
Neuroplasticity and Multilevel System of Connections Determine the Integrative Role of Nucleus Accumbens in the Brain Reward System

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Structure and Function of Nucleus Accumbens
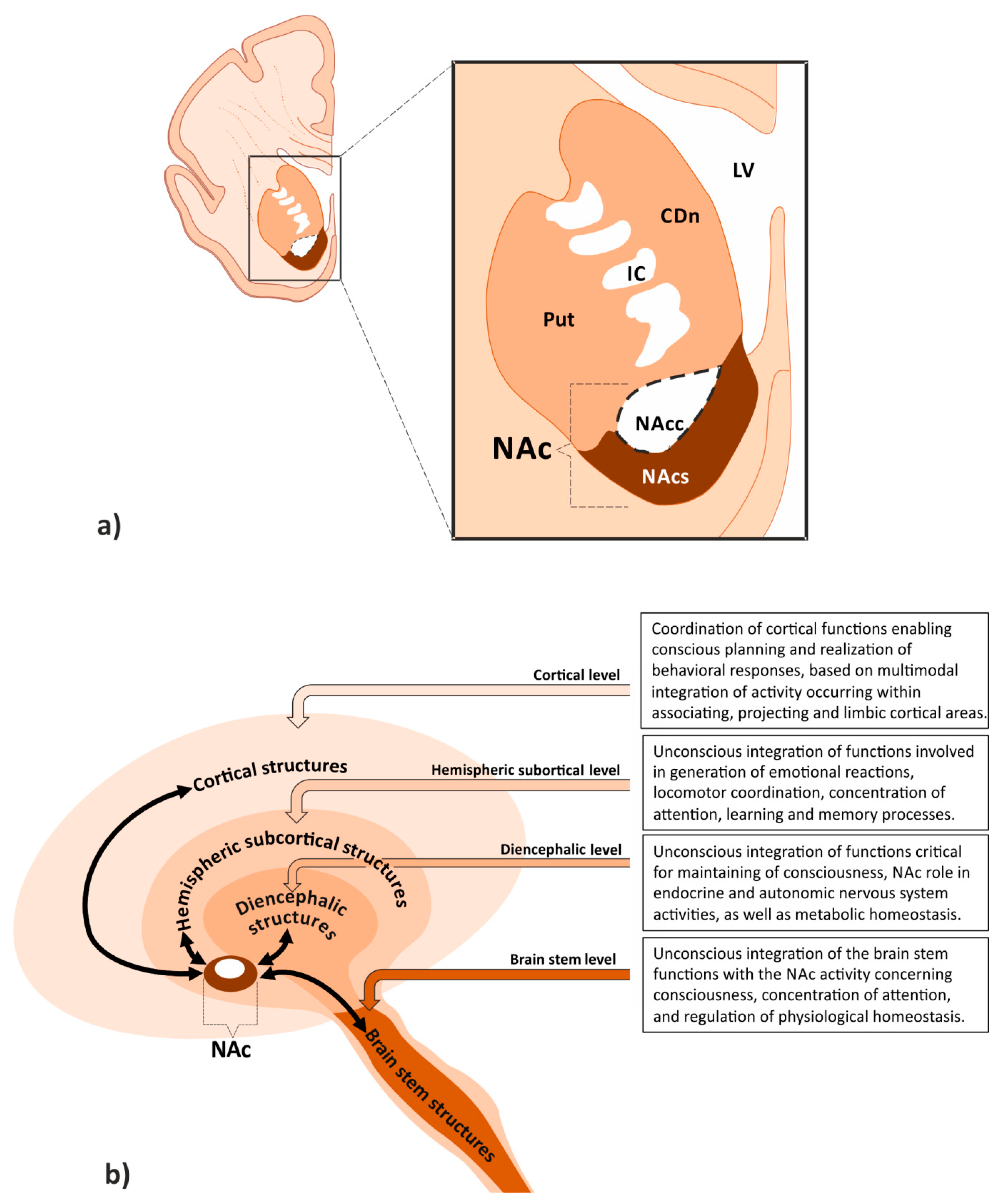
1.1. Localization of Nucleus Accumbens on the Border between Motor and Limbic Areas Suggests Its Integrative Role in the Brain Reward System
1.2. Morphological and Molecular Characteristics of the Accumbal Neurons Determine Their Integrative Role in the Brain Reward System
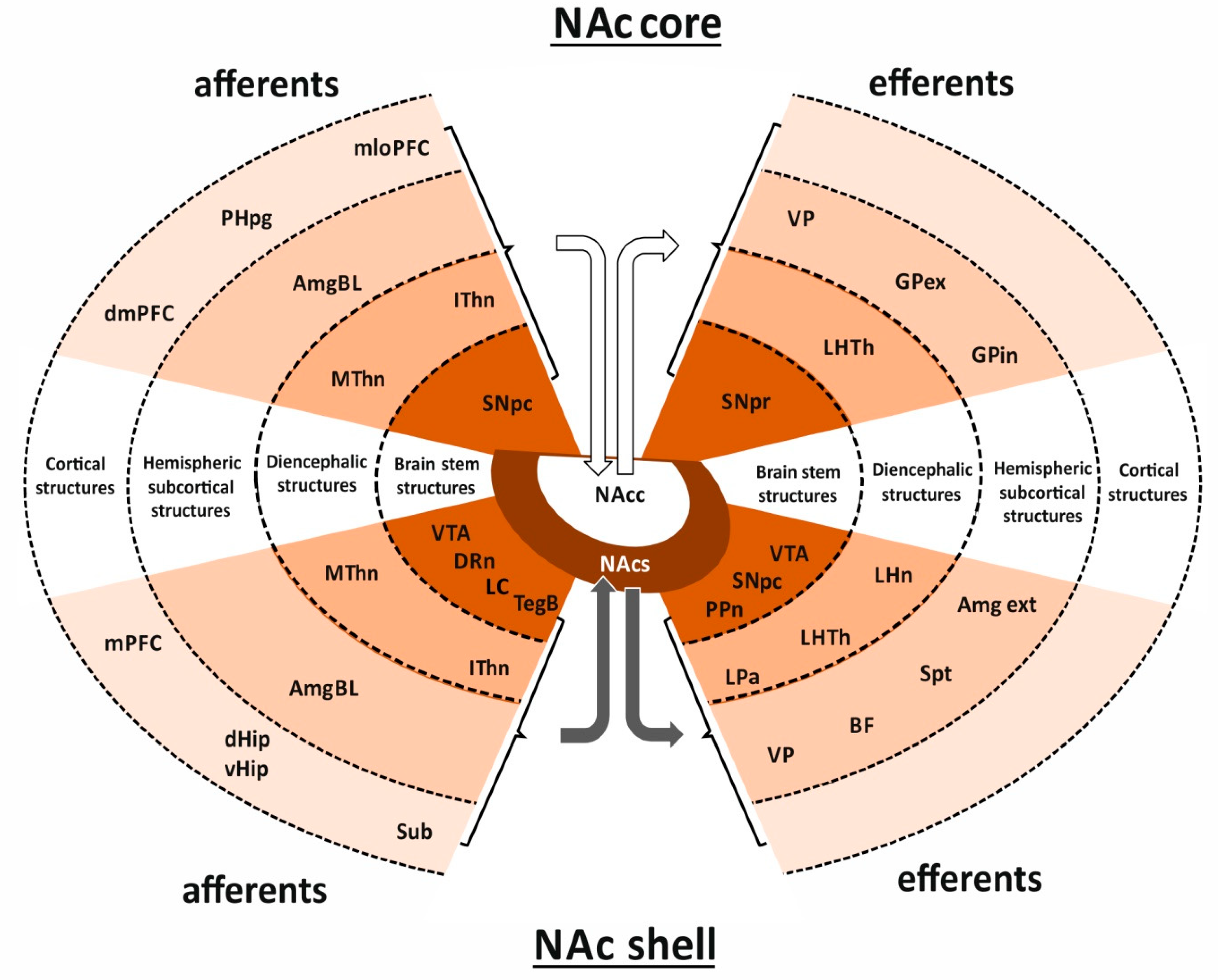
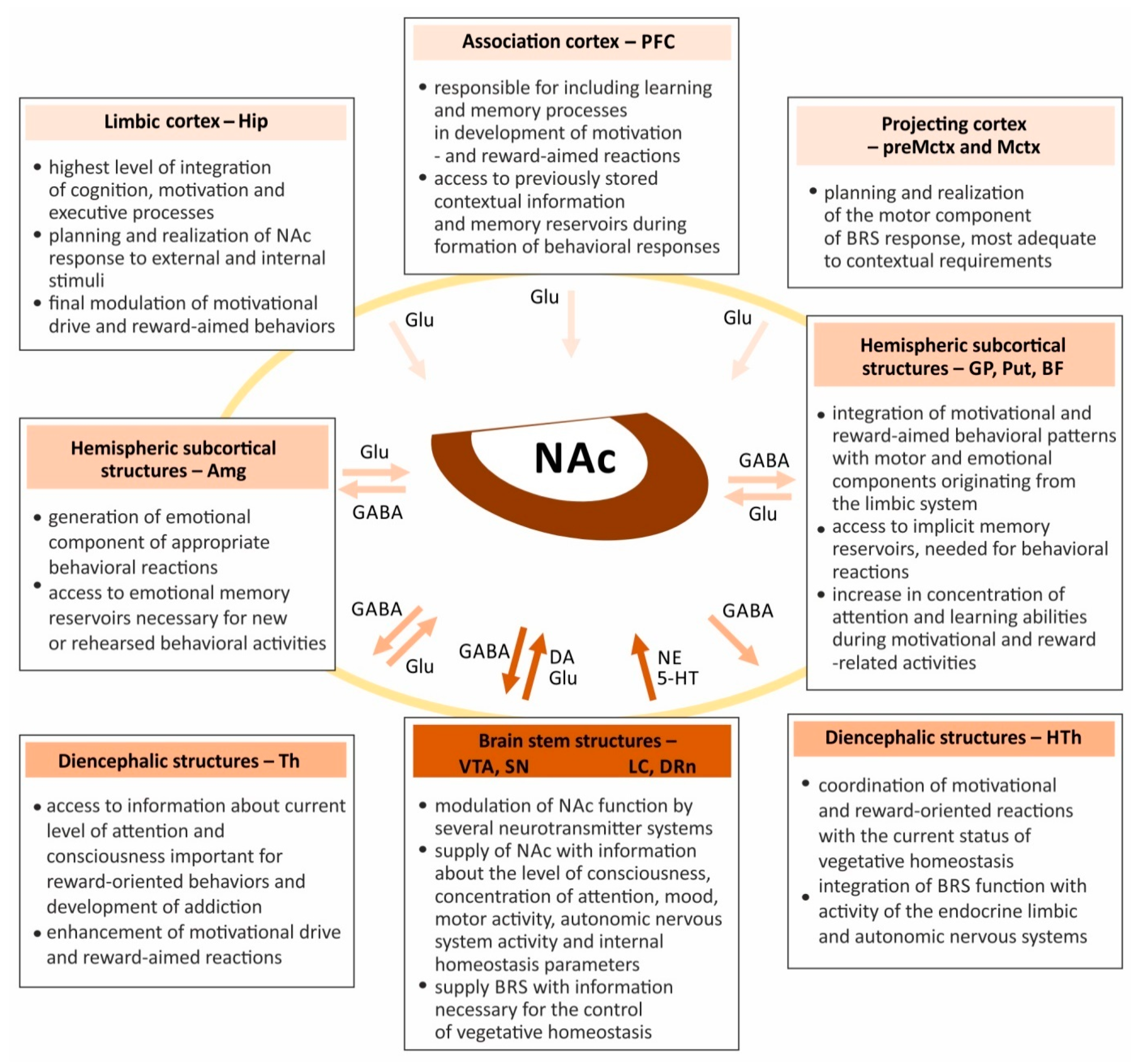
1.3. Nucleus Accumbens Connections Enable Coordination of Limbic, Motor and Vegetative Functions
1.4. Integrative Role of the Nucleus Accumbens Requires Cooperation of Several Neurotransmitter Systems and Receptors, Which Modulate Synaptic Plasticity and Determine the Effects of Drugs on Behavioral Responses
1.4.1. Dopaminergic System
1.4.2. Glutamatergic System
1.4.3. GABA-ergic System
1.4.4. Serotoninergic System
1.4.5. Noradrenergic System
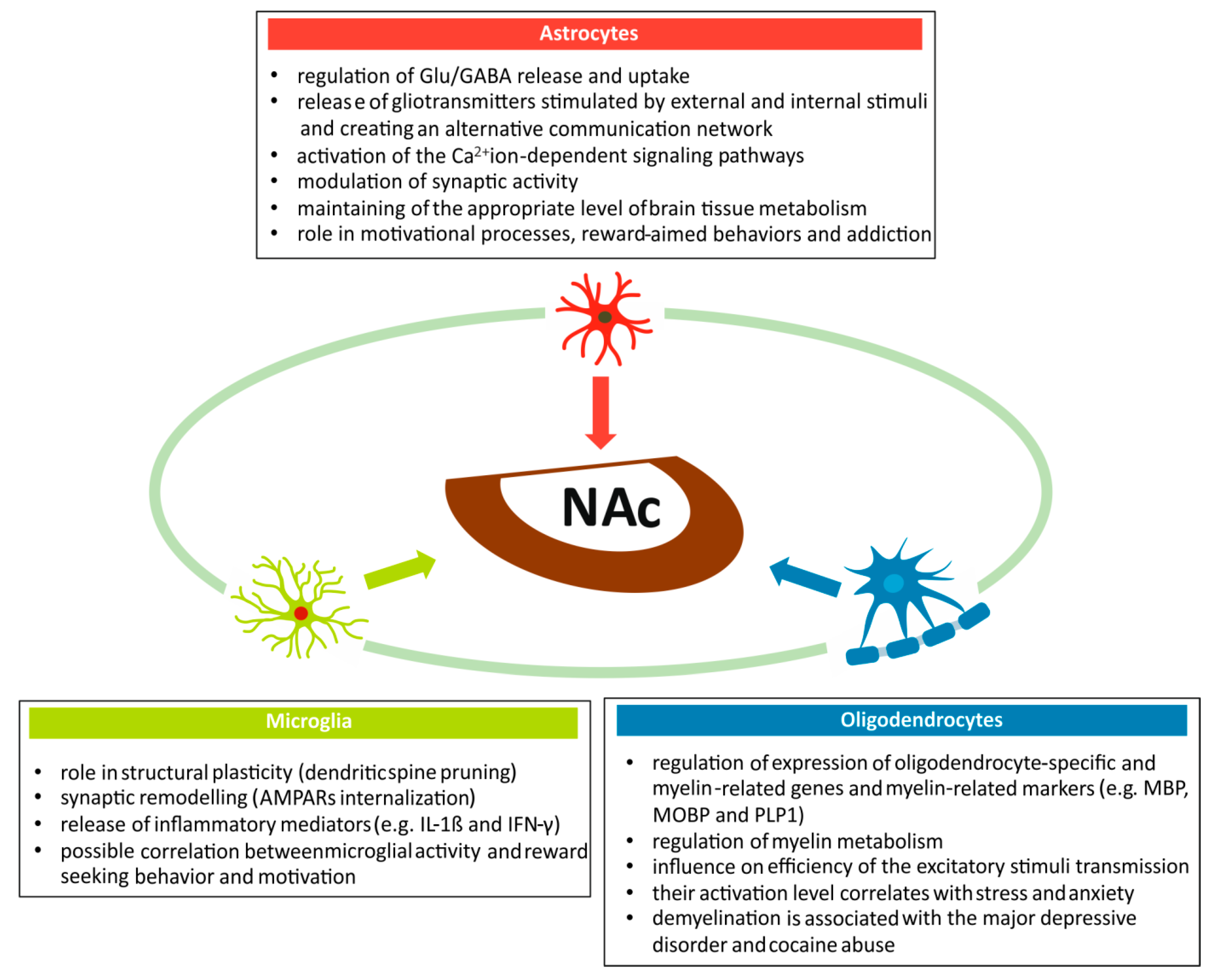
2. Neuroglia Participates in a Wide Spectrum of Physiological and Pathological Processes within the Nucleus Accumbens
3. Nucleus Accumbens Is Responsible for Executive Behaviors Aimed at Motivation, Survival, and Reward Achievement
4. Stress, Psychostimulants and Experience Impact NAc Function during Early Development and Adolescence
4.1. Mechanisms and Effects of Stress Acting on Nucleus Accumbens during Early Development and Adolescence
4.2. Mechanisms and Effects of Nucleus Accumbens Exposition to Addictive Substances during Development
4.3. Reward Mechanisms in Adolescence—Role of the Neurotransmitters and Neurotrophic Factors
4.4. Influence of Learning and Gained Experience on the NAc Function in Adolescence
5. NAc Participates in Elaboration of Aversive Reactions
6. Mechanisms of Neuroplasticity within Nucleus Accumbens
6.1. Morphological Changes in the Dendritic Tree and Dendritic Spines of the Accumbal Neurons Are Triggered by Both External and Internal Stimuli
6.2. Mechanisms of Synaptic Neuroplasticity within the Spino-Dendritic System of the Reward-Related Brain Areas Require Changes in Gene Expression and Activation of Specific Signaling Pathways
7. Role of Neurotransmitters Transporters and BDNF in Synaptic Plasticity within Nucleus Accumbens
7.1. Neurotransmitter Transporters
7.2. DA Transporter (DAT)
7.2.1. Glu Transporters
7.2.2. GABA Transporters
7.2.3. 5-HT Transporter (SERT)
7.2.4. NE Transporter
7.2.5. DAT and SERT Knock-Out Models in Studies on NAc and BRS
7.3. Brain-Derived Neurotrophic Factor
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| α1AR | α1-adrenergic receptor |
| α2AR | α2-adrenergic receptor |
| Amg | amygdala |
| AmgBL | basolateral part of amygdala |
| Amgex | extended amygdala |
| AMPARs | α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid-type glutamate receptors |
| BDNF | brain-derived neurotrophic factor |
| BF | basal forebrain |
| BRS | brain reward system |
| CDn | caudate nucleus |
| cAMP | cyclic adenosine monophosphate |
| CREB | cAMP-response element-binding protein |
| DA | dopamine |
| dHip | dorsal hippocampus |
| dmPFC | dorso-medial prefrontal cortex |
| D1R | D1 dopamine receptor |
| D2R | D2 dopamine receptor |
| DRn | dorsal raphe nucleus |
| ERK | extracellular signal-regulated kinase |
| GABA | γ-aminobutyric acid |
| GDP | guanosine diphosphate |
| Glu | glutamate |
| GP | globus pallidus |
| GPex | external globus pallidus |
| GPin | internal globus pallidus |
| GTP | guanosine triphosphate |
| Hip | hippocampus |
| 5-HT | serotonin |
| HTh | hypothalamus |
| IFN-γ | interferon gamma |
| IL-1β | interleukin-1 beta |
| IThn | intralaminar thalamic nuclei |
| LC | locus coeruleus |
| LTP | long-term potentiation |
| LHn | lateral habenular nucleus |
| LHTh | lateral hypothalamus |
| LPa | lateral preoptic area |
| LV | lateral ventricle |
| MBP | myelin basic protein |
| Mctx | motor cortex |
| MDD | major depressive disorder |
| MDThn | mediodorsal thalamic nucleus |
| mloPFC | medial and lateral orbital prefrontal cortex |
| MOBP | myelin-associated oligodendrocyte basic protein |
| mPFC | medial prefrontal cortex |
| MThn | midline thalamic nuclei |
| NAc | nucleus accumbens |
| NAcc | core of nucleus accumbens |
| NAcs | shell of nucleus accumbens |
| NE | norepinephrine |
| NF-κB | nuclear factor-kappa B |
| NPAS2 | neuronal PAS domain 2 protein |
| PFC | prefrontal cortex |
| PHpg | para-hippocampal gyrus |
| PLP1 | proteolipid protein 1 |
| PPn | pedunculopontine nucleus |
| preMctx | premotor cortex |
| PSD95 | postsynaptic density of protein 95 |
| Put | putamen |
| SNpc | substantia nigra pars compacta |
| SNpr | substantia nigra pars reticulate |
| Spt | septum |
| Sub | subiculum |
| TegB | bulbar tegmentum |
| Th | thalamus |
| TrkB | tyrosine receptor kinase B |
| VGLUT | vesicular glutamate transporter |
| vHip | ventral hippocampus |
| VP | ventral pallidum |
| VTA | ventral tegmental area |
References
- Koob, G.F.; Volkow, N.D. Neurocircuitry of addiction. Neuropsychopharmacology 2010, 35, 217–238. [Google Scholar] [CrossRef] [Green Version]
- Salgado, S.; Kaplitt, M.G. The nucleus accumbens: A comprehensive review. Stereotact. Funct. Neurosurg. 2015, 93, 75–93. [Google Scholar] [CrossRef] [PubMed]
- Groenewegen, H.J.; Wrigth, C.I.; Beijer, A.V.J. The nucleus accumbens: Gateway for limbic structures to reach the motor system? Prog. Brain Res. 1996, 107, 485–511. [Google Scholar]
- Heimer, L.; Zahm, D.S.; Churchill, L.; Kalivas, P.W.; Wohltmann, C. Specificity in the projection patterns of accumbal core and shell in the rat. Neuroscience 1991, 41, 89–125. [Google Scholar] [CrossRef]
- Jongen-Rêlo, A.L.; Voorn, P.; Groenewegen, H.J. Immunohistochemical Characterization of the Shell and Core Territories of the Nucleus Accumbens in the Rat. Eur. J. Neurosci. 1994, 6, 1255–1264. [Google Scholar] [CrossRef]
- Zahm, D.S.; Heimer, L. Specificity in the efferent projections of the nucleus accumbens in the rat: Comparison of the rostral pole projection patterns with those of the core and shell. J. Comp. Neurol. 1993, 327, 220–232. [Google Scholar] [CrossRef]
- Sazdanović, M.; Sazdanović, P.; Živanović-Mačužić, I.; Jakovljević, V.; Jeremić, D.; Peljto, A.; Toševski, J. Neuroni humanog nukleusa akumbensa. Vojnosanit. Pregl. 2011, 68, 655–660. [Google Scholar] [CrossRef] [PubMed]
- Meredith, G.E.; Agolia, R.; Arts, M.P.M.; Groenewegen, H.J.; Zahm, D.S. Morphological differences between projection neurons of the core and shell in the nucleus accumbens of the rat. Neuroscience 1992, 50, 149–162. [Google Scholar] [CrossRef]
- Gangarossa, G.; Espallergues, J.; D’Exaerde, A.D.K.; El Mestikawy, S.; Gerfen, C.R.; Hervé, D.; Girault, J.-A.; Valjent, E. Distribution and compartmental organization of GABAergic medium-sized spiny neurons in the mouse Nucleus Accumbens. Front. Neural Circuits 2013, 7, 22. [Google Scholar] [CrossRef] [Green Version]
- Jones, D.L.; Mogenson, G.J. Nucleus accumbens to globus pallidus GABA projection: Electrophysiological and iontophoretic investigations. Brain Res. 1980, 188, 93–105. [Google Scholar] [CrossRef]
- Meredith, G.E.; Blank, B.; Groenewegen, H.J. The distribution and compartmental organization of the cholinergic neurons in nucleus accumbens of the rat. Neuroscience 1989, 31, 327–345. [Google Scholar] [CrossRef]
- Deutch, A.Y.; Cameron, D.S. Pharmacological characterization of dopamine systems in the nucleus accumbens core and shell. Neuroscience 1992, 46, 49–56. [Google Scholar] [CrossRef]
- Patel, S.; Roberts, J.; Moorman, J.; Reavill, C. Localization of serotonin-4 receptors in the striatonigral pathway in rat brain. Neuroscience 1995, 69, 1159–1167. [Google Scholar] [CrossRef]
- McKittrick, C.R.; Abercrombie, E.D. Catecholamine mapping within nucleus accumbens: Differences in basal and amphetamine-stimulated efflux of norepinephrine and dopamine in shell and core. J. Neurochem. 2007, 100, 1247–1256. [Google Scholar] [CrossRef]
- Nirenberg, M.J.; Chan, J.; Pohorille, A.; Vaughan, R.A.; Uhl, G.R.; Kuhar, M.J.; Pickel, V.M. The dopamine transporter: Comparative ultrastructure of dopaminergic axons in limbic and motor compartments of the nucleus accumbens. J. Neurosci. 1997, 17, 6899–6907. [Google Scholar] [CrossRef] [Green Version]
- Pifl, C.; Agneter, E.; Drobny, H.; Sitte, H.H.; Singer, E.A. Amphetamine reverses or blocks the operation of the human noradrenaline transporter depending on its concentration: Superfusion studies on transfected cells. Neuropharmacology 1999, 38, 157–165. [Google Scholar] [CrossRef]
- Seiden, L.S.; Sabol, K.E.; Ricaurte, G.A. Amphetamine: Effects on catecholamine systems and behavior. Annu. Rev. Pharmacol. Toxicol. 1993, 33, 639–677. [Google Scholar] [CrossRef] [PubMed]
- Tong, J.; Hornykiewicz, O.; Kish, S.J. Identification of a noradrenaline-rich subdivision of the human nucleus accumbens. J. Neurochem. 2006, 96, 349–354. [Google Scholar] [CrossRef]
- Versteeg, D.H.G.; Van der Gugten, J.; De Jong, W.; Palkovits, M. Regional concentrations of noradrenaline and dopamine in rat brain. Brain Res. 1976, 113, 563–574. [Google Scholar] [CrossRef]
- DeFrance, J.F.; Sikes, R.W.; Gottesfeld, Z. Regional Distribution of Catecholamines in Nucleus Accumbens of the Rabbit. J. Neurochem. 1983, 40, 291–293. [Google Scholar] [CrossRef] [PubMed]
- Pifl, C.; Schingnitz, G.; Hornykiewicz, O. Effect of 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine on the regional distribution of brain monoamines in the rhesus monkey. Neuroscience 1991, 44, 591–605. [Google Scholar] [CrossRef]
- Tye, K.M. Glutamate Inputs to the Nucleus Accumbens: Does Source Matter? Neuron 2012, 76, 671–673. [Google Scholar] [CrossRef] [Green Version]
- Prensa, L.; Richard, S.; Parent, A. Chemical anatomy of the human ventral striatum and adjacent basal forebrain structures. J. Comp. Neurol. 2003, 460, 345–367. [Google Scholar] [CrossRef] [PubMed]
- Voorn, P.; Gerfen, C.R.; Groenewegen, H.J. Compartmental organization of the ventral striatum of the rat: Immunohistochemical distribution of enkephalin, substance P, dopamine, and calcium-binding protein. J. Comp. Neurol. 1989, 289, 189–201. [Google Scholar] [CrossRef] [PubMed]
- Berendse, H.W.; Groenewegen, H.J. Organization of the thalamostriatal projections in the rat, with special emphasis on the ventral striatum. J. Comp. Neurol. 1990, 299, 187–228. [Google Scholar] [CrossRef]
- Churchill, L.; Cross, R.S.; Pazdernik, T.L.; Nelson, S.R.; Zahm, D.S.; Heimer, L.; Kalivas, P.W. Patterns of glucose use after bicuculline-induced convulsions in relationship to γ-aminobutyric acid and μ-opioid receptors in the ventral pallidum-functional markers for the ventral pallidum. Brain Res. 1992, 581, 39–45. [Google Scholar] [CrossRef]
- Mega, M.S.; Cummings, J.L.; Salloway, S.; Malloy, P. The limbic system: An anatomic, phylogenetic, and clinical perspective. J. Neuropsychiatry Clin. Neurosci. 1997, 9, 315–330. [Google Scholar]
- MacLean, P.D. The triune brain, emotion, and scientific bias. In The Neuroscience Second Study Program; Schmitt, F.O., Ed.; New York Rockefeller University Press, Birkhäuser: Boston, MA, USA, 1970; pp. 336–349. [Google Scholar]
- Yakovlev, P.I.; Lecours, A.R. The myelogenetic cycles of regional maturation of the brain. In Regional Development of the Brain in Early Life; Minkowski, A., Ed.; Blackwell Science: Oxford, UK, 1967; pp. 3–70. [Google Scholar]
- Brog, J.S.; Salyapongse, A.; Deutch, A.Y.; Zahm, D.S. The patterns of afferent innervation of the core and shell in the “Accumbens” part of the rat ventral striatum: Immunohistochemical detection of retrogradely transported fluoro-gold. J. Comp. Neurol. 1993, 338, 255–278. [Google Scholar] [CrossRef]
- Ito, N.; Ishida, H.; Fumio, M.; Naito, H. Microelectrode study of projections from the amygdaloid complex to the nucleus accumbens in the cat. Brain Res. 1974, 67, 338–341. [Google Scholar] [CrossRef]
- Groenewegen, H.J.; Russchen, F.T. Organization of the efferent projections of the nucleus accumbens to pallidal, hypothalamic, and mesencephalic structures: A tracing and immunohistochemical study in the cat. J. Comp. Neurol. 1984, 223, 347–367. [Google Scholar] [CrossRef]
- Russchen, F.T.; Bakst, I.; Amaral, D.G.; Price, J.L. The amygdalostriatal projections in the monkey. An anterograde tracing study. Brain Res. 1985, 329, 241–257. [Google Scholar] [CrossRef]
- Williams, D.J.; Crossman, A.R.; Slater, P. The efferent projections of the nucleus accumbens in the rat. Brain Res. 1977, 130, 217–227. [Google Scholar] [CrossRef]
- Nauta, W.J.H.; Smith, G.P.; Faull, R.L.M.; Domesick, V.B. Efferent connections and nigral afferents of the nucleus accumbens septi in the rat. Neuroscience 1978, 3, 385–401. [Google Scholar] [CrossRef]
- Wright, C.I.; Groenewegen, H.J. Patterns of overlap and segregation between insular cortical, intermediodorsal thalamic and basal amygdaloid afferents in the nucleus accumbens of the rat. Neuroscience 1996, 73, 359–373. [Google Scholar] [CrossRef]
- Zahm, D.S.; Williams, E.; Wohltmann, C. Ventral striatopallidothalamic projection: IV. Relative involvements of neurochemically distinct subterritories in the ventral pallidum and adjacent parts of the rostroventral forebrain. J. Comp. Neurol. 1996, 364, 340–362. [Google Scholar] [CrossRef]
- O’Donnell, P.; Lavín, A.; Enquist, L.W.; Grace, A.A.; Card, J.P. Interconnected parallel circuits between rat nucleus accumbens and thalamus revealed by retrograde transynaptic transport of pseudorabies virus. J. Neurosci. 1997, 17, 2143–2167. [Google Scholar] [CrossRef] [Green Version]
- Alexander, G.E.; DeLong, M.R.; Strick, P.L. Parallel Organization of Functionally Segregated Circuits Linking Basal Ganglia and Cortex. Annu. Rev. Neurosci. 1986, 9, 357–381. [Google Scholar] [CrossRef] [PubMed]
- Alexander, G.E.; Crutcher, M.D.; Delong, M.R. Chapter 6 Basal ganglia-thalamocortical circuits: Parallel substrates for motor, oculomotor, “prefrontal” and “limbic” functions. Prog. Brain Res. 1991, 85, 119–146. [Google Scholar] [CrossRef]
- Scofield, M.D.; Heinsbroek, J.A.; Gipson, C.D.; Kupchik, Y.M.; Spencer, S.; Smith, A.C.W.; Roberts-Wolfe, D.; Kalivas, P.W. The nucleus accumbens: Mechanisms of addiction across drug classes reflect the importance of glutamate homeostasis. Pharmacol. Rev. 2016, 68, 816–871. [Google Scholar] [CrossRef] [Green Version]
- Cooper, S.; Robison, A.J.; Mazei-Robison, M.S. Reward Circuitry in Addiction. Neurotherapeutics 2017, 14, 687–697. [Google Scholar] [CrossRef] [Green Version]
- Russo, S.J.; Dietz, D.M.; Dumitriu, D.; Morrison, J.H.; Malenka, R.C.; Nestler, E.J. The addicted synapse: Mechanisms of synaptic and structural plasticity in nucleus accumbens. Trends Neurosci. 2010, 33, 267–276. [Google Scholar] [CrossRef] [Green Version]
- Dietz, D.M.; Dietz, K.C.; Nestler, E.J.; Russo, S.J. Molecular mechanisms of psychostimulant-induced structural plasticity. Pharmacopsychiatry 2009, 42, S69–S78. [Google Scholar] [CrossRef] [Green Version]
- Fallon, J.H.; Moore, R.Y. Catecholamine innervation of the basal forebrain IV. Topography of the dopamine projection to the basal forebrain and neostriatum. J. Comp. Neurol. 1978, 180, 545–579. [Google Scholar] [CrossRef]
- Phillipson, O.T.; Griffiths, A.C. The topographic order of inputs to nucleus accumbens in the rat. Neuroscience 1985, 16, 275–296. [Google Scholar] [CrossRef]
- Han, X.; Jing, M.-Y.; Zhao, T.-Y.; Wu, N.; Song, R.; Li, J. Role of dopamine projections from ventral tegmental area to nucleus accumbens and medial prefrontal cortex in reinforcement behaviors assessed using optogenetic manipulation. Metab. Brain Dis. 2017, 32, 1491–1502. [Google Scholar] [CrossRef]
- Gerfen, C.R.; Surmeier, D.J. Modulation of Striatal Projection Systems by Dopamine. Annu. Rev. Neurosci. 2011, 34, 441–466. [Google Scholar] [CrossRef] [Green Version]
- Nirenberg, M.J.; Vaughan, R.A.; Uhl, G.R.; Kuhar, M.J.; Pickel, V.M. The dopamine transporter is localized to dendritic and axonal plasma membranes of nigrostriatal dopaminergic neurons. J. Neurosci. 1996, 16, 436–447. [Google Scholar] [CrossRef]
- Zahm, D.S.; Brog, J.S. On the significance of subterritories in the “accumbens” part of the rat ventral striatum. Neuroscience 1992, 50, 751–767. [Google Scholar] [CrossRef]
- Bardo, M.T.; Hammer, R.P. Autoradiographic localization of dopamine D1 and D2 receptors in rat nucleus accumbens: Resistance to differential rearing conditions. Neuroscience 1991, 45, 281–290. [Google Scholar] [CrossRef]
- Hart, A.S.; Rutledge, R.; Glimcher, P.W.; Phillips, P. Phasic dopamine release in the rat nucleus accumbens symmetrically encodes a reward prediction error term. J. Neurosci. 2014, 34, 698–704. [Google Scholar] [CrossRef] [Green Version]
- Mohebi, A.; Pettibone, J.R.; Hamid, A.A.; Wong, J.M.T.; Vinson, L.T.; Patriarchi, T.; Tian, L.; Kennedy, R.T.; Berke, J.D. Dissociable dopamine dynamics for learning and motivation. Nature 2019, 570, 65–70. [Google Scholar] [CrossRef]
- Hamid, A.A.; Pettibone, J.R.; Mabrouk, O.S.; Hetrick, V.L.; Schmidt, R.; Vander Weele, C.M.; Kennedy, R.T.; Aragona, B.J.; Berke, J.D. Mesolimbic dopamine signals the value of work. Nat. Neurosci. 2015, 19, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Berke, J.D. What does dopamine mean? Nat. Neurosci. 2018, 21, 787–793. [Google Scholar] [CrossRef]
- Asl, M.M.; Vahabie, A.H.; Valizadeh, A. Review paper: Dopaminergic modulation of synaptic plasticity, its role in neuropsychiatric disorders, and its computational modeling. Basic Clin. Neurosci. 2019, 10, 1–12. [Google Scholar]
- Smith, R.J.; Lobo, M.K.; Spencer, S.; Kalivas, P.W. Cocaine-induced adaptations in D1 and D2 accumbens projection neurons (a dichotomy not necessarily synonymous with direct and indirect pathways). Curr. Opin. Neurobiol. 2013, 23, 546–552. [Google Scholar] [CrossRef] [Green Version]
- Creed, M.; Ntamati, N.R.; Chandra, R.; Lobo, M.K.; Lüscher, C. Convergence of Reinforcing and Anhedonic Cocaine Effects in the Ventral Pallidum. Neuron 2016, 92, 214–226. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Milovanovic, M.; Zhao, Y.; Wolf, M.E. Acute and chronic dopamine receptor stimulation modulates AMPA receptor trafficking in nucleus accumbens neurons cocultured with prefrontal cortex neurons. J. Neurosci. 2008, 28, 4216–4230. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Wolf, M.E. Nucleus accumbens neurons exhibit synaptic scaling that is occluded by repeated dopamine pre-exposure. Eur. J. Neurosci. 2009, 30, 539–550. [Google Scholar] [CrossRef]
- Li, M.-Y.; Yan, Q.-S.; Coffey, L.L.; Reith, M.E.A. Extracellular Dopamine, Norepinephrine, and Serotonin in the Nucleus Accumbens of Freely Moving Rats during Intracerebral Dialysis with Cocaine and Other Monoamine Uptake Blockers. J. Neurochem. 1996, 66, 559–568. [Google Scholar] [CrossRef] [PubMed]
- Sesack, S.R.; Carr, D.B.; Omelchenko, N.; Pinto, A. Anatomical Substrates for Glutamate-Dopamine Interactions: Evidence for Specificity of Connections and Extrasynaptic Actions. Ann. N. Y. Acad. Sci. 2003, 1003, 36–52. [Google Scholar] [CrossRef]
- Chao, S.Z.; Ariano, M.A.; Peterson, D.A.; Wolf, M.E. D1 dopamine receptor stimulation increases GluR1 surface expression in nucleus accumbens neurons. J. Neurochem. 2002, 83, 704–712. [Google Scholar] [CrossRef] [Green Version]
- Mangiavacchi, S.; Wolf, M.E. D1 dopamine receptor stimulation increases the rate of AMPA receptor insertion onto the surface of cultured nucleus accumbens neurons through a pathway dependent on protein kinase A. J. Neurochem. 2004, 88, 1261–1271. [Google Scholar] [CrossRef]
- Sanna, A.; Fattore, L.; Badas, P.; Corona, G.; Diana, M. The hypodopaminergic state ten years after: Transcranial magnetic stimulation as a tool to test the dopamine hypothesis of drug addiction. Curr. Opin. Pharmacol. 2021, 56, 61–67. [Google Scholar] [CrossRef]
- Popescu, A.; Marian, M.; Drăgoi, A.; Costea, R.-V. Understanding the genetics and neurobiological pathways behind addiction (Review). Exp. Ther. Med. 2021, 21, 544. [Google Scholar] [CrossRef]
- Bimpisidis, Z.; Wallén-Mackenzie, Å. Neurocircuitry of Reward and Addiction: Potential Impact of Dopamine–Glutamate Co-release as Future Target in Substance Use Disorder. J. Clin. Med. 2019, 8, 1887. [Google Scholar] [CrossRef] [Green Version]
- Samaha, A.N.; Khoo, S.Y.-S.; Ferrario, C.R.; Robinson, T.E. Dopamine ‘ups and downs’ in addiction revisited. Trends Neurosci. 2021, 44, 516–526. [Google Scholar] [CrossRef]
- Nestler, E.J.; Lüscher, C. The Molecular Basis of Drug Addiction: Linking Epigenetic to Synaptic and Circuit Mechanisms. Neuron 2019, 102, 48–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solinas, M.; Belujon, P.; Fernagut, P.O.; Jaber, M.; Thiriet, N. Dopamine and addiction: What have we learned from 40 years of research. J. Neural Transm. 2019, 126, 481–516. [Google Scholar] [CrossRef]
- Liu, J.; Gandhi, P.J.; Pavuluri, R.; Shelkar, G.P.; Dravid, S.M. Glutamate delta-1 receptor regulates cocaine-induced plasticity in the nucleus accumbens. Transl. Psychiatry 2018, 8, 219. [Google Scholar] [CrossRef] [PubMed]
- Schotanus, S.M.; Chergui, K. Dopamine D1 receptors and group I metabotropic glutamate receptors contribute to the induction of long-term potentiation in the nucleus accumbens. Neuropharmacology 2008, 54, 837–844. [Google Scholar] [CrossRef]
- Tukey, D.S.; Lee, M.; Xu, D.; Eberle, S.E.; Goffer, Y.; Manders, T.R.; Ziff, E.B.; Wang, J. Differential effects of natural rewards and pain on vesicular glutamate transporter expression in the nucleus accumbens. Mol. Brain 2013, 6, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imperato, A.; Honoré, T.; Jensen, L.H. Dopamine release in the nucleus caudatus and in the nucleus accumbens is under glutamatergic control through non-NMDA receptors: A study in freely-moving rats. Brain Res. 1990, 530, 223–228. [Google Scholar] [CrossRef]
- Mogenson, G.J.; Nielsen, M. A study of the contribution of hippocampal-accumbens-subpallidal projections to locomotor activity. Behav. Neural Biol. 1984, 42, 38–51. [Google Scholar] [CrossRef]
- Cornish, J.L.; Kalivas, P.W. Glutamate transmission in the nucleus accumbens mediates relapse in cocaine addiction. J. Neurosci. 2000, 20, RC89. [Google Scholar] [CrossRef] [Green Version]
- Di Ciano, P.; Everitt, B.J. Dissociable effects of antagonism of NMDA and AMPA/KA receptors in the nucleus accumbens core and shell on cocaine-seeking behavior. Neuropsychopharmacology 2001, 25, 341–360. [Google Scholar] [CrossRef] [Green Version]
- Kelley, A.E.; Smith-Roe, S.L.; Holahan, M.R. Response-reinforcement learning is dependent on N-methyl-D-aspartate receptor activation in the nucleus accumbens core. Proc. Natl. Acad. Sci. USA 1997, 94, 12174–12179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolf, M.E.; Ferrario, C.R. AMPA receptor plasticity in the nucleus accumbens after repeated exposure to cocaine. Neurosci. Biobehav. Rev. 2010, 35, 185–211. [Google Scholar] [CrossRef] [Green Version]
- Schotanus, S.M.; Chergui, K. Long-term potentiation in the nucleus accumbens requires both NR2A- and NR2B-containing N-methyl-D-aspartate receptors. Eur. J. Neurosci. 2008, 27, 1957–1964. [Google Scholar] [CrossRef]
- Hu, H.; Real, E.; Takamiya, K.; Kang, M.G.; Ledoux, J.; Huganir, R.L.; Malinow, R. Emotion Enhances Learning via Norepinephrine Regulation of AMPA-Receptor Trafficking. Cell 2007, 131, 160–173. [Google Scholar] [CrossRef] [Green Version]
- Mishra, D.; Zhang, X.; Chergui, K. Ethanol Disrupts the Mechanisms of Induction of Long-Term Potentiation in the Mouse Nucleus Accumbens. Alcohol. Clin. Exp. Res. 2012, 36, 2117–2125. [Google Scholar] [CrossRef] [Green Version]
- Pennartz, C.M.A.; Boeijinga, P.H.; da Silva, F.H.L. Locally evoked potentials in slices of the rat nucleus accumbens: NMDA and non-NMDA receptor mediated components and modulation by GABA. Brain Res. 1990, 529, 30–41. [Google Scholar] [CrossRef] [Green Version]
- Bredt, D.S.; Nicoll, R.A. AMPA receptor trafficking at excitatory synapses. Neuron 2003, 40, 361–379. [Google Scholar] [CrossRef] [Green Version]
- Shepherd, J.D.; Huganir, R.L. The Cell Biology of Synaptic Plasticity: AMPA Receptor Trafficking. Annu. Rev. Cell Dev. Biol. 2007, 23, 613–643. [Google Scholar] [CrossRef] [Green Version]
- Derkach, V.A.; Oh, M.C.; Guire, E.S.; Soderling, T.R. Regulatory mechanisms of AMPA receptors in synaptic plasticity. Nat. Rev. Neurosci. 2007, 8, 101–113. [Google Scholar] [CrossRef] [PubMed]
- Song, I.; Huganir, R.L. Regulation of AMPA receptors during synaptic plasticity. Trends Neurosci. 2002, 25, 578–588. [Google Scholar] [CrossRef]
- Malenka, R.C.; Nicoll, R.A. Silent synapses speak up. Neuron 1997, 19, 473–476. [Google Scholar] [CrossRef] [Green Version]
- Grueter, B.A.; Rothwell, P.E.; Malenka, R.C. Integrating synaptic plasticity and striatal circuit function in addiction. Curr. Opin. Neurobiol. 2012, 22, 545–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.H.; Lin, Y.; Mu, P.; Lee, B.R.; Brown, T.E.; Wayman, G.; Marie, H.; Liu, W.; Yan, Z.; Sorg, B.A.; et al. In vivo cocaine experience generates silent synapses. Neuron 2009, 63, 40–47. [Google Scholar] [CrossRef] [Green Version]
- Knackstedt, L.A.; Kalivas, P.W. Glutamate and reinstatement. Curr. Opin. Pharmacol. 2009, 9, 59–64. [Google Scholar] [CrossRef] [Green Version]
- Boudreau, A.C.; Wolf, M.E. Behavioral sensitization to cocaine is associated with increased AMPA receptor surface expression in the nucleus accumbens. J. Neurosci. 2005, 25, 9144–9151. [Google Scholar] [CrossRef]
- Boudreau, A.C.; Reimers, J.M.; Milovanovic, M.; Wolf, M.E. Cell surface AMPA receptors in the rat nucleus accumbens increase during cocaine withdrawal but internalize after cocaine challenge in association with altered activation of mitogen-activated protein kinases. J. Neurosci. 2007, 27, 10621–10635. [Google Scholar] [CrossRef] [Green Version]
- Kourrich, S.; Rothwell, P.E.; Klug, J.R.; Thomas, M.J. Cocaine experience controls bidirectional synaptic plasticity in the nucleus accumbens. J. Neurosci. 2007, 27, 7921–7928. [Google Scholar] [CrossRef] [Green Version]
- Marie, N.; Canestrelli, C.; Noble, F. Transfer of neuroplasticity from nucleus accumbens core to shell is required for cocaine reward. PLoS ONE 2012, 7, e30241. [Google Scholar] [CrossRef] [PubMed]
- Beckley, J.T.; Laguesse, S.; Phamluong, K.; Morisot, N.; Wegner, S.A.; Ron, D. The first alcohol drink triggers mTORC1-dependent synaptic plasticity in nucleus accumbens dopamine D1 receptor neurons. J. Neurosci. 2016, 36, 701–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abrahao, K.P.; Ariwodola, O.J.; Butler, T.R.; Rau, A.R.; Skelly, M.J.; Carter, E.; Alexander, N.P.; McCool, B.A.; Souza-Formigoni, M.L.O.; Weiner, J.L. Locomotor sensitization to ethanol impairs NMDA receptor-dependent synaptic plasticity in the nucleus accumbens and increases ethanol self-administration. J. Neurosci. 2013, 33, 4834–4842. [Google Scholar] [CrossRef] [Green Version]
- Záborsky, L.; Cullinan, W.E. Projections from the nucleus accumbens to cholinergic neurons of the ventral pallidum: A correlated light and electron microscopic double-immunolabeling study in rat. Brain Res. 1992, 570, 92–101. [Google Scholar] [CrossRef]
- Avena, N.M.; Rada, P.V. Cholinergic modulation of food and drug satiety and withdrawal. Physiol. Behav. 2012, 106, 332–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Witten, I.B.; Lin, S.C.; Brodsky, M.; Prakash, R.; Diester, I.; Anikeeva, P.; Gradinaru, V.; Ramakrishnan, C.; Deisseroth, K. Cholinergic interneurons control local circuit activity and cocaine conditioning. Science 2010, 330, 1677–1681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wachtel, H.; Anden, N.E. Motor activity of rats following intracerebral injections of drugs influencing GABA mechanisms. Naunyn. Schmiedebergs. Arch. Pharmacol. 1978, 302, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.L.; Mogenson, G.J.; Wu, M. Injections of dopaminergic, cholinergic, serotoninergic and gabaergic drugs into the nucleus accumbens: Effects on locomotor activity in the rat. Neuropharmacology 1981, 20, 29–37. [Google Scholar] [CrossRef]
- Brodnik, Z.D.; Batra, A.; Oleson, E.B.; Espana, R.A. Local GABAA Receptor-Mediated Suppression of Dopamine Release within the Nucleus Accumbens. ACS Chem. Neurosci. 2019, 10, 1978–1985. [Google Scholar] [CrossRef]
- Pitman, K.A.; Puil, E.; Borgland, S.L. GABAB modulation of dopamine release in the nucleus accumbens core. Eur. J. Neurosci. 2014, 40, 3472–3480. [Google Scholar] [CrossRef]
- Watanabe, Y.; Aono, Y.; Komiya, M.; Waddington, J.L.; Saigusa, T. Stimulation of accumbal GABA B receptors inhibits delta1- and delta2-opioid receptor-mediated dopamine efflux in the nucleus accumbens of freely moving rats. Eur. J. Pharmacol. 2018, 837, 88–95. [Google Scholar] [CrossRef]
- Saigusa, T.; Aono, Y.; Sekino, R.; Uchida, T.; Takada, K.; Oi, Y.; Koshikawa, N.; Cools, A.R. In vivo neurochemical evidence that newly synthesised GABA activates GABA B, but not GABA A, receptors on dopaminergic nerve endings in the nucleus accumbens of freely moving rats. Neuropharmacology 2012, 62, 907–913. [Google Scholar] [CrossRef] [PubMed]
- Rada, P.V.; Mark, G.P.; Hoebel, B.G. In vivo modulation of acetylcholine in the nucleus accumbens of freely moving rats: II. Inhibition by γ-aminobutyric acid. Brain Res. 1993, 619, 105–110. [Google Scholar] [CrossRef]
- Aono, Y.; Watanabe, Y.; Ishikawa, M.; Kuboyama, N.; Waddington, J.L.; Saigusa, T. In vivo neurochemical evidence that stimulation of accumbal GABA A and GABA B receptors each reduce acetylcholine efflux without affecting dopamine efflux in the nucleus accumbens of freely moving rats. Synapse 2019, 73, e22081. [Google Scholar] [CrossRef]
- Rahman, S.; McBride, W.J. Involvement of GABA and cholinergic receptors in the nucleus accumbens on feedback control of somatodendritic dopamine release in the ventral tegmental area. J. Neurochem. 2002, 80, 646–654. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Heida, T.; Stegenga, J.; Zhao, Y.; Moser, A.; Tronnier, V.; Feuerstein, T.J.; Hofmann, U.G. High-frequency electrical stimulation suppresses cholinergic accumbens interneurons in acute rat brain slices through GABAB receptors. Eur. J. Neurosci. 2014, 40, 3653–3662. [Google Scholar] [CrossRef] [PubMed]
- Manz, K.M.; Baxley, A.G.; Zurawski, Z.; Hamm, H.E.; Grueter, B.A. Heterosynaptic GABAB Receptor Function within Feedforward Microcircuits Gates Glutamatergic Transmission in the Nucleus Accumbens Core. J. Neurosci. 2019, 39, 9277–9293. [Google Scholar] [CrossRef]
- Ding, Z.M.; Ingraham, C.M.; Rodd, Z.A.; McBride, W.J. The reinforcing effects of ethanol within the nucleus accumbens shell involve activation of local GABA and serotonin receptors. J. Psychopharmacol. 2015, 29, 725–733. [Google Scholar] [CrossRef] [Green Version]
- Mishra, D.; Chergui, K. Ethanol inhibits excitatory neurotransmission in the nucleus accumbens of adolescent mice through GABAA and GABAB receptors. Addict. Biol. 2013, 18, 605–613. [Google Scholar] [CrossRef]
- Yoon, S.S.; Kim, J.A.; Lee, B.H.; Choi, K.H.; Shim, I.; Choi, S.H.; Hwang, M.; Yang, C.H. Role for GABA agonists in the nucleus accumbens in regulating morphine self-administration. Neurosci. Lett. 2009, 462, 289–293. [Google Scholar] [CrossRef]
- Varani, A.P.; Pedrón, V.T.; Aon, A.J.; Höcht, C.; Acosta, G.B.; Bettler, B.; Balerio, G.N. Nicotine-induced molecular alterations are modulated by GABA B receptor activity. Addict. Biol. 2018, 23, 230–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahraei, H.; Askaripour, M.; Esmaeilpour, K.; Shahsavari, F.; Rajabi, S.; Moradi-Kor, N. GABAB receptor activation ameliorates spatial memory impairments in stress-exposed rats. Neuropsychiatr. Dis. Treat. 2019, 15, 1497–1506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopes, A.P.F.; da Cunha, I.C.; Steffens, S.M.; Ferraz, A.; Vargas, J.C.; de Lima, T.C.M.; Neto, J.M.; Faria, M.S.; Paschoalini, M.A. GABAA and GABAB agonist microinjections into medial accumbens shell increase feeding and induce anxiolysis in an animal model of anxiety. Behav. Brain Res. 2007, 184, 142–149. [Google Scholar] [CrossRef]
- Wong, L.S.; Eshel, G.; Dreher, J.; Ong, J.; Jackson, D.M. Role of dopamine and GABA in the control of motor activity elicited from the rat nucleus accumbens. Pharmacol. Biochem. Behav. 1991, 38, 829–835. [Google Scholar] [CrossRef]
- Stratford, T.R.; Kelley, A.E. GABA in the nucleus accumbens shell participates in the central regulation of feeding behavior. J. Neurosci. 1997, 17, 4434–4440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zangen, A.; Nakash, R.; Overstreet, D.H.; Yadid, G. Association between depressive behavior and absence of serotonin-dopamine interaction in the nucleus accumbens. Psychopharmacology 2001, 155, 434–439. [Google Scholar] [CrossRef]
- Browne, C.J.; Abela, A.R.; Chu, D.; Li, Z.; Ji, X.; Lambe, E.K.; Fletcher, P.J. Dorsal raphe serotonin neurons inhibit operant responding for reward via inputs to the ventral tegmental area but not the nucleus accumbens: Evidence from studies combining optogenetic stimulation and serotonin reuptake inhibition. Neuropsychopharmacology 2019, 44, 793–804. [Google Scholar] [CrossRef]
- Walsh, J.J.; Christoffel, D.J.; Heifets, B.D.; Ben-Dor, G.A.; Selimbeyoglu, A.; Hung, L.W.; Deisseroth, K.; Malenka, R.C. 5-HT release in nucleus accumbens rescues social deficits in mouse autism model. Nature 2018, 560, 589–594. [Google Scholar] [CrossRef] [PubMed]
- Dölen, G.; Darvishzadeh, A.; Huang, K.W.; Malenka, R.C. Social reward requires coordinated activity of nucleus accumbens oxytocin and serotonin. Nature 2013, 501, 179–184. [Google Scholar] [CrossRef] [Green Version]
- López-Arnau, R.; Buenrostro-Jáuregui, M.; Camarasa, J.; Pubill, D.; Escubedo, E. Effect of the combination of mephedrone plus ethanol on serotonin and dopamine release in the nucleus accumbens and medial prefrontal cortex of awake rats. Naunyn. Schmiedebergs. Arch. Pharmacol. 2018, 391, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Teneud, L.M.; Baptista, T.; Murzi, E.; Hoebel, B.G.; Hernandez, L. Systemic and local cocaine increase extracellular serotonin in the nucleus accumbens. Pharmacol. Biochem. Behav. 1996, 53, 747–752. [Google Scholar] [CrossRef]
- Canal, C.E.; Murnane, K.S. The serotonin 5-HT2C receptor and the non-addictive nature of classic hallucinogens. J. Psychopharmacol. 2017, 31, 127–143. [Google Scholar] [CrossRef] [Green Version]
- Robison, A.J.; Nestler, E.J. Transcriptional and epigenetic mechanisms of addiction. Nat. Rev. Neurosci. 2011, 12, 623–637. [Google Scholar] [CrossRef] [Green Version]
- Bubar, M.J.; Cunningham, K.A. Prospects for serotonin 5-HT2R pharmacotherapy in psychostimulant abuse. Prog. Brain Res. 2008, 172, 319–346. [Google Scholar] [PubMed]
- McMahon, L.R.; Cunningham, K.A. Antagonism of 5-hydroxytryptamine2A receptors attenuates the behavioral effects of cocaine in rats. J. Pharmacol. Exp. Ther. 2001, 297, 357–363. [Google Scholar] [PubMed]
- McMahon, L.R.; Filip, M.; Cunningham, K.A. Differential regulation of the mesoaccumbens circuit by serotonin 5-hydroxytryptamine (5-HT)2A and 5-HT2C receptors. J. Neurosci. 2001, 21, 7781–7787. [Google Scholar] [CrossRef] [Green Version]
- Sawyer, E.K.; Mun, J.; Nye, J.A.; Kimmel, H.L.; Voll, R.J.; Stehouwer, J.S.; Rice, K.C.; Goodman, M.M.; Howell, L.L. Neurobiological changes mediating the effects of chronic fluoxetine on cocaine use. Neuropsychopharmacology 2012, 37, 1816–1824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Auclair, A.; Drouin, C.; Cotecchia, S.; Glowinski, J.; Tassin, J.P. 5-HT2A and α1b-adrenergic receptors entirely mediate dopamine release, locomotor response and behavioural sensitization to opiates and psychostimulants. Eur. J. Neurosci. 2004, 20, 3073–3084. [Google Scholar] [CrossRef]
- Broderick, P.A.; Olabisi, O.A.; Rahni, D.N.; Zhou, Y. Cocaine acts on accumbens monoamines and locomotor behavior via a 5-HT 2A/2C receptor mechanism as shown by ketanserin: 24-h follow-up studies. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2004, 28, 547–557. [Google Scholar] [CrossRef] [PubMed]
- Murnane, K.S.; Andersen, M.L.; Rice, K.C.; Howell, L.L. Selective serotonin 2A receptor antagonism attenuates the effects of amphetamine on arousal and dopamine overflow in non-human primates. J. Sleep Res. 2013, 22, 581–588. [Google Scholar] [CrossRef] [PubMed]
- Higgins, G.A.; Fletcher, P.J. Therapeutic Potential of 5-HT2C Receptor Agonists for Addictive Disorders. ACS Chem. Neurosci. 2015, 6, 1071–1088. [Google Scholar] [CrossRef]
- Howell, L.L.; Cunningham, K.A. Serotonin 5-HT2 receptor interactions with dopamine function: Implications for therapeutics in cocaine use disorder. Pharmacol. Rev. 2015, 67, 176–197. [Google Scholar] [CrossRef] [Green Version]
- Müller, C.P.; Homberg, J.R. The role of serotonin in drug use and addiction. Behav. Brain Res. 2015, 277, 146–192. [Google Scholar] [CrossRef] [PubMed]
- Bubar, M.J.; Stutz, S.J.; Cunningham, K.A. 5-HT2c Receptors localize to dopamine and gaba neurons in the rat mesoaccumbens pathway. PLoS ONE 2011, 6, e20508. [Google Scholar] [CrossRef] [Green Version]
- Bubar, M.J.; Cunningham, K.A. Distribution of serotonin 5-HT2C receptors in the ventral tegmental area. Neuroscience 2007, 146, 286–297. [Google Scholar] [CrossRef] [Green Version]
- Cathala, A.; Devroye, C.; Maitre, M.; Piazza, P.V.; Abrous, D.N.; Revest, J.M.; Spampinato, U. Serotonin2C receptors modulate dopamine transmission in the nucleus accumbens independently of dopamine release: Behavioral, neurochemical and molecular studies with cocaine. Addict. Biol. 2015, 20, 445–457. [Google Scholar] [CrossRef] [PubMed]
- Auclair, A.L.; Cathala, A.; Sarrazin, F.; Depoortère, R.; Piazza, P.V.; Newman-Tancredi, A.; Spampinato, U. The central serotonin2B receptor: A new pharmacological target to modulate the mesoaccumbens dopaminergic pathway activity. J. Neurochem. 2010, 114, 1323–1332. [Google Scholar] [CrossRef]
- Devroye, C.; Cathala, A.; Di Marco, B.; Caraci, F.; Drago, F.; Piazza, P.V.; Spampinato, U. Central serotonin2B receptor blockade inhibits cocaine-induced hyperlocomotion independently of changes of subcortical dopamine outflow. Neuropharmacology 2015, 97, 329–337. [Google Scholar] [CrossRef]
- Hoplight, B.J.; Vincow, E.S.; Neumaier, J.F. Cocaine increases 5-HT1B mRNA in rat nucleus accumbens shell neurons. Neuropharmacology 2007, 52, 444–449. [Google Scholar] [CrossRef]
- Crupi, R.; Marino, A.; Cuzzocrea, S. New Therapeutic Strategy for Mood Disorders. Curr. Med. Chem. 2011, 18, 4284–4298. [Google Scholar] [CrossRef]
- Bubar, M.J.; McMahon, L.R.; De Deurwaerdère, P.; Spampinato, U.; Cunningham, K.A. Selective serotonin reuptake inhibitors enhance cocaine-induced locomotor activity and dopamine release in the nucleus accumbens. Neuropharmacology 2003, 44, 342–353. [Google Scholar] [CrossRef]
- Caffino, L.; Mottarlini, F.; Targa, G.; Verheij, M.M.M.; Homberg, J.; Fumagalli, F. Long access to cocaine self-administration dysregulates the glutamate synapse in the nucleus accumbens core of serotonin transporter knockout rats. Br. J. Pharmacol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Caffino, L.; Verheij, M.M.M.; Roversi, K.; Targa, G.; Mottarlini, F.; Popik, P.; Nikiforuk, A.; Golebiowska, J.; Fumagalli, F.; Homberg, J.R. Hypersensitivity to amphetamine’s psychomotor and reinforcing effects in serotonin transporter knockout rats: Glutamate in the nucleus accumbens. Br. J. Pharmacol. 2020, 177, 4532–4547. [Google Scholar] [CrossRef] [PubMed]
- Hamon, M.; Blier, P. Monoamine neurocircuitry in depression and strategies for new treatments. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2013, 45, 54–63. [Google Scholar] [CrossRef]
- Diepenbroek, C.; Rijnsburger, M.; Eggels, L.; van Megen, K.M.; Ackermans, M.T.; Fliers, E.; Kalsbeek, A.; Serlie, M.J.; la Fleur, S.E. Infusion of fluoxetine, a serotonin reuptake inhibitor, in the shell region of the nucleus accumbens increases blood glucose concentrations in rats. Neurosci. Lett. 2017, 637, 85–90. [Google Scholar] [CrossRef]
- Cenci, M.A.; Kalén, P.; Mandel, R.J.; Björklund, A. Regional differences in the regulation of dopamine and noradrenaline release in medial frontal cortex, nucleus accumbens and caudate-putamen: A microdialysis study in the rat. Brain Res. 1992, 581, 217–228. [Google Scholar] [CrossRef]
- Vanderschuren, L.J.M.J.; Achterberg, E.J.M.; Trezza, V. The neurobiology of social play and its rewarding value in rats. Neurosci. Biobehav. Rev. 2016, 70, 86–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, F.C.; Liang, K.C. Interactions of the dorsal hippocampus, medial prefrontal cortex and nucleus accumbens in formation of fear memory: Difference in inhibitory avoidance learning and contextual fear conditioning. Neurobiol. Learn. Mem. 2014, 112, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Pal, P.; Raj, S.S.; Mohan, M.; Pal, G.K. Modulation of feeding and drinking behaviour by catecholamines injected into nucleus accumbens in rats. Indian J. Physiol. Pharmacol. 2000, 44, 24–32. [Google Scholar]
- Zhang, Y.; Qu, H.; Zhou, Y.; Wang, Y.; Zhang, D.; Yang, X.; Yang, C.X.; Xu, M.Y. The involvement of norepinephrine in pain modulation in the nucleus accumbens of morphine-dependent rats. Neurosci. Lett. 2015, 585, 6–11. [Google Scholar] [CrossRef]
- Reith, M.E.A.; Li, M.Y.; Yan, Q.S. Extracellular dopamine, norepinephrine, and serotonin in the ventral tegmental area and nucleus accumbens of freely moving rats during intracerebral dialysis following systemic administration of cocaine and other uptake blockers. Psychopharmacology 1997, 134, 309–317. [Google Scholar] [CrossRef]
- Vanderschuren, L.J.M.J.; Wardeh, G.; De Vries, T.J.; Mulder, A.H.; Schoffelmeer, A.N.M. Opposing role of dopamine D1 and D2 receptors in modulation of rat nucleus accumbens noradrenaline release. J. Neurosci. 1999, 19, 4123–4131. [Google Scholar] [CrossRef] [Green Version]
- Karkhanis, A.N.; Locke, J.L.; Mccool, B.A.; Weiner, J.L.; Jones, S.R. Social isolation rearing increases nucleus accumbens dopamine and norepinephrine responses to acute ethanol in adulthood. Alcohol. Clin. Exp. Res. 2014, 38, 2770–2779. [Google Scholar] [CrossRef] [Green Version]
- Mitrano, D.A.; Jackson, K.; Finley, S.; Seeley, A. α1b-Adrenergic Receptor Localization and Relationship to the D1-Dopamine Receptor in the Rat Nucleus Accumbens. Neuroscience 2018, 371, 126–137. [Google Scholar] [CrossRef] [PubMed]
- Ihalainen, J.A.; Tanila, H. In vivo regulation of dopamine and noradrenaline release by alpha2A-adrenoceptors in the mouse nucleus accumbens. J. Neurochem. 2004, 91, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Scofield, M.D. Exploring the Role of Astroglial Glutamate Release and Association with Synapses in Neuronal Function and Behavior. Biol. Psychiatry 2018, 84, 778–786. [Google Scholar] [CrossRef] [PubMed]
- Figley, C.R.; Stroman, P.W. The role(s) of astrocytes and astrocyte activity in neurometabolism, neurovascular coupling, and the production of functional neuroimaging signals. Eur. J. Neurosci. 2011, 33, 577–588. [Google Scholar] [CrossRef]
- Filosa, J.A.; Bonev, A.D.; Nelson, M.T. Calcium dynamics in cortical astrocytes and arterioles during neurovascular coupling. Circ. Res. 2004, 95, e73–e81. [Google Scholar] [CrossRef] [Green Version]
- Bull, C.; Freitas, K.C.C.; Zou, S.; Poland, R.S.; Syed, W.A.; Urban, D.J.; Minter, S.C.; Shelton, K.L.; Hauser, K.F.; Negus, S.S.; et al. Rat nucleus accumbens core astrocytes modulate reward and the motivation to self-administer ethanol after abstinence. Neuropsychopharmacology 2014, 39, 2835–2845. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez-Martos, M.; Girard, B.; Mendonça-Netto, S.; Perroy, J.; Valjent, E.; Maldonado, R.; Martin, M. Cafeteria diet induces neuroplastic modifications in the nucleus accumbens mediated by microglia activation. Addict. Biol. 2018, 23, 735–749. [Google Scholar] [CrossRef] [PubMed]
- Guegan, T.; Cutando, L.; Ayuso, E.; Santini, E.; Fisone, G.; Bosch, F.; Martinez, A.; Valjent, E.; Maldonado, R.; Martin, M. Operant behavior to obtain palatable food modifies neuronal plasticity in the brain reward circuit. Eur. Neuropsychopharmacol. 2013, 23, 146–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewitus, G.M.; Konefal, S.C.; Greenhalgh, A.D.; Pribiag, H.; Augereau, K.; Stellwagen, D. Microglial TNF-α Suppresses Cocaine-Induced Plasticity and Behavioral Sensitization. Neuron 2016, 90, 483–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Dietz, K.; Hodes, G.E.; Russo, S.J.; Casaccia, P. Widespread transcriptional alternations in oligodendrocytes in the adult mouse brain following chronic stress. Dev. Neurobiol. 2018, 78, 152–162. [Google Scholar] [CrossRef] [PubMed]
- Bonnefil, V.; Dietz, K.; Amatruda, M.; Wentling, M.; Aubry, A.V.; Dupree, J.L.; Temple, G.; Park, H.J.; Burghardt, N.S.; Casaccia, P.; et al. Region-specific myelin differences define behavioral consequences of chronic social defeat stress in mice. eLife 2019, 8, e40855. [Google Scholar] [CrossRef] [PubMed]
- Sacchet, M.D.; Gotlib, I.H. Myelination of the brain in major depressive disorder: An in vivo quantitative magnetic resonance imaging study. Sci. Rep. 2017, 7, 2200. [Google Scholar] [CrossRef]
- Kristiansen, L.V.; Bannon, M.J.; Meador-Woodruff, J.H. Expression of transcripts for myelin related genes in postmortem brain from cocaine abusers. Neurochem. Res. 2009, 34, 46–54. [Google Scholar] [CrossRef] [Green Version]
- Albertson, D.N.; Pruetz, B.; Schmidt, C.J.; Kuhn, D.M.; Kapatos, G.; Bannon, M.J. Gene expression profile of the nucleus accumbens of human cocaine abusers: Evidence for dysregulation of myelin. J. Neurochem. 2004, 88, 1211–1219. [Google Scholar] [CrossRef]
- Kovalevich, J.; Corley, G.; Yen, W.; Rawls, S.M.; Langford, D. Cocaine-induced loss of white matter proteins in the adult mouse nucleus accumbens is attenuated by administration of a β-lactam antibiotic during cocaine withdrawal. Am. J. Pathol. 2012, 181, 1921–1927. [Google Scholar] [CrossRef] [Green Version]
- Berridge, K.C.; Robinson, T.E.; Aldridge, J.W. Dissecting components of reward: “Liking”, “wanting”, and learning. Curr. Opin. Pharmacol. 2009, 9, 65–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swanson, L.W. Cerebral hemisphere regulation of motivated behavior. Brain Res. 2000, 886, 113–164. [Google Scholar] [CrossRef]
- Setlow, B. The nucleus accumbens and learning and memory. J. Neurosci. Res. 1997, 49, 515–521. [Google Scholar] [CrossRef]
- Kelley, A.E.; Baldo, B.A.; Pratt, W.E.; Will, M.J. Corticostriatal-hypothalamic circuitry and food motivation: Integration of energy, action and reward. Physiol. Behav. 2005, 86, 773–795. [Google Scholar] [CrossRef] [PubMed]
- Everitt, B.J. Sexual motivation: A neural and behavioural analysis of the mechanisms underlying appetitive and copulatory responses of male rats. Neurosci. Biobehav. Rev. 1990, 14, 217–232. [Google Scholar] [CrossRef]
- Kuhnen, C.M.; Knutson, B. The neural basis of financial risk taking. Neuron 2005, 47, 763–770. [Google Scholar] [CrossRef] [Green Version]
- Carr, G.D.; White, N.M. Conditioned place preference from intra-accumbens but not intra-caudate amphetamine injections. Life Sci. 1983, 33, 2551–2557. [Google Scholar] [CrossRef]
- Everitt, B.J.; Morris, K.A.; O’Brien, A.; Robbins, T.W. The basolateral amygdala-ventral striatal system and conditioned place preference: Further evidence of limbic-striatal interactions underlying reward-related processes. Neuroscience 1991, 42, 1–18. [Google Scholar] [CrossRef]
- McCullough, L.D.; Sokolowski, J.D.; Salamone, J.D. A neurochemical and behavioral investigation of the involvement of nucleus accumbens dopamine in instrumental avoidance. Neuroscience 1993, 52, 919–925. [Google Scholar] [CrossRef]
- Berns, G.S.; McClure, S.M.; Pagnoni, G.; Montague, P.R. Predictability modulates human brain response to reward. J. Neurosci. 2001, 21, 2793–2798. [Google Scholar] [CrossRef] [Green Version]
- Rebec, G.V.; Grabner, C.P.; Johnson, M.; Pierce, R.C.; Bardo, M.T. Transient increases in catecholaminergic activity in medial prefrontal cortex and nucleus accumbens shell during novelty. Neuroscience 1997, 76, 707–714. [Google Scholar] [CrossRef]
- Belin, D.; Jonkman, S.; Dickinson, A.; Robbins, T.W.; Everitt, B.J. Parallel and interactive learning processes within the basal ganglia: Relevance for the understanding of addiction. Behav. Brain Res. 2009, 199, 89–102. [Google Scholar] [CrossRef] [PubMed]
- Saddoris, M.P.; Sugam, J.A.; Cacciapaglia, F.; Carelli, R.M. Rapid dopamine dynamics in the accumbens core and shell: Learning and action. Front. Biosci.-Elit. Ed. 2013, 5, 273–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meredith, G.E.; Baldo, B.A.; Andrezjewski, M.E.; Kelley, A.E. The structural basis for mapping behavior onto the ventral striatum and its subdivisions. Brain Struct. Funct. 2008, 213, 17–27. [Google Scholar] [CrossRef] [Green Version]
- Floresco, S.B. The nucleus accumbens: An interface between cognition, emotion, and action. Annu. Rev. Psychol. 2015, 66, 25–32. [Google Scholar] [CrossRef]
- Parkinson, J.A.; Olmstead, M.C.; Burns, L.H.; Robbins, T.W.; Everitt, B.J. Dissociation in effects of lesions of the nucleus accumbens core and shell on appetitive Pavlovian approach behavior and the potentiation of conditioned reinforcement and locomotor activity by D-amphetamine. J. Neurosci. 1999, 19, 2401–2411. [Google Scholar] [CrossRef] [PubMed]
- van der Plasse, G.; Schrama, R.; van Seters, S.P.; Vanderschuren, L.J.M.J.; Westenberg, H.G.M. Deep brain stimulation reveals a dissociation of consummatory and motivated behaviour in the medial and lateral nucleus accumbens shell of the rat. PLoS ONE 2012, 7, e33455. [Google Scholar] [CrossRef] [Green Version]
- Alderson, H.L.; Parkinson, J.A.; Robbins, T.W.; Everitt, B.J. The effects of excitotoxic lesions of the nucleus accumbens core or shell regions on intravenous heroin self-administration in rats. Psychopharmacology 2001, 153, 455–463. [Google Scholar] [CrossRef]
- Bossert, J.M.; Poles, G.C.; Wihbey, K.A.; Koya, E.; Shaham, Y. Differential effects of blockade of dopamine D1-family receptors in nucleus accumbens core or shell on reinstatement of heroin seeking induced by contextual and discrete cues. J. Neurosci. 2007, 27, 12655–12663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bossert, J.M.; Gray, S.M.; Lu, L.; Shaham, Y. Activation of group II metabotropic glutamate receptors in the nucleus accumbens shell attenuates context-induced relapse to heroin seeking. Neuropsychopharmacology 2006, 31, 2197–2209. [Google Scholar] [CrossRef]
- Corbit, L.H.; Muir, J.L.; Balleine, B.W. The role of the nucleus accumbens in instrumental conditioning: Evidence of a functional dissociation between accumbens core and shell. J. Neurosci. 2001, 21, 3251–3260. [Google Scholar] [CrossRef] [Green Version]
- Basar, K.; Sesia, T.; Groenewegen, H.; Steinbusch, H.W.M.; Visser-Vandewalle, V.; Temel, Y. Nucleus accumbens and impulsivity. Prog. Neurobiol. 2010, 92, 533–557. [Google Scholar] [CrossRef] [PubMed]
- Cardinal, R.N.; Cheung, T.H.C. Nucleus accumbens core lesions retard instrumental learning and performance with delayed reinforcement in the rat. BMC Neurosci. 2005, 6, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parkinson, J.A.; Willoughby, P.J.; Robbins, T.W.; Everitt, B.J. Disconnection of the anterior cingulate cortex and nucleus accumbens core impairs pavlovian approach behavior: Further evidence for limbic cortical-ventral striatopallidal systems. Behav. Neurosci. 2000, 114, 42–63. [Google Scholar] [CrossRef] [PubMed]
- Maldonado-Irizarry, C.S.; Kelley, A.E. Excitatory amino acid receptors within nucleus accumbens subregions differentially mediate spatial learning in the rat. Behav. Pharmacol. 1995, 6, 527–539. [Google Scholar] [CrossRef]
- Salamone, J.D. The involvement of nucleus accumbens dopamine in appetitive and aversive motivation. Behav. Brain Res. 1994, 61, 117–133. [Google Scholar] [CrossRef]
- Wager, T.D.; Davidson, M.L.; Hughes, B.L.; Lindquist, M.A.; Ochsner, K.N. Prefrontal-Subcortical Pathways Mediating Successful Emotion Regulation. Neuron 2008, 59, 1037–1050. [Google Scholar] [CrossRef] [Green Version]
- Sturman, D.A.; Moghaddam, B. Striatum processes reward differently in adolescents versus adults. Proc. Natl. Acad. Sci. USA 2012, 109, 1719–1724. [Google Scholar] [CrossRef] [Green Version]
- Galvan, A.; Hare, T.A.; Parra, C.E.; Penn, J.; Voss, H.; Glover, G.; Casey, B.J. Earlier development of the accumbens relative to orbitofrontal cortex might underlie risk-taking behavior in adolescents. J. Neurosci. 2006, 26, 6885–6892. [Google Scholar] [CrossRef] [Green Version]
- Avital, A.; Richter-Levin, G. Exposure to juvenile stress exacerbates the behavioural consequences of exposure to stress in the adult rat. Int. J. Neuropsychopharmacol. 2005, 8, 163–173. [Google Scholar] [CrossRef] [Green Version]
- Caldji, C.; Diorio, J.; Meaney, M.J. Variations in maternal care in infancy regulate the development of stress reactivity. Biol. Psychiatry 2000, 48, 1164–1174. [Google Scholar] [CrossRef]
- Badowska-Szalewska, E.; Lietzau, G.; Moryś, J.; Kowiański, P. Role of brain-derived neurotrophic factor in shaping the behavioural response to environmental stressors. Folia Morphol. 2021, 80, 487–504. [Google Scholar] [CrossRef]
- Bazak, N.; Kozlovsky, N.; Kaplan, Z.; Matar, M.; Golan, H.; Zohar, J.; Richter-Levin, G.; Cohen, H. Pre-pubertal stress exposure affects adult behavioral response in association with changes in circulating corticosterone and brain-derived neurotrophic factor. Psychoneuroendocrinology 2009, 34, 844–858. [Google Scholar] [CrossRef] [PubMed]
- Bahtiyar, S.; Karaca, K.G.; Henckens, M.J.A.G.; Roozendaal, B. Norepinephrine and glucocorticoid effects on the brain mechanisms underlying memory accuracy and generalization. Mol. Cell. Neurosci. 2020, 108, 103537. [Google Scholar] [CrossRef] [PubMed]
- Daskalakis, N.P.; de Kloet, R.; Yehuda, R.; Malaspina, D.; Kranz, T.M. Early life stress effects on glucocorticoid—BDNF interplay in the hippocampus. Front. Mol. Neurosci. 2015, 8, 68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fenoglio, K.A.; Brunson, K.L.; Baram, T.Z. Hippocampal neuroplasticity induced by early-life stress: Functional and molecular aspects. Front. Neuroendocrinol. 2006, 27, 180–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dent, G.; Choi, D.C.; Herman, J.P.; Levine, S. GABAergic circuits and the stress hyporesponsive period in the rat: Ontogeny of glutamic acid decarboxylase (GAD) 67 mRNA expression in limbic-hypothalamic stress pathways. Brain Res. 2007, 1138, 1–9. [Google Scholar] [CrossRef]
- Rosenfeld, P.; Wetmore, J.B.; Levine, S. Effects of repeated maternal separations on the adrenocortical response to stress of preweanling rats. Physiol. Behav. 1992, 52, 787–791. [Google Scholar] [CrossRef]
- Sapolsky, R.M.; Meaney, M.J. Maturation of the adrenocortical stress response: Neuroendocrine control mechanisms and the stress hyporesponsive period. Brain Res. 1986, 396, 64–76. [Google Scholar] [CrossRef]
- Schmidt, M.; Enthoven, L.; Van Der Mark, M.; Levine, S.; De Kloet, E.R.; Oitzl, M.S. The postnatal development of the hypothalamic-pituitary-adrenal axis in the mouse. Int. J. Dev. Neurosci. 2003, 21, 125–132. [Google Scholar] [CrossRef]
- Schoenfeld, N.M.; Leathem, J.H.; Rabii, J. Maturation of adrenal stress responsiveness in the rat. Neuroendocrinology 1980, 31, 101–105. [Google Scholar] [CrossRef]
- McCormick, C.M.; Mathews, I.Z.; Thomas, C.; Waters, P. Investigations of HPA function and the enduring consequences of stressors in adolescence in animal models. Brain Cogn. 2010, 72, 73–85. [Google Scholar] [CrossRef]
- Bennett, M.R.; Lagopoulos, J. Stress and trauma: BDNF control of dendritic-spine formation and regression. Prog. Neurobiol. 2014, 112, 80–99. [Google Scholar] [CrossRef] [PubMed]
- Borges, J.V.; de Freitas, B.S.; Antoniazzi, V.; Santos, C.D.S.D.; Vedovelli, K.; Pires, V.N.; Paludo, L.; de Lima, M.N.M.; Bromberg, E. Social isolation and social support at adulthood affect epigenetic mechanisms, brain-derived neurotrophic factor levels and behavior of chronically stressed rats. Behav. Brain Res. 2019, 366, 36–44. [Google Scholar] [CrossRef]
- Wei, J.; Wang, J.; Dwyer, J.B.; Mangold, J.; Cao, J.; Leslie, F.M.; Li, M.D. Gestational nicotine treatment modulates cell death/survival-related pathways in the brains of adolescent female rats. Int. J. Neuropsychopharmacol. 2011, 14, 91–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, J.; Dwyer, J.B.; Mangold, J.E.; Wang, J.; Wei, J.; Leslie, F.M.; Li, M.D. Modulation of cell adhesion systems by prenatal nicotine exposure in limbic brain regions of adolescent female rats. Int. J. Neuropsychopharmacol. 2011, 14, 157–174. [Google Scholar] [CrossRef] [Green Version]
- Counotte, D.S.; Spijker, S.; Van De Burgwal, L.H.; Hogenboom, F.; Schoffelmeer, A.N.M.; De Vries, T.J.; Smit, A.B.; Pattij, T. Long-lasting cognitive deficits resulting from adolescent nicotine exposure in rats. Neuropsychopharmacology 2009, 34, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Natividad, L.A.; Tejeda, H.A.; Torres, O.V.; O’Dell, L.E. Nicotine withdrawal produces a decrease in extracellular levels of dopamine in the nucleus accumbens that is lower in adolescent versus adult male rats. Synapse 2010, 64, 136–145. [Google Scholar] [CrossRef] [Green Version]
- O’Dell, L.E. A psychobiological framework of the substrates that mediate nicotine use during adolescence. Neuropharmacology 2009, 56, 263–278. [Google Scholar] [CrossRef] [Green Version]
- Sershen, H.; Shearman, E.; Fallon, S.; Chakraborty, G.; Smiley, J.; Lajtha, A. The effects of acetaldehyde on nicotine-induced transmitter levels in young and adult brain areas. Brain Res. Bull. 2009, 79, 458–462. [Google Scholar] [CrossRef] [PubMed]
- Shearman, E.; Fallon, S.; Sershen, H.; Lajtha, A. Nicotine-induced monoamine neurotransmitter changes in the brain of young rats. Brain Res. Bull. 2008, 76, 626–639. [Google Scholar] [CrossRef]
- McDonald, C.G.; Eppolito, A.K.; Brielmaier, J.M.; Smith, L.N.; Bergstrom, H.C.; Lawhead, M.R.; Smith, R.F. Evidence for elevated nicotine-induced structural plasticity in nucleus accumbens of adolescent rats. Brain Res. 2007, 1151, 211–218. [Google Scholar] [CrossRef]
- Soderstrom, K.; Qin, W.; Williams, H.; Taylor, D.A.; McMillen, B.A. Nicotine increases FosB expression within a subset of reward- and memory-related brain regions during both peri- and post-adolescence. Psychopharmacology 2007, 191, 891–897. [Google Scholar] [CrossRef]
- Ernst, M.; Pine, D.S.; Hardin, M. Triadic model of the neurobiology of motivated behavior in adolescence. Psychol. Med. 2006, 36, 299–312. [Google Scholar] [CrossRef]
- Watt, M.J.; Weber, M.A.; Davies, S.R.; Forster, G.L. Impact of juvenile chronic stress on adult cortico-accumbal function: Implications for cognition and addiction. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2017, 79, 136–154. [Google Scholar] [CrossRef]
- Birnie, M.T.; Kooiker, C.L.; Short, A.K.; Bolton, J.L.; Chen, Y.; Baram, T.Z. Plasticity of the Reward Circuitry After Early-Life Adversity: Mechanisms and Significance. Biol. Psychiatry 2020, 87, 875–884. [Google Scholar] [CrossRef]
- Sellings, L.H.L.; Baharnouri, G.; McQuade, L.E.; Clarke, P.B.S. Rewarding and aversive effects of nicotine are segregated within the nucleus accumbens. Eur. J. Neurosci. 2008, 28, 342–352. [Google Scholar] [CrossRef]
- Laviolette, S.R.; Lauzon, N.M.; Bishop, S.F.; Sun, N.; Tan, H. Dopamine signaling through D1-like versus D2-like receptors in the nucleus accumbens core versus shell differentially modulates nicotine reward sensitivity. J. Neurosci. 2008, 28, 8025–8033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soares-Cunha, C.; de Vasconcelos, N.A.P.; Coimbra, B.; Domingues, A.V.; Silva, J.M.; Loureiro-Campos, E.; Gaspar, R.; Sotiropoulos, I.; Sousa, N.; Rodrigues, A.J. Nucleus accumbens medium spiny neurons subtypes signal both reward and aversion. Mol. Psychiatry 2020, 25, 3241–3255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, J.; Zhang, S.; Wang, H.L.; Barker, D.J.; Miranda-Barrientos, J.; Morales, M. VTA glutamatergic inputs to nucleus accumbens drive aversion by acting on GABAergic interneurons. Nat. Neurosci. 2016, 19, 725–733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Hasani, R.; McCall, J.G.; Shin, G.; Gomez, A.; Schmitz, G.P.; Bernardi, J.M.; Pyo, C.-O.; Park, S.I.; Marcinkiewcz, C.; Crowley, N.A.; et al. Distinct Subpopulations of Nucleus Accumbens Dynorphin Neurons Drive Aversion and Reward. Neuron 2015, 87, 1063–1077. [Google Scholar] [CrossRef] [Green Version]
- Isola, R.; Zhang, H.; Tejwani, G.A.; Neff, N.H.; Hadjiconstantinou, M. Acute nicotine changes dynorphin and prodynorphin mRNA in the striatum. Psychopharmacology 2009, 201, 507–516. [Google Scholar] [CrossRef]
- Isola, R.; Zhang, H.; Tejwani, G.A.; Neff, N.H.; Hadjiconstantinou, M. Dynorphin and prodynorphin mRNA changes in the striatum during nicotine withdrawal. Synapse 2008, 62, 448–455. [Google Scholar] [CrossRef]
- Qiao, H.; Li, M.-X.; Xu, C.; Chen, H.-B.; An, S.-C.; Ma, X.-M. Dendritic Spines in Depression: What We Learned from Animal Models. Neural Plast. 2016, 2016, 20–24. [Google Scholar] [CrossRef] [Green Version]
- Muhammad, A.; Carroll, C.; Kolb, B. Stress during development alters dendritic morphology in the nucleus accumbens and prefrontal cortex. Neuroscience 2012, 216, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Monroy, E.; Hernández-Torres, E.; Flores, G. Maternal separation disrupts dendritic morphology of neurons in prefrontal cortex, hippocampus, and nucleus accumbens in male rat offspring. J. Chem. Neuroanat. 2010, 40, 93–101. [Google Scholar] [CrossRef]
- Lai, K.O.; Ip, N.Y. Structural plasticity of dendritic spines: The underlying mechanisms and its dysregulation in brain disorders. Biochim. Biophys. Acta-Mol. Basis Dis. 2013, 1832, 2257–2263. [Google Scholar] [CrossRef] [Green Version]
- Bailey, C.H.; Kandel, E.R.; Harris, K.M. Structural components of synaptic plasticity and memory consolidation. Cold Spring Harb. Perspect. Biol. 2015, 7, a021758. [Google Scholar] [CrossRef] [Green Version]
- Bourne, J.N.; Harris, K.M. Balancing Structure and Function at Hippocampal Dendritic Spines. Annu. Rev. Neurosci. 2008, 31, 47–67. [Google Scholar] [CrossRef] [Green Version]
- Kasai, H.; Matsuzaki, M.; Noguchi, J.; Yasumatsu, N.; Nakahara, H. Structure-stability-function relationships of dendritic spines. Trends Neurosci. 2003, 26, 360–368. [Google Scholar] [CrossRef]
- Marsden, W.N. Synaptic plasticity in depression: Molecular, cellular and functional correlates. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2013, 43, 168–184. [Google Scholar] [CrossRef]
- Wang, Z.Z.; Yang, W.X.; Zhang, Y.; Zhao, N.; Zhang, Y.Z.; Liu, Y.Q.; Xu, Y.; Wilson, S.P.; O’Donnell, J.M.; Zhang, H.T.; et al. Phosphodiesterase-4D Knock-down in the Prefrontal Cortex Alleviates Chronic Unpredictable Stress-Induced Depressive-Like Behaviors and Memory Deficits in Mice. Sci. Rep. 2015, 5, 11332. [Google Scholar] [CrossRef] [Green Version]
- Gray, J.D.; Rubin, T.G.; Hunter, R.G.; McEwen, B.S. Hippocampal gene expression changes underlying stress sensitization and recovery. Mol. Psychiatry 2014, 19, 1171–1178. [Google Scholar] [CrossRef] [Green Version]
- Golden, S.A.; Russo, S.J. Mechanisms of psychostimulant-induced structural plasticity. Cold Spring Harb. Perspect. Med. 2012, 2, a011957. [Google Scholar] [CrossRef] [Green Version]
- Christian, D.T.; Wang, X.; Chen, E.L.; Sehgal, L.K.; Ghassemlou, M.N.; Miao, J.J.; Estepanian, D.; Araghi, C.H.; Stutzmann, G.E.; Wolf, M.E. Dynamic Alterations of Rat Nucleus Accumbens Dendritic Spines over 2 Months of Abstinence from Extended-Access Cocaine Self-Administration. Neuropsychopharmacology 2017, 42, 748–756. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Cahill, M.E.; Werner, C.T.; Christoffel, D.J.; Golden, S.A.; Xie, Z.; Loweth, J.A.; Marinelli, M.; Russo, S.J.; Penzes, P.; et al. Kalirin-7 mediates cocaine-induced AMPA receptor and spine plasticity, enabling incentive sensitization. J. Neurosci. 2013, 33, 11012–11022. [Google Scholar] [CrossRef] [Green Version]
- Bobadilla, A.C.; Dereschewitz, E.; Vaccaro, L.; Heinsbroek, J.A.; Scofield, M.D.; Kalivas, P.W. Cocaine and sucrose rewards recruit different seeking ensembles in the nucleus accumbens core. Mol. Psychiatry 2020, 25, 3150–3163. [Google Scholar] [CrossRef]
- Stankeviciute, N.M.; Scofield, M.D.; Kalivas, P.W.; Gipson, C.D. Rapid, transient potentiation of dendritic spines in context-induced relapse to cocaine seeking. Addict. Biol. 2014, 19, 972–974. [Google Scholar] [CrossRef] [Green Version]
- Cahill, M.E.; Bagot, R.C.; Gancarz, A.M.; Walker, D.M.; Sun, H.S.; Wang, Z.J.; Heller, E.A.; Feng, J.; Kennedy, P.J.; Koo, J.W.; et al. Bidirectional Synaptic Structural Plasticity after Chronic Cocaine Administration Occurs through Rap1 Small GTPase Signaling. Neuron 2016, 89, 566–582. [Google Scholar] [CrossRef] [Green Version]
- Goldin, M.; Segal, M. Protein kinase C and ERK involvement in dendritic spine plasticity in cultured rodent hippocampal neurons. Eur. J. Neurosci. 2003, 17, 2529–2539. [Google Scholar] [CrossRef] [PubMed]
- Anderson, E.M.; Wissman, A.M.; Chemplanikal, J.; Buzin, N.; Guzman, D.; Larson, E.B.; Neve, R.L.; Nestler, E.J.; Cowan, C.W.; Self, D.W. BDNF-TrkB controls cocaine-induced dendritic spines in rodent nucleus accumbens dissociated from increases in addictive behaviors. Proc. Natl. Acad. Sci. USA 2017, 114, 9469–9474. [Google Scholar] [CrossRef] [Green Version]
- Fox, M.E.; Chandra, R.; Menken, M.S.; Larkin, E.J.; Nam, H.; Engeln, M.; Francis, T.C.; Lobo, M.K. Dendritic remodeling of D1 neurons by RhoA/Rho-kinase mediates depression-like behavior. Mol. Psychiatry 2018, 25, 1022–1034. [Google Scholar] [CrossRef] [PubMed]
- Rigoni, D.; Avalos, M.P.; Boezio, M.J.; Guzmán, A.S.; Calfa, G.D.; Perassi, E.M.; Pierotti, S.M.; Bisbal, M.; Garcia-Keller, C.; Cancela, L.M.; et al. Stress-induced vulnerability to develop cocaine addiction depends on cofilin modulation. Neurobiol. Stress 2021, 15, 100349. [Google Scholar] [CrossRef] [PubMed]
- Caffino, L.; Giannotti, G.; Malpighi, C.; Racagni, G.; Fumagalli, F. Short-term withdrawal from developmental exposure to cocaine activates the glucocorticoid receptor and alters spine dynamics. Eur. Neuropsychopharmacol. 2015, 25, 1832–1841. [Google Scholar] [CrossRef] [PubMed]
- DePoy, L.M.; Gourley, S.L. Synaptic Cytoskeletal Plasticity in the Prefrontal Cortex Following Psychostimulant Exposure. Traffic 2015, 16, 919–940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newey, S.E.; Velamoor, V.; Govek, E.E.; Van Aelst, L. Rho GTPases, dendritic structure, and mental retardation. J. Neurobiol. 2005, 64, 58–74. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, A.Y.; Harms, M.B.; Luo, L. Small GTPases Rac and Rho in the maintenance of dendritic spines and branches in hippocampal pyramidal neurons. J. Neurosci. 2000, 20, 5329–5338. [Google Scholar] [CrossRef] [PubMed]
- Peck, J.W.; Oberst, M.; Bouker, K.B.; Bowden, E.; Burbelo, P.D. The RhoA-binding protein, rhophilin-2, regulates actin cytoskeleton organization. J. Biol. Chem. 2002, 277, 43924–43932. [Google Scholar] [CrossRef] [Green Version]
- Luo, L. Actin Cytoskeleton Regulation in Neuronal Morphogenesis and Structural Plasticity. Annu. Rev. Cell Dev. Biol. 2002, 18, 601–635. [Google Scholar] [CrossRef] [Green Version]
- Spijker, S.; Houtzager, S.W.J.; De Gunst, M.C.M.; De Boer, W.P.H.; Schoffelmeer, A.N.M.; Smit, A.B. Morphine exposure and abstinence define specific stages of gene expression in the rat nucleus accumbens. FASEB J. 2004, 18, 848–850. [Google Scholar] [CrossRef] [PubMed]
- Parekh, P.K.; Logan, R.W.; Ketchesin, K.D.; Becker-Krail, D.; Shelton, M.A.; Hildebrand, M.A.; Barko, K.; Huang, Y.H.; McClung, C.A. Cell-type-specific regulation of nucleus accumbens synaptic plasticity and cocaine reward sensitivity by the circadian protein, NPAS2. J. Neurosci. 2019, 39, 4657–4667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koepsell, H. General Overview of Organic Cation Transporters in Brain. In Handbook of Experimental Pharmacology; Springer: Berlin/Heidelberg, Germany, 2021. [Google Scholar]
- Hovde, M.J.; Larson, G.H.; Vaughan, R.A.; Foster, J.D. Model systems for analysis of dopamine transporter function and regulation. Neurochem. Int. 2019, 123, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Bahi, A. Dopamine transporter gene expression within the nucleus accumbens plays important role in the acquisition and reinstatement of ethanol-seeking behavior in mice. Behav. Brain Res. 2020, 381, 112475. [Google Scholar] [CrossRef] [PubMed]
- McGinnis, M.M.; Siciliano, C.A.; Jones, S.R. Dopamine D3 autoreceptor inhibition enhances cocaine potency at the dopamine transporter. J. Neurochem. 2016, 138, 821–829. [Google Scholar] [CrossRef]
- Gaval-Cruz, M.; Goertz, R.B.; Puttick, D.J.; Bowles, D.E.; Meyer, R.C.; Hall, R.A.; Ko, D.; Paladini, C.A.; Weinshenker, D. Chronic loss of noradrenergic tone produces β-arrestin2-mediated cocaine hypersensitivity and alters cellular D2 responses in the nucleus accumbens. Addict. Biol. 2016, 21, 35–48. [Google Scholar] [CrossRef] [Green Version]
- Siciliano, C.A.; Fordahl, S.C.; Jones, S.R. Cocaine self-administration produces long-lasting alterations in dopamine transporter responses to cocaine. J. Neurosci. 2016, 36, 7807–7816. [Google Scholar] [CrossRef] [Green Version]
- Mateo, Y.; Budygin, E.A.; John, C.E.; Jones, S.R. Role of serotonin in cocaine effects in mice with reduced dopamine transporter function. Proc. Natl. Acad. Sci. USA 2004, 101, 372–377. [Google Scholar] [CrossRef] [Green Version]
- Siciliano, C.A.; Saha, K.; Calipari, E.S.; Fordahl, S.C.; Chen, R.; Khoshbouei, H.; Jones, S.R. Amphetamine reverses escalated cocaine intake via restoration of dopamine transporter conformation. J. Neurosci. 2018, 38, 484–497. [Google Scholar] [CrossRef]
- Martin, B.J.; Naughton, B.J.; Thirtamara-Rajamani, K.; Yoon, D.J.; Han, D.D.; Devries, A.C.; Gu, H.H. Dopamine transporter inhibition is necessary for cocaine-induced increases in dendritic spine density in the nucleus accumbens. Synapse 2011, 65, 490–496. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Zeng, F.; Hu, Y.; Li, X. A mini-review of the role of glutamate transporter in drug addiction. Front. Neurol. 2019, 10, 1123. [Google Scholar] [CrossRef]
- Qi, C.; Guo, B.; Ren, K.; Yao, H.; Wang, M.; Sun, T.; Cai, G.; Liu, H.; Li, R.; Luo, C.; et al. Chronic inflammatory pain decreases the glutamate vesicles in presynaptic terminals of the nucleus accumbens. Mol. Pain 2018, 14, 1744806918781259. [Google Scholar] [CrossRef]
- Roberts, B.M.; Doig, N.M.; Brimblecombe, K.R.; Lopes, E.F.; Siddorn, R.E.; Threlfell, S.; Connor-Robson, N.; Bengoa-Vergniory, N.; Pasternack, N.; Wade-Martins, R.; et al. GABA uptake transporters support dopamine release in dorsal striatum with maladaptive downregulation in a parkinsonism model. Nat. Commun. 2020, 11, 4958. [Google Scholar] [CrossRef]
- Dannenhoffer, C.A.; Werner, D.F.; Varlinskaya, E.I.; Spear, L.P. Adolescent intermittent ethanol exposure does not alter responsiveness to ifenprodil or expression of vesicular GABA and glutamate transporters. Dev. Psychobiol. 2021, 63, 903–914. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Wang, G.; Ma, K.; Cui, S.; Wang, J.H. GABAergic neurons in nucleus accumbens are correlated to resilience and vulnerability to chronic stress for major depression. Oncotarget 2017, 8, 35933–35945. [Google Scholar] [CrossRef] [Green Version]
- Kasahara, Y.; Sakakibara, Y.; Hiratsuka, T.; Moriya, Y.; Lesch, K.P.; Hall, F.S.; Uhl, G.R.; Sora, I. Repeated methamphetamine treatment increases spine density in the nucleus accumbens of serotonin transporter knockout mice. Neuropsychopharmacol. Rep. 2019, 39, 130–133. [Google Scholar] [CrossRef] [PubMed]
- Verheij, M.M.M.; Karel, P.; Cools, A.R.; Homberg, J.R. Reduced cocaine-induced serotonin, but not dopamine and noradrenaline, release in rats with a genetic deletion of serotonin transporters. Eur. Neuropsychopharmacol. 2014, 24, 1850–1854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simmler, L.D.; Anacker, A.M.J.; Levin, M.H.; Vaswani, N.M.; Gresch, P.J.; Nackenoff, A.G.; Anastasio, N.C.; Stutz, S.J.; Cunningham, K.A.; Wang, J.; et al. Blockade of the 5-HT transporter contributes to the behavioural, neuronal and molecular effects of cocaine. Br. J. Pharmacol. 2017, 174, 2716–2738. [Google Scholar] [CrossRef]
- Carboni, E.; Silvagni, A. Dopamine reuptake by norepinephrine neurons: Exception or rule? Crit. Rev. Neurobiol. 2004, 16, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Carboni, E.; Silvagni, A.; Vacca, C.; Di Chiara, G. Cumulative effect of norepinephrine and dopamine carrier blockade on extracellular dopamine increase in the nucleus accumbens shell, bed nucleus of stria terminalis and prefrontal cortex. J. Neurochem. 2006, 96, 473–481. [Google Scholar] [CrossRef]
- Toth, M. Use of Mice with Targeted Genetic Inactivation in the Serotonergic System for the Study of Anxiety. In Serotonin Receptors in Neurobiology; CRC Press: Boca Raton, FL, USA; Taylor & Francis: Abingdon, UK, 2007; pp. 181–195. ISBN 0849339774. [Google Scholar]
- Biala, G. Behavioral and pharmacological characteristics of mice lacking the dopamine transporter. Postepy Hig. Med. Dosw. 2004, 58, 560–564. [Google Scholar]
- Jaber, M. Controle par la dopamine de fonctions neuroendocrines. Nouvelles donnees basees sur l’etude d’animaux transgeniques. Ann. Endocrinol. 1997, 58, 427–435. [Google Scholar]
- Lanfumey, L.; La Cour, C.M.; Froger, N.; Hamon, M. 5-HT-HPA Interactions in Two Models of Transgenic Mice Relevant to Major Depression. Neurochem. Res. 2000, 25, 1199–1206. [Google Scholar] [CrossRef]
- Nonkes, L.J.P.; Van Bussel, I.P.G.; Verheij, M.M.M.; Homberg, J.R. The interplay between brain 5-hydroxytryptamine levels and cocaine addiction. Behav. Pharmacol. 2011, 22, 723–738. [Google Scholar] [CrossRef]
- Karel, P.; Calabrese, F.; Riva, M.; Brivio, P.; Van der Veen, B.; Reneman, L.; Verheij, M.; Homberg, J. d-Cycloserine enhanced extinction of cocaine-induced conditioned place preference is attenuated in serotonin transporter knockout rats. Addict. Biol. 2018, 23, 120–129. [Google Scholar] [CrossRef]
- Verheij, M.M.M.; Contet, C.; Karel, P.; Latour, J.; van der Doelen, R.H.A.; Geenen, B.; van Hulten, J.A.; Meyer, F.; Kozicz, T.; George, O.; et al. Median and Dorsal Raphe Serotonergic Neurons Control Moderate Versus Compulsive Cocaine Intake. Biol. Psychiatry 2018, 83, 1024–1035. [Google Scholar] [CrossRef]
- Gainetdinov, R.R.; Jones, S.R.; Caron, M.G. Functional hyperdopaminergia in dopamine transporter knock-out mice. Biol. Psychiatry 1999, 46, 303–311. [Google Scholar] [CrossRef]
- Bobadilla, A.C.; Garcia-Keller, C.; Chareunsouk, V.; Hyde, J.; Camacho, D.M.; Heinsbroek, J.A.; Kalivas, P.W. Accumbens brain-derived neurotrophic factor (BDNF) transmission inhibits cocaine seeking. Addict. Biol. 2019, 24, 860–873. [Google Scholar] [CrossRef]
- Mottarlini, F.; Racagni, G.; Brambilla, P.; Fumagalli, F.; Caffino, L. Repeated cocaine exposure during adolescence impairs recognition memory in early adulthood: A role for BDNF signaling in the perirhinal cortex. Dev. Cogn. Neurosci. 2020, 43, 100789. [Google Scholar] [CrossRef]
- Li, X.; Wolf, M.E. Multiple faces of BDNF in cocaine addiction. Behav. Brain Res. 2015, 279, 240–254. [Google Scholar] [CrossRef] [Green Version]
- Kowiański, P.; Lietzau, G.; Czuba, E.; Waśkow, M.; Steliga, A.; Moryś, J. BDNF: A Key Factor with Multipotent Impact on Brain Signaling and Synaptic Plasticity. Cell. Mol. Neurobiol. 2018, 38, 579–593. [Google Scholar] [CrossRef]
- Badowska-Szalewska, E.; Ludkiewicz, B.; Krawczyk, R.; Moryś, J. Exposure to mild stress and brain derived neurotrophin factor (BDNF) immunoreactivity in the hypothalamic paraventricular (PVN) and supraoptic (SON) nuclei: Comparison between aged and adult rats. J. Chem. Neuroanat. 2016, 78, 57–64. [Google Scholar] [CrossRef]
- Perreault, M.L.; Fan, T.; O’Dowd, B.F.; George, S.R. Enhanced brain-derived neurotrophic factor signaling in the nucleus accumbens of juvenile rats. Dev. Neurosci. 2013, 35, 384–395. [Google Scholar] [CrossRef] [Green Version]
- Autry, A.E.; Monteggia, L.M. Brain-derived neurotrophic factor and neuropsychiatric disorders. Pharmacol. Rev. 2012, 64, 238–258. [Google Scholar] [CrossRef] [Green Version]
- Fumagalli, F.; Moro, F.; Caffino, L.; Orrù, A.; Cassina, C.; Giannotti, G.; Di Clemente, A.; Racagni, G.; Riva, M.A.; Cervo, L. Region-specific effects on BDNF expression after contingent or non-contingent cocaine i.v. self-administration in rats. Int. J. Neuropsychopharmacol. 2013, 16, 913–918. [Google Scholar] [CrossRef] [Green Version]
- Graham, D.L.; Edwards, S.; Bachtell, R.K.; DiLeone, R.J.; Rios, M.; Self, D.W. Dynamic BDNF activity in nucleus accumbens with cocaine use increases self-administration and relapse. Nat. Neurosci. 2007, 10, 1029–1037. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bayassi-Jakowicka, M.; Lietzau, G.; Czuba, E.; Steliga, A.; Waśkow, M.; Kowiański, P. Neuroplasticity and Multilevel System of Connections Determine the Integrative Role of Nucleus Accumbens in the Brain Reward System. Int. J. Mol. Sci. 2021, 22, 9806. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22189806
Bayassi-Jakowicka M, Lietzau G, Czuba E, Steliga A, Waśkow M, Kowiański P. Neuroplasticity and Multilevel System of Connections Determine the Integrative Role of Nucleus Accumbens in the Brain Reward System. International Journal of Molecular Sciences. 2021; 22(18):9806. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22189806
Chicago/Turabian StyleBayassi-Jakowicka, Martyna, Grazyna Lietzau, Ewelina Czuba, Aleksandra Steliga, Monika Waśkow, and Przemysław Kowiański. 2021. "Neuroplasticity and Multilevel System of Connections Determine the Integrative Role of Nucleus Accumbens in the Brain Reward System" International Journal of Molecular Sciences 22, no. 18: 9806. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22189806