize starch. Carbohydr. Polym. 2020, 247, 116681. [Google Scholar] [CrossRef] [PubMed]
- Kwak, M.S.; Oh, M.-J.; Paek, K.-H.; Shin, J.S.; Bae, J.M. Dissected effect of a transit peptide of the ADP-glucose pyrophosphorylase gene from sweetpotato (ibAGP2) in increasing foreign protein accumulation. Plant Cell Rep. 2008, 27, 1359–1367. [Google Scholar] [CrossRef]
- Kim, T.-W.; Goo, Y.-M.; Lee, C.-H.; Lee, B.-H.; Bae, J.-M.; Lee, S.-W. The sweetpotato ADP-glucose pyrophosphorylase gene (ibAGP1) promoter confers high-level expression of the GUS reporter gene in the potato tuber. Comptes Rendus Biol. 2009, 332, 876–885. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wu, Y.; Zhang, Y.; Yang, J.; Fan, W.; Zhang, H.; Zhao, S.; Yuan, L.; Zhang, P. CRISPR/Cas9-Based Mutagenesis of Starch Biosynthetic Genes in Sweet Potato (Ipomoea batatas) for the Improvement of Starch Quality. Int. J. Mol. Sci. 2019, 20, 4702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahata, Y.; Tanaka, M.; Otani, M.; Katayama, K.; Kitahara, K.; Nakayachi, O.; Nakayama, H.; Yoshinaga, M. Inhibition of the expression of the starch synthase II gene leads to lower pasting temperature in sweetpotato starch. Plant Cell Rep. 2010, 29, 535–543. [Google Scholar] [CrossRef]
- Kimura, T.; Otani, M.; Noda, T.; Ideta, O.; Shimada, T.; Saito, A. Absence of amylose in sweet potato [Ipomoea batatas (L.) Lam.] following the introduction of granule-bound starch synthase I cDNA. Plant Cell Rep. 2001, 20, 663–666. [Google Scholar] [CrossRef]
- Noda, T.; Kimura, T.; Otani, M.; Ideta, O.; Shimada, T.; Saito, A.; Suda, I. Physicochemical properties of amylose-free starch from transgenic sweet potato. Carbohydr. Polym. 2002, 49, 253–260. [Google Scholar] [CrossRef]
- Otani, M.; Hamada, T.; Katayama, K.; Kitahara, K.; Kim, S.-H.; Takahata, Y.; Suganuma, T.; Shimada, T. Inhibition of the gene expression for granule-bound starch synthase I by RNA interference in sweet potato plants. Plant Cell Rep. 2007, 26, 1801–1807. [Google Scholar] [CrossRef]
- Kitahara, K.; Nakamura, Y.; Otani, M.; Hamada, T.; Nakayachi, O.; Takahata, Y. Carbohydrate components in sweetpotato storage roots: Their diversities and genetic improvement. Breed. Sci. 2017, 67, 62–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, W.; Yang, J.; Hong, Y.; Liu, G.; Zheng, J.; Gu, Z.; Zhang, P. Impact of amylose content on starch physicochemical properties in transgenic sweet potato. Carbohydr. Polym. 2015, 122, 417–427. [Google Scholar] [CrossRef]
- Zhang, B.; Zhou, W.; Qiao, D.; Zhang, P.; Zhao, S.; Zhang, L.; Xie, F. Changes in Nanoscale Chain Assembly in Sweet Potato Starch Lamellae by Downregulation of Biosynthesis Enzymes. J. Agric. Food Chem. 2019, 67, 6302–6312. [Google Scholar] [CrossRef] [Green Version]
- Shimada, T.; Otani, M.; Hamada, T.; Kim, S.-H. Increase of amylose content of sweetpotato starch by RNA interference of the starch branching enzyme II gene (IbSBEII). Plant Biotechnol. 2006, 23, 85–90. [Google Scholar] [CrossRef] [Green Version]
- Kitahara, K.; Hamasuna, K.; Nozuma, K.; Otani, M.; Hamada, T.; Shimada, T.; Fujita, K.; Suganuma, T. Physicochemical properties of amylose-free and high-amylose starches from transgenic sweetpotatoes modified by RNA interference. Carbohydr. Polym. 2007, 69, 233–240. [Google Scholar] [CrossRef]
- Toyosawa, Y.; Kawagoe, Y.; Matsushima, R.; Crofts, N.; Ogawa, M.; Fukuda, M.; Kumamaru, T.; Okazaki, Y.; Kusano, M.; Saito, K.; et al. Deficiency of Starch Synthase IIIa and IVb Alters Starch Granule Morphology from Polyhedral to Spherical in Rice Endosperm. Plant Physiol. 2016, 170, 1255. [Google Scholar] [CrossRef] [Green Version]
- Miura, S.; Crofts, N.; Saito, Y.; Hosaka, Y.; Oitome, N.F.; Watanabe, T.; Kumamaru, T.; Fujita, N. Starch Synthase IIa-Deficient Mutant Rice Line Produces Endosperm Starch With Lower Gelatinization Temperature Than Japonica Rice Cultivars. Front. Plant Sci. 2018, 9, 645. [Google Scholar] [CrossRef] [Green Version]
- Firon, N.; LaBonte, D.; Villordon, A.; Kfir, Y.; Solis, J.; Lapis, E.; Perlman, T.S.; Doron-Faigenboim, A.; Hetzroni, A.; Althan, L.; et al. Transcriptional profiling of sweetpotato (Ipomoea batatas) roots indicates down-regulation of lignin biosynthesis and up-regulation of starch biosynthesis at an early stage of storage root formation. BMC Genom. 2013, 14, 460. [Google Scholar] [CrossRef] [Green Version]
- Choi, H.M.; Yoo, B. Steady and dynamic shear rheology of sweet potato starch–xanthan gum mixtures. Food Chem. 2009, 116, 638–643. [Google Scholar] [CrossRef]
- Park, S.-U.; Lee, C.-J.; Kim, S.-E.; Lim, Y.-H.; Lee, H.-U.; Nam, S.-S.; Kim, H.S.; Kwak, S.-S. Selection of flooding stress tolerant sweetpotato cultivars based on biochemical and phenotypic characterization. Plant Physiol. Biochem. 2020, 155, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Ren, Z.; He, S.; Zhao, N.; Zhai, H.; Liu, Q. A sucrose non-fermenting-1-related protein kinase-1 gene, IbSnRK1, improves starch content, composition, granule size, degree of crystallinity and gelatinization in transgenic sweet potato. Plant Biotechnol. J. 2019, 17, 21–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Moeinzadeh, M.H.; Kuhl, H.; Helmuth, J.; Xiao, P.; Haas, S.; Liu, G.; Zheng, J.; Sun, Z.; Fan, W.; et al. Haplotype-resolved sweet potato genome traces back its hexaploidization history. Nat. Plants 2017, 3, 696–703. [Google Scholar] [CrossRef] [PubMed]
- Gao, C. Genome engineering for crop improvement and future agriculture. Cell 2021, 184, 1621–1635. [Google Scholar] [CrossRef] [PubMed]
- Bertoft, E. Understanding Starch Structure: Recent Progress. Agronomy 2017, 7, 56. [Google Scholar] [CrossRef]
- Qu, J.; Xu, S.; Zhang, Z.; Chen, G.; Zhong, Y.; Liu, L.; Zhang, R.; Xue, J.; Guo, D. Evolutionary, structural and expression analysis of core genes involved in starch synthesis. Sci. Rep. 2018, 8, 12736. [Google Scholar] [CrossRef]
- Nakatani, M.; Komeichi, M. Relationship between starch content and activity of starch synthase and ADP-glucose pyrophosphorylase in tuberous root of sweet potato. JPN J. Crop Sci. 1992, 61, 463–468. [Google Scholar] [CrossRef] [Green Version]
- He, S.; Hao, X.; Wang, S.; Zhou, W.; Ma, Q.; Lu, X.; Chen, L.; Zhang, P. A newly-identified inactive starch synthase simultaneously regulates starch synthesis and carbon allocation in storage roots of cassava (Manihot esculenta Crantz). Cold Spring Harb. Lab. 2020. [Google Scholar] [CrossRef] [Green Version]
- Zeeman, S.C.; Kossmann, J.; Smith, A.M. Starch: Its metabolism, evolution, and biotechnological modification in plants. Annu. Rev. Plant Biol. 2010, 61, 209–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehrpouyan, S.; Menon, U.; Tetlow, I.J.; Emes, M.J. Protein phosphorylation regulates maize endosperm starch synthase IIa activity and protein−protein interactions. Plant J. 2021, 105, 1098–1112. [Google Scholar] [CrossRef]
- Boyer, C.D.; Preiss, J. Evidence for independent genetic control of the multiple forms of maize endosperm branching enzymes and starch synthases. Plant Physiol. 1981, 67, 1141–1145. [Google Scholar] [CrossRef] [Green Version]
- Mishra, B.P.; Kumar, R.; Mohan, A.; Gill, K.S. Conservation and divergence of Starch Synthase III genes of monocots and dicots. PLoS ONE 2017, 12, e0189303. [Google Scholar] [CrossRef] [Green Version]
- Crumpton-Taylor, M.; Pike, M.; Lu, K.-J.; Hylton, C.M.; Feil, R.; Eicke, S.; Lunn, J.E.; Zeeman, S.C.; Smith, A.M. Starch synthase 4 is essential for coordination of starch granule formation with chloroplast division during Arabidopsis leaf expansion. New Phytol. 2013, 200, 1064–1075. [Google Scholar] [CrossRef] [PubMed]
- Abt, M.R.; Pfister, B.; Sharma, M.; Eicke, S.; Bürgy, L.; Neale, I.; Seung, D.; Zeeman, S.C. STARCH SYNTHASE5, a Noncanonical Starch Synthase-Like Protein, Promotes Starch Granule Initiation in Arabidopsis. Plant Cell 2020, 32, 2543–2565. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Luo, K.; Li, S.; Peng, D.; Tang, D.; Lu, H.; Zhao, Y.; Lv, C.; Wang, J. Genetic Variation and Sequence Diversity of Starch Biosynthesis and Sucrose Metabolism Genes in Sweet Potato. Agronomy 2020, 10, 627. [Google Scholar] [CrossRef]
- Nabemoto, M.; Watanabe, R.; Ohsu, M.; Sato, K.; Otani, M.; Nakayachi, O.; Watanabe, M. Molecular characterization of genes encoding isoamylase-type debranching enzyme in tuberous root of sweet potato, Ipomoea batatas (L.) Lam. Plant Biotechnol. 2016, 33, 351–359. [Google Scholar] [CrossRef] [Green Version]
- Noda, T.; Kobayashi, T.; Suda, I. Effect of soil temperature on starch properties of sweet potatoes. Carbohydr. Polym. 2001, 44, 239–246. [Google Scholar] [CrossRef]
- Tian, S.; Rickard, J.; Blanshard, J. Physicochemical properties of sweet potato starch. J. Sci. Food Agric. 1991, 57, 459–491. [Google Scholar] [CrossRef]
- Chen, Z.; Schols, H.A.; Voragen, A.G.J. Physicochemical Properties of Starches Obtained from Three Varieties of Chinese Sweet Potatoes. J. Food Sci. 2003, 68, 431–437. [Google Scholar] [CrossRef]
- Toyama, J.; Ishiguro, K.; Noda, T.; Kumagai, T.; Yamakawa, O. Influence of Delayed Harvest Time on Physico-chemical Properties of Sweetpotato Starch. Starch-Stärke 2003, 55, 558–563. [Google Scholar] [CrossRef]
- Tester, R.F.; Morrison, W.R. Swelling and gelatinization of cereal starches. I. Effects of amylopectin, amylose, and lipids. Cereal Chem. 1990, 67, 551–557. [Google Scholar]
- Stevenson, D.G.; Jane, J.-L.; Inglett, G.E. Structures and physicochemical properties of starch from immature seeds of soybean varieties (Glycine max (L.) Merr.) exhibiting normal, low-linolenic or low-saturated fatty acid oil profiles at maturity. Carbohydr. Polym. 2007, 70, 149–159. [Google Scholar] [CrossRef]
- Tortoe, C.; Akonor, P.T.; Koch, K.; Menzel, C.; Adofo, K. Amylose and Amylopectin Molecular Fractions and Chain Length Distribution of Amylopectin in Twelve Varieties of Ghanaian Sweet Potato (Ipomoea batatas) Flours. Int. J. Food Prop. 2017, 20, 3225–3233. [Google Scholar] [CrossRef] [Green Version]
- Jane, J.; Chen, Y.Y.; Lee, L.F.; McPherson, A.E.; Wong, K.S.; Radosavljevic, M.; Kasemsuwan, T. Effects of Amylopectin Branch Chain Length and Amylose Content on the Gelatinization and Pasting Properties of Starch. Cereal Chem. 1999, 76, 629–637. [Google Scholar] [CrossRef]
- Buléon, A.; Colonna, P.; Planchot, V.; Ball, S. Starch granules: Structure and biosynthesis. Int. J. Biol. Macromol. 1998, 23, 85–112. [Google Scholar] [CrossRef] [Green Version]
- Imberty, A.; Chanzy, H.; Perez, S.; Buleon, A.; Tran, V. New three-dimensional structure for A-type starch. Macromolecules 1987, 20, 2634–2636. [Google Scholar] [CrossRef]
- Imberty, A.; Perez, S. A revisit to the three-dimensional structure of B-type starch. Biopolymers 1988, 27, 1205–1221. [Google Scholar] [CrossRef]
- Koroteeva, D.A.; Kiseleva, V.I.; Krivandin, A.V.; Shatalova, O.V.; Błaszczak, W.; Bertoft, E.; Piyachomkwan, K.; Yuryev, V.P. Structural and thermodynamic properties of rice starches with different genetic background: Part 2. Defectiveness of different supramolecular structures in starch granules. Int. J. Biol. Macromol. 2007, 41, 534–547. [Google Scholar] [CrossRef]
- Srichuwong, S.; Sunarti, T.C.; Mishima, T.; Isono, N.; Hisamatsu, M. Starches from different botanical sources I: Contribution of amylopectin fine structure to thermal properties and enzyme digestibility. Carbohydr. Polym. 2005, 60, 529–538. [Google Scholar] [CrossRef]
- Noda, T.; Isono, N.; Krivandin, A.V.; Shatalova, O.V.; Blaszczak, W.; Yuryev, V.P. Origin of defects in assembled supramolecular structures of sweet potato starches with different amylopectin chain-length distribution. Carbohydr. Polym. 2009, 76, 400–409. [Google Scholar] [CrossRef]
- Teerawanichpan, P.; Lertpanyasampatha, M.; Netrphan, S.; Varavinit, S.; Boonseng, O.; Narangajavana, J. Influence of Cassava Storage Root Development and Environmental Conditions on Starch Granule Size Distribution. Starch-Stärke 2008, 60, 696–705. [Google Scholar] [CrossRef]
- Lv, Z.; Yu, K.; Jin, S.; Ke, W.; Fei, C.; Cui, P.; Lu, G. Starch Granules Size Distribution of Sweet Potato and Their Relationship with Quality of Dried and Fried Products. Starch-Stärke 2019, 71, 1800175. [Google Scholar] [CrossRef]
- Zhu, F.; Wang, S. Physicochemical properties, molecular structure, and uses of sweetpotato starch. Trends Food Sci. Technol. 2014, 36, 68–78. [Google Scholar] [CrossRef]
- Tanaka, M.; Takahata, Y.; Nakayama, H.; Nakatani, M.; Tahara, M. Altered carbohydrate metabolism in the storage roots of sweetpotato plants overexpressing the SRF1 gene, which encodes a Dof zinc finger transcription factor. Planta 2009, 230, 737–746. [Google Scholar] [CrossRef]
- Tanaka, M. Recent Progress in Molecular Studies on Storage Root Formation in Sweetpotato (Ipomoea batatas). JARQ-JPN Agric. Res. Q. 2016, 50, 293–299. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Chen, X.; Ma, P.a.; Zhang, S.; Zeng, C.; Jiang, X.; Wang, W. Ethylene Responsive Factor MeERF72 Negatively Regulates Sucrose synthase 1 Gene in Cassava. Int. J. Mol. Sci. 2018, 19, 1281. [Google Scholar] [CrossRef] [Green Version]
- Baguma, Y.; Sun, C.; Boren, M.; Olsson, H.; Rosenqvist, S.; Mutisya, J.; Rubaihayo, P.R.; Jansson, C. Sugar-mediated semidian oscillation of gene expression in the cassava storage root regulates starch synthesis. Plant Signal. Behav. 2008, 3, 439–445. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Chang, L.; Tong, Z.; Wang, D.; Yin, Q.; Wang, D.; Jin, X.; Yang, Q.; Wang, L.; Sun, Y.; et al. Proteomics Profiling Reveals Carbohydrate Metabolic Enzymes and 14-3-3 Proteins Play Important Roles for Starch Accumulation during Cassava Root Tuberization. Sci. Rep. 2016, 6, 19643. [Google Scholar] [CrossRef] [Green Version]
- Ren, Z.-T.; Zhao, H.-Y.; He, S.-Z.; Zhai, H.; Zhao, N.; Liu, Q.-C. Overexpression of IbSnRK1 enhances nitrogen uptake and carbon assimilation in transgenic sweetpotato. J. Integr. Agric. 2018, 17, 296–305. [Google Scholar] [CrossRef] [Green Version]
- Duan, W.; Wang, Q.; Zhang, H.; Xie, B.; Li, A.; Hou, F.; Dong, S.; Wang, B.; Qin, Z.; Zhang, L. Differences between nitrogen-tolerant and nitrogen-susceptible sweetpotato cultivars in photosynthate distribution and transport under different nitrogen conditions. PLoS ONE 2018, 13, e0194570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Yang, J.; Zhang, M.; Fan, W.; Firon, N.; Pattanaik, S.; Yuan, L.; Zhang, P. Altered Phenylpropanoid Metabolism in the Maize Lc-Expressed Sweet Potato (Ipomoea batatas) Affects Storage Root Development. Sci. Rep. 2016, 6, 18645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, S.H.; Shen, H.M.; Wu, J.H. Role of calonyctin on free sugars in relation to starch accumulation in developing sweet potatoes. J. Plant Growth Regul. 1996, 15, 27–31. [Google Scholar] [CrossRef]
- Zhu, F. Composition, structure, physicochemical properties, and modifications of cassava starch. Carbohydr. Polym. 2015, 122, 456–480. [Google Scholar] [CrossRef]
- Dupuis, J.H.; Liu, Q. Potato Starch: A Review of Physicochemical, Functional and Nutritional Properties. Am. J. Potato Res. 2019, 96, 127–138. [Google Scholar] [CrossRef]
- Juhász, R.; Salgó, A. Pasting behavior of amylose, amylopectin and their mixtures as determined by RVA curves and first derivatives. Starch-Stärke 2008, 60, 70–78. [Google Scholar] [CrossRef]
- Batey, I. Interpretation of RVA Curves; American Association of Cereal Chemists, Inc. (AACC): St. Paul, MN, USA, 2007; pp. 19–30. [Google Scholar]
- Katayama, K.; Komae, K.; Kohyama, K.; Kato, T.; Tamiya, S.; Komaki, K. New Sweet Potato Line having low Gelatinization Temperature and altered Starch Structure. Starch-Starke 2002, 54, 51–57. [Google Scholar] [CrossRef]
- Genkina, N.K.; Noda, T.; Koltisheva, G.I.; Wasserman, L.A.; Tester, R.F.; Yuryev, V.P. Effects of growth temperature on some structural properties of crystalline lamellae in starches extracted from sweet potatoes (Sunnyred and Ayamurasaki). Starch-Starke 2003, 55, 350–357. [Google Scholar] [CrossRef]
- Ahmed, S.; Zhou, X.; Pang, Y.; Xu, Y.; Tong, C.; Bao, J. Genetic diversity of potato genotypes estimated by starch physicochemical properties and microsatellite markers. Food Chem. 2018, 257, 368–375. [Google Scholar] [CrossRef]
- Tong, C.; Ru, W.; Wu, L.; Wu, W.; Bao, J. Fine structure and relationships with functional properties of pigmented sweet potato starches. Food Chem. 2020, 311, 126011. [Google Scholar] [CrossRef] [PubMed]
- McGrane, S.J.; Mainwaring, D.E.; Cornell, H.J.; Rix, C.J. The role of hydrogen bonding in amylose gelation. Starch-Starke 2004, 56, 122–131. [Google Scholar] [CrossRef]
- Srichuwong, S.; Jane, J.-l. Physicochemical properties of starch affected by molecular composition and structures: A review. Food Sci. Biotechnol. 2007, 16, 663. [Google Scholar]
- Zhu, L.-J.; Liu, Q.-Q.; Wilson, J.D.; Gu, M.-H.; Shi, Y.-C. Digestibility and physicochemical properties of rice (Oryza sativa L.) flours and starches differing in amylose content. Carbohydr. Polym. 2011, 86, 1751–1759. [Google Scholar] [CrossRef]
- Geigenberger, P. Regulation of starch biosynthesis in response to a fluctuating environment. Plant Physiol. 2011, 155, 1566–1577. [Google Scholar] [CrossRef] [Green Version]
- Lee, B.-H.; Lee, Y.-T. Physicochemical and structural properties of different colored sweet potato starches. Starch-Stärke 2017, 69, 1600001. [Google Scholar] [CrossRef]
- Tester, R.F.; Karkalas, J.; Qi, X. Starch—Composition, fine structure and architecture. J. Cereal Sci. 2004, 39, 151–165. [Google Scholar] [CrossRef]
- Rolland-Sabaté, A.; Sánchez, T.; Buléon, A.; Colonna, P.; Jaillais, B.; Ceballos, H.; Dufour, D. Structural characterization of novel cassava starches with low and high-amylose contents in comparison with other commercial sources. Food Hydrocoll. 2012, 27, 161–174. [Google Scholar] [CrossRef]
- Vamadevan, V.; Bertoft, E. Structure-function relationships of starch components. Starch-Stärke 2015, 67, 55–68. [Google Scholar] [CrossRef]
- Srichuwong, S.; Orikasa, T.; Matsuki, J.; Shiina, T.; Kobayashi, T.; Tokuyasu, K. Sweet potato having a low temperature-gelatinizing starch as a promising feedstock for bioethanol production. Biomass Bioenergy 2012, 39, 120–127. [Google Scholar] [CrossRef]
- Blennow, A.; Engelsen, S.B.; Nielsen, T.H.; Baunsgaard, L.; Mikkelsen, R. Starch phosphorylation: A new front line in starch research. Trends Plant Sci. 2002, 7, 445–450. [Google Scholar] [CrossRef]
- Mahlow, S.; Orzechowski, S.; Fettke, J. Starch phosphorylation: Insights and perspectives. Cell. Mol. Life Sci. 2016, 73, 2753–2764. [Google Scholar] [CrossRef]
- Carpenter, M.A.; Joyce, N.I.; Genet, R.A.; Cooper, R.D.; Murray, S.R.; Noble, A.D.; Butler, R.C.; Timmerman-Vaughan, G.M. Starch phosphorylation in potato tubers is influenced by allelic variation in the genes encoding glucan water dikinase, starch branching enzymes I and II, and starch synthase III. Front. Plant Sci. 2015, 6, 143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- You, Y.; Zhang, M.; Yang, W.; Li, C.; Liu, Y.; Li, C.; He, J.; Wu, W. Starch phosphorylation and the in vivo regulation of starch metabolism and characteristics. Int. J. Biol. Macromol. 2020, 159, 823–831. [Google Scholar] [CrossRef]
- Baunsgaard, L.; Lütken, H.; Mikkelsen, R.; Glaring, M.A.; Pham, T.T.; Blennow, A. A novel isoform of glucan, water dikinase phosphorylates pre-phosphorylated α-glucans and is involved in starch degradation in Arabidopsis. Plant J. 2005, 41, 595–605. [Google Scholar] [CrossRef]
- Edner, C.; Li, J.; Albrecht, T.; Mahlow, S.; Hejazi, M.; Hussain, H.; Kaplan, F.; Guy, C.; Smith, S.M.; Steup, M.; et al. Glucan, Water Dikinase Activity Stimulates Breakdown of Starch Granules by Plastidial beta-amylases. Plant Physiol. 2007, 145, 17. [Google Scholar] [CrossRef] [Green Version]
- Kötting, O.; Pusch, K.; Tiessen, A.; Geigenberger, P.; Steup, M.; Ritte, G. Identification of a Novel Enzyme Required for Starch Metabolism in Arabidopsis Leaves. The Phosphoglucan, Water Dikinase. Plant Physiol. 2005, 137, 242–252. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Dees, D.; Dechesne, A.; Huang, X.-F.; Visser, R.G.F.; Trindade, L.M. Starch phosphorylation plays an important role in starch biosynthesis. Carbohydr. Polym. 2017, 157, 1628–1637. [Google Scholar] [CrossRef]
- Zhou, W.; He, S.; Naconsie, M.; Ma, Q.; Zeeman, S.C.; Gruissem, W.; Zhang, P. Alpha-Glucan, Water Dikinase 1 Affects Starch Metabolism and Storage Root Growth in Cassava (Manihot esculenta Crantz). Sci. Rep. 2017, 7, 9863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kötting, O.; Santelia, D.; Edner, C.; Eicke, S.; Marthaler, T.; Gentry, M.S.; Comparot-Moss, S.; Chen, J.; Smith, A.M.; Steup, M.; et al. STARCH-EXCESS4 Is a Laforin-Like Phosphoglucan Phosphatase Required for Starch Degradation in Arabidopsis thaliana. Plant Cell 2009, 21, 334–346. [Google Scholar] [CrossRef] [Green Version]
- Hejazi, M.; Fettke, J.; Kotting, O.; Zeeman, S.C.; Steup, M. The Laforin-Like Dual-Specificity Phosphatase SEX4 from Arabidopsis Hydrolyzes Both C6-and C3-Phosphate Esters Introduced by Starch-Related Dikinases and Thereby Affects Phase Transition of alpha-Glucans. Plant Physiol. 2010, 152, 711–722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gentry, M.S.; Dowen, R.H., III; Worby, C.A.; Mattoo, S.; Ecker, J.R.; Dixon, J.E. The phosphatase laforin crosses evolutionary boundaries and links carbohydrate metabolism to neuronal disease. J. Cell Biol. 2007, 178, 477–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Hostettler, C.E.; Damberger, F.F.; Kossmann, J.; Lloyd, J.R.; Zeeman, S.C. Modification of Cassava Root Starch Phosphorylation Enhances Starch Functional Properties. Front. Plant Sci. 2018, 9, 1562. [Google Scholar] [CrossRef] [PubMed]
- Raigond, P.; Ezekiel, R.; Raigond, B. Resistant starch in food: A review. J. Sci. Food Agric. 2015, 95, 1968–1978. [Google Scholar] [CrossRef]
- Fallahi, P.; Mussoline, W.A.; Athmanathan, A.; Trupia, S.; Wilkie, A.C. Potential Value-Added Products from Industrial Sweetpotato Syrup Processing. Ind. Biotechnol. 2016, 12, 343–349. [Google Scholar] [CrossRef]
- Sagnelli, D.; Hebelstrup, K.H.; Leroy, E.; Rolland-Sabaté, A.; Guilois, S.; Kirkensgaard, J.J.K.; Mortensen, K.; Lourdin, D.; Blennow, A. Plant-crafted starches for bioplastics production. Carbohydr. Polym. 2016, 152, 398–408. [Google Scholar] [CrossRef]
- Lauer, M.K.; Smith, R.C. Recent advances in starch-based films toward food packaging applications: Physicochemical, mechanical, and functional properties. Compr. Rev. Food Sci. Food Saf. 2020, 19, 3031–3083. [Google Scholar] [CrossRef]
- Colussi, R.; Pinto, V.Z.; El Halal, S.L.M.; Biduski, B.; Prietto, L.; Castilhos, D.D.; Zavareze, E.d.R.; Dias, A.R.G. Acetylated rice starches films with different levels of amylose: Mechanical, water vapor barrier, thermal, and biodegradability properties. Food Chem. 2017, 221, 1614–1620. [Google Scholar] [CrossRef]
- Thakur, R.; Pristijono, P.; Scarlett, C.J.; Bowyer, M.; Singh, S.P.; Vuong, Q.V. Starch-based films: Major factors affecting their properties. Int. J. Biol. Macromol. 2019, 132, 1079–1089. [Google Scholar] [CrossRef]
- Cano, A.; Jiménez, A.; Cháfer, M.; Gónzalez, C.; Chiralt, A. Effect of amylose:amylopectin ratio and rice bran addition on starch films properties. Carbohydr. Polym. 2014, 111, 543–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGlashan, S.A.; Halley, P.J. Preparation and characterisation of biodegradable starch-based nanocomposite materials. Polym. Int. 2003, 52, 1767–1773. [Google Scholar] [CrossRef]
- Shen, X.L.; Wu, J.M.; Chen, Y.; Zhao, G. Antimicrobial and physical properties of sweet potato starch films incorporated with potassium sorbate or chitosan. Food Hydrocoll. 2010, 24, 285–290. [Google Scholar] [CrossRef]
- Kumar, S.; Mukherjee, A.; Dutta, J. Chitosan based nanocomposite films and coatings: Emerging antimicrobial food packaging alternatives. Trends Food Sci. Technol. 2020, 97, 196–209. [Google Scholar] [CrossRef]
- Yu, M.; Ji, N.; Wang, Y.; Dai, L.; Xiong, L.; Sun, Q. Starch-based nanoparticles: Stimuli responsiveness, toxicity, and interactions with food components. Compr. Rev. Food Sci. Food Saf. 2021, 20, 1075–1100. [Google Scholar] [CrossRef]
- Oh, S.-M.; Lee, B.-H.; Seo, D.-H.; Choi, H.-W.; Kim, B.-Y.; Baik, M.-Y. Starch nanoparticles prepared by enzymatic hydrolysis and self-assembly of short-chain glucans. Food Sci. Biotechnol. 2020, 29, 585–598. [Google Scholar] [CrossRef] [PubMed]
- Odeniyi, M.A.; Omoteso, O.A.; Adepoju, A.O.; Jaiyeoba, K.T. Starch nanoparticles in drug delivery: A review. Polim. Med. 2018, 48, 41–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Q.; Gong, M.; Li, Y.; Xiong, L. Effect of retrogradation time on preparation and characterization of proso millet starch nanoparticles. Carbohydr. Polym. 2014, 111, 133–138. [Google Scholar] [CrossRef]
- Šárka, E.; Dvořáček, V. Waxy starch as a perspective raw material (a review). Food Hydrocoll. 2017, 69, 402–409. [Google Scholar] [CrossRef]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A Programmable Dual-RNA–Guided DNA Endonuclease in Adaptive Bacterial Immunity. Science 2012, 337, 816–826. [Google Scholar] [CrossRef]
- Korotkova, A.M.; Gerasimova, S.V.; Shumny, V.K.; Khlestkina, E.K. Crop genes modified using the CRISPR/Cas system. Russ. J. Genet. Appl. Res. 2017, 7, 822–832. [Google Scholar] [CrossRef]
- Manghwar, H.; Lindsey, K.; Zhang, X.; Jin, S. CRISPR/Cas System: Recent Advances and Future Prospects for Genome Editing. Trends Plant Sci. 2019, 24, 1102–1125. [Google Scholar] [CrossRef] [Green Version]
- Metje-Sprink, J.; Menz, J.; Modrzejewski, D.; Sprink, T. DNA-Free Genome Editing: Past, Present and Future. Front. Plant Sci. 2019, 9, 1957. [Google Scholar] [CrossRef]
- Zhu, H.; Li, C.; Gao, C. Applications of CRISPR–Cas in agriculture and plant biotechnology. Nat. Rev. Mol. Cell Biol. 2020, 21, 661–677. [Google Scholar] [CrossRef] [PubMed]
- Anzalone, A.V.; Randolph, P.B.; Davis, J.R.; Sousa, A.A.; Koblan, L.W.; Levy, J.M.; Chen, P.J.; Wilson, C.; Newby, G.A.; Raguram, A.; et al. Search-and-replace genome editing without double-strand breaks or donor DNA. Nature 2019, 576, 149–157. [Google Scholar] [CrossRef]
- Anzalone, A.V.; Koblan, L.W.; Liu, D.R. Genome editing with CRISPR–Cas nucleases, base editors, transposases and prime editors. Nat. Biotechnol. 2020, 38, 824–844. [Google Scholar] [CrossRef]
- Botticella, E.; Sestili, F.; Sparla, F.; Moscatello, S.; Marri, L.; Cuesta-Seijo, J.A.; Falini, G.; Battistelli, A.; Trost, P.; Lafiandra, D. Combining mutations at genes encoding key enzymes involved in starch synthesis affects the amylose content, carbohydrate allocation and hardness in the wheat grain. Plant Biotechnol. J. 2018, 16, 1723–1734. [Google Scholar] [CrossRef] [Green Version]
- Minkenberg, B.; Wheatley, M.; Yang, Y. Chapter Seven—CRISPR/Cas9-Enabled Multiplex Genome Editing and Its Application. In Progress in Molecular Biology and Translational Science; Weeks, D.P., Yang, B., Eds.; Academic Press: Cambridge, MA, USA, 2017; Volume 149, pp. 111–132. [Google Scholar]
- Walkowiak, S.; Gao, L.; Monat, C.; Haberer, G.; Kassa, M.T.; Brinton, J.; Ramirez-Gonzalez, R.H.; Kolodziej, M.C.; Delorean, E.; Thambugala, D.; et al. Multiple wheat genomes reveal global variation in modern breeding. Nature 2020, 588, 277–283. [Google Scholar] [CrossRef]
- Xu, X.; Pan, S.; Cheng, S.; Zhang, B.; Mu, D.; Ni, P.; Zhang, G.; Yang, S.; Li, R.; Wang, J.; et al. Genome sequence and analysis of the tuber crop potato. Nature 2011, 475, 189–195. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Jayarathna, S.; Turesson, H.; Fält, A.-S.; Nestor, G.; González, M.N.; Olsson, N.; Beganovic, M.; Hofvander, P.; Andersson, R.; et al. Amylose starch with no detectable branching developed through DNA-free CRISPR-Cas9 mediated mutagenesis of two starch branching enzymes in potato. Sci. Rep. 2021, 11, 4311. [Google Scholar] [CrossRef] [PubMed]
- McKenzie, M.J.; Chen, R.K.Y.; Harris, J.C.; Ashworth, M.J.; Brummell, D.A. Post-translational regulation of acid invertase activity by vacuolar invertase inhibitor affects resistance to cold-induced sweetening of potato tubers. Plant Cell Environ. 2013, 36, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Datir, S.S. Invertase inhibitors in potato: Towards a biochemical and molecular understanding of cold-induced sweetening. Crit. Rev. Food Sci. Nutr. 2020, 1–15. [Google Scholar] [CrossRef]
- Park, S.H.; Na, Y.; Kim, J.; Kang, S.D.; Park, K.-H. Properties and applications of starch modifying enzymes for use in the baking industry. Food Sci. Biotechnol. 2017, 27, 299–312. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.P.; Newby, G.A.; Liu, D.R. Precision genome editing using cytosine and adenine base editors in mammalian cells. Nat. Protoc. 2021, 16, 1089–1128. [Google Scholar] [CrossRef] [PubMed]
- Isobe, S.; Shirasawa, K.; Hirakawa, H. Current status in whole genome sequencing and analysis of Ipomoea spp. Plant Cell Rep. 2019, 38, 1365–1371. [Google Scholar] [CrossRef] [Green Version]
- Torti, S.; Schlesier, R.; Thümmler, A.; Bartels, D.; Römer, P.; Koch, B.; Werner, S.; Panwar, V.; Kanyuka, K.; Wirén, N.v.; et al. Transient reprogramming of crop plants for agronomic performance. Nat. Plants 2021, 7, 159–171. [Google Scholar] [CrossRef]
- Santa-Maria, M.; Pecota, K.V.; Yencho, C.G.; Allen, G.; Sosinski, B. Rapid shoot regeneration in industrial ‘high starch’ sweetpotato (Ipomoea batatas L.) genotypes. Plant Cell Tissue Organ Cult. (PCTOC) 2009, 97, 109–117. [Google Scholar] [CrossRef]
- Ren, Q.; Sretenovic, S.; Liu, S.; Tang, X.; Huang, L.; He, Y.; Liu, L.; Guo, Y.; Zhong, Z.; Liu, G.; et al. PAM-less plant genome editing using a CRISPR–SpRY toolbox. Nat. Plants 2021, 7, 25–33. [Google Scholar] [CrossRef]
- Han, Z.; Shi, R.; Sun, D.-W. Effects of novel physical processing techniques on the multi-structures of starch. Trends Food Sci. Technol. 2020, 97, 126–135. [Google Scholar] [CrossRef]
- Ojogbo, E.; Ogunsona, E.O.; Mekonnen, T.H. Chemical and physical modifications of starch for renewable polymeric materials. Mater. Today Sustain. 2020, 7–8, 100028. [Google Scholar] [CrossRef]
- Kumar, P.; Prakash, K.S.; Jan, K.; Swer, T.L.; Jan, S.; Verma, R.; Deepika, K.; Dar, M.Z.; Verma, K.; Bashir, K. Effects of gamma irradiation on starch granule structure and physicochemical properties of brown rice starch. J. Cereal Sci. 2017, 77, 194–200. [Google Scholar] [CrossRef]
- Tao, Y.; Yan, B.; Fan, D.; Zhang, N.; Ma, S.; Wang, L.; Wu, Y.; Wang, M.; Zhao, J.; Zhang, H. Structural changes of starch subjected to microwave heating: A review from the perspective of dielectric properties. Trends Food Sci. Technol. 2020, 99, 593–607. [Google Scholar] [CrossRef]
- Thirumdas, R.; Trimukhe, A.; Deshmukh, R.R.; Annapure, U.S. Functional and rheological properties of cold plasma treated rice starch. Carbohydr. Polym. 2017, 157, 1723–1731. [Google Scholar] [CrossRef] [PubMed]
- Rönspies, M.; Dorn, A.; Schindele, P.; Puchta, H. CRISPR–Cas-mediated chromosome engineering for crop improvement and synthetic biology. Nat. Plants 2021, 7, 566–573. [Google Scholar] [CrossRef]
- Shoeb, E.; Badar, U.; Venkataraman, S.; Hefferon, K. Chapter 10—CRISPR/Cas9 and Cas13a systems: A promising tool for plant breeding and plant defence. In CRISPR and RNAi Systems; Abd-Elsalam, K.A., Lim, K.-T., Eds.; Elsevier: Amsterdam, The Netherlands, 2021; pp. 211–231. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Technique | Cultivar | Improved Trait/Observation | Reference |
|---|---|---|---|---|
| GBSSI | Co-suppression; RNAi | Kokei-14 | Reduction of amylose content | [18] |
| Tuberous roots free of amylose generated in transgenic plants, compared to 17–18% in wild-type plants (WT). | ||||
| Amylose-free starch obtained from transgenic plants. | [19] | |||
| Decreased content of DP 6–8 amylopectin. | ||||
| The amylose-free starch exhibits altered values of gelatinization properties, with increased To at 71 °C, Tp at 74 °C, and ΔH at 17.2 J/g. | ||||
| Irregular pasting curve was observed in amylose-free starch | ||||
| Amylose-free transgenic plants | [20] | |||
| No significant difference in storage root yield between transgenic and non-transgenic plants | ||||
| Amylose-free; lacking long chain amylopectin (DP > 100) and decreased DP 6–7 amylopectin in transgenic plants | [21] | |||
| RNAi | Xushu-22 (High-starch) | Amylose content in transgenic (waxy) lines decreased to 5.7% from 30.4% (Xushu22) | [22] | |
| Larger granule size was observed in waxy starch from transgenic plants. | ||||
| - Fewer short chain of amylopectin CLDs. | ||||
| The waxy transgenic plants show weak Type II distribution compared to high amylose starch | [23] | |||
| Large decrease in amylose and increase in long chain amylopectin. | ||||
| CRISPR/Cas9 | Xushu-22 | Lower amylose content (5.8–22.4%) in mutant lines compared to WT (27.2%) | [16] | |
| Taizhong-6 (Carotenoid rich) | Lower amylose content (5.5–14.8%) in mutant lines compared to WT (25.7%) | |||
| SSI | Overexpression and RNAi | Lizixiang (Low starch) | Increased starch content and granule size and reduced the proportion of amylose in IbSSI overexpressing plants. Starch content reduced in RNAi plants. | [7] |
| SSII | RNAi | Quick Sweet | Lower pasting temperature (Pte) and breakdown (BD) of starch from transgenic plants compared to WT. | [17] |
| Higher amylopectin at DP 6–11 but lower at DP 13–25. | ||||
| Cracking on the hilum was observed in starch granules from mutants. | ||||
| SBEII | RNAi | Kokei-14 | Higher in amylose content but slightly lower in starch yield, and showed different shapes of starch grains compared to WT. | [24] |
| Increased levels of amylose, phosphate contents, amylopectin at DP12–15 and DP24–33, and altered crystalline structure. Decreased amylopectin at DP >100, DP6–11, compared to WT. | [25] | |||
| CRISPR/Cas9 | Xushu-22 | Higher amylose content (38.0–40.3%) compared to WT (27.2%) | [16] | |
| Fewer short chain amylopectin (DP6–12) | ||||
| Taizhong-6 | Higher amylose content (37.4–37.8%) compared to WT (25.7%) | [16] | ||
| SBEI SBEII | RNAi | Xushu-22 | High amylose content in transgenic plants (50.3–65.5%). | [22] |
| Larger granule size in high-amylose starch (90 µm) compared to WT (5–60 µm). | ||||
| Less short chain amylopectin compared to WT. | ||||
| Higher PTe (85–89 °C) compared to WT (80 °C). | ||||
| Lower BD (58.0–75.7 cP) and higher setback (SB) (911–1810 cP) compared to WT (BD = 403 cP, SB = 425 cP). | ||||
| Lower gelatinization level and higher retrogradation ability compared to WT. | ||||
| Increased Type II semicrystalline lamellae. | [23] | |||
| Chain length distributions (CLDs) of the high-amylose composes of three fractions, DP 32, DP 33–200, and the combination of first and second fractions. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lyu, R.; Ahmed, S.; Fan, W.; Yang, J.; Wu, X.; Zhou, W.; Zhang, P.; Yuan, L.; Wang, H. Engineering Properties of Sweet Potato Starch for Industrial Applications by Biotechnological Techniques including Genome Editing. Int. J. Mol. Sci. 2021, 22, 9533. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179533
Lyu R, Ahmed S, Fan W, Yang J, Wu X, Zhou W, Zhang P, Yuan L, Wang H. Engineering Properties of Sweet Potato Starch for Industrial Applications by Biotechnological Techniques including Genome Editing. International Journal of Molecular Sciences. 2021; 22(17):9533. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179533
Chicago/Turabian StyleLyu, Ruiqing, Sulaiman Ahmed, Weijuan Fan, Jun Yang, Xiaoyun Wu, Wenzhi Zhou, Peng Zhang, Ling Yuan, and Hongxia Wang. 2021. "Engineering Properties of Sweet Potato Starch for Industrial Applications by Biotechnological Techniques including Genome Editing" International Journal of Molecular Sciences 22, no. 17: 9533. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179533