
How Are Epigenetic Modifications Related to Cardiovascular Disease in Older Adults?

 ,
,  , , , and
, , , and 
Abstract
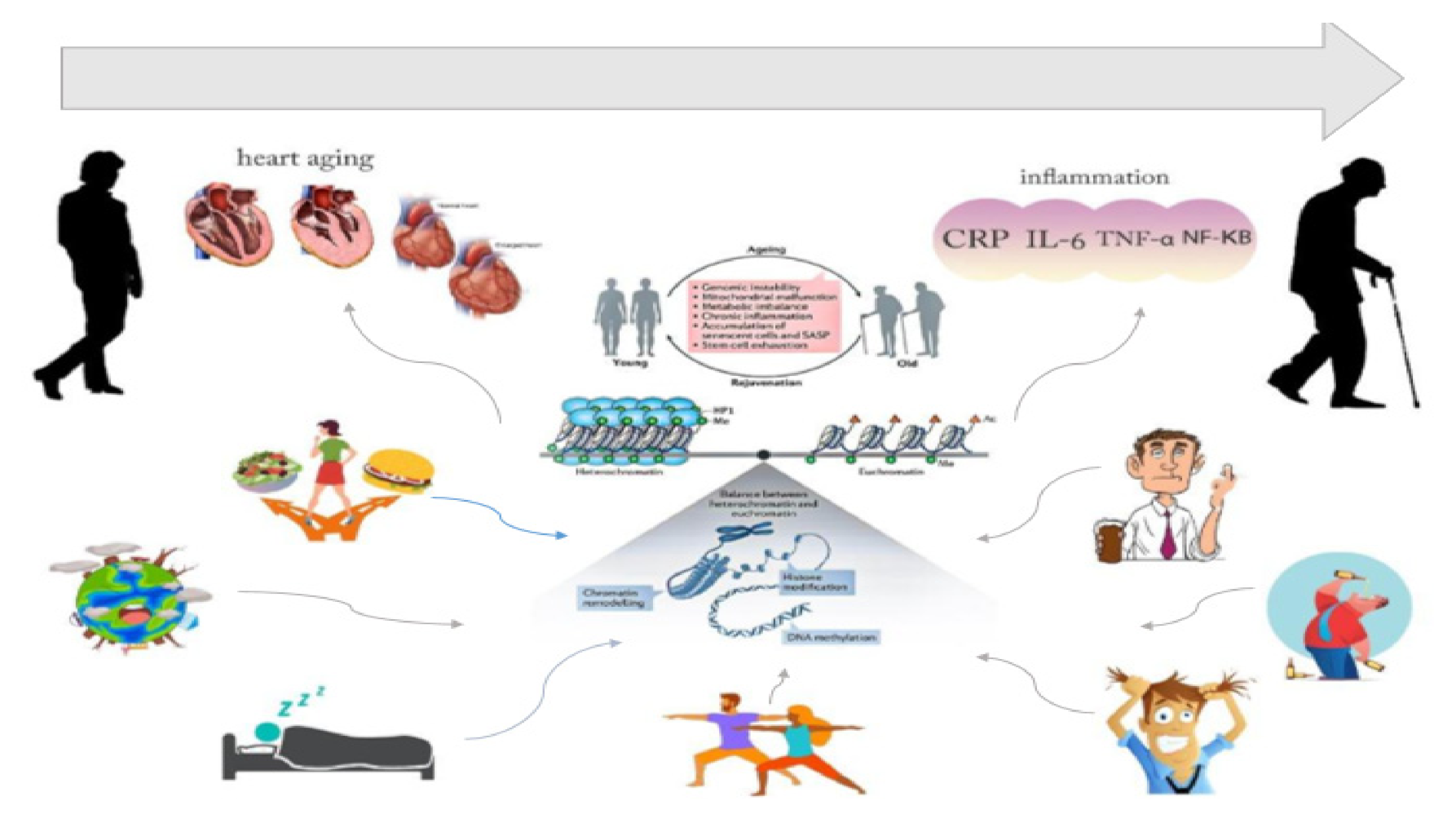
:1. Introduction
Aging, Cardiovascular Disease, and Epigenetics
2. Aging, CVD, and Epigenetic Modification
2.1. Age-Related DNA Methylation
2.2. The Effect of Aging on Histone Modifications
2.3. Aging-Related microRNAs (miRNAs)
3. Aging, Epigenetic Modification, and Inflammation
3.1. C-Reactive Protein (CRP)
3.2. Interleukin 6 (IL-6)
3.3. Tumor Necrosis Factor α (TNF-α)
4. Effect of Lifestyle and Environmental Factors on Epigenetic Modification in Older Adults with CVD
4.1. Nutritional Habits and Food Consumption
4.1.1. Nutrients
4.1.2. Phytochemicals and Representative Compounds
4.1.3. Trace Elements and Vitamins
4.2. Physical Activity
4.3. Smoking
4.4. Alcohol Consumption
4.5. Psychological Stress and Insufficient Sleep
4.6. Environmental Factors
4.6.1. Arsenic
4.6.2. Air Pollution
4.6.3. Aromatic Hydrocarbons and Other Organic Pollutants
4.6.4. Shift Work
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Ünsal, A.; Demir, G. The prevalence of chronic disease and drug use in the elderly in central Kirşehir. Turk. Geriatr. Derg. 2010, 13, 244–251. [Google Scholar]
- Lunenfeld, B. The ageing male: Demographics and challenges. World J. Urol. 2002, 20, 11–16. [Google Scholar] [CrossRef]
- Seals, D.R.; Justice, J.N.; Larocca, T.J. Physiological geroscience: Targeting function to increase healthspan and achieve optimal longevity. J. Physiol. 2016, 594, 2001–2024. [Google Scholar] [CrossRef]
- Mensah, G.A.; Brown, D.W. An overview of cardiovascular disease burden in the United States. Health Aff. 2007, 26, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.C.; Rabinovitch, P.S.; Kaeberlein, M. MTOR is a key modulator of ageing and age-related disease. Nature 2013, 493, 338–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otsuka, F.; Vorpahl, M.; Nakano, M.; Foerst, J.; Newell, J.B.; Sakakura, K.; Kutys, R.; Ladich, E.; Finn, A.V.; Kolodgie, F.D.; et al. Pathology of second-generation everolimus-eluting stents versus first-generation sirolimus- and paclitaxel-eluting stents in humans. Circulation 2014, 129, 211–223. [Google Scholar] [CrossRef] [Green Version]
- Fabrizio, P.; Pozza, F.; Pletcher, S.D.; Gendron, C.M.; Longo, V.D. Regulation of longevity and stress resistance by Sch9 in yeast. Science 2001, 292, 288–290. [Google Scholar] [CrossRef] [Green Version]
- Argano, C.; Scichilone, N.; Natoli, G.; Nobili, A.; Corazza, G.R.; Mannucci, P.M.; Perticone, F.; Corrao, S. Pattern of comorbidities and 1-year mortality in elderly patients with COPD hospitalized in internal medicine wards: Data from the RePoSI Registry. Intern. Emerg. Med. 2021, 16, 389–400. [Google Scholar] [CrossRef]
- Murphy, S.L.; Kratz, A.L.; Schepens Niemiec, S.L. Assessing Fatigability in the Lab and in Daily Life in Older Adults With Osteoarthritis Using Perceived, Performance, and Ecological Measures. J. Gerontol.-Ser. A Biol. Sci. Med. Sci. 2017, 72, 115–120. [Google Scholar] [CrossRef]
- Franceschi, C.; Campisi, J. Chronic inflammation (Inflammaging) and its potential contribution to age-associated diseases. J. Gerontol.-Ser. A Biol. Sci. Med. Sci. 2014, 69, S4–S9. [Google Scholar] [CrossRef]
- Fabbri, L.M.; Rabe, K.F. From COPD to Chronic Systemic Inflammatory Syndrome? Lancet 2007, 370, 797–799. Available online: http://0-search-ebscohost-com.brum.beds.ac.uk/login.aspx?direct=true&db=cin20&AN=105642877&site=ehost-live (accessed on 16 August 2021). [CrossRef]
- Barabási, A.L.; Gulbahce, N.; Loscalzo, J. Network medicine: A network-based approach to human disease. Nat. Rev. Genet. 2011, 12, 56–68. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Wang, J.; Wang, L.; Feng, G.; Li, G.; Yu, M.; Li, Y.; Liu, C.; Yuan, X.; Zang, G.; et al. Impaired lipid metabolism by age-dependent DNA methylation alterations accelerates aging. Proc. Natl. Acad. Sci. USA 2020, 117, 4328–4336. [Google Scholar] [CrossRef]
- Costantino, S.; Ambrosini, S.; Paneni, F. The epigenetic landscape in the cardiovascular complications of diabetes. J. Endocrinol. Investig. 2019, 42, 505–511. [Google Scholar] [CrossRef]
- Pagiatakis, C.; Musolino, E.; Gornati, R.; Bernardini, G.; Papait, R. Epigenetics of aging and disease: A brief overview. Aging Clin. Exp. Res. 2019, 33, 737–745. [Google Scholar] [CrossRef] [Green Version]
- Tosato, M.; Zamboni, V.; Ferrini, A.; Cesari1, M. The Aging Process and Potential Interventions to Extend Life Expectancy. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pmc/articles/pmc2685272/ (accessed on 24 July 2021).
- Kennedy, B.K.; Berger, S.L.; Brunet, A.; Campisi, J.; Cuervo, A.M.; Epel, E.S.; Franceschi, C.; Lithgow, G.J.; Morimoto, R.I.; Pessin, J.E.; et al. Commentary Geroscience: Linking Aging to Chronic Disease. Cell 2014, 159, 709–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, A.T.; Quach, A.; Wilson, J.G.; Reiner, A.P.; Aviv, A.; Raj, K.; Hou, L.; Baccarelli, A.A.; Li, Y.; Stewart, J.D.; et al. DNA methylation GrimAge strongly predicts lifespan and healthspan. Aging 2019, 11, 303–327. [Google Scholar] [CrossRef]
- Flavahan, W.A.; Gaskell, E.; Bernstein, B.E. Epigenetic plasticity and the hallmarks of cancer. Science 2017, 357, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papait, R.; Serio, S.; Pagiatakis, C.; Rusconi, F.; Carullo, P.; Mazzola, M.; Salvarani, N.; Miragoli, M.; Condorelli, G. Histone methyltransferase G9a is required for cardiomyocyte homeostasis and hypertrophy. Circulation 2017, 136, 1233–1246. [Google Scholar] [CrossRef]
- Ou, H.L.; Schumacher, B. DNA damage responses and p53 in the aging process. Blood 2018, 131, 488–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christensen, B.C.; Houseman, E.A.; Marsit, C.J.; Zheng, S.; Wrensch, M.R.; Wiemels, J.L.; Nelson, H.H.; Karagas, M.R.; Padbury, J.F.; Bueno, R.; et al. Aging and environmental exposures alter tissue-specific DNA methylation dependent upon CpG island context. PLoS Genet. 2009, 5, e1000602. [Google Scholar] [CrossRef] [Green Version]
- Roetker, N.S.; Pankow, J.S.; Bressler, J.; Morrison, A.C.; Boerwinkle, E. Prospective Study of Epigenetic Age Acceleration and Incidence of Cardiovascular Disease Outcomes in the ARIC Study (Atherosclerosis Risk in Communities). Circ. Genom. Precis. Med. 2018, 11, e001937. [Google Scholar] [CrossRef] [Green Version]
- Hannum, G.; Guinney, J.; Zhao, L.; Zhang, L.; Hughes, G.; Sadda, S.; Klotzle, B.; Bibikova, M.; Fan, J.-B.; Gao, Y.; et al. Genome-wide Methylation Profiles Reveal Quantitative Views of Human Aging Rates. Mol. Cell 2013, 49, 359–367. [Google Scholar] [CrossRef] [Green Version]
- Fraga, M.F.; Ballestar, E.; Paz, M.F.; Ropero, S.; Setien, F.; Ballestar, M.L.; Heine-Suñer, D.; Cigudosa, J.C.; Urioste, M.; Benitez, J.; et al. Epigenetic differences arise during the lifetime of monozygotic twins. Proc. Natl. Acad. Sci. USA 2005, 102, 10604–10609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grant, C.D.; Jafari, N.; Hou, L.; Li, Y.; Stewart, J.D.; Zhang, G.; Lamichhane, A.; Manson, J.E.; Baccarelli, A.A.; Whitsel, E.A.; et al. A longitudinal study of DNA methylation as a potential mediator of age-related diabetes risk. GeroScience 2017, 39, 475–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horvath, S.; Ritz, B.R. Increased Epigenetic Age and Granulocyte Counts in the Blood of Parkinson’s Disease Patients. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pmc/articles/PMC4712337/ (accessed on 24 July 2021).
- Slieker, R.C.; van Iterson, M.; Luijk, R.; Beekman, M.; Zhernakova, D.V.; Moed, M.H.; Mei, H.; van Galen, M.; Deelen, P.; Bonder, M.J.; et al. Age-related accrual of methylomic variability is linked to fundamental ageing mechanisms. Genome Biol. 2016, 17, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Lim, U.; Song, M. ADNA Methylation as a Biomarker of Aging in Epidemiologic Studies. Methods Mol. Biol. 2018, 1856, 219–231. [Google Scholar] [CrossRef] [PubMed]
- Marioni, R.E.; Shah, S.; McRae, A.F.; Chen, B.H.; Colicino, E.; Harris, S.E.; Gibson, J.; Henders, A.K.; Redmond, P.; Cox, S.R.; et al. DNA methylation age of blood predicts all-cause mortality in later life. Genome Biol. 2015, 16, 1–12. [Google Scholar] [CrossRef] [Green Version]
- McClay, J.L.; Aberg, K.A.; Clark, S.L.; Nerella, S.; Kumar, G.; Xie, L.Y.; Hudson, A.D.; Harada, A.; Hultman, C.M.; Magnusson, P.K.; et al. A methylome-wide study of aging using massively parallel sequencing of the methyl-CpG-enriched genomic fraction from blood in over 700 subjects. Hum. Mol. Genet. 2014, 23, 1175–1185. [Google Scholar] [CrossRef] [Green Version]
- Kalebic, T. Epigenetic transitions: Towards therapeutic targets. Expert Opin. Ther. Targets 2003, 7, 693–699. [Google Scholar] [CrossRef]
- McKay, J.A.; Mathers, J.C. Diet induced epigenetic changes and their implications for health. Acta Physiol. 2011, 202, 103–118. [Google Scholar] [CrossRef]
- Zhang, Y.; Zeng, C. Role of DNA methylation in cardiovascular diseases. Clin. Exp. Hypertens. 2016, 38, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Pelisek, J.; Jin, Z.G. Atherosclerosis Is an Epigenetic Disease. Trends Endocrinol. Metab. 2018, 29, 739–742. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Azhar, G.; Wei, J.Y. The Expression of microRNA and microRNA Clusters in the Aging Heart. PLoS ONE 2012, 7, e34688. [Google Scholar] [CrossRef] [Green Version]
- Thum, T.; Gross, C.; Fiedler, J.; Fischer, T.; Kissler, S.; Bussen, M.; Galuppo, P.; Just, S.; Rottbauer, W.; Frantz, S.; et al. MicroRNA-21 contributes to myocardial disease by stimulating MAP kinase signalling in fibroblasts. Nature 2008, 456, 980–984. [Google Scholar] [CrossRef] [PubMed]
- Ettig, R.; Kepper, N.; Stehr, R.; Wedemann, G.; Rippe, K. Dissecting DNA-Histone Interactions in the Nucleosome by Molecular Dynamics Simulations of DNA Unwrapping. Biophys J. 2011, 101, 1999–2008. [Google Scholar] [CrossRef] [Green Version]
- Maro, G.S.; Han, S.; Banko, M.R.; Gozani, O.; Brunet, A. HHS Public Access. Popul. Stud. 2011, 466, 383–387. [Google Scholar]
- Saul, D.; Kosinsky, R.L. Epigenetics of Aging and Aging-Associated Diseases. Int. J. Mol. Sci. 2021, 22, 401. [Google Scholar] [CrossRef]
- Mitchell, G.F. Arterial Stiffness and Hypertension. Hypertension 2014, 64, 13–18. [Google Scholar] [CrossRef] [Green Version]
- Dallapiccola, B.; Mingarelli, R.; Digilio, M.C.; Marino, B.; Novelli, G. Genetics of congenital heart diseases. G. Ital. Cardiol. 1994, 24, 155–166. [Google Scholar]
- Kubben, N.; Misteli, T. Shared molecular and cellular mechanisms of premature ageing and ageing-associated diseases. Nat. Rev. Mol. Cell Biol. 2017, 18, 595–609. [Google Scholar] [CrossRef]
- Etchegaray, J.P.; Mostoslavsky, R. Interplay between Metabolism and Epigenetics: A Nuclear Adaptation to Environmental Changes. Mol. Cell 2016, 62, 695–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greißel, A.; Culmes, M.; Burgkart, R.; Zimmermann, A.; Eckstein, H.H.; Zernecke, A.; Pelisek, J. Histone acetylation and methylation significantly change with severity of atherosclerosis in human carotid plaques. Cardiovasc. Pathol. 2016, 25, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Lee, Y.; Yeom, K.H.; Nam, J.W.; Heo, I.; Rhee, J.K.; Sohn, S.Y.; Cho, Y.; Zhang, B.T.; Kim, V.N. Molecular Basis for the Recognition of Primary microRNAs by the Drosha-DGCR8 Complex. Cell 2006, 125, 887–901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartel, D.P. Review MicroRNAs: Target Recognition and Regulatory Functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [Green Version]
- D’andrade, P.N.; Fulmer-Smentek, S. Agilent MicroRNA microarray profiling system. Methods Mol. Biol. 2012, 822, 85–102. [Google Scholar] [PubMed]
- Gupta, S.K.; Foinquinos, A.; Thum, S.; Remke, J.; Zimmer, K.; Bauters, C.; de Groote, P.; Boon, R.A.; de Windt, L.J.; Preissl, S.; et al. Preclinical Development of a MicroRNA-Based Therapy for Elderly Patients With Myocardial Infarction. J. Am. Coll. Cardiol. 2016, 68, 1557–1571. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Liu, Y.J.; Liu, T.; Zhang, H.; Yang, S.J. Plasma microRNA-21 is a potential diagnostic biomarker of acute myocardial infarction. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 323–329. [Google Scholar]
- Kontaraki, J.E.; Marketou, M.E.; Zacharis, E.A.; Parthenakis, F.I.; Vardas, P.E. Differential expression of vascular smooth muscle-modulating microRNAs in human peripheral blood mononuclear cells: Novel targets in essential hypertension. J. Hum. Hypertens. 2014, 28, 510–516. [Google Scholar] [CrossRef]
- Gao, Y.; Dai, M.; Liu, H.; He, W.; Lin, S.; Yuan, T.; Chen, H.; Dai, S. Diagnostic value of circulating miR-21: An update meta-analysis in various cancers and validation in endometrial cancer. Oncotarget 2016, 7, 68894–68908. [Google Scholar] [CrossRef] [Green Version]
- Osipova, J.; Fischer, D.C.; Dangwal, S.; Volkmann, I.; Widera, C.; Schwarz, K.; Lorenzen, J.M.; Schreiver, C.; Jacoby, U.; Heimhalt, M.; et al. Diabetes-associated MicroRNAs in pediatric patients with type 1 diabetes mellitus: A cross-sectional cohort study. J. Clin. Endocrinol. Metab. 2014, 99, E1661–E1665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, S.; Khanna, S.; Hussain, S.R.A.; Biswas, S.; Azad, A.; Rink, C.; Gnyawali, S.; Shilo, S.; Nuovo, G.J.; Sen, C.K. MicroRNA expression in response to murine myocardial infarction: MiR-21 regulates fibroblast metalloprotease-2 via phosphatase and tensin homologue. Cardiovasc. Res. 2009, 82, 21–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boon, R.A.; Seeger, T.; Heydt, S.; Fischer, A.; Hergenreider, E.; Horrevoets, A.J.G.; Vinciguerra, M.; Rosenthal, N.; Sciacca, S.; Pilato, M.; et al. MicroRNA-29 in aortic dilation: Implications for aneurysm formation. Circ. Res. 2011, 109, 1115–1119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, H.; Hata, A. MicroRNA regulation of smooth muscle gene expression and phenotype. Curr. Opin. Hematol. 2012, 19, 224–231. [Google Scholar] [CrossRef]
- Wu, X.; Du, X.; Yang, Y.; Liu, X.; Liu, X.; Zhang, N.; Li, Y.; Jiang, X.; Jiang, Y.; Yang, Z. Inhibition of miR-122 reduced atherosclerotic lesion formation by regulating NPAS3-mediated endothelial to mesenchymal transition. Life Sci. 2021, 265, 118816. [Google Scholar] [CrossRef]
- Hulsmans, M.; Sinnaeve, P.; Van Der Schueren, B.; Mathieu, C.; Janssens, S.; Holvoet, P. Decreased miR-181a expression in monocytes of obese patients is associated with the occurrence of metabolic syndrome and coronary artery disease. J. Clin. Endocrinol. Metab. 2012, 97, 1213–1218. [Google Scholar] [CrossRef] [Green Version]
- Marques, F.Z.; Campain, A.E.; Tomaszewski, M.; Zukowska-Szczechowska, E.; Yang, Y.H.J.; Charchar, F.J.; Morris, B.J. Gene expression profiling reveals renin mRNA overexpression in human hypertensive kidneys and a role for microRNAs. Hypertension 2011, 58, 1093–1098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olivieri, F.; Spazzafumo, L.; Santini, G.; Lazzarini, R.; Albertini, M.C.; Rippo, M.R.; Galeazzi, R.; Abbatecola, A.M.; Marcheselli, F.; Monti, D.; et al. Age-related differences in the expression of circulating microRNAs: MiR-21 as a new circulating marker of inflammaging. Mech. Ageing Dev. 2012, 133, 675–685. [Google Scholar] [CrossRef] [PubMed]
- Paneni, F.; Beckman, J.A.; Creager, M.A.; Cosentino, F. Diabetes and vascular disease: Pathophysiology, clinical consequences, and medical therapy: Part i. Eur. Heart J. 2013, 34, 2436–2446. [Google Scholar] [CrossRef]
- Donato, A.J.; Pierce, G.L.; Lesniewski, L.A.; Seals, D.R. Role of NFkappaB in age-related vascular endothelial dysfunction in humans. Aging 2009, 1, 678–680. [Google Scholar] [CrossRef]
- Fabbri, M.; Paone, A.; Calore, F.; Galli, R.; Gaudio, E.; Santhanam, R.; Lovat, F.; Fadda, P.; Mao, C.; Nuovo, G.J.; et al. MicroRNAs bind to Toll-like receptors to induce prometastatic inflammatory response. Proc. Natl. Acad. Sci. USA 2012, 109, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Olivieri, F.; Rippo, M.R.; Monsurrò, V.; Salvioli, S.; Capri, M.; Procopio, A.D.; Franceschi, C. MicroRNAs linking inflamm-aging, cellular senescence and cancer. Ageing Res. Rev. 2013, 12, 1056–1068. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, Y.; Saito, T.; Ogihara, T.; Ishigaki, Y.; Yamada, T.; Imai, J.; Uno, K.; Gao, J.; Kaneko, K.; Shimosawa, T.; et al. Blockade of the nuclear factor-κB pathway in the endothelium prevents insulin resistance and prolongs life spans. Circulation 2012, 125, 1122–1133. [Google Scholar] [CrossRef] [Green Version]
- Kundu, J.K.; Surh, Y.J. Emerging avenues linking inflammation and cancer. Free Radic. Biol. Med. 2012, 52, 2013–2037. [Google Scholar] [CrossRef] [PubMed]
- Olivieri, F.; Lazzarini, R.; Recchioni, R.; Marcheselli, F.; Rippo, M.R.; Di Nuzzo, S.; Albertini, M.C.; Graciotti, L.; Babini, L.; Mariotti, S.; et al. MiR-146a as marker of senescence-Associated pro-inflammatory status in cells involved in vascular remodelling. Age 2013, 35, 1157–1172. [Google Scholar] [CrossRef] [Green Version]
- Cheng, M.; Wu, G.; Song, Y.; Wang, L.; Tu, L.; Zhang, L.; Zhang, C. Celastrol-induced suppression of the MiR-21/ERK signalling pathway attenuates cardiac fibrosis and dysfunction. Cell Physiol. Biochem. 2016, 38, 1928–1938. Available online: https://www.karger.com/Article/Abstract/445554 (accessed on 30 July 2021). [CrossRef]
- Huang, Z.P.; Wang, D.Z. MiR-22 in cardiac remodeling and disease. Trends Cardiovasc. Med. 2014, 24, 267–272. [Google Scholar] [CrossRef] [Green Version]
- van Almen, G.C.; Verhesen, W.; van Leeuwen, R.E.W.; van de Vrie, M.; Eurlings, C.; Schellings, M.W.M.; Swinnen, M.; Cleutjens, J.P.M.; van Zandvoort, M.A.M.J.; Heymans, S.; et al. MicroRNA-18 and microRNA-19 regulate CTGF and TSP-1 expression in age-related heart failure. Aging Cell 2011, 10, 769–779. [Google Scholar] [CrossRef] [Green Version]
- Du, W.W.; Li, X.; Li, T.; Li, H.; Khorshidi, A.; Liu, F.; Yang, B.B. The microRNA miR-17-3p inhibits mouse cardiac fibroblast senescence by targeting Par4. J. Cell Sci. 2015, 128, 293–304. [Google Scholar]
- Xia, W.; Chen, H.; Chen, D.; Ye, Y.; Xie, C.; Hou, M. PD-1 inhibitor inducing exosomal miR-34a-5p expression mediates the cross talk between cardiomyocyte and macrophage in immune checkpoint inhibitor–related cardiac dysfunction. J. Immunother. Cancer 2020, 8, e001293. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pmc/articles/PMC7594538/ (accessed on 30 July 2021). [CrossRef] [PubMed]
- Tabuchi, T.; Satoh, M.; Itoh, T.; Nakamura, M. MicroRNA-34a regulates the longevity-associated protein SIRT1 in coronary artery disease: Effect of statins on SIRT1 and microRNA-34a expression. Clin. Sci. 2012, 123, 161–171. [Google Scholar] [CrossRef] [PubMed]
- de Yébenes, V.G.; Briones, A.M.; Martos-Folgado, I.; Mur, S.M.; Oller, J.; Bilal, F.; González-Amor, M.; Méndez-Barbero, N.; Silla-Castro, J.C.; Were, F.; et al. Aging-Associated miR-217 Aggravates Atherosclerosis and Promotes Cardiovascular Dysfunction. Arterioscler. Thromb. Vasc. Biol. 2020, 40, 2408–2424. [Google Scholar] [CrossRef] [PubMed]
- Van Rooij, E.; Sutherland, L.B.; Thatcher, J.E.; DiMaio, J.M.; Naseem, R.H.; Marshall, W.S.; Hill, J.A.; Olson, E.N. Dysregulation of microRNAs after myocardial infarction reveals a role of miR-29 in cardiac fibrosis. Proc. Natl. Acad. Sci. USA 2008, 105, 13027–13032. [Google Scholar] [CrossRef] [Green Version]
- Xiong, X.D.; Cho, M.; Cai, X.P.; Cheng, J.; Jing, X.; Cen, J.M.; Liu, X.; Yang, X.L.; Suh, Y. A common variant in pre-miR-146 is associated with coronary artery disease risk and its mature miRNA expression. Mutat. Res.-Fundam. Mol. Mech. Mutagenesis 2014, 761, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Hazra, S.; Henson, G.D.; Morgan, R.G.; Breevoort, S.R.; Ives, S.J.; Richardson, R.S.; Donato, A.J.; Lesniewski, L.A. Experimental reduction of miR-92a mimics arterial aging. Exp. Gerontol. 2016, 83, 165–170. [Google Scholar] [CrossRef] [Green Version]
- Pin, A.L.; Houle, F.; Guillonneau, M.; Paquet, É.R.; Simard, M.J.; Huot, J. MIR-20a represses endothelial cell migration by targeting MKK3 and inhibiting p38 MAP kinase activation in response to VEGF. Angiogenesis 2012, 15, 593–608. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Lian, Y.; Wen, X.; Guo, J.; Wang, Z.; Jiang, S.; Hu, Y. Expression of miR-126 and its potential function in coronary artery disease. Afr. Health Sci. 2017, 17, 474–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuo, J.; Tsai, H.; Lin, C.; Lee, C.; Lee, P.; Ruan, Y.; Chiu, J.; Lee, D. Low Levels of MicroRNA-10a in Cardiovascular Endothelium and Blood Serum Are Related to Human Atherosclerotic Disease. Cardiol. Res. Pract. 2021, 2021. [Google Scholar] [CrossRef] [PubMed]
- Loscalzo, J.; Handy, D.E. Epigenetic modifications: Basic mechanisms and role in cardiovascular disease (2013 Grover Conference series). Pulm. Circ. 2014, 4, 169–174. [Google Scholar] [CrossRef] [Green Version]
- Nie, X.; Fan, J.; Li, H.; Yin, Z.; Zhao, Y.; Dai, B.; Dong, N.; Chen, C.; Wang, D.W. miR-217 Promotes Cardiac Hypertrophy and Dysfunction by Targeting PTEN. Mol. Ther.-Nucleic Acids 2018, 12, 254–266. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.G.; Zheng, B.; Han, M.; Fang, X.M.; Li, H.X.; Miao, S.B.; Su, M.; Han, Y.; Shi, H.J.; Wen, J.K. MiR-146a and Krüppel-like factor 4 form a feedback loop to participate in vascular smooth muscle cell proliferation. EMBO Rep. 2011, 12, 56–62. [Google Scholar] [CrossRef] [Green Version]
- Danielson, L.S.; Park, D.S.; Rotllan, N.; Chamorro-Jorganes, A.; Guijarro, M.V.; Fernandez-Hernando, C.; Fishman, G.I.; Phoon, C.K.L.; Hernando, E. Cardiovascular dysregulation of miR-17-92 causes a lethal hypertrophic cardiomyopathy and arrhythmogenesis. FASEB J. 2013, 27, 1460–1467. [Google Scholar] [CrossRef] [Green Version]
- Franceschi, C.; Bonafè, M.; Valensin, S.; Olivieri, F.; De Luca, M.; Ottaviani, E.; De Benedictis, G. Inflamm-aging. An evolutionary perspective on immunosenescence. Ann. N.Y. Acad. Sci. 2000, 908, 244–254. [Google Scholar] [CrossRef]
- Silveira, E.A.; Vaseghi, G.; de Carvalho Santos, A.S.; Kliemann, N.; Masoudkabir, F.; Noll, M.; Mohammadifard, N.; Sarrafzadegan, N.; de Oliveira, C. Visceral obesity and its shared role in cancer and cardiovascular disease: A scoping review of the pathophysiology and pharmacological treatments. Int. J. Mol. Sci. 2020, 21, 1–18. [Google Scholar]
- Incalza, M.A.; Oria, R.D.; Natalicchio, A.; Perrini, S.; Laviola, L.; Giorgino, F. Oxidative stress and reactive oxygen species in endothelial dysfunction associated with cardiovascular and metabolic diseases. Vasc. Pharmacol. 2017, 100, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Chae, C.U.; Lee, R.T.; Rifai, N.; Ridker, P.M. Blood pressure and inflammation in apparently healthy men. Hypertension 2001, 38, 399–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hage, F.G.; Szalai, A.J. C-Reactive Protein Gene Polymorphisms, C-Reactive Protein Blood Levels, and Cardiovascular Disease Risk. J. Am. Coll. Cardiol. 2007, 50, 1115–1122. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.V.; Lazarus, A.; Smith, J.A.; Chuang, Y.H.; Zhao, W.; Turner, S.T.; Kardia, S.L.R. Gene-specific DNA methylation association with serum levels of C-reactive protein in African Americans. PLoS ONE 2013, 8, e73480. [Google Scholar]
- Lepeule, J.; Baccarelli, A.; Tarantini, L.; Motta, V.; Cantone, L.; Litonjua, A.A.; Sparrow, D.; Vokonas, P.S.; Schwartz, J. Gene promoter methylation is associated with lung function in the elderly: The normative aging study. Epigenetics 2012, 7, 261–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aslibekyan, S.; Agha, G.; Colicino, E.; Do, A.N.; Lahti, J.; Ligthart, S.; Marioni, R.E.; Marzi, C.; Mendelson, M.M.; Tanaka, T.; et al. Association of methylation signals with incident coronary heart disease in an epigenome-wide assessment of circulating tumor necrosis factor. JAMA Cardiol. 2018, 3, 463–472. [Google Scholar] [CrossRef] [Green Version]
- Marques-Rocha, J.L.; Milagro, F.I.; Mansego, M.L.; Mourão, D.M.; Martínez, J.A.; Bressan, J. LINE-1 methylation is positively associated with healthier lifestyle but inversely related to body fat mass in healthy young individuals. Epigenetics 2016, 11, 49–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hermsdorff, H.H.; Mansego, M.L.; Campión, J.; Milagro, F.I.; Zulet, M.A.; Martínez, J.A. TNF-alpha promoter methylation in peripheral white blood cells: Relationship with circulating TNFα, truncal fat and n-6 PUFA intake in young women. Cytokine 2013, 64, 265–271. [Google Scholar] [CrossRef]
- Zhao, Y.; Shao, F. NLRC5: A NOD-like receptor protein with many faces in immune regulation. Cell Res. 2012, 22, 1099–1101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, Y.; Wang, Y.; Chen, F.; Huang, Y.; Zhu, S.; Leng, Q.; Wang, H.; Shi, Y.; Qian, Y. NLRC5 regulates MHC class I antigen presentation in host defense against intracellular pathogens. Cell Res. 2012, 22, 836–847. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Mao, D.; Roswit, W.T.; Jin, X.; Patel, A.C.; Patel, D.A.; Agapov, E.; Wang, Z.; Tidwell, R.M.; Atkinson, J.J.; et al. PARP9-DTX3L ubiquitin ligase targets host histone H2BJ and viral 3C protease to enhance interferon signaling and control viral infection. Nat. Immunol. 2015, 16, 1215–1227. [Google Scholar] [CrossRef]
- Melzer, D.; Perry, J.R.; Hernandez, D.; Corsi, A.M.; Stevens, K.; Rafferty, I.; Lauretani, F.; Murray, A.; Gibbs, J.R.; Paolisso, G.; et al. A genome-wide association study identifies protein quantitative trait loci (pQTLs). PLoS Genet. 2008, 4, e1000072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, D.T.; Zee, R.Y.L.; Danik, J.S.; Kozlowski, P.; Chasman, D.I.; Lazarus, R.; Cook, N.R.; Ridker, P.M.; Kwiatkowski, D.J. Association of common CRP gene variants with CRP levels and cardiovascular events. Ann. Hum. Genet. 2005, 69, 623–638. [Google Scholar] [CrossRef]
- Wannamethee, G.G.; Whincup, P.H.; Rumley, A.; Lowe, G.D.O. Inter-relationships of interleukin-6, cardiovascular risk factors and the metabolic syndrome among older men. J. Thromb. Haemost. 2007, 5, 1637–1643. [Google Scholar] [CrossRef]
- Urschel, K.; Cicha, I. TNF-α in the cardiovascular system: From physiology to therapy. Int. J. Interferon Cytokine Mediat. Res. 2015, 2015, 9–25. Available online: https://www.dovepress.com/tnf-alpha-in-the-cardiovascular-system-from-physiology-to-therapy-peer-reviewed-fulltext-article-IJICMR?source=content_type%3Areact%7Cfirst_level_url%3Aarticle%7Csection%3Amain_content%7Cbutton%3Abody_link (accessed on 30 July 2021).
- Chong, Z.Z.; Wang, S.; Shang, Y.C.; Maiese, K. Targeting cardiovascular disease with novel SIRT1 pathways. Future Cardiol. 2012, 8, 89–100. [Google Scholar] [CrossRef] [Green Version]
- Shen, J.; Han, X.; Ren, H.; Han, X.; Sun, W.; Gu, Y.; Qiao, J.; Dong, Q. Levels of Histone H3 Acetylation in Peripheral Blood Mononuclear Cells of Acute Cerebral Infarction Patients. Available online: https://europepmc.org/article/med/25327859 (accessed on 24 July 2021).
- Costantino, S.; Camici, G.G.; Mohammed, S.A.; Volpe, M.; Lüscher, T.F.; Paneni, F. Epigenetics and cardiovascular regenerative medicine in the elderly. Int. J. Cardiol. 2018, 250, 207–214. [Google Scholar] [CrossRef]
- Jaffe, I.Z.; Mendelsohn, M.E. Angiotensin II and aldosterone regulate gene transcription via functional mineralocortocoid receptors in human coronary artery smooth muscle cells. Circ. Res. 2005, 96, 643–650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Recchioni, R.; Marcheselli, F.; Antonicelli, R.; Mensà, E.; Lazzarini, R.; Procopio, A.D.; Olivieri, F. Epigenetic effects of physical activity in elderly patients with cardiovascular disease. Exp. Gerontol. 2017, 100, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Ling, C.; Rönn, T. Epigenetics in Human Obesity and Type 2 Diabetes. Cell Metab. 2019, 29, 1028–1044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loh, M.; Zhou, L.; Ng, H.K.; Chambers, J.C. Epigenetic disturbances in obesity and diabetes: Epidemiological and functional insights. Mol. Metab. 2019, 27, S33–S41. [Google Scholar] [CrossRef]
- Evans, L.W.; Stratton, M.S.; Ferguson, B.S. Dietary natural products as epigenetic modifiers in aging-associated inflammation and disease. Nat. Prod. Rep. 2020, 37, 653–676. [Google Scholar] [CrossRef] [PubMed]
- Alegría-Torres, J.A.; Baccarelli, A.; Bollati, V. Epigenetics and lifestyle. Epigenomics 2011, 3, 267–277. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, A.P.S.; Rosa, L.P.S.; Silveira, E.A. PPARG2 Pro12Ala polymorphism influences body composition changes in severely obese patients consuming extra virgin olive oil: A randomized clinical trial. Nutr. Metab. 2018, 15, 1–13. [Google Scholar] [CrossRef]
- Amenyah, S.D.; Ward, M.; Lees-murdock, D.J.; Strain, J.J.; Mcnulty, H.; Hughes, C.F.; Dollin, C.; Walsh, C.P. Nutritional Epigenomics and Age-Related Disease. Curr. Dev. Nutr. 2020, 4, nzaa097. [Google Scholar] [CrossRef]
- Rosanoff, A.; Weaver, C.M.; Rude, R.K. Suboptimal magnesium status in the United States: Are the health consequences underestimated? Nutr. Rev. 2012, 70, 153–164. [Google Scholar] [CrossRef]
- Silva-Martínez, G.A.; Rodríguez-Ríos, D.; Alvarado-Caudillo, Y.; Vaquero, A.; Esteller, M.; Carmona, F.J.; Moran, S.; Nielsen, F.C.; Wickström-Lindholm, M.; Wrobel, K.; et al. Arachidonic and oleic acid exert distinct effects on the DNA methylome. Epigenetics 2016, 11, 321–334. [Google Scholar] [CrossRef] [Green Version]
- González-Becerra, K.; Ramos-Lopez, O.; Barrón-Cabrera, E.; Riezu-Boj, J.I.; Milagro, F.I.; Martínez-López, E.; Martínez, J.A. Fatty acids, epigenetic mechanisms and chronic diseases: A systematic review. Lipids Health Dis. 2019, 18, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Zhao, F.; Zhang, J.; Chang, N. Epigenetic modification of Nrf2 by sulforaphane increases the antioxidative and anti-inflammatory capacity in a cellular model of Alzheimer’s disease. Eur. J. Pharmacol. 2018, 824, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Teiten, M.H.; Dicato, M.; Diederich, M. Curcumin as a regulator of epigenetic events. Mol. Nutr. Food Res. 2013, 57, 1619–1629. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Y.; Tang, D.; Du, Y.L.; Cao, Y.; Nie, Y.Q.; Cao, J.; Zhou, Y.J. Fatty liver mediated by PPAR-α DNA methylation can be reversed by a methylation inhibitor and curcumin. J. Dig. Dis. 2018, 19, 21–430. [Google Scholar] [CrossRef]
- Sasidharan Nair, V.; Song, M.H.; Oh, K.I. Vitamin C Facilitates Demethylation of the Foxp3 Enhancer in a Tet-Dependent Manner. J. Immunol. 2016, 196, 2119–2131. [Google Scholar] [CrossRef] [Green Version]
- Esteban, M.A.; Wang, T.; Qin, B.; Yang, J.; Qin, D.; Cai, J.; Li, W.; Weng, Z.; Chen, J.; Ni, S.; et al. Vitamin C Enhances the Generation of Mouse and Human Induced Pluripotent Stem Cells. Cell Stem Cell 2010, 6, 71–79. [Google Scholar] [CrossRef] [Green Version]
- Camarena, V.; Wang, G. The epigenetic role of vitamin C in health and disease. Cell. Mol. Life Sci. 2016, 73, 1645–1658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González, S.; Huerta, J.M.; Álvarez-Uría, J.; Fernández, S.; Patterson, Á.M.; Lasheras, C. Serum selenium is associated with plasma homocysteine concentrations in elderly humans. J. Nutr. 2004, 134, 1736–1740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.; Dong, Y.L.; Li, T.; Xiong, W.; Zhang, X.; Wang, P.J.; Huang, J.Q. Dietary Selenium Regulates microRNAs in Metabolic Disease: Recent Progress. Nutrients 2021, 13, 1527. [Google Scholar] [CrossRef]
- Speckmann, B.; Grune, T. Epigenetic effects of selenium and their implications for health. Epigenetics 2015, 10, 179–190. [Google Scholar] [CrossRef]
- Fiorito, G.; Guarrera, S.; Valle, C.; Ricceri, F.; Russo, A.; Grioni, S.; Mattiello, A.; Di Gaetano, C.; Rosa, F.; Modica, F.; et al. B-vitamins intake, DNA-methylation of One Carbon Metabolism and homocysteine pathway genes and myocardial infarction risk: The EPICOR study. Nutr. Metab. Cardiovasc. Dis. 2014, 24, 483–488. [Google Scholar] [CrossRef]
- Won, J.L.; Shim, J.Y.; Zhu, B.T. Mechanisms for the inhibition of DNA methyltransferases by tea catechins and bioflavonoids. Mol. Pharmacol. 2005, 68, 1018–1030. [Google Scholar]
- Grazioli, E.; Dimauro, I.; Mercatelli, N.; Wang, G.; Pitsiladis, Y.; Di Luigi, L.; Caporossi, D. Physical activity in the prevention of human diseases: Role of epigenetic modifications. BMC Genom. 2017, 18 (Suppl. 8), 111–123. [Google Scholar] [CrossRef] [Green Version]
- Xie, Z.; Rahman, I.; Goniewicz, M.L.; Li, D. Perspectives on Epigenetics Alterations Associated with Smoking and Vaping. Function 2021, 2, 1–6. [Google Scholar] [CrossRef]
- Ciafrè, S.; Carito, V.; Ferraguti, G.; Greco, A.; Chaldakov, G.N.; Fiore, M.; Ceccanti, M. How alcohol drinking affects our genes: An epigenetic point of view. Biochem. Cell Biol. 2019, 97, 345–356. [Google Scholar] [CrossRef] [Green Version]
- Brody, G.H.; Yu, T.; Chen, E.; Beach, S.R.H.; Miller, G.E. Family-centered prevention ameliorates the longitudinal association between risky family processes and epigenetic aging. J. Child. Psychol. Psychiatry Allied Discip. 2016, 57, 566–574. [Google Scholar] [CrossRef] [Green Version]
- Boks, M.P.; van Mierlo, H.C.; Rutten, B.P.; Radstake, T.R.; De Witte, L.; Geuze, E.; Horvath, S.; Schalkwyk, L.C.; Vinkers, C.H.; Broen, J.C.; et al. Longitudinal changes of telomere length and epigenetic age related to traumatic stress and post-traumatic stress disorder. Psychoneuroendocrinology 2015, 51, 506–512. [Google Scholar] [CrossRef] [Green Version]
- Lahtinen, A.; Puttonen, S.; Vanttola, P.; Viitasalo, K.; Sulkava, S.; Pervjakova, N.; Joensuu, A.; Salo, P.; Toivola, A.; Härmä, M.; et al. A distinctive DNA methylation pattern in insufficient sleep. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Reichard, J.F.; Puga, A. Effects of arsenic exposure on DNA methylation and epigenetic gene regulation. Epigenomics 2010, 2, 87–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chahine, T.; Baccarelli, A.; Litonjua, A.; Wright, R.O.; Suh, H.; Gold, D.R.; Sparrow, D.; Vokonas, P.; Schwartz, J. Particulate air pollution, oxidative stress genes, and heart rate variability in an elderly cohort. Environ. Health Perspect. 2007, 115, 1617–1622. [Google Scholar] [CrossRef]
- Mordukhovich, I.; Coull, B.; Kloog, I.; Koutrakis, P.; Vokonas, P.; Schwartz, J. Exposure to sub-chronic and long-term particulate air pollution and heart rate variability in an elderly cohort: The Normative Aging Study. Environ. Health 2015, 14, 87. [Google Scholar] [CrossRef] [Green Version]
- Castro, R.; Rivera, I.; Struys, E.A.; Jansen, E.E.W.; Ravasco, P.; Camilo, M.E.; Blom, H.J.; Jakobs, C.; De Almeida, I.T. Increased homocysteine and S-adenosylhomocysteine concentrations and DNA hypomethylation in vascular disease. Clin. Chem. 2003, 49, 1292–1296. [Google Scholar] [CrossRef] [PubMed]
- Bollati, V.; Marinelli, B.; Apostoli, P.; Bonzini, M.; Nordio, F.; Hoxha, M.; Pegoraro, V.; Motta, V.; Tarantini, L.; Cantone, L.; et al. Exposure to metal-rich particulate matter modifies the expression of candidate MicroRNAs in peripheral blood leukocytes. Environ. Health Perspect. 2010, 118, 763–768. [Google Scholar] [CrossRef] [Green Version]
- Duca, R.C.; Grova, N.; Ghosh, M.; Do, J.M.; Hoet, P.H.M.; Vanoirbeek, J.A.J.; Appenzeller, B.M.R.; Godderis, L. Exposure to Polycyclic Aromatic Hydrocarbons Leads to Non-monotonic Modulation of DNA and RNA (hydroxy)methylation in a Rat Model. Sci. Rep. 2018, 8, 1–9. [Google Scholar]
- White, A.J.; Kresovich, J.K.; Xu, Z.; Sandler, D.P.; Taylor, J.A. Shift work, DNA methylation and epigenetic age. Int. J. Epidemiol. 2019, 48, 1536–1544. [Google Scholar] [CrossRef] [PubMed]
- Bhatti, P.; Zhang, Y.; Song, X.; Makar, K.W.; Sather, C.L.; Kelsey, K.T.; Houseman, E.A.; Wang, P. Nightshift work and genome-wide DNA methylation. Chronobiol. Int. 2015, 32, 103–112. [Google Scholar] [CrossRef]
- Bartsch, H.; Nair, J. Oxidative stress and lipid peroxidation-derived DNA-lesions in inflammation driven carcinogenesis. Cancer Detect. Prev. 2004, 28, 385–391. [Google Scholar] [CrossRef]
- Kieć-Wilk, B.; Razny, U.; Mathers, J.; Dembińska-Kieć, A. DNA Methylation, Induced by Beta-Carotene and Arachidonic Acid, Plays a Regulatory Role in the Pro-Angiogenic VEGF-Receptor (KDR) Gene Expression in Endothelial Cells. Available online: http://www.jpp.krakow.pl/journal/archive/12_09/pdf/49_12_09_article.pdf (accessed on 30 July 2021).
- Dashwood, R.; Ho, E. Dietary Histone Deacetylase Inhibitors: From Cells to Mice to Man; Elsevier: Amsterdam, The Netherlands, 2007; Available online: https://0-www-sciencedirect-com.brum.beds.ac.uk/science/article/pii/S1044579X07000247 (accessed on 30 July 2021).
- Arora, I.; Sharma, M.; Sun, L.Y.; Tollefsbol, T.O. The epigenetic link between polyphenols, aging and age-related diseases. Genes 2020, 11, 1094. [Google Scholar] [CrossRef]
- Zhang, F.F.; Cardarelli, R.; Carroll, J.; Zhang, S.; Fulda, K.G.; Gonzalez, K.; Vishwanatha, J.K.; Morabia, A.; Santella, R.M. Physical activity and global genomic DNA methylation in a cancer-free population. Epigenetics 2011, 6, 293–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuasa, Y.; Nagasaki, H.; Akiyama, Y.; Hashimoto, Y.; Takizawa, T.; Kojima, K.; Kawano, T.; Sugihara, K.; Imai, K.; Nakachi, K. DNA methylation status is inversely correlated with green tea intake and physical activity in gastric cancer patients. Int. J. Cancer 2009, 124, 2677–2682. [Google Scholar] [CrossRef] [PubMed]
- Andersen, A.; Gerrard, M.; Gibbons, F.X.; Beach, S.R.H.; Philibert, R. An Examination of Risk Factors for Tobacco and Cannabis Smoke Exposure in Adolescents Using an Epigenetic Biomarker. Front Psychiatry 2021, 24, 688384. [Google Scholar] [CrossRef]
- Liu, F.; Killian, J.K.; Yang, M.; Walker, R.L.; Hong, J.A.; Zhang, M.; Davis, S.; Zhang, Y.; Hussain, M.; Xi, S.; et al. Epigenomic alterations and gene expression profiles in respiratory epithelia exposed to cigarette smoke condensate. Oncogene 2010, 29, 3650–3664. [Google Scholar] [CrossRef]
- Toyooka, S.; Tokumo, M.; Shigematsu, H.; Matsuo, K.; Asano, H.; Gazdar, A.F.; Shimizu, N. Mutational and Epigenetic Evidence for Independent Pathways for Lung Adenocarcinomas Arising in Smokers and Never Smokers. Cancer Res. 2006, 66, 1371–1376. [Google Scholar] [CrossRef] [Green Version]
- Miller, C.A.; Sweatt, J.D. Covalent Modification of DNA Regulates Memory Formation. Neuron 2007, 53, 857–869. [Google Scholar] [CrossRef] [Green Version]
- McGowan, P.O.; Sasaki, A.; D’Alessio, A.C.; Dymov, S.; Labonté, B.; Szyf, M.; Turecki, G.; Meaney, M.J. Epigenetic regulation of the glucocorticoid receptor in human brain associates with childhood abuse. Nat. Neurosci. 2009, 12, 342–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murgatroyd, C.; Patchev, A.V.; Wu, Y.; Micale, V.; Bockmühl, Y.; Fischer, D.; Holsboer, F.; Wotjak, C.T.; Almeida, O.F.X.; Spengler, D. Dynamic DNA methylation programs persistent adverse effects of early-life stress. Nat. Neurosci. 2009, 12, 1559–1566. [Google Scholar] [CrossRef] [PubMed]
- de Moura, M.B.; Santana-Santos, L.; Van Houten, B. Mitochondrial dysfunction in neurodegenerative diseases and cancer. Environ. Mol. Mutagenesis 2010, 51, 391–405. [Google Scholar] [CrossRef]
- Skinner, M.K. Environmental epigenomics and disease susceptibility. EMBO Rep. 2011, 12, 620–622. [Google Scholar] [CrossRef] [Green Version]
- Miller, C.A.; Gavin, C.F.; White, J.A.; Parrish, R.R.; Honasoge, A.; Yancey, C.R.; Rivera, I.M.; Rubio, M.D.; Rumbaugh, G.; Sweatt, J.D. Cortical DNA Methylation Maintains Remote Memory. Available online: https://0-www-nature-com.brum.beds.ac.uk/articles/nn.2560 (accessed on 7 June 2021).
- Pilsner, J.R.; Liu, X.; Ahsan, H.; Ilievski, V.; Slavkovich, V.; Levy, D.; Factor-Litvak, P.; Graziano, J.H.; Gamble, M.V. Genomic methylation of peripheral blood leukocyte DNA: Influences of arsenic and folate in Bangladeshi adults. Am. J. Clin. Nutr. 2007, 86, 1179–1186. [Google Scholar] [CrossRef] [Green Version]
- Pilsner, J.R.; Liu, X.; Ahsan, H.; Ilievski, V.; Slavkovich, V.; Levy, D.; Factor-Litvak, P.; Graziano, J.H.; Gamble, M.V. Folate deficiency, hyperhomocysteinemia, low university creatinine, and hypomethylation of leukocyte DNA are risk factors for arsenic-induced skin lesions. Environ. Health Perspect. 2009, 117, 254–260. [Google Scholar] [CrossRef] [PubMed]
- Chanda, S.; Dasgupta, U.B.; GuhaMazumder, D.; Gupta, M.; Chaudhuri, U.; Lahiri, S.; Das, S.; Ghosh, N.; Chatterjee, D. DNA hypermethylation of promoter of gene p53 and p16 in arsenic-exposed people with and without malignancy. Toxicol. Sci. 2006, 89, 431–437. [Google Scholar] [CrossRef] [Green Version]
- Kunovac, A.; Hathaway, Q.A.; Pinti, M.V.; Taylor, A.D.; Hollander, J.M. Cardiovascular adaptations to particle inhalation exposure: Molecular mechanisms of the toxicology. Am. J. Physiol.-Heart Circ. Physiol. 2020, 319, H282–H305. [Google Scholar] [CrossRef] [PubMed]
- Costa, G. The Problem: Shiftwork. Chronobiol. Int. 1997, 14, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Bajpe, P.K.; Heynen, G.J.J.E.; Mittempergher, L.; Grernrum, W.; de Rink, I.A.; Nijkamp, W.; Beijersbergen, R.L.; Bernards, R.; Huang, S. The Corepressor CTBP2 Is a Coactivator of Retinoic Acid Receptor/Retinoid X Receptor in Retinoic Acid Signaling. Mol. Cell. Biol. 2013, 33, 3343–3353. [Google Scholar] [CrossRef] [Green Version]
- Rodenhiser, D.; Mann, M. Epigenetics and human disease: Translating basic biology into clinical applications. CMAJ 2006, 174, 341–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khalil, C.A. The emerging role of epigenetics in cardiovascular disease. Ther. Adv. Chronic Dis. 2014, 5, 178–187. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Tissue | miRNA | Molecular Targets | Functions | |
|---|---|---|---|---|
| Aging heart | miR-21 [68] | Increase | ERK-MAP kinase signaling | Profibrotic (role on cardiac fibroblast (CFS)) |
| Aging heart | miR-22 [69] | Increase | Mimecan/osteoglycin | Accelerate CF senescence and migration |
| Aging heart | miR-18 and miR-19 [70] | Decrease | Thrombospondin-1 and connective tissue growth factor | Anti-heart failure-related fibrosis during aging |
| Aging heart | miR-17-3P [71] | Decrease | PAR-4 | Reduce CF cellular senescence |
| Aging heart | miR-34a [72] | Increase | Phosphatase 1 nuclear targeting subunit | Increase age-related cardiomyocyte apoptosis and cardiac dysfunction |
| Vascular aging | miR-34a [73] | Increase | SIRT1 | Stimulate senescence in endothelial cells and vascular smooth muscle cells |
| Vascular aging | miR-217 [74] | Increase | SIRT1 | Stimulate endothelial senescence, decrease nitric oxide |
| Vascular aging | miR-29 [75] | Decrease | Collagen and elastin | Extracellular matrix impairment (risk of age-related aortic aneurism) |
| Vascular aging | miR-146 [76] | Increase | IRAK and NOX4 | Proinflammation or antioxidative stress |
| Vascular aging | miR-92 [77] | Decrease | TNF receptor 1 and collagen type1 | Reduction in mimic arterial aging |
| Vascular aging | mir-20a [78] | Decrease | MKK3, activation of p38 MAP kinase | Inhibit endothelial cell migration by the inhibition of MKK3 and the activation of p38 MAP kinase |
| Vascular aging | mir-126 [79] | Decrease | PLGF | Increase cell apoptosis, decrease proliferation, endothelial cell migration |
| Vascular aging | mir-10a [80] | Decrease | p53/Rb network, including p53 regulator MDM4, Rb regulator RB1CC1, p21 regulator TFAP2C, p53 | Endothelial progenitor cell dysfunction |
| Vascular aging | mir-21 [81] | Decrease | PTEN, SPRY1, SPRY2 | Regeneration of endothelial progenitor cells |
| Vascular aging | mir- 217 [82] | Increase | SIRT1, FOXO1, eNOS | Impair endothelial angiogenesis |
| Vascular aging | mir-146a [83] | Decrease | Toll-like receptor 4 (TLR4) | Senescent endothelial cells |
| Vascular aging | mir-17-92 [84] | Decrease | Chk1/2, G-H2AX, ATM | Regulators of chromatin-related proteins |
| Vascular aging | miR-204-3p [56] | Decrease | PDGF | Vascular smooth muscle cell growth upon injury |
| Epigenetic Modifications | Sites | Affected Gene |
|---|---|---|
| DNA methylation | KLK10, LIM, LMO, D1D, CD7, CD22, CD27, CD59 and CD82, IL1R2, IL2RA, IL19, IL21R, IL32, GPR21, GPR65, GPR81, GPR84, and GPR171 | CRP [99] |
| BAF155, Inil, c-Myc, BAF170, Max, NRSF, and Nrf1 | IL-6 [100] | |
| NLRC5 and DTX3L/PARP9, IFN-γ, and ABO | TNF-α [101] | |
| Histone modification | H3K4me3 | SIRT1, FoxO3, NF-κB, and p53 [102] TNF-α [103], SET1A/B, SET7, MLL1/2, MLL3/4, LL1, and VEGFA [104] |
| H3K4me3 and H3K9ac | TNF-α [103] | |
| H3K9me2 | VSMC [105] |
| Item | Epigenetic Changes | ||||
|---|---|---|---|---|---|
| DNA Methylation | Histone Modification | microRNA Involvement | DNMT Enzymes | ||
| Nutritional habits and food consumption | Polyunsaturated fatty acids [141] | + | |||
| Arachidonic acid and oleic acid [142] | + | ||||
| Diets rich in fruits and vegetables—sulforaphane— for example, broccoli, cauliflower, cabbage, kale [143] | + | + | |||
| Folate and vitamin B12 intake [121] | + | + | + | + | |
| Polyphenols (green tea, tea, and coffee are a rich source of polyphenols) [144] | + | - | |||
| Magnesium and selenium [122] | + | + | |||
| Curcumin | + | ||||
| Ascorbic acid | + | ||||
| Physical activity [145,146] | + | + | + | ||
| Tobacco smoke [147,148,149] | + | + | + | ||
| Alcohol consumption [110] | + | ||||
| Psychological stress and sleep deficiency [132,150,151,152,153,154,155] | + | ||||
| Environmental pollutants | Arsenic [156,157,158] | + | |||
| Air pollution [159] | + | + | |||
| Aromatic hydrocarbons and other organic pollutants | + | ||||
| Shift work [160] | + | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gharipour, M.; Mani, A.; Amini Baghbahadorani, M.; de Souza Cardoso, C.K.; Jahanfar, S.; Sarrafzadegan, N.; de Oliveira, C.; Silveira, E.A. How Are Epigenetic Modifications Related to Cardiovascular Disease in Older Adults? Int. J. Mol. Sci. 2021, 22, 9949. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22189949
Gharipour M, Mani A, Amini Baghbahadorani M, de Souza Cardoso CK, Jahanfar S, Sarrafzadegan N, de Oliveira C, Silveira EA. How Are Epigenetic Modifications Related to Cardiovascular Disease in Older Adults? International Journal of Molecular Sciences. 2021; 22(18):9949. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22189949
Chicago/Turabian StyleGharipour, Mojgan, Arya Mani, Mona Amini Baghbahadorani, Camila Kellen de Souza Cardoso, Shayesteh Jahanfar, Nizal Sarrafzadegan, Cesar de Oliveira, and Erika Aparecida Silveira. 2021. "How Are Epigenetic Modifications Related to Cardiovascular Disease in Older Adults?" International Journal of Molecular Sciences 22, no. 18: 9949. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22189949