
Improvement Effects of Myelophil on Symptoms of Chronic Fatigue Syndrome in a Reserpine-Induced Mouse Model



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
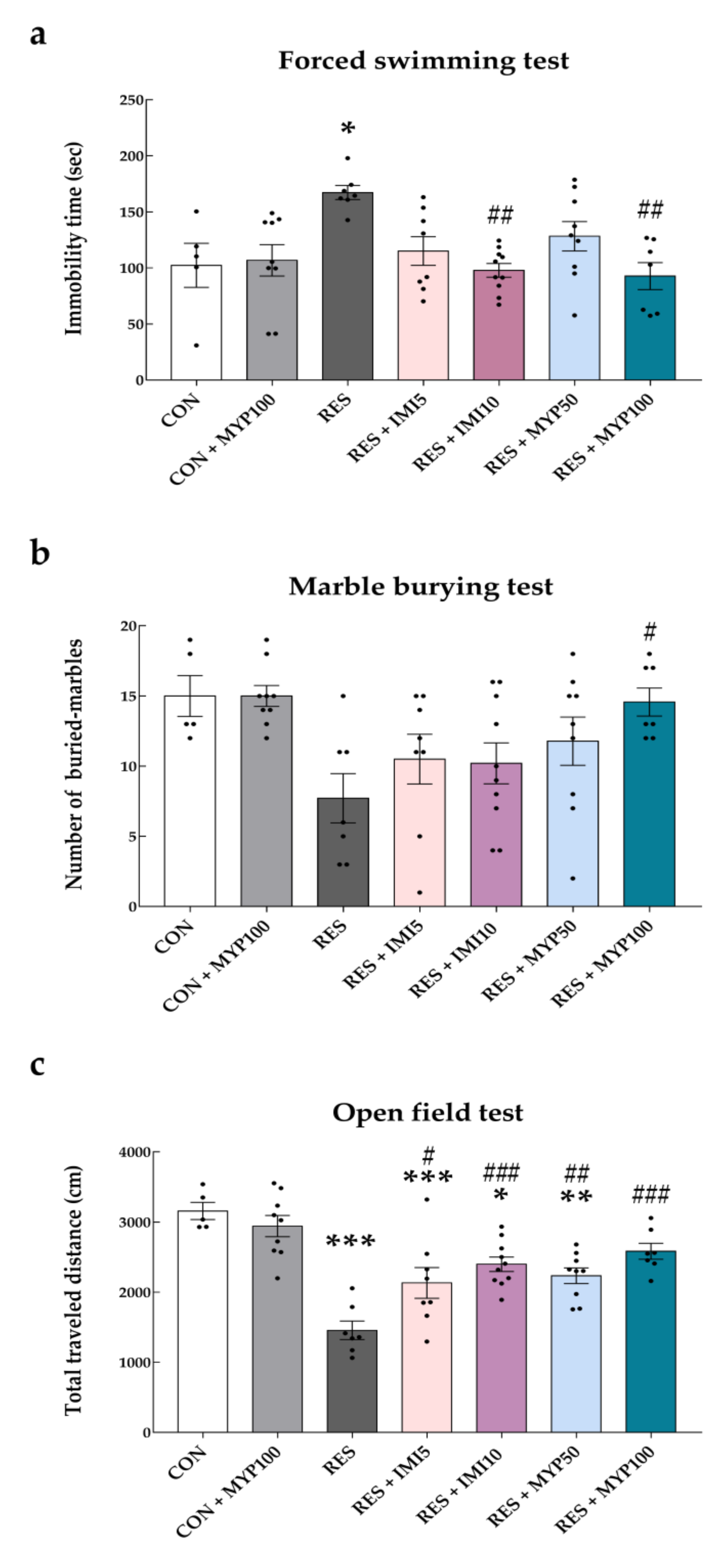
2.1. Improvement Effect of MYP Treatment on Depression-Like Behaviors
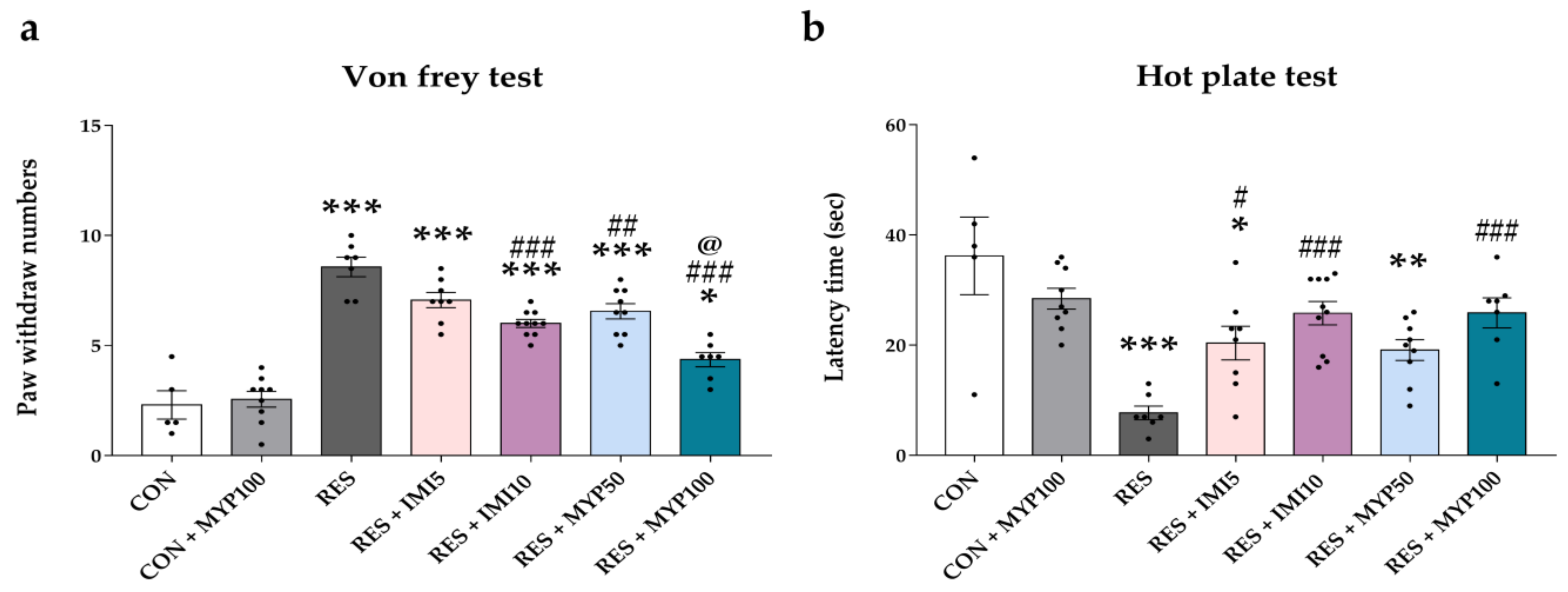
2.2. Alteration of Mechanical and Thermal Hypersensitivity by MYP Treatment
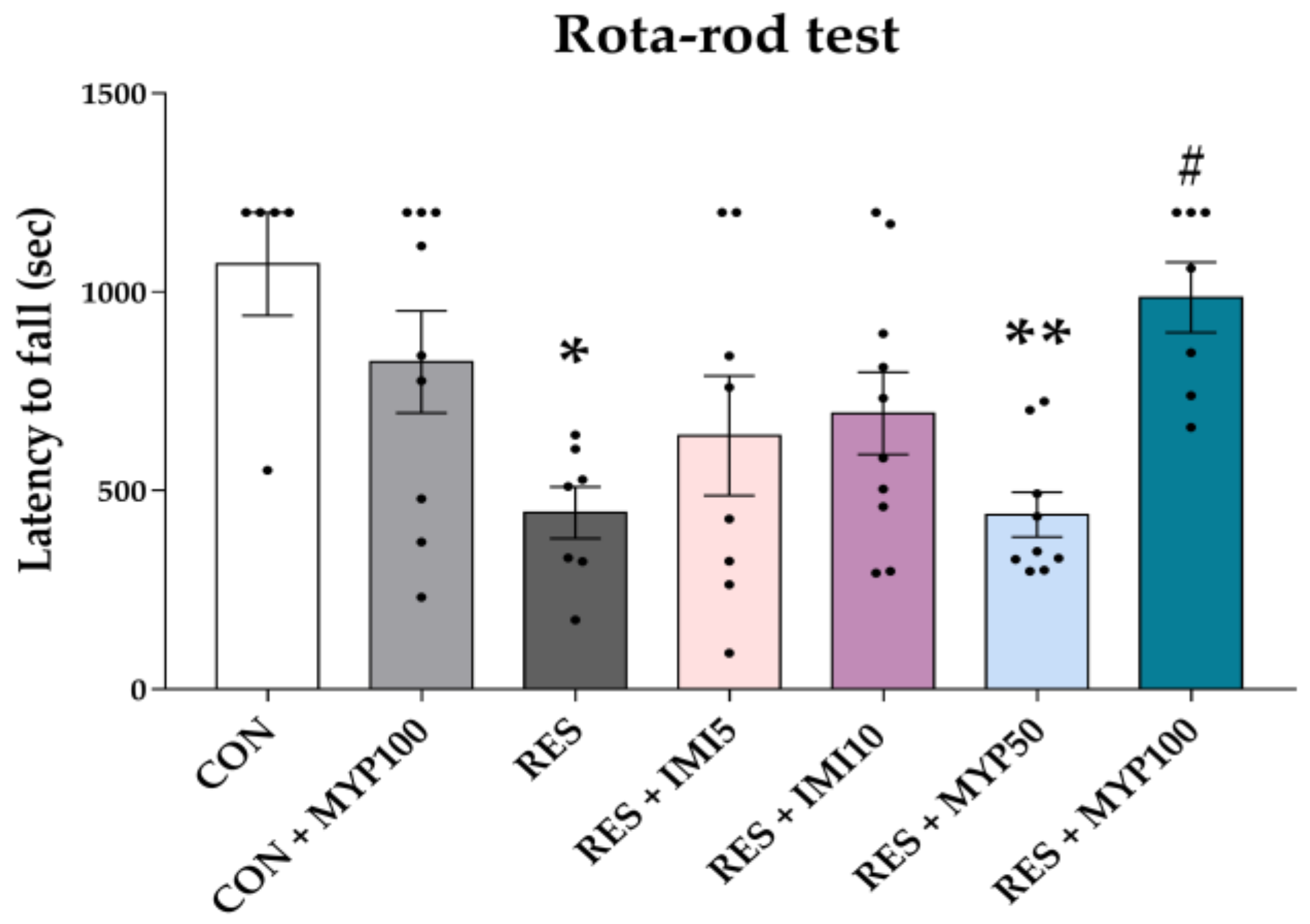
2.3. Improvement Effect of MYP Treatment on Fatigue Behavior
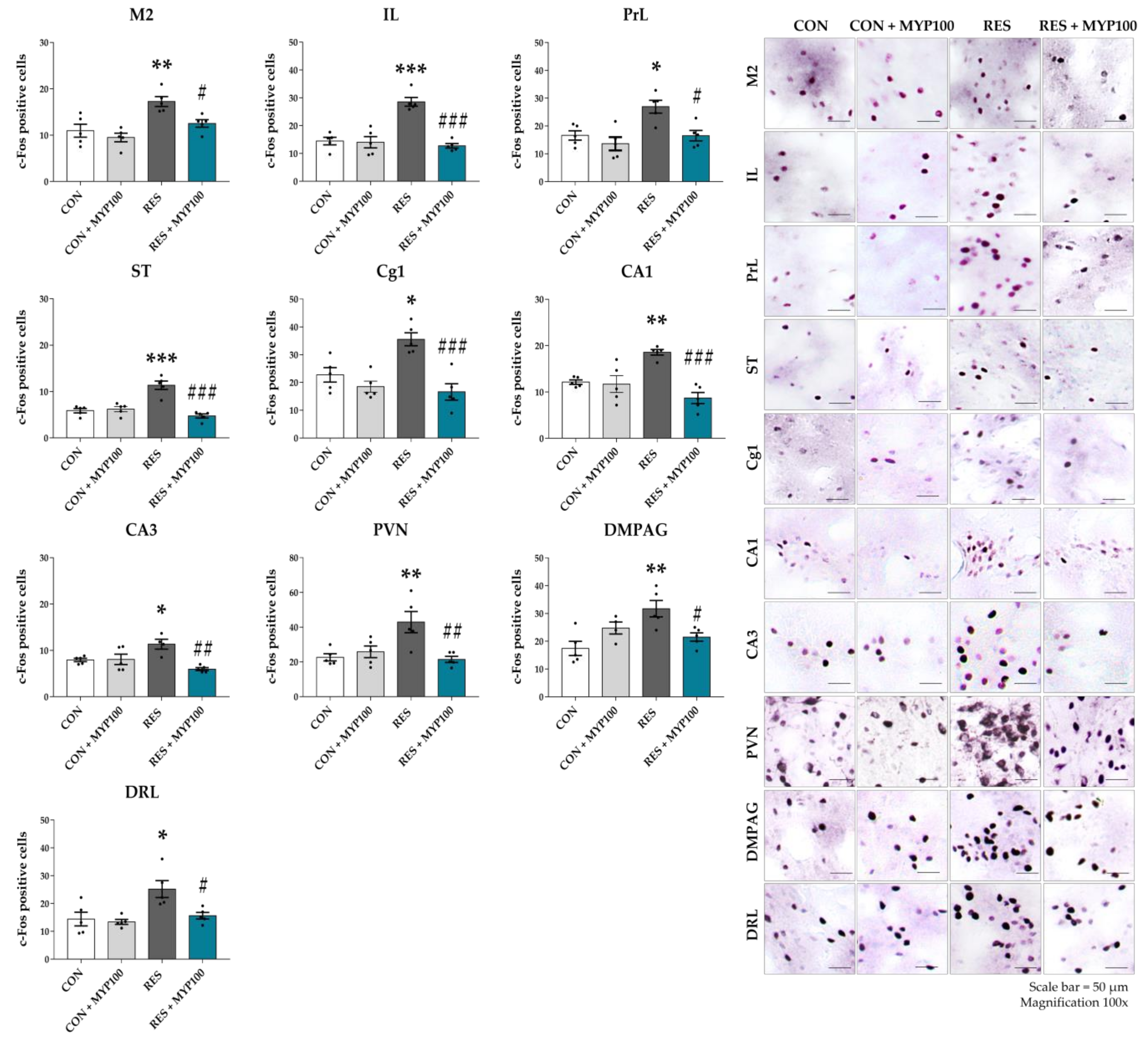
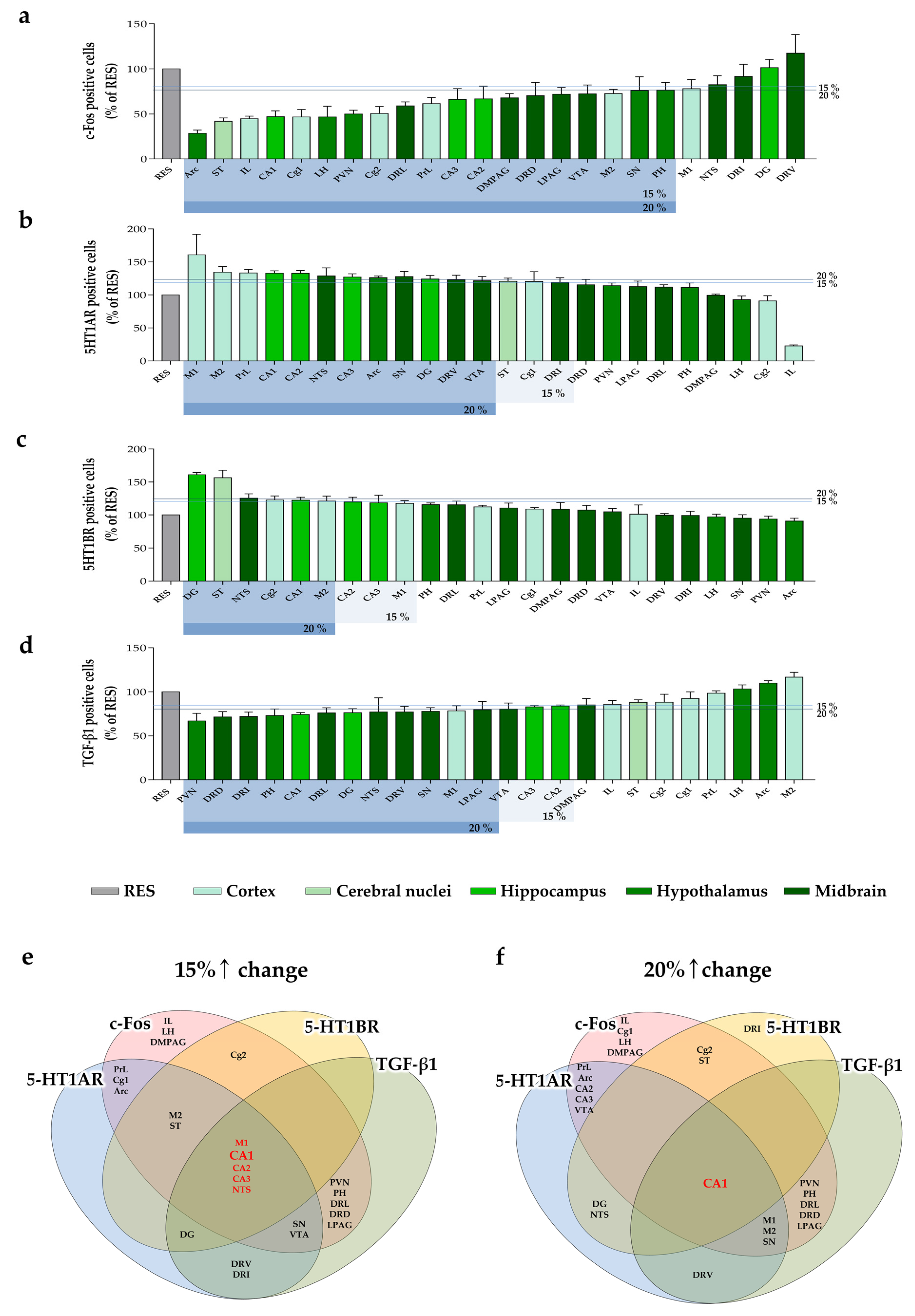
2.4. Changes of c-Fos Expression by MYP Treatment in the Brain
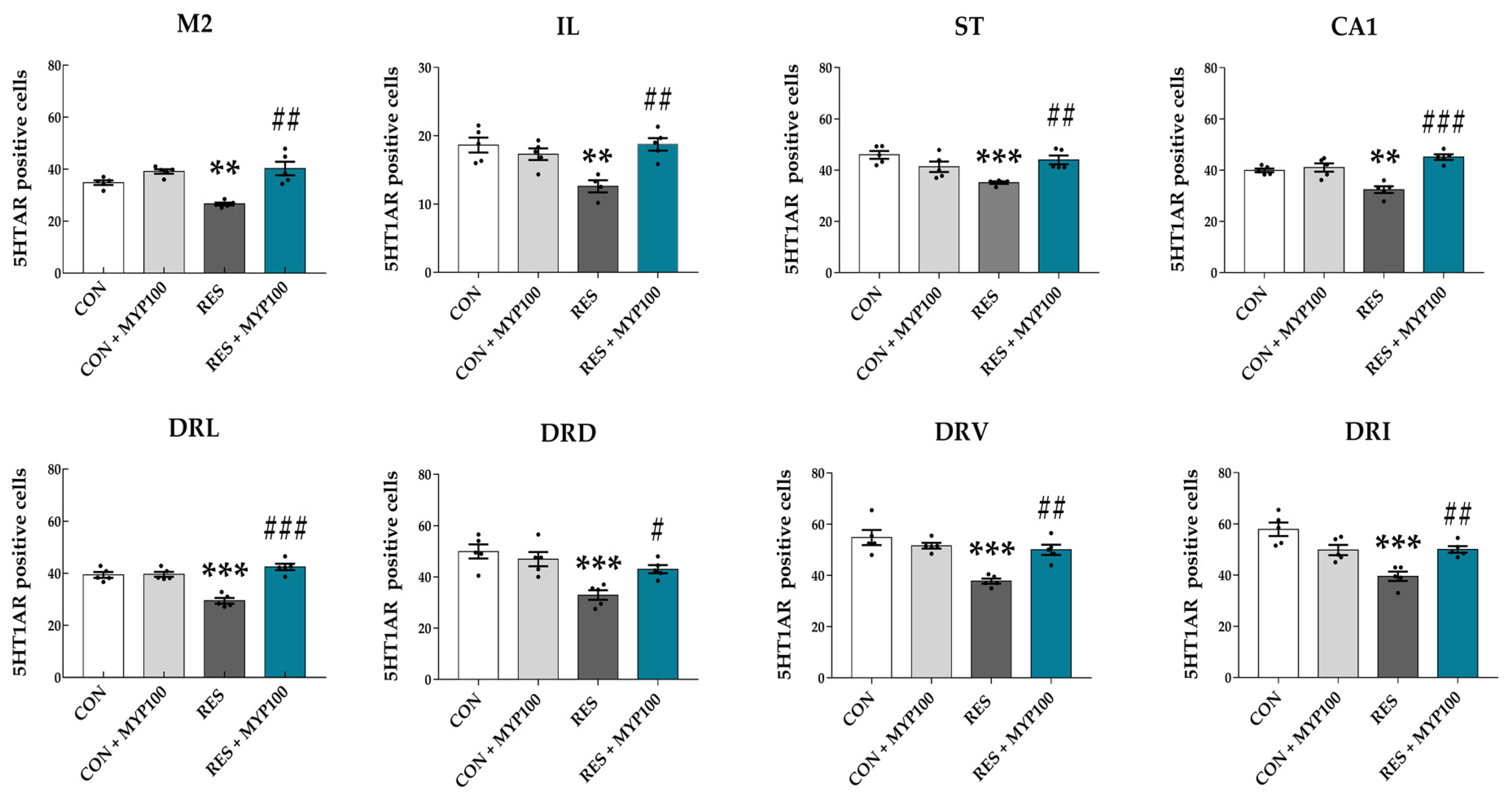
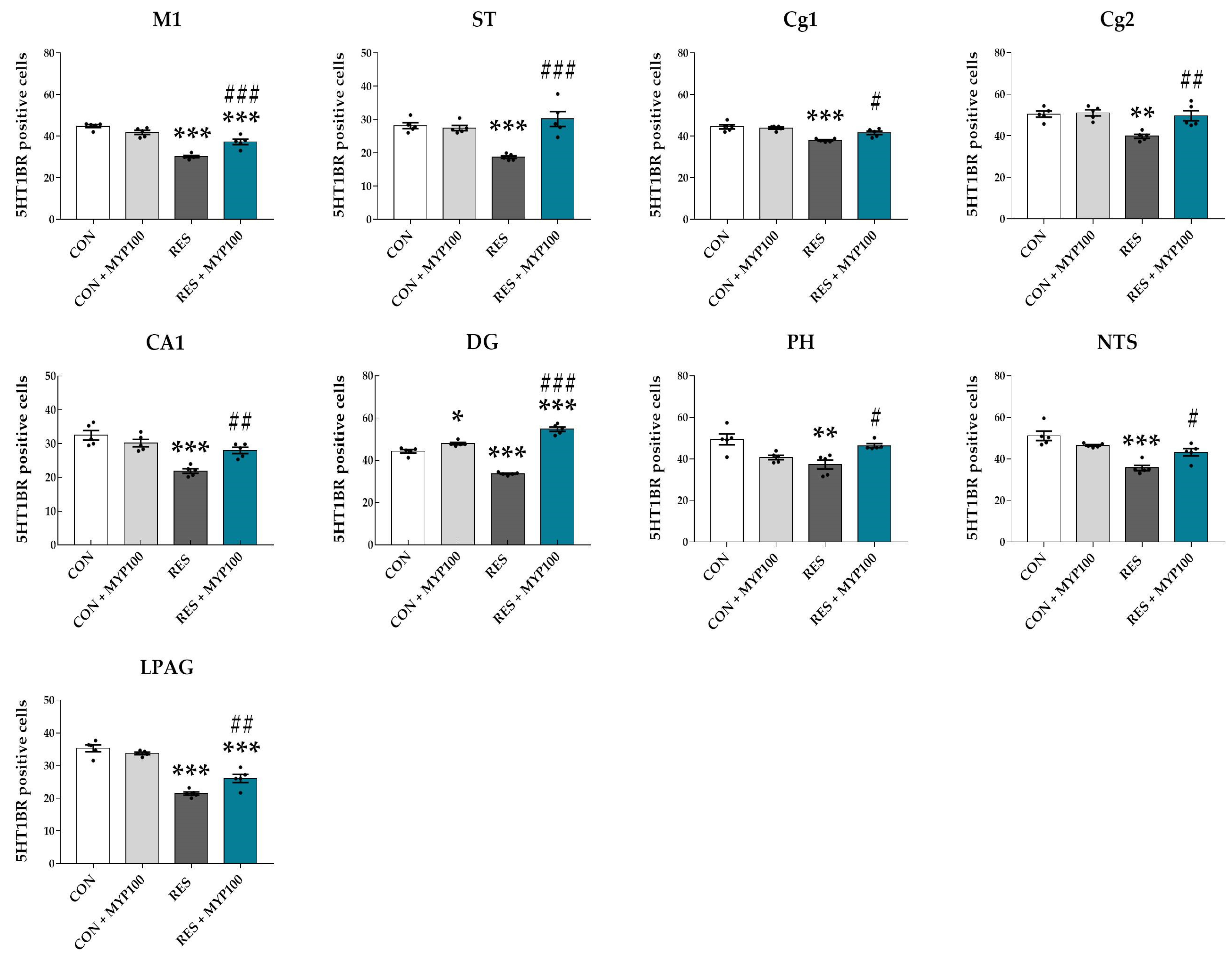
2.5. Regulation of 5-HT1A/B Receptors Expression by MYP Treatment in the Brain
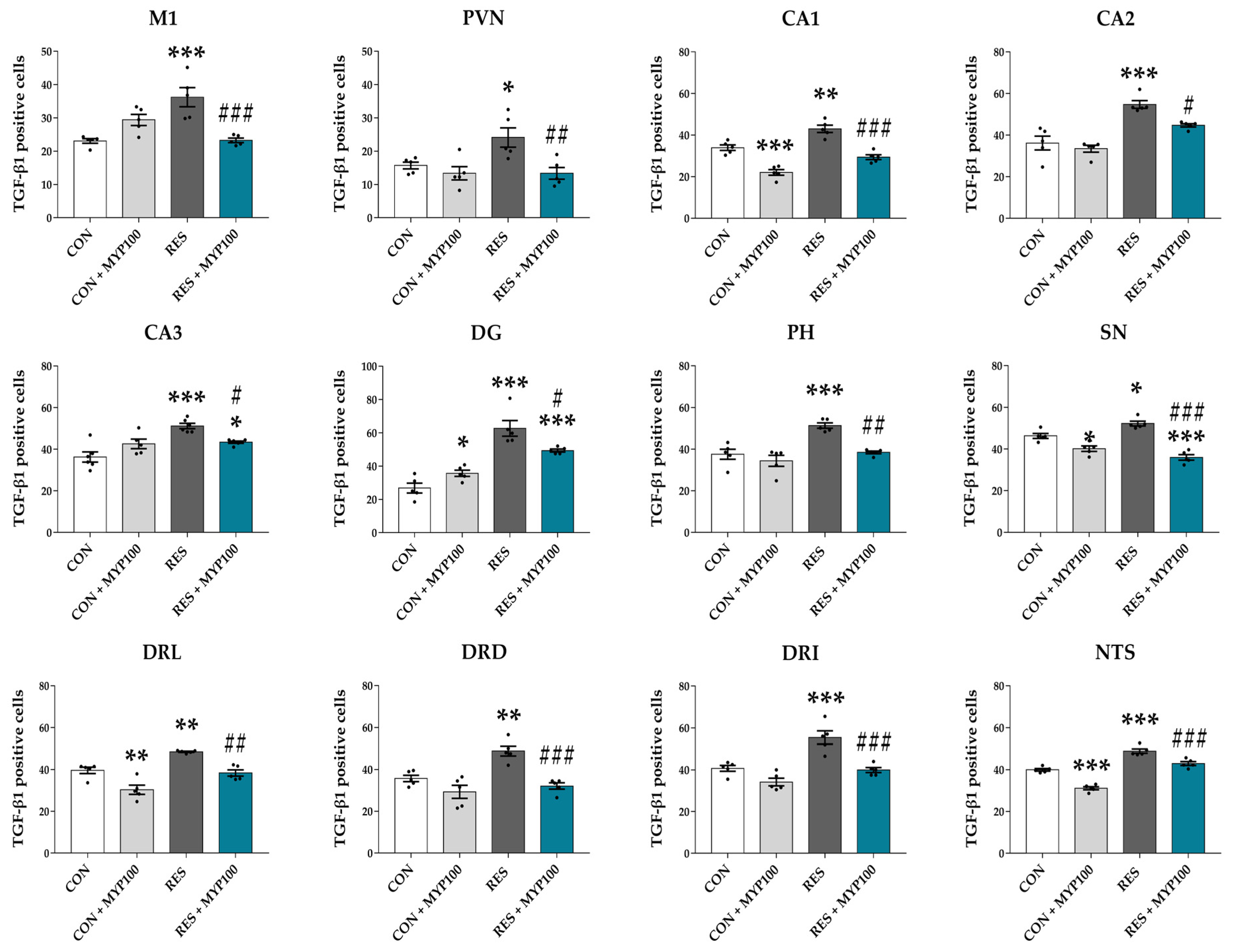
2.6. Reduction of TGF-β1 Expression by MYP Treatment in the Brain
2.7. Analyze Hub Brain Regions According to the Changes of c-Fos, 5-HT1A/B Receptors, and TGF-β1 Expression by MYP Treatment
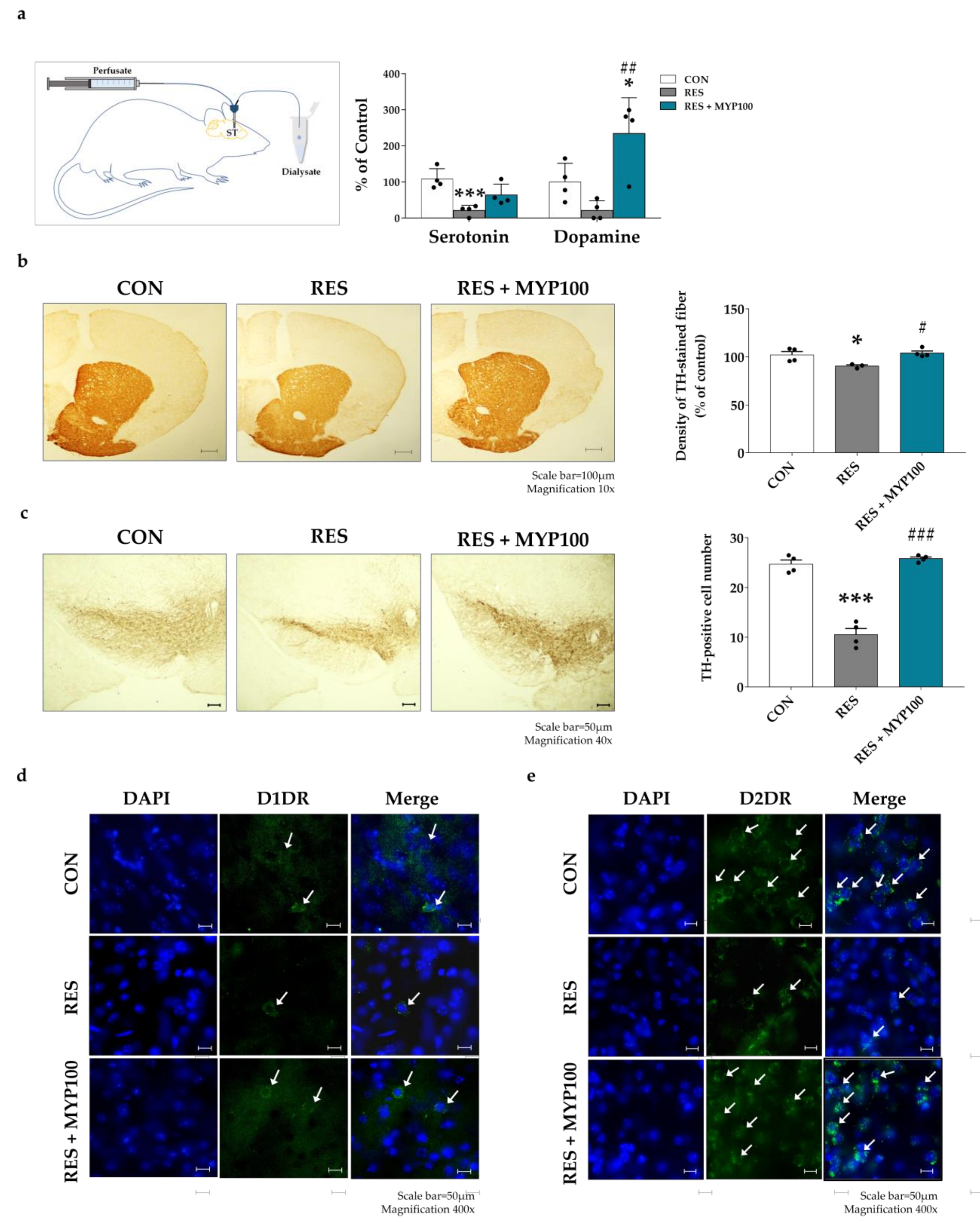
2.8. Reduction of TGF-β1 and Iba1 Expression by MYP Treatment in the Hippocampus
2.9. Effect of MYP Treatment on Dopamine and 5-HT Production in ST
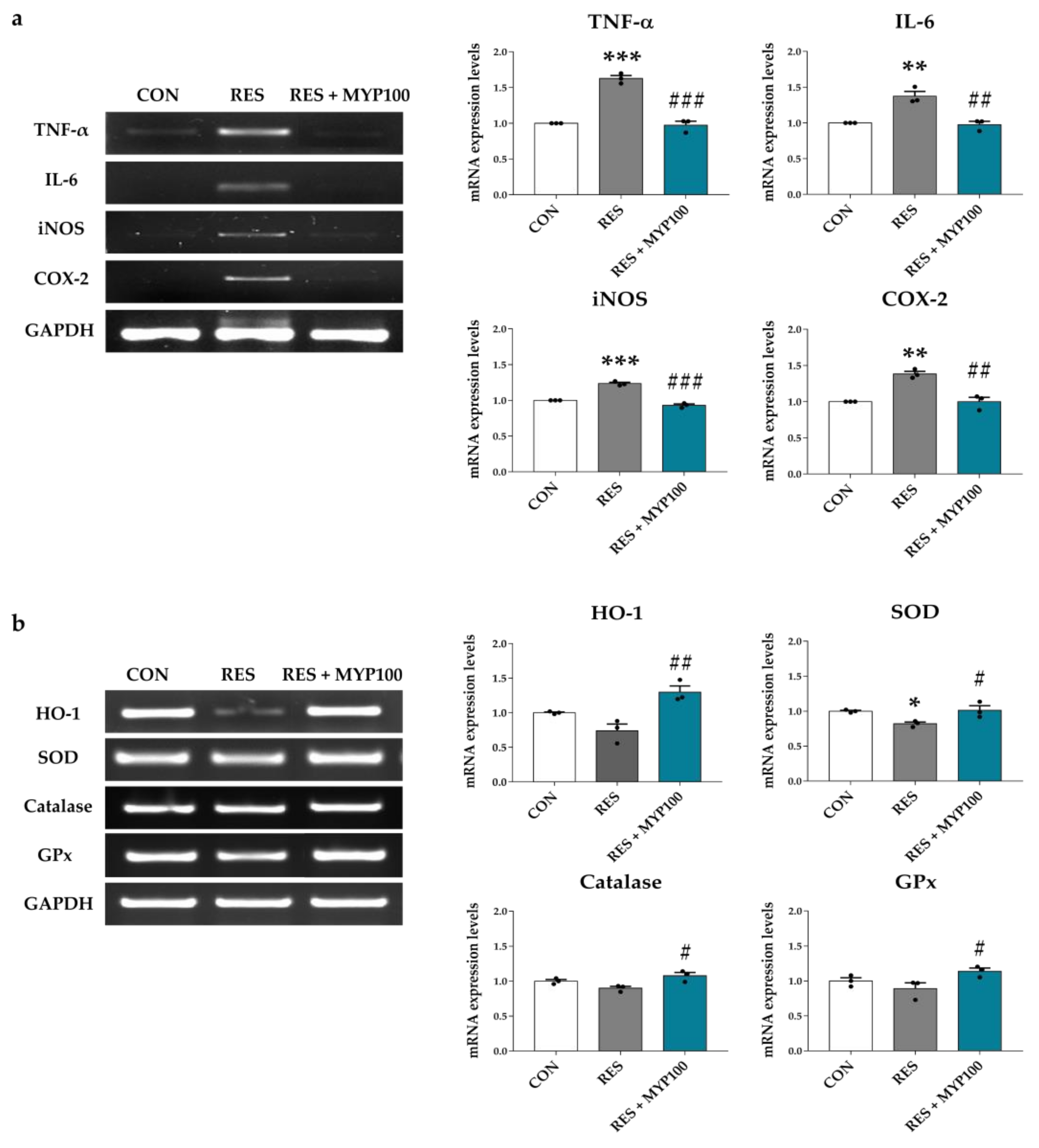
2.10. Anti-Oxidative and Anti-Inflammation Effects of MYP in Internal Organs
3. Discussion
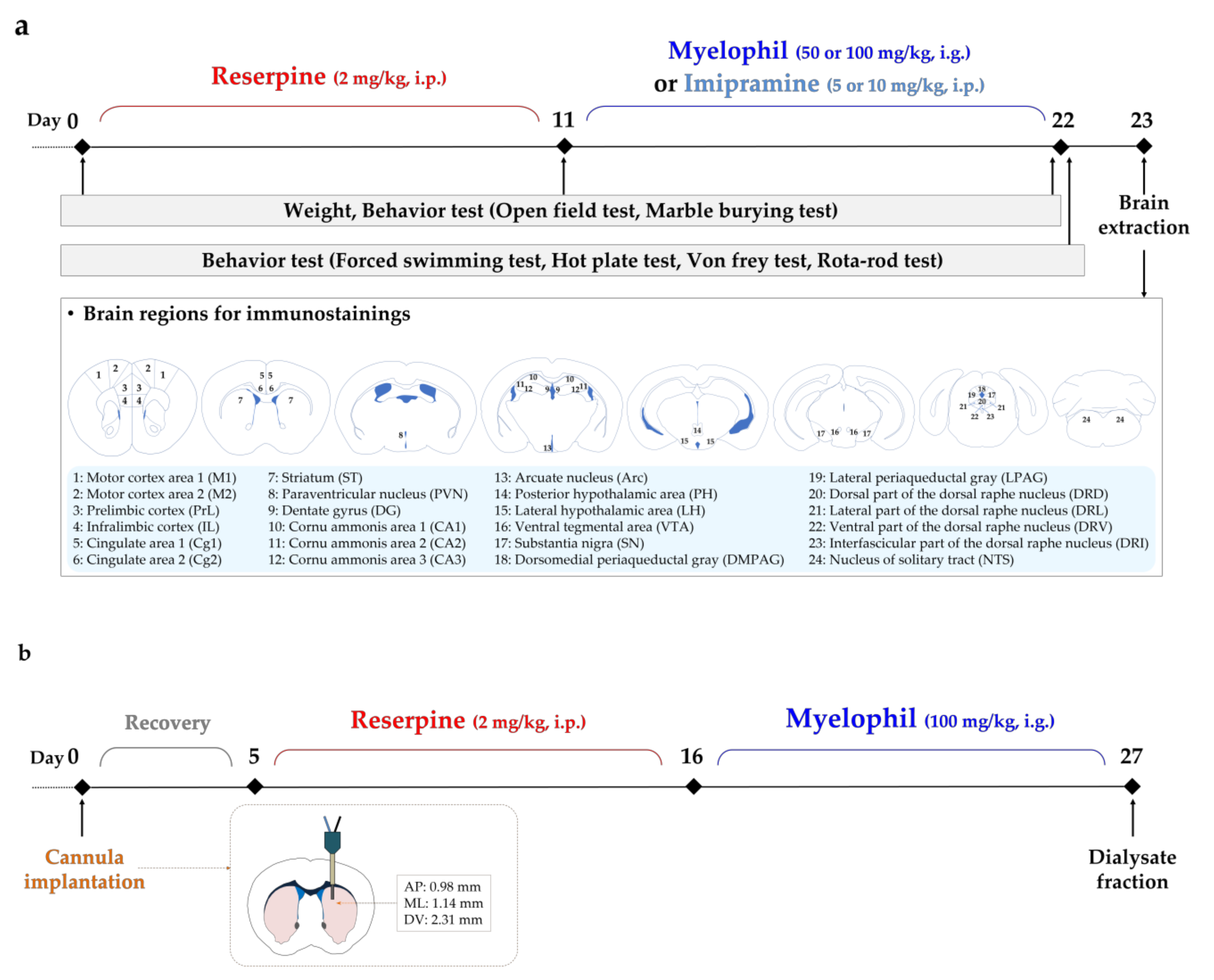
4. Materials and Methods
4.1. Animals
4.2. Reserpine Injection and MYP Administration
4.3. Behavioral Test
4.3.1. OFT
4.3.2. MBT
4.3.3. FST
4.3.4. Von Frey Test
4.3.5. Hot Plate Test
4.3.6. Rota-Rod Test
4.4. Immunostainings
4.5. Brain Microdialysis
4.6. Liquid Chromathography (LC)-Tandem Mass Spectrometer (MS/MS) Analysis
4.7. Reverse Transcription-Polymerase Chain Reaction (RT-PCR)
4.8. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Boulazreg, S.; Rokach, A. The Lonely, Isolating, and Alienating Implications of Myalgic Encephalomyelitis/Chronic Fatigue Syndrome. Healthcare 2020, 8, 413. [Google Scholar] [CrossRef]
- Fukuda, K.; Stephen, E.; Ian, H.; Michael, C.S.; James, G.D. The chronic fatigue syndrome: A comprehensive approach to its definition and study. International Chronic Fatigue Syndrome Study Group. Ann. Intern. Med. 1994, 121, 953–959. [Google Scholar] [CrossRef] [PubMed]
- Abbi, B.; Natelson, B.H. Is chronic fatigue syndrome the same illness as fibromyalgia: Evaluating the ′single syndrome′ hypothesis. QJM 2013, 106, 3–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maes, M.; Ringel, K.; Kubera, M.; Anderson, G.; Morris, G.; Galecki, P.; Geffard, M. In myalgic encephalomyelitis/chronic fatigue syndrome, increased autoimmune activity against 5-HT is associated with immuno-inflammatory pathways and bacterial translocation. J. Affect Disord. 2013, 150, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Corbitt, M.; Eaton-Fitch, N.; Staines, D.; Cabanas, H.; Marshall-Gradisnik, S. A systematic review of cytokines in chronic fatigue syndrome/myalgic encephalomyelitis/systemic exertion intolerance disease (CFS/ME/SEID). BMC Neurol. 2019, 19, 207. [Google Scholar] [CrossRef]
- Lee, J.S.; Jeon, Y.-J.; Park, S.-Y.; Son, C.-G. An Adrenalectomy Mouse Model Reflecting Clinical Features for Chronic Fatigue Syndrome. Biomolecules 2020, 10, 71. [Google Scholar] [CrossRef] [Green Version]
- Ohba, T.; Domoto, S.; Tanak, M.; Nakamura, S.; Shimazawa, M.; Hara, H. Myalgic Encephalomyelitis/Chronic Fatigue Syndrome Induced by Repeated Forced Swimming in Mice. Biol. Pharm. Bull. 2019, 42, 1140–1145. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Hu, Y.; Liu, P.; Zhao, H.-X.; Zhou, X.-J.; Wei, Y.-M. Effects of a Chinese traditional formula Kai Xin San (KXS) on chronic fatigue syndrome mice induced by forced wheel running. J. Ethnopharmacol. 2012, 139, 19–25. [Google Scholar] [CrossRef]
- Gupta, A.; Vij, G.; Chopra, K. Possible role of oxidative stress and immunological activation in mouse model of chronic fatigue syndrome and its attenuation by olive extract. J. Neuroimmunol. 2010, 226, 3–7. [Google Scholar] [CrossRef]
- Yaffe, D.; Forrest, L.R.; Schuldiner, S. The ins and outs of vesicular monoamine transporters. J. Gen. Physiol. 2018, 150, 671–682. [Google Scholar] [CrossRef] [Green Version]
- Fujimiya, M.; Nakazawa, M.; Maeda, T.; Kimura, H.; Okumiya, K.; Kitahama, K. Effect of reserpine on 5-hydroxytryptophan (5HTP)-immunoreactive neurons in the rat brain. Histochemistry 1994, 101, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Eiden, L.E.; Weihe, E. VMAT2: A dynamic regulator of brain monoaminergic neuronal function interacting with drugs of abuse. Ann. N. Y. Acad. Sci. 2011, 1216, 86–98. [Google Scholar] [CrossRef] [Green Version]
- Hao, Y.; Ge, H.; Sun, M.; Gao, Y. Selecting an Appropriate Animal Model of Depression. Int. J. Mol. Sci. 2019, 20, 4827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brum, E.D.S.; Fialho, M.F.P.; Fischer, S.P.M.; Hartman, D.D.; Goncalves, D.F.; Scussel, R.; Machado-de-Avila, R.A.; Corte, C.L.D.; Soares, F.A.A.; Oliviera, S.M. Relevance of Mitochondrial Dysfunction in the Reserpine-Induced Experimental Fibromyalgia Model. Mol. Neurobiol. 2020, 57, 4202–4217. [Google Scholar] [CrossRef]
- Lee, J.S.; Kim, H.-G.; Han, J.-M.; Lee, J.-S.; Son, S.-W.; Ahn, Y.-C.; Son, C.-G. Myelophil ameliorates brain oxidative stress in mice subjected to restraint stress. Prog. Neuropsychopharmacol. Biol. Psychiatry 2012, 39, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Joung, J.-Y.; Lee, J.-S.; Cho, J.-H.; Lee, D.-S.; Ahn, Y.-C.; Son, C.-G. The Efficacy and Safety of Myelophil, an Ethanol Extract Mixture of Astragali Radix and Salviae Radix, for Chronic Fatigue Syndrome: A Randomized Clinical Trial. Front Pharmacol. 2019, 10, 991. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.G.; Lee, J.-S.; Han, J.-M.; Lee, J.-S.; Choi, M.-K.; Son, S.-W.; Kim, Y.-K.; Son, C.-G. Myelophil attenuates brain oxidative damage by modulating the hypothalamus-pituitary-adrenal (HPA) axis in a chronic cold-stress mouse model. J Ethnopharmacol. 2013, 148, 505–514. [Google Scholar] [CrossRef]
- Martinez, M.; Phillips, P.J.; Herbert, J. Adaptation in patterns of c-fos expression in the brain associated with exposure to either single or repeated social stress in male rats. Eur. J. Neurosci. 1998, 10, 20–33. [Google Scholar] [CrossRef]
- Xu, Y.; Day, T.A.; Buller, K.M. The central amygdala modulates hypothalamic-pituitary-adrenal axis responses to systemic interleukin-1beta administration. Neuroscience 1999, 94, 175–183. [Google Scholar] [CrossRef]
- Dielenberg, R.A.; Hunt, G.E.; McGregor, I.S. “When a rat smells a cat”: The distribution of Fos immunoreactivity in rat brain following exposure to a predatory odor. Neuroscience 2001, 104, 1085–1097. [Google Scholar] [CrossRef]
- Lee, W.K.; Kim, Y.; Jang, H.; Sim, J.H.; Choi, H.J.; Shin, Y.; Choi, J.J. Exogenous Transforming Growth Factor-beta in Brain-Induced Symptoms of Central Fatigue and Suppressed Dopamine Production in Mice. Int. J. Mol. Sci. 2021, 22, 2580. [Google Scholar] [CrossRef] [PubMed]
- Simon, A.R.; Severgnini, M.; Takahashi, S.; Rozo, L.; Andrahbi, B.; Agyeman, A.; Cochran, B.H.; Day, R.M.; Fanburg, B.L. 5-HT induction of c-fos gene expression requires reactive oxygen species and Rac1 and Ras GTPases. Cell Biochem. Biophys. 2005, 42, 263–276. [Google Scholar] [CrossRef]
- Wyller, V.B.; Nguyen, C.B.; Ludviksen, J.A.; Mollnes, T.E. Transforming growth factor beta (TGF-beta) in adolescent chronic fatigue syndrome. J. Transl. Med. 2017, 15, 245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, L.V.; Buckland, M.; Murphy, G.; Taylor, N.; Vleck, V.; Mein, C.; Wozniak, E.; Smuk, M.; White, P.D. Cytokine responses to exercise and activity in patients with chronic fatigue syndrome: Case-control study. Clin. Exp. Immunol. 2017, 190, 360–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, Y.; Li, Q. The variation of the 5-hydroxytryptamine system between chronic unpredictable mild stress rats and chronic fatigue syndrome rats induced by forced treadmill running. Neuroreport 2017, 28, 630–637. [Google Scholar] [CrossRef] [PubMed]
- Cercignani, M.; Dipasquale, O.; Bogdan, I.; Carandini, T.; Scott, J.; Rashid, W.; Sabri, O.; Hesse, S.; Rullmann, M.; Lopiano, L.; et al. Cognitive fatigue in multiple sclerosis is associated with alterations in the functional connectivity of monoamine circuits. Brain Commun. 2021, 3, fcab023. [Google Scholar] [CrossRef] [PubMed]
- Kong, J.; Li, X.; Yao, S. Cognition changes of chronic fatigue syndrome caused by liver stasis and spleen deficiency. Chin. J. Behav. Med. Sci. Chin. 2B06 2006, 15, j492–j493. [Google Scholar]
- Glaser, R.; Padgett, D.A.; Litsky, M.L.; Baiocchi, R.A.; Yang, E.V.; Chen, M.; Yeh, P.-E.; Klimas, N.G.; Marshall, G.D.; Whiteside, T.; et al. Stress-associated changes in the steady-state expression of latent Epstein–Barr virus: Implications for chronic fatigue syndrome and cancer. Brain Behav. Immun. 2005, 19, 91–103. [Google Scholar] [CrossRef]
- Swain, M.G. Fatigue in liver disease: Pathophysiology and clinical management. Can. J. Gastroenterol. 2006, 20, 181–188. [Google Scholar] [CrossRef]
- Cleare, A.J. Chronic fatigue syndrome. BMJ Clin. Evid. 2015, 9, 1011. [Google Scholar]
- Lee, J.S.; Kim, H.-G.; Han, J.-M.; Kim, Y.-A.; Son, C.-G. Anti-fatigue effect of Myelophil in a chronic forced exercise mouse model. Eur. J. Pharmacol. 2015, 764, 100–108. [Google Scholar] [CrossRef]
- Tomas, C.; Newton, J.; Watson, S. A review of hypothalamic-pituitary-adrenal axis function in chronic fatigue syndrome. ISRN Neurosci. 2013, 2013, 784520. [Google Scholar] [CrossRef] [Green Version]
- Brurberg, K.G.; Fonhus, M.S.; Larun, L.; Flottorp, S.; Malterud, K. Case definitions for chronic fatigue syndrome/myalgic encephalomyelitis (CFS/ME): A systematic review. BMJ Open 2014, 4, e003973. [Google Scholar] [CrossRef]
- Akazawa, K.H.; Cui, Y.; Tanaka, M.; Kataoka, Y.; Yoneda, Y.; Watanabe, Y. Mapping of regional brain activation in response to fatigue-load and recovery in rats with c-Fos immunohistochemistry. Neurosci. Res. 2010, 66, 372–379. [Google Scholar] [CrossRef]
- Inoue, T.; Tsuchiya, K.; Koyama, T. Regional changes in dopamine and serotonin activation with various intensity of physical and psychological stress in the rat brain. Pharmacol. Biochem. Behav. 1994, 49, 911–920. [Google Scholar] [CrossRef]
- Stein, D.J.; Westenberg, H.G.; Liebowitz, M.R. Social anxiety disorder and generalized anxiety disorder: Serotonergic and dopaminergic neurocircuitry. J. Clin. Psychiatry 2002, 63, 12–19. [Google Scholar] [PubMed]
- Wrase, J.; Reimold, M.; Puls, I.; Kienast, T.; Heinz, A. Serotonergic dysfunction: Brain imaging and behavioral correlates. Cognit. Affect. Behav. Neurosci. 2006, 6, 53–61. [Google Scholar] [CrossRef] [Green Version]
- Höltje, M.; von Jagov, B.; Pahner, I.; Lautenschlager, M.; Hortnagl, H.; Nurnberg, B.; Jahn, R.; Ahnert-Hilger, G. The neuronal monoamine transporter VMAT2 is regulated by the trimeric GTPase Go2. J. Neurosci. 2000, 20, 2131–2141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lukasiewicz, S.; Blasiak, E.; Szafran-Pilch, K.; Dziedzecka-Wasylewska, M. Dopamine D2 and serotonin 5-HT1A receptor interaction in the context of the effects of antipsychotics—In vitro studies. J. Neurochem. 2016, 137, 549–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asan, E.; Steinke, M.; Lesch, K.-P. Serotonergic innervation of the amygdala: Targets, receptors, and implications for stress and anxiety. Histochem. Cell Biol. 2013, 139, 785–813. [Google Scholar] [CrossRef] [PubMed]
- Turco, K. Mapping the Serotonergic System: Topographical Organization of Serotonergic Projections from the Dorsal Raphe Nucleus. Ph.D. Thesis, University of Coimbra, Coimbra, Portugal, 2013. [Google Scholar]
- Nakatomi, Y.; Mizuno, K.; Ishii, A.; Wada, Y.; Tanaka, M.; Tazawa, S.; Onoe, K.; Fukuda, S.; Kawabe, J.; Takahashi, K.; et al. Neuroinflammation in patients with chronic fatigue syndrome/myalgic encephalomyelitis: An 11C-(R)-PK11195 PET study. J. Nucl. Med. 2014, 55, 945–950. [Google Scholar] [CrossRef] [Green Version]
- Noda, M.; Ifuku, M.; Hossain, S.; Katafuchi, T. Glial Activation and Expression of the Serotonin Transporter in Chronic Fatigue Syndrome. Front. Psychiatry 2018, 9, 589. [Google Scholar] [CrossRef]
- Ariza, M.E. Myalgic Encephalomyelitis/Chronic Fatigue Syndrome: The Human Herpesviruses Are Back! Biomolecules 2021, 11, 185. [Google Scholar] [CrossRef]
- Dobryakova, E.; Genova, H.M.; DeLuca, J.; Wylie, G.R. The dopamine imbalance hypothesis of fatigue in multiple sclerosis and other neurological disorders. Front. Neurol. 2015, 6, 52. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Yang, J.-m.; Hu, T.-t.; Xu, T.-j.; Xu, W.-p.; Wei, W. Elevated dopamine D2 receptor in prefrontal cortex of CUMS rats is associated with downregulated cAMP-independent signaling pathway. Can. J. Physiol. Pharmacol. 2013, 91, 750–758. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Cao, F.; Liu, Q.; Li, X.; Xu, G.; Liu, G.; Zhang, Y.; Yang, X.; Yi, S.; Xu, F.; et al. Behavioral, inflammatory and neurochemical disturbances in LPS and UCMS-induced mouse models of depression. Behav. Brain Res. 2019, 364, 494–502. [Google Scholar] [CrossRef]
- Lesné, S.; Docagne, F.; Gabriel, C.; Liot, G.; Lahiri, D.K.; Buée, L.; Plawinski, L.; Delacourte, A.; MacKenzie, E.T.; Buisson, A.; et al. Transforming growth factor-beta 1 potentiates amyloid-beta generation in astrocytes and in transgenic mice. J. Biol. Chem. 2003, 278, 18408–18418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montoya, J.G.; Holmes, T.H.; Anderson, J.N.; Maecker, H.T.; Rosenverg-Hansson, Y.; Valencia, I.J.; Chu, V.L.; Younger, J.W.; Tota, C.M.; Davis, M.M. Cytokine signature associated with disease severity in chronic fatigue syndrome patients. Proc. Natl. Acad. Sci. USA 2017, 114, E7150–E7158. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.G.; Lee, J.S.; Lee, J.S.; Han, J.M.; Son, C.G. Hepatoprotective and antioxidant effects of Myelophil on restraint stress-induced liver injury in BALB/c mice. J. Ethnopharmacol. 2012, 142, 113–120. [Google Scholar] [CrossRef]
- Saury, J.-M. The role of the hippocampus in the pathogenesis of Myalgic Encephalomyelitis/Chronic Fatigue Syndrome (ME/CFS). Med. Hypothe. 2016, 86, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Garg, V.; Gupta, S.; Kulkarni, S.K. Role of antioxidants in chronic fatigue syndrome in mice. Indian J. Exp. Biol. 2002, 40, 1240–1244. [Google Scholar] [PubMed]
- Feuerstein, G.; Shusterman, N.; Ruffolo, R. Carvedilol update IV: Prevention of oxidative stress, cardiac remodeling and progression of congestive heart failure. Medicam. Actual. 1997, 33, 453–473. [Google Scholar]
- Deacon, R.M. Digging and marble burying in mice: Simple methods for in vivo identification of biological impacts. Nat. Protoc. 2006, 1, 122–124. [Google Scholar] [CrossRef] [PubMed]
- Eckeli, A.L.; Dach, F.; Rodrigues, A.L. Acute treatments with GMP produce antidepressant-like effects in mice. Neuroreport 2000, 11, 1839–1843. [Google Scholar] [CrossRef]
- Derrien, M.; Noble, F.; Maldonado, R.; Roques, B.P. Cholecystokinin-A but not cholecystokinin-B receptor stimulation induces endogenous opioid-dependent antinociceptive effects in the hot plate test in mice. Neurosci. Lett. 1993, 160, 193–196. [Google Scholar] [CrossRef]
- Paxinos, G.; Franklin, K.B. Paxinos and Franklin′s the Mouse Brain in Stereotaxic Coordinates; Academic Press: Cambridge, MA, USA, 2019. [Google Scholar]












Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, J.-H.; Won, S.-K.; Eom, G.-H.; Lee, D.-S.; Park, B.-J.; Lee, J.-S.; Son, C.-G.; Park, J.-Y. Improvement Effects of Myelophil on Symptoms of Chronic Fatigue Syndrome in a Reserpine-Induced Mouse Model. Int. J. Mol. Sci. 2021, 22, 10199. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910199
Song J-H, Won S-K, Eom G-H, Lee D-S, Park B-J, Lee J-S, Son C-G, Park J-Y. Improvement Effects of Myelophil on Symptoms of Chronic Fatigue Syndrome in a Reserpine-Induced Mouse Model. International Journal of Molecular Sciences. 2021; 22(19):10199. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910199
Chicago/Turabian StyleSong, Ji-Hye, Seul-Ki Won, Geun-Hyang Eom, Da-Som Lee, Byung-Jin Park, Jin-Seok Lee, Chang-Gue Son, and Ji-Yeun Park. 2021. "Improvement Effects of Myelophil on Symptoms of Chronic Fatigue Syndrome in a Reserpine-Induced Mouse Model" International Journal of Molecular Sciences 22, no. 19: 10199. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910199