
Adipose-Derived Lipid-Binding Proteins: The Good, the Bad and the Metabolic Diseases


1
Inserm, UMR 1297, Institute of Metabolic and Cardiovascular Diseases (I2MC), 31432 Toulouse, France
2
Paul Sabatier University, Université de Toulouse, 31330 Toulouse, France
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2021, 22(19), 10460; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910460
Submission received: 24 August 2021
/
Revised: 21 September 2021
/
Accepted: 22 September 2021
/
Published: 28 September 2021
(This article belongs to the Special Issue Recent Advances in Molecular Research of Metabolic Disorders)
Abstract
:Adipose tissue releases a large range of bioactive factors called adipokines, many of which are involved in inflammation, glucose homeostasis and lipid metabolism. Under pathological conditions such as obesity, most of the adipokines are upregulated and considered as deleterious, due to their pro-inflammatory, pro-atherosclerotic or pro-diabetic properties, while only a few are downregulated and would be designated as beneficial adipokines, thanks to their counteracting properties against the onset of comorbidities. This review focuses on six adipose-derived lipid-binding proteins that have emerged as key factors in the development of obesity and diabetes: Retinol binding protein 4 (RBP4), Fatty acid binding protein 4 (FABP4), Apolipoprotein D (APOD), Lipocalin-2 (LCN2), Lipocalin-14 (LCN14) and Apolipoprotein M (APOM). These proteins share structural homology and capacity to bind small hydrophobic molecules but display opposite effects on glucose and lipid metabolism. RBP4 and FABP4 are positively associated with metabolic syndrome, while APOD and LCN2 are ubiquitously expressed proteins with deleterious or beneficial effects, depending on their anatomical site of expression. LCN14 and APOM have been recently identified as adipokines associated with healthy metabolism. Recent findings on these lipid-binding proteins exhibiting detrimental or protective roles in human and murine metabolism and their involvement in metabolic diseases are also discussed.
1. Introduction
Obesity is still a major public health concern since almost half a century, and the global pandemic is overwhelming every country, even the least industrialized countries. In a 2015 study collecting obesity data from 195 countries, the Global Burden of Disease 2015 Obesity group estimated that 603 million adults and 107 million children were obese; meaning that, all over the world, one human being in 10 is obese [1].
Obesity is characterized by an excess of adipose tissue, originated from an imbalance between energy intake and energy expenditure [2]. Besides its prominent role in fatty acids trafficking, adipose tissue releases a wide range of factors called adipokines with local paracrine, and broad systemic endocrine effects on whole body homeostasis. In obese individuals, secretion of a majority of adipokines is increased. Among those, several promote inflammation and insulin resistance, which participate in the onset of cardiovascular diseases, diabetes and cancer. Till now, only a few adipokines display low levels in obesity, some having atheroprotective, anti-inflammatory and insulin-sensitizing properties such as adiponectin, the most well-known beneficial adipokine [3].
Among this swarm of factors, six are lipid-binding adipokines: Retinol binding protein 4 (RBP4), Fatty acid binding protein 4 (FABP4), Apolipoprotein D (APOD), Lipocalin-2 (LCN2), Lipocalin-14 (LCN14) and Apolipoprotein M (APOM). These lipid-binding proteins are all part of the calycin superfamily, a family of proteins with structural similarities, which is composed of the lipocalin family, the fatty acid binding protein family, the avidin family and the metalloproteinase inhibitor family [4,5].
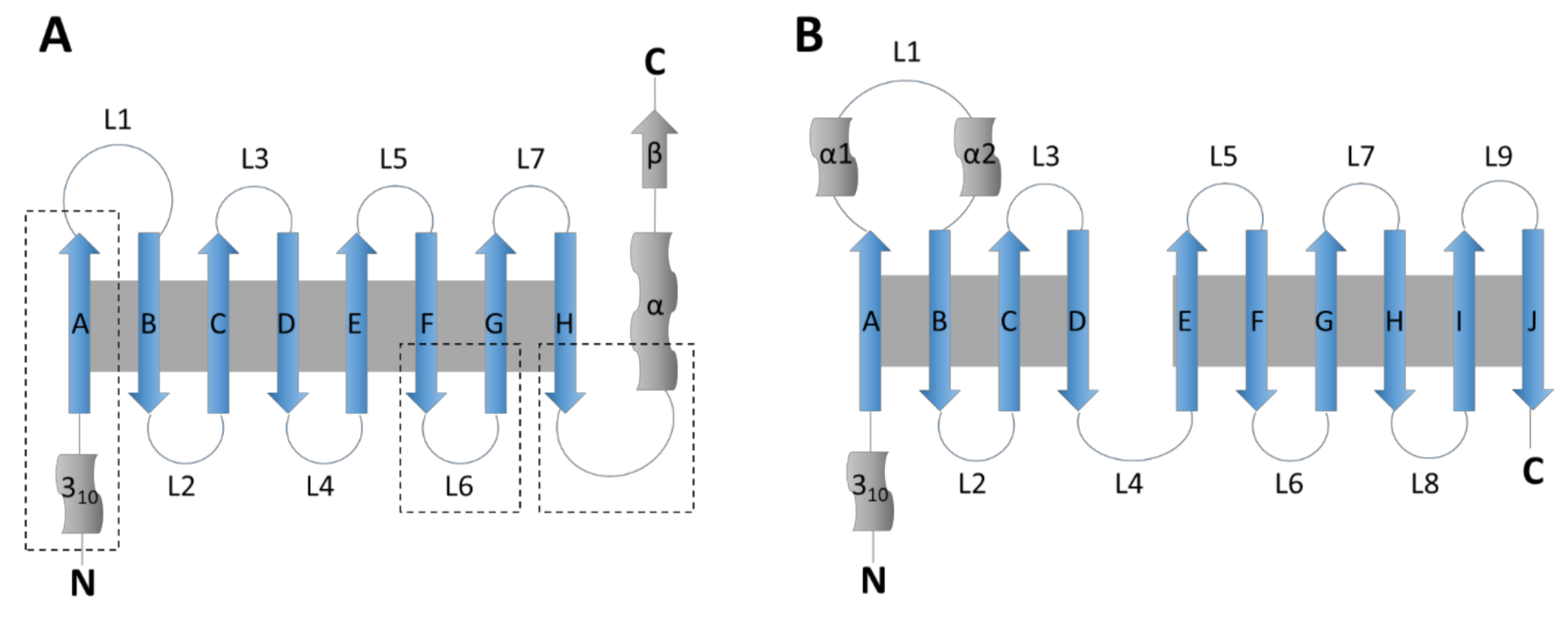
The lipocalin family was named by Pervaiz and Brew in 1987 and is composed of small extracellular proteins, generally of 160–180 amino acids, with a sequence homology barely reaching 20%. All members of this family, including RBP4, LCN2, LCN14, APOD and APOM, share a conserved tertiary structure formed by a β-barrel of eight β-strands linked by seven loops (Figure 1). The family is divided in two groups, kernel and outlier lipocalins [5]. Kernel lipocalins like RBP4, LCN2 or APOD have three highly conserved structural regions, outlier lipocalins like APOM and LCN14 only have one or two of these conserved regions [5].
The fatty acid binding proteins (FABPs) family is composed of nine proteins of approximately 15 kDa. All of them are intracellular proteins of 126–137 amino acids [6]. They were firstly identified in liver (FABP1), intestine (FABP2), heart and skeletal muscle (FABP3), adipose tissue (FABP4), epidermis (FABP5), ileum (FABP6), brain (FABP7), peripheral nervous system (Myelin P2/FABP8) and testis (FABP9), but they show no tissue-specificity. FABPs share a conserved tertiary structure formed by a β-barrel of 10 antiparallel β-strands (Figure 1). They also present two α-helixes (named A1 and A2) forming a lid over the β-barrel [7].
For lipocalins and FABPs, this specific ‘coffee-cup’ structure forms a hydrophobic pocket and allows binding to small lipidic molecules. RBP4, FABP4, APOD, LCN2, LCN14 and APOM ligands and main biological functions are summarized in Table 1.
RBP4 and FABP4 could be considered as detrimental to metabolism because they exhibit high circulating levels and adipose tissue gene expression with obesity related disorders. APOD and LCN2 are ubiquitously expressed proteins with deleterious or beneficial effects, depending on their site of expression. LCN14 and APOM are newly identified adipokines associated with healthy adipose tissue and metabolism.
In this review, we will focus on these six adipokines and their implication in metabolic disorders such as obesity and diabetes.
2. RBP4
RBP4 was identified in the 1960s by Kanai et al. as the plasma transporter of retinol complexed with transthyretin (TTR). This 21 kDa lipocalin is mainly expressed in liver and adipose tissue, and various other tissues at lower levels, and is the main—if not the only—transporter of vitamin A in vivo [8]. Some gain-of-function mutations on RBP4 promoter were reported to be associated with a higher risk of developing obesity or type 2 diabetes [15,16] while a non-coding mutation in RBP4 promoter is associated with dyslipidemia [17]. Two specific membrane receptors for RBP4 have been discovered so far: the signaling receptor and transporter of retinol 6 (STRA6), expressed widely but not in liver, and the liver-expressed retinol-binding protein receptor-2 (L-RBPR or RBPR2) [18,19].
Several publications report (i) high RBP4 concentration in plasma or high levels of RBP4 mRNA in adipose tissue of obese, diabetic individuals and their close relatives [20,21]; (ii) a positive association between serum RBP4, insulin resistance and adiposity [20,22]; (iii) a negative association (or inverse correlation) between serum RBP4 and HDL levels [20]. Similar results are observed in obese children [23]. Others also report a decrease in circulating RBP4 associated with an increase of insulin sensitivity during weight loss, subsequent to exercise training, low calorie diet or gastrectomy [20,22]. Moreover, RBP4 plasma concentrations are positively associated with blood pressure and atherosclerotic plaque in obese patients [24]. Additional recent reports in humans are reviewed in [8,9].
Furthermore, RBP4 gene expression in human adipose tissue and heart positively correlates with expression of inflammatory markers such as the mannose receptor C-type 1 (MRC1), C-C motif chemokine ligand 18 (CCL18) or Toll-like receptor 4 (TLR4), and with cardiac hypertrophy markers such as the natriuretic peptides [25,26]. RBP4-induced TLR4 expression activates the NLRP3 inflammasome in mice, which leads to the expression of inflammatory cytokines such as IL-1β, promotes insulin resistance and cardiac hypertrophy [26,27]. Animal models give further insight into the role of RBP4 in obesity and diabetes. Adipose Glut4-deficient diabetic mice display elevated Rbp4 mRNA level in adipose tissue and increased plasma RBP4, as well as ob/ob mice and mice fed with a high fat diet (HFD) [28]. Mice overexpressing human or murine RBP4 in adipocytes and fed with HFD are more prone to develop obesity, insulin resistance and hepatic steatosis than wildtype littermates [28,29]. These mice also have higher levels of inflammatory markers like tumor necrosis factor α (TNFα) [29]. Mice overexpressing Rbp4 in blood vessels musculature exhibit a higher blood pressure and an impaired eNOS-mediated vasodilatation [30]. Interestingly, these results could only be obtained in adipose tissue and muscle, as mice overexpressing Rbp4 specifically in liver do not display an impaired glucose metabolism, suggesting that RBP4 might have different roles in hepatic, adipose and vascular functions [31].
In several studies, insulin sensitivity in mice was improved when targeting RBP4. When administrated fenretinide, a synthetic retinol derivate which prevents RBP4 binding to TTR, rosiglitazone, or TTR antisense oligonucleotides which lower TTR levels and induce RBP4 clearance, insulin sensitivity in mice was restored [28,32].
So far, RBP4 has been presented as a deleterious adipokine which promotes insulin resistance, obesity and cardiovascular diseases. Even if the RBP4 signaling is not entirely deciphered, many publications tend to demonstrate the negative effect of RBP4 on metabolism and how it can be a promising therapeutic target. In addition, a recent study from Fenzl et al. showed that RBP4 is critical in humans and mice for adipose adaptative thermogenesis during a cold exposure [33]. This discovery highlights new perspectives on RBP4 role in adipose tissue and metabolism, maybe not as detrimental as it first appeared.
3. FABP4
Also called the adipocyte lipid-binding protein (ALBP), adipocyte FABP (A-FABP) or adipocyte protein 2 (aP2), FABP4 is a 15 kDa protein that binds fatty acids and structurally related lipids. It is mainly expressed in mature adipocytes, macrophages and capillary endothelial cells. In adipocytes, this calycin protein is located in the cytoplasmic compartment of the adipose cell where it interacts with the adipose triglyceride lipase (ATGL) co-activator CGi-58 and the hormone-sensitive lipase (HSL) to enhance lipolysis [10]. It is secreted in the blood stream and the adipocyte is the prevailing contributor to circulating levels [10].
In humans, serum levels of FABP4 are higher in obese, type 1 diabetic, type 2 diabetic and gestational diabetic individuals compared to non-obese, non-diabetic subjects [34,35,36,37]. Gastric bypass surgery reduces circulating FABP4 by 42% in obese patients with type 2 diabetes, whereas behavioral and nutritional intervention alone does not reduce the serum levels of FABP4 [35]. Cohort studies including obese or diabetic patients with their family revealed that FABP4 levels were high with first-degree relatives of type 1 diabetic patients [37] and in children of obese parents, even if they were not obese themselves [38]. FABP4 plasma levels are positively associated with BMI, waist circumference and metabolic syndrome, but also with inflammatory markers like CRP or IL-6 in type 2 diabetic subjects [39]. Taken together, these observations lead to the conclusion of a detrimental role of FABP4 on metabolism. Interestingly, a recent study focusing on type 1 diabetic patients revealed that low FABP4 plasma levels, due to a functional, low-expression variant in the gene promoter, was associated with a 2.4-fold higher risk of cardiovascular disease [40]. Further comparative studies on the role of FABP4 between type 1 and type 2 diabetes are required to elucidate these differences.
Adverse effects of FABP4 in patients with obesity, metabolic syndrome or diabetes are supported by in vitro and in vivo observations. Mice with diet or genetically induced obesity and lacking FABP4 have lower plasma glucose, triacylglycerol and cholesterol and better insulin sensitivity than control littermates [41,42]. Furthermore, apolipoprotein E (Apoe)-deficient mice which also lack FABP4 are less prone to develop atherosclerosis than Apoe-deficient mice with FABP4 [43]. Several studies report that FABP4 is the main responsible factor for fatty acids phagocytosis by macrophages, leading to the formation of foam cells [44]. TLR4 agonists such as lipopolysaccharide trigger the transactivation of the c-Jun N-terminal kinase (JNK) and FABP4, which leads to the recruitment of c-Jun on the FABP4 promoter and launches a positive feedback loop to accumulate FABP4 [45]. Concomitantly, FABP4 and TLR4 activation promote the nuclear factor Kappa-B (NF-κB) signaling pathway [46]. More than a fatty acid lipocalin implicated in lipids trafficking, recent advances show FABP4 as a pro-inflammatory adipokine.
Like RBP4, FABP4 has all the traits of a harmful adipokine promoting metabolic diseases. While further studies are still needed to complete FABP4 role in type 1 diabetes and in inflammation, this deleterious adipose-derived lipid-binding protein may be a new target to treat type 2 diabetes.
4. APOD
APOD was first identified by McConathy and Alaupovic in 1973 on HDL surface. It is a 169 amino acids protein with two glycosylation sites responsible for its varying molecular weight ranging from 20 to 32 kDa [47]. APOD was reported as produced by the adipose tissue in mice and humans [48,49] but also by a large variety of tissues [50]. It is able to associate with lecithin-cholesterol acyl-transferase (LCAT) and Apolipoproteins A-I or A-II, and transport cholesterol onto HDL particles [51,52]. As a lipocalin, APOD binds arachidonic acid and, with a lower affinity, progesterone and pregnenolone [53,54]. APOD gene polymorphisms were found in several populations, some of these variants being associated with a modified risk of Alzheimer’s disease or type 2 diabetes [55,56].
In a cohort of moderate to severely obese women, Desmarais et al. showed that, despite a differential expression of APOD between omental, mesenteric and round ligament adipose depots, APOD protein levels in fat tissues are positively correlated with the insulin sensitivity index QUICKI and negatively correlated with circulating levels of IL-6 and TNFα. In abdominal adipose tissue, higher APOD protein levels are associated with lower BMI, waist circumference and less insulin resistance [48]. However, APOD expression in cells extracted from amniotic fluid of obese women during pregnancy is nine-fold higher than in lean pregnant women [57]. These variations of APOD mRNA level between obese non-pregnant women and obese pregnant women might be driven by estradiol regulation on APOD expression, as circulating estradiol concentration increases during pregnancy and downregulates APOD expression in breast cancer [58]. In gestational diabetes, APOD placenta and plasma levels are higher in women with gestational diabetes than women without, whether they are overweight, obese [59] or not [60].
Apod-deficient mice display an altered lipidomic profile with elevated plasma triglycerides and insulin, without changes in glucose homeostasis evaluated by glucose tolerance test [61]; whereas mice overexpressing human APOD develop insulin resistance and hepatic steatosis with aging, but not with obesity or inflammation [62,63]. Hepatic steatosis after human APOD overexpression in mice is due to an upregulation of PPARγ, leading to an increased formation of lipid droplets in hepatocytes [64]. Obese and diabetic models such as the ob/ob and db/db mice exhibit lower levels of plasma APODthan heterozygous control littermates [65,66], which might be due to an altered interaction between APOD and the leptin receptor [65,67]. Similar results were obtained in obese male rats, in which Apod expression was downregulated in the reproductive system after a high fat diet [68].
On another hand, Apod-deficient mice have a higher sensitivity to oxidative stress in brain and develop locomotor and learning issues, while an overexpression of human APOD in brain rescued this phenotype [69]. In murine NIH/3T3 cells, a study tested various stress conditions in order to evaluate Apod regulation. The authors show that hydrogen peroxide H2O2 upregulates Apod expression in these adipocytes [70]. Taken together, these data suggest antioxidant properties of APOD.
Such characteristics have made APOD a debated topic in the field of obesity, diabetes and cardiovascular diseases. Further studies are needed to clarify the role of APOD in metabolic diseases, especially in obesity and type 2 diabetes, including men. So far, only the neuroprotective and antioxidant properties of APOD are known [11,71].
5. LCN2
The human neutrophil gelatinase-associated lipocalin NGAL, siderocalin, murine 24p3 protein or LCN2 was first identified in the early 1990s as a small protein secreted by neutrophils and bound to the matrix metalloproteinase (MMP)-9 [12,72]. It is a 25 kDa lipocalin able to bind iron and siderophores, bacterial iron-binding metabolites which captures iron from its host transporter proteins, therefore inducing a bacteriostatic effect [73]. This unique characteristic among the lipocalin family explains the wide expression pattern of LCN2 in pathogen-exposed tissues such as salivary glands, stomach, appendix, colon, trachea, lung, heart, kidney, uterus, prostate, blood cells and bone marrow [74,75,76].
Recent studies have demonstrated that plasma LCN2 is elevated in patients with acute myocardial infarction and is associated with poor outcomes like cardiogenic shock or increased risk of mortality [77,78]. As LCN2 levels also positively correlate with kidney disease, it has been proposed as a novel biomarker for cardiac event and as a predictive marker for acute kidney injury following a cardiac surgery [79].
LCN2 is highly expressed in liver [80] and adipose tissue [81], so its role in metabolic diseases has been explored. In human cohort studies, plasma level and mRNA expression of LCN2 in liver and adipose tissue are higher in obese and diabetic subjects compared with lean individuals [80,82,83]. Despite higher levels in men than women, this difference remains after adjustment for sex [80]. Wang et al. also reported a positive correlation between serum LCN2 and indexes of adiposity (e.g., BMI, waist circumference, body fat percentage) and indexes of insulin resistance (e.g., HOMA-IR, fasting insulin and glucose levels) but this is not confirmed by another study [82]. LCN2 plasma levels and expression in adipose tissue are also positively associated with inflammatory markers in humans, hs-CRP and IL-6 [80,82]. An 8-week rosiglitazone treatment decreases the plasma levels of CRP as well as LCN2. Petropoulou et al. showed that circulating LCN2 increases after a meal in lean individuals, but not in obese subjects, and this increase correlates with a reduced hunger feeling [84], suggesting an anorexigenic effect of LCN2 in lean humans. This was supported by human LCN2 administration in monkeys, which displayed an acute 21% reduced food intake without short-time side effects.
However, these observations are in contradiction with a recent study, which shows an increase of food intake in Lcn2-overexpressing mice [85]. The authors also demonstrate high insulin levels in plasma from these mice. Moreover, db/db mice and streptozotocin-treated mice have higher circulating LCN2 and mRNA levels in liver and adipose tissue than wild-type littermates [80,86].
Several publications are in contradiction regarding the role of LCN2 in insulin resistance in mice. The same year, while Law et al. reported an improved insulin sensitivity in Lcn2-deficient mice compared to wildtype littermates [87], Guo et al. showed that Lcn2 deficiency in mice promotes insulin resistance [88]. This paradox was studied by another group which reported increased insulin levels and increased insulin sensitivity in Lcn2-overexpressing mice [89]. The authors gave evidence of an improved β-cell function and insulin secretion in these mice, thereby increasing plasma insulin, a hallmark of insulin resistance. Moreover, LCN2 promotes muscle differentiation, improves thermogenic function of brown adipose tissue (BAT) and triggers beiging in white adipose tissue [90,91]. Interestingly, activation of BAT consequently to LCN2 signal is independent of the classical β-adrenergic/AMPK pathway and involves p38/MAPK [92].
Further studies are needed to get an overview of all LCN2 effects on metabolic pathways. While LCN2 is considered as a predictive marker for cardiac outcomes, its importance in immunity response has also been considered. Regarding its role in the onset of diabetes, a novel hypothesis is emerging: LCN2 may be upregulated during the earliest stage of diabetes to promote β-cell function and insulin sensitivity but this protective mechanism is eventually overwhelmed when diabetes settles.
6. LCN14
LCN14 was first identified in mice as an odorant binding protein (OBP) with two isoforms, 2a and 2b. In mice, it shares a 50% homology with the OBP-1 variant produced by hepatocytes, also known as lipocalin-13. LCN14 is mainly expressed in white adipose tissue and poorly expressed in brown adipose tissue. In high fat diet-induced obese mice as well as diabetic db/db mice, LCN14 expression in the adipose tissue is lower than in wildtype, normal chow diet-fed mice [93].
Lee et al. showed that LCN14 is a beneficial adipokine which is able to enhance insulin sensitivity of liver and adipose tissue [93]. Overexpression reduces isoprenaline induced lipolysis and therefore might limit hepatic gluconeogenesis [93].
Murine LCN14 shares a 60% homology with human OBP2a and OBP2b [94,95], which also share similarities with rat OBP2. These two human isoforms, similar at 90%, display ability to bind olfactory molecules in the nasal tractus but so far, their expression in the adipose tissue has not been studied [95,96].
Overall, the putative positive role of LCN14 in metabolic homeostasis now deserves additional studies.
7. APOM
APOM was discovered in 1999 by Xu and Dahlbäck as a novel apolipoprotein mainly secreted by liver and kidney, and at lower level by adipose tissues [13]. Plasma APOM is mostly found as monomers on HDL, yet a recent study gave evidence of an in vitro homo- or heterodimerization of APOM when HDL particles are incubated in high-glucose buffer, though the authors could not find any APOM dimers in serums of diabetic subjects [97]. The human APOM protein is a lipocalin with one glycation site, so that its molecular weight varies from 21 to 26 kDa, but the murine APOM protein has an uncommon β-barrel lipocalin structure formed by only seven β-strands and no glycation site [98]. Whether this rare structure influences the murine APOM binding and affinity for its ligands is still an open question. In plasma, APOM is the main chaperone for S1P on HDL, also it binds retinol, retinoic acid, oxidized phospholipids and some fatty acids [13]. S1P is a bioactive lipid with diverse effects due to its five different G-protein coupled receptors, S1P1 to S1P5. S1P1 to S1P3 are widely distributed, while S1P4 is almost exclusively expressed in the lymphatic system and S1P5 in the nervous system. When bound to S1P1 or S1P3, S1P displays protective effects against insulin resistance and atherosclerosis, whereas S1P bound on S1P2 promotes insulin resistance and atherosclerosis [99,100]. Moreover, some genetic variants of the APOM gene reported in the Chinese population associate low plasma APOM levels with a higher risk of developing type 2 diabetes [101]. Therefore, the APOM/S1P complex as well as free APOM might be interesting targets in the metabolic diseases research area.
Plasma APOM levels are lower in obese, metabolic syndrome or type 2 diabetic subjects, and in women with gestational diabetes compared to lean, non-diabetic individuals. APOM expression in adipose tissue and liver is concordant with circulating APOM levels [60,102,103]. However, in type 1 diabetic patients, plasma APOM levels are not different to plasma levels from non-diabetic controls, in two cohort studies [104,105]. When submitted to a hypocaloric diet inducing a significant weight loss and improvements in the metabolic profile, APOM expression and secretion from the adipose tissue of obese individuals increase during calorie restriction [102]. Further details on human cohorts are reviewed in [14].
Animal studies provide further insight into the role of APOM in metabolic diseases. Goto-Kakizaki rats, a non-obese, spontaneous model of diabetic rats display better insulin sensitivity when overexpressing Apom, concomitantly with an increased insulin secretion and glucose utilization [106]. Concordant results were obtained in Apom-deficient mice which present impaired insulin sensitivity, high plasma inflammatory markers such as IL-6 or IL-1β, and hepatic NF-κB [99,107]. Yao et al. also recently demonstrated that APOM is expressed in murine and human macrophages, interacts with the scavenger receptor BI to promote cholesterol efflux from the phagocytic cells and protects against atherosclerosis [108,109]. More, Apom-deficient mice display blood hypertension and cardiac hypertrophy compared to wildtype animals [110].
Taken together, these observations indicate that APOM might be beneficial in the field of cardiac and metabolic diseases. However, APOM binding to S1P may complicate the elucidation of APOM part on insulin sensitivity. Christoffersen et al. showed that Apom-deficient mice have a more active BAT and improved insulin sensitivity [111], a phenotype also observed in S1P2-deficient mice [112]. Whether APOM only acts as a chaperone for S1P or is by itself a beneficial cardiometabolic adipokine is still a matter of debate, the latter being more and more considered.
8. Clinical Perspectives
Targeting RBP4, FABP4 or their signaling pathway to treat diabetes or atherosclerosis has been a profuse topic for the last few years. Torabi et al. designed calcium-phosphate nanoparticles conjugated with a single-stranded DNA aptamer which specifically binds to RBP4. This engineered nanoparticle ligand inhibits RBP4 binding to its natural ligand, vitamin A [113]. Other groups have tested already commercially available drugs, used for their antidiabetic properties, and studied the variations of RBP4 and FABP4 levels: both sodium-glucose cotransporter 2 (SGLT2) and dipeptidyl peptidase 4 (DDP4) inhibitors in combination with metformin can decrease RBP4 and FABP4 levels [114], while metformin alone reduces RBP4 and FABP4 levels and inflammatory markers [115]. Angiotensin II receptor blockers (namely candesartan, olmesartan, valsartan and telmisartan) [116], atorvastatin [117] and omega-3 fatty acids (namely eicosapentaenoic and docosahexaenoic acids) [118] induce a 8–20% decrease of FABP4 plasma levels in diabetic, hypertensive or dyslipidemic patients, whereas treatment of human adipocytes in vitro with bisphenol A induces an increase of FABP4 expression [119].
So far, two groups have successfully used monoclonal antibodies directed against FABP4 which improve insulin sensitivity or attenuate inflammation [120,121], while others have developed novel small inhibiting molecules which stick in FABP4 hydrophobic pocket as molecular lures and prevent its effects [122,123].
9. Conclusions and Perspectives
Adipose tissue secretes a large range of bioactive molecules, including lipids. In healthy states, these factors maintain metabolic homeostasis. When adipose tissue expands, many exhibit increased secretion and are implicated in metabolic disturbances, and some that show lower secretion might contribute to maintain “healthy” metabolism. Among the six lipid-binding adipokines discussed in this review, most display effects closely associated with their ligand’s biological properties. In addition, these lipid-binding proteins have a large diversity of ligands, from fatty acids to sphingolipids, odorant volatile molecules to haeme metabolites. To understand the complex mechanisms by which those lipid-binding adipokines exert their effect, identification of other ligands warrants further investigations. Consideration should also be given to the possibility that these adipose-derived lipid-binding proteins have ligand-independent roles in metabolic diseases.
Funding
Some of the work discussed here has received support from Inserm, Paul Sabatier University, the Innovative Medicines Initiative Joint Undertaking (grant agreement n° 115372), the Commission of the European Communities (FP6-513946 DiOGenes), F-CRIN/FORCE network, and the Société Française de Nutrition. Laurie Frances was funded by Inserm and the Occitanie/Pyrénées-Méditerranée region, France.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Collaborators, G.B.D.O.; Afshin, A.; Forouzanfar, M.H.; Reitsma, M.B.; Sur, P.; Estep, K.; Lee, A.; Marczak, L.; Mokdad, A.H.; Moradi-Lakeh, M.; et al. Health Effects of Overweight and Obesity in 195 Countries over 25 Years. N. Engl. J. Med. 2017, 377, 13–27. [Google Scholar] [CrossRef]
- Mittendorfer, B. Origins of metabolic complications in obesity: Adipose tissue and free fatty acid trafficking. Curr. Opin. Clin. Nutr. Metab. Care 2011, 14, 535–541. [Google Scholar] [CrossRef] [Green Version]
- Bluher, M.; Rudich, A.; Kloting, N.; Golan, R.; Henkin, Y.; Rubin, E.; Schwarzfuchs, D.; Gepner, Y.; Stampfer, M.J.; Fiedler, M.; et al. Two patterns of adipokine and other biomarker dynamics in a long-term weight loss intervention. Diabetes Care 2012, 35, 342–349. [Google Scholar] [CrossRef] [Green Version]
- Flower, D.R.; North, A.C.; Attwood, T.K. Structure and sequence relationships in the lipocalins and related proteins. Protein Sci. 1993, 2, 753–761. [Google Scholar] [CrossRef] [Green Version]
- Flower, D.R.; North, A.C.; Sansom, C.E. The lipocalin protein family: Structural and sequence overview. Biochim. Biophys. Acta 2000, 1482, 9–24. [Google Scholar] [CrossRef]
- Glatz, J.F.; van der Vusse, G.J. Cellular fatty acid-binding proteins: Their function and physiological significance. Prog. Lipid Res. 1996, 35, 243–282. [Google Scholar] [CrossRef]
- Scapin, G.; Young, A.C.; Kromminga, A.; Veerkamp, J.H.; Gordon, J.I.; Sacchettini, J.C. High resolution X-ray studies of mammalian intestinal and muscle fatty acid-binding proteins provide an opportunity for defining the chemical nature of fatty acid: Protein interactions. Mol. Cell. Biochem. 1993, 123, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Steinhoff, J.S.; Lass, A.; Schupp, M. Biological Functions of RBP4 and Its Relevance for Human Diseases. Front. Physiol. 2021, 12, 659977. [Google Scholar] [CrossRef] [PubMed]
- Nono Nankam, P.A.; Bluher, M. Retinol-binding protein 4 in obesity and metabolic dysfunctions. Mol. Cell. Endocrinol. 2021, 531, 111312. [Google Scholar] [CrossRef]
- Lee, C.H.; Lui, D.T.W.; Lam, K.S.L. Adipocyte Fatty Acid-Binding Protein, Cardiovascular Diseases and Mortality. Front. Immunol. 2021, 12, 589206. [Google Scholar] [CrossRef] [PubMed]
- Rassart, E.; Desmarais, F.; Najyb, O.; Bergeron, K.F.; Mounier, C. Apolipoprotein D. Gene 2020, 756, 144874. [Google Scholar] [CrossRef] [PubMed]
- Krizanac, M.; Mass Sanchez, P.B.; Weiskirchen, R.; Asimakopoulos, A. A Scoping Review on Lipocalin-2 and Its Role in Non-Alcoholic Steatohepatitis and Hepatocellular Carcinoma. Int. J. Mol. Sci. 2021, 22, 2865. [Google Scholar] [CrossRef] [PubMed]
- Tavernier, G.; Caspar-Bauguil, S.; Viguerie, N. Apolipoprotein M: New connections with diet, adipose tissue and metabolic syndrome. Curr. Opin. Lipidol. 2020, 31, 8–14. [Google Scholar] [CrossRef]
- Christoffersen, C. Apolipoprotein M-A Marker or an Active Player in Type II Diabetes? Front. Endocrinol. 2021, 12, 665393. [Google Scholar] [CrossRef] [PubMed]
- van Hoek, M.; Dehghan, A.; Zillikens, M.C.; Hofman, A.; Witteman, J.C.; Sijbrands, E.J. An RBP4 promoter polymorphism increases risk of type 2 diabetes. Diabetologia 2008, 51, 1423–1428. [Google Scholar] [CrossRef] [Green Version]
- Shajarian, M.; Rafiee, L.; Naji-Esfahani, H.; Haghjooy-Javanmard, S.; Nizal, S. Association of RBP4 gene variants with adverse lipid profile and obesity. Gene 2015, 561, 1–5. [Google Scholar] [CrossRef]
- Aref-Eshghi, E.; Hurley, O.; Sun, G.; Simms, A.; Godwin, M.; Duke, P.; Araee, M.; Mahdavian, M.; Asghari, S. Genetic associations in community context: A mixed model approach identifies a functional variant in the RBP4 gene associated with HDL-C dyslipidemia. BMC Med. Genet. 2018, 19, 205. [Google Scholar] [CrossRef]
- Kawaguchi, R.; Yu, J.; Honda, J.; Hu, J.; Whitelegge, J.; Ping, P.; Wiita, P.; Bok, D.; Sun, H. A membrane receptor for retinol binding protein mediates cellular uptake of vitamin A. Science 2007, 315, 820–825. [Google Scholar] [CrossRef]
- Alapatt, P.; Guo, F.; Komanetsky, S.M.; Wang, S.; Cai, J.; Sargsyan, A.; Rodriguez Diaz, E.; Bacon, B.T.; Aryal, P.; Graham, T.E. Liver retinol transporter and receptor for serum retinol-binding protein (RBP4). J. Biol. Chem. 2013, 288, 1250–1265. [Google Scholar] [CrossRef] [Green Version]
- Graham, T.E.; Yang, Q.; Bluher, M.; Hammarstedt, A.; Ciaraldi, T.P.; Henry, R.R.; Wason, C.J.; Oberbach, A.; Jansson, P.A.; Smith, U.; et al. Retinol-binding protein 4 and insulin resistance in lean, obese, and diabetic subjects. N. Engl. J. Med. 2006, 354, 2552–2563. [Google Scholar] [CrossRef]
- Hammarstedt, A.; Graham, T.E.; Kahn, B.B. Adipose tissue dysregulation and reduced insulin sensitivity in non-obese individuals with enlarged abdominal adipose cells. Diabetol. Metab. Syndr. 2012, 4, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Huang, Y.; Gao, J.; Sun, H.; Jayachandran, M.; Qu, S. Changes of serum retinol-binding protein 4 associated with improved insulin resistance after laparoscopic sleeve gastrectomy in Chinese obese patients. Diabetol. Metab. Syndr. 2020, 12, 7. [Google Scholar] [CrossRef] [PubMed]
- Reinehr, T.; Stoffel-Wagner, B.; Roth, C.L. Retinol-binding protein 4 and its relation to insulin resistance in obese children before and after weight loss. J. Clin. Endocrinol. Metab. 2008, 93, 2287–2293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dessein, P.H.; Tsang, L.; Norton, G.R.; Woodiwiss, A.J.; Solomon, A. Retinol binding protein 4 concentrations relate to enhanced atherosclerosis in obese patients with rheumatoid arthritis. PLoS ONE 2014, 9, e92739. [Google Scholar] [CrossRef] [Green Version]
- Kilicarslan, M.; de Weijer, B.A.; Simonyte Sjodin, K.; Aryal, P.; Ter Horst, K.W.; Cakir, H.; Romijn, J.A.; Ackermans, M.T.; Janssen, I.M.; Berends, F.J.; et al. RBP4 increases lipolysis in human adipocytes and is associated with increased lipolysis and hepatic insulin resistance in obese women. FASEB J. 2020, 34, 6099–6110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, W.; Wang, H.; Zhang, L.; Cao, Y.; Bao, J.Z.; Liu, Z.X.; Wang, L.S.; Yang, Q.; Lu, X. Retinol-Binding Protein 4 Induces Cardiomyocyte Hypertrophy by Activating TLR4/MyD88 Pathway. Endocrinology 2016, 157, 2282–2293. [Google Scholar] [CrossRef] [Green Version]
- Moraes-Vieira, P.M.; Yore, M.M.; Sontheimer-Phelps, A.; Castoldi, A.; Norseen, J.; Aryal, P.; Simonyte Sjodin, K.; Kahn, B.B. Retinol binding protein 4 primes the NLRP3 inflammasome by signaling through Toll-like receptors 2 and 4. Proc. Natl. Acad. Sci. USA 2020, 117, 31309–31318. [Google Scholar] [CrossRef]
- Yang, Q.; Graham, T.E.; Mody, N.; Preitner, F.; Peroni, O.D.; Zabolotny, J.M.; Kotani, K.; Quadro, L.; Kahn, B.B. Serum retinol binding protein 4 contributes to insulin resistance in obesity and type 2 diabetes. Nature 2005, 436, 356–362. [Google Scholar] [CrossRef]
- Lee, S.A.; Yuen, J.J.; Jiang, H.; Kahn, B.B.; Blaner, W.S. Adipocyte-specific overexpression of retinol-binding protein 4 causes hepatic steatosis in mice. Hepatology 2016, 64, 1534–1546. [Google Scholar] [CrossRef] [Green Version]
- Kraus, B.J.; Sartoretto, J.L.; Polak, P.; Hosooka, T.; Shiroto, T.; Eskurza, I.; Lee, S.A.; Jiang, H.; Michel, T.; Kahn, B.B. Novel role for retinol-binding protein 4 in the regulation of blood pressure. FASEB J. 2015, 29, 3133–3140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fedders, R.; Muenzner, M.; Weber, P.; Sommerfeld, M.; Knauer, M.; Kedziora, S.; Kast, N.; Heidenreich, S.; Raila, J.; Weger, S.; et al. Liver-secreted RBP4 does not impair glucose homeostasis in mice. J. Biol. Chem. 2018, 293, 15269–15276. [Google Scholar] [CrossRef] [Green Version]
- Zemany, L.; Bhanot, S.; Peroni, O.D.; Murray, S.F.; Moraes-Vieira, P.M.; Castoldi, A.; Manchem, P.; Guo, S.; Monia, B.P.; Kahn, B.B. Transthyretin Antisense Oligonucleotides Lower Circulating RBP4 Levels and Improve Insulin Sensitivity in Obese Mice. Diabetes 2015, 64, 1603–1614. [Google Scholar] [CrossRef] [Green Version]
- Fenzl, A.; Kulterer, O.C.; Spirk, K.; Mitulovic, G.; Marculescu, R.; Bilban, M.; Baumgartner-Parzer, S.; Kautzky-Willer, A.; Kenner, L.; Plutzky, J.; et al. Intact vitamin A transport is critical for cold-mediated adipose tissue browning and thermogenesis. Mol. Metab. 2020, 42, 101088. [Google Scholar] [CrossRef]
- Ebert, T.; Roth, I.; Richter, J.; Tonjes, A.; Kralisch, S.; Lossner, U.; Kratzsch, J.; Bluher, M.; Stumvoll, M.; Fasshauer, M. Different associations of adipokines in lean and healthy adults. Horm. Metab. Res. 2014, 46, 41–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jahansouz, C.; Xu, H.; Kizy, S.; Thomas, A.J.; Josephrajan, A.; Hertzel, A.V.; Foncea, R.; Connett, J.C.; Billington, C.J.; Jensen, M.; et al. Serum FABP4 concentrations decrease after Roux-en-Y gastric bypass but not after intensive medical management. Surgery 2019, 165, 571–578. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Zhang, D.; Xu, J.; Chen, C.; Deng, D.; Pan, F.; Dong, L.; Li, S.; Ye, S. Circulating FABP4, nesfatin-1, and osteocalcin concentrations in women with gestational diabetes mellitus: A meta-analysis. Lipids Health Dis. 2020, 19, 199. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Shu, L.; Wu, X.; Liu, Y.; Cheong, L.Y.; Liao, B.; Xiao, X.; Hoo, R.L.; Zhou, Z.; Xu, A. Fatty acid binding protein 4 promotes autoimmune diabetes by recruitment and activation of pancreatic islet macrophages. JCI Insight 2021, 6, e141814. [Google Scholar] [CrossRef]
- Zachariah, J.P.; Quiroz, R.; Enserro, D.; Andersson, C.; Keaney, J.F., Jr.; Sullivan, L.M.; Vasan, R.S. Association of Parental Obesity and Diabetes Mellitus With Circulating Adipokines in Nonobese Nondiabetic Offspring. J. Am. Heart Assoc. 2017, 6, e004973. [Google Scholar] [CrossRef]
- Bagheri, R.; Qasim, A.N.; Mehta, N.N.; Terembula, K.; Kapoor, S.; Braunstein, S.; Schutta, M.; Iqbal, N.; Lehrke, M.; Reilly, M.P. Relation of plasma fatty acid binding proteins 4 and 5 with the metabolic syndrome, inflammation and coronary calcium in patients with type-2 diabetes mellitus. Am. J. Cardiol. 2010, 106, 1118–1123. [Google Scholar] [CrossRef] [Green Version]
- Dahlstrom, E.H.; Saksi, J.; Forsblom, C.; Uglebjerg, N.; Mars, N.; Thorn, L.M.; Harjutsalo, V.; Rossing, P.; Ahluwalia, T.S.; Lindsberg, P.J.; et al. The low-expression variant of FABP4 is associated with cardiovascular disease in type 1 diabetes. Diabetes 2021. [Google Scholar] [CrossRef]
- Baar, R.A.; Dingfelder, C.S.; Smith, L.A.; Bernlohr, D.A.; Wu, C.; Lange, A.J.; Parks, E.J. Investigation of in vivo fatty acid metabolism in AFABP/aP2(-/-) mice. Am. J. Physiol. Endocrinol. Metab. 2005, 288, E187–E193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maeda, K.; Cao, H.; Kono, K.; Gorgun, C.Z.; Furuhashi, M.; Uysal, K.T.; Cao, Q.; Atsumi, G.; Malone, H.; Krishnan, B.; et al. Adipocyte/macrophage fatty acid binding proteins control integrated metabolic responses in obesity and diabetes. Cell Metab. 2005, 1, 107–119. [Google Scholar] [CrossRef] [Green Version]
- Makowski, L.; Boord, J.B.; Maeda, K.; Babaev, V.R.; Uysal, K.T.; Morgan, M.A.; Parker, R.A.; Suttles, J.; Fazio, S.; Hotamisligil, G.S.; et al. Lack of macrophage fatty-acid-binding protein aP2 protects mice deficient in apolipoprotein E against atherosclerosis. Nat. Med. 2001, 7, 699–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erbay, E.; Babaev, V.R.; Mayers, J.R.; Makowski, L.; Charles, K.N.; Snitow, M.E.; Fazio, S.; Wiest, M.M.; Watkins, S.M.; Linton, M.F.; et al. Reducing endoplasmic reticulum stress through a macrophage lipid chaperone alleviates atherosclerosis. Nat. Med. 2009, 15, 1383–1391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hui, X.; Li, H.; Zhou, Z.; Lam, K.S.; Xiao, Y.; Wu, D.; Ding, K.; Wang, Y.; Vanhoutte, P.M.; Xu, A. Adipocyte fatty acid-binding protein modulates inflammatory responses in macrophages through a positive feedback loop involving c-Jun NH2-terminal kinases and activator protein-1. J. Biol. Chem. 2010, 285, 10273–10280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, F.; Chen, G.; Yang, Y.; Lei, M. Fatty acid-binding protein 4 silencing protects against lipopolysaccharide-induced cardiomyocyte hypertrophy and apoptosis by inhibiting the Toll-like receptor 4-nuclear factor-kappaB pathway. J. Int. Med. Res. 2021, 49, 300060521998233. [Google Scholar] [CrossRef] [PubMed]
- Boyles, J.K.; Notterpek, L.M.; Wardell, M.R.; Rall, S.C., Jr. Identification, characterization, and tissue distribution of apolipoprotein D in the rat. J. Lipid Res. 1990, 31, 2243–2256. [Google Scholar] [CrossRef]
- Desmarais, F.; Bergeron, K.F.; Lacaille, M.; Lemieux, I.; Bergeron, J.; Biron, S.; Rassart, E.; Joanisse, D.R.; Mauriege, P.; Mounier, C. High ApoD protein level in the round ligament fat depot of severely obese women is associated with an improved inflammatory profile. Endocrine 2018, 61, 248–257. [Google Scholar] [CrossRef]
- Yoshida, K.; Cleaveland, E.S.; Nagle, J.W.; French, S.; Yaswen, L.; Ohshima, T.; Brady, R.O.; Pentchev, P.G.; Kulkarni, A.B. Molecular cloning of the mouse apolipoprotein D gene and its upregulated expression in Niemann-Pick disease type C mouse model. DNA Cell Biol. 1996, 15, 873–882. [Google Scholar] [CrossRef] [PubMed]
- Drayna, D.T.; McLean, J.W.; Wion, K.L.; Trent, J.M.; Drabkin, H.A.; Lawn, R.M. Human apolipoprotein D gene: Gene sequence, chromosome localization, and homology to the alpha 2u-globulin superfamily. DNA 1987, 6, 199–204. [Google Scholar] [CrossRef]
- Fielding, P.E.; Fielding, C.J. A cholesteryl ester transfer complex in human plasma. Proc. Natl. Acad. Sci. USA 1980, 77, 3327–3330. [Google Scholar] [CrossRef] [Green Version]
- Steyrer, E.; Kostner, G.M. Activation of lecithin-cholesterol acyltransferase by apolipoprotein D: Comparison of proteoliposomes containing apolipoprotein D, A-I or C-I. Biochim. Biophys. Acta 1988, 958, 484–491. [Google Scholar] [CrossRef]
- Eichinger, A.; Nasreen, A.; Kim, H.J.; Skerra, A. Structural insight into the dual ligand specificity and mode of high density lipoprotein association of apolipoprotein D. J. Biol. Chem. 2007, 282, 31068–31075. [Google Scholar] [CrossRef] [Green Version]
- Morais Cabral, J.H.; Atkins, G.L.; Sanchez, L.M.; Lopez-Boado, Y.S.; Lopez-Otin, C.; Sawyer, L. Arachidonic acid binds to apolipoprotein D: Implications for the protein’s function. FEBS Lett. 1995, 366, 53–56. [Google Scholar] [CrossRef] [Green Version]
- Baker, W.A.; Hitman, G.A.; Hawrami, K.; McCarthy, M.I.; Riikonen, A.; Tuomilehto-Wolf, E.; Nissinen, A.; Tuomilehto, J.; Mohan, V.; Viswanathan, M.; et al. Apolipoprotein D gene polymorphism: A new genetic marker for type 2 diabetic subjects in Nauru and south India. Diabet. Med. 1994, 11, 947–952. [Google Scholar] [CrossRef]
- Shibata, N.; Nagata, T.; Shinagawa, S.; Ohnuma, T.; Shimazaki, H.; Komatsu, M.; Kuerban, B.; Tomson, K.; Nakayama, K.; Yamada, H.; et al. Genetic association between APOA1 and APOD polymorphisms and Alzheimer’s disease in a Japanese population. J. Neural Transm. 2013, 120, 1599–1603. [Google Scholar] [CrossRef] [PubMed]
- Edlow, A.G.; Vora, N.L.; Hui, L.; Wick, H.C.; Cowan, J.M.; Bianchi, D.W. Maternal obesity affects fetal neurodevelopmental and metabolic gene expression: A pilot study. PLoS ONE 2014, 9, e88661. [Google Scholar] [CrossRef] [PubMed]
- Simard, J.; Dauvois, S.; Haagensen, D.E.; Levesque, C.; Merand, Y.; Labrie, F. Regulation of progesterone-binding breast cyst protein GCDFP-24 secretion by estrogens and androgens in human breast cancer cells: A new marker of steroid action in breast cancer. Endocrinology 1990, 126, 3223–3231. [Google Scholar] [CrossRef] [PubMed]
- Navarro, A.; Alonso, A.; Garrido, P.; Gonzalez, C.; Gonzalez Del Rey, C.; Ordonez, C.; Tolivia, J. Increase in placental apolipoprotein D as an adaptation to human gestational diabetes. Placenta 2010, 31, 25–31. [Google Scholar] [CrossRef]
- Ravnsborg, T.; Andersen, L.L.; Trabjerg, N.D.; Rasmussen, L.M.; Jensen, D.M.; Overgaard, M. First-trimester multimarker prediction of gestational diabetes mellitus using targeted mass spectrometry. Diabetologia 2016, 59, 970–979. [Google Scholar] [CrossRef] [Green Version]
- Jimenez-Palomares, M.; Cozar-Castellano, I.; Ganfornina, M.D.; Sanchez, D.; Perdomo, G. Genetic deficiency of apolipoprotein D in the mouse is associated with nonfasting hypertriglyceridemia and hyperinsulinemia. Metabolism 2011, 60, 1767–1774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desmarais, F.; Bergeron, K.F.; Rassart, E.; Mounier, C. Apolipoprotein D overexpression alters hepatic prostaglandin and omega fatty acid metabolism during the development of a non-inflammatory hepatic steatosis. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2019, 1864, 522–531. [Google Scholar] [CrossRef]
- Do Carmo, S.; Fournier, D.; Mounier, C.; Rassart, E. Human apolipoprotein D overexpression in transgenic mice induces insulin resistance and alters lipid metabolism. Am. J. Physiol. Endocrinol. Metab. 2009, 296, E802–E811. [Google Scholar] [CrossRef] [Green Version]
- Labrie, M.; Lalonde, S.; Najyb, O.; Thiery, M.; Daneault, C.; Des Rosiers, C.; Rassart, E.; Mounier, C. Apolipoprotein D Transgenic Mice Develop Hepatic Steatosis through Activation of PPARgamma and Fatty Acid Uptake. PLoS ONE 2015, 10, e0130230. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Chang, G.Q.; Leibowitz, S.F. Apolipoprotein D interacts with the long-form leptin receptor: A hypothalamic function in the control of energy homeostasis. FASEB J. 2001, 15, 1329–1331. [Google Scholar] [CrossRef] [Green Version]
- Perdomo, G.; Kim, D.H.; Zhang, T.; Qu, S.; Thomas, E.A.; Toledo, F.G.; Slusher, S.; Fan, Y.; Kelley, D.E.; Dong, H.H. A role of apolipoprotein D in triglyceride metabolism. J. Lipid Res. 2010, 51, 1298–1311. [Google Scholar] [CrossRef] [Green Version]
- El-Darzi, N.; Mast, N.; Petrov, A.M.; Dao, T.; Astafev, A.A.; Saadane, A.; Prendergast, E.; Schwarz, E.; Bederman, I.; Pikuleva, I.A. Studies of ApoD(-/-) and ApoD(-/-)ApoE(-/-) mice uncover the APOD significance for retinal metabolism, function, and status of chorioretinal blood vessels. Cell Mol. Life Sci. 2021, 78, 963–983. [Google Scholar] [CrossRef]
- Lim, W.; Bae, H.; Song, G. Differential expression of apolipoprotein D in male reproductive system of rats by high-fat diet. Andrology 2016, 4, 1115–1122. [Google Scholar] [CrossRef] [PubMed]
- Ganfornina, M.D.; Do Carmo, S.; Lora, J.M.; Torres-Schumann, S.; Vogel, M.; Allhorn, M.; Gonzalez, C.; Bastiani, M.J.; Rassart, E.; Sanchez, D. Apolipoprotein D is involved in the mechanisms regulating protection from oxidative stress. Aging Cell 2008, 7, 506–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Do Carmo, S.; Levros, L.C., Jr.; Rassart, E. Modulation of apolipoprotein D expression and translocation under specific stress conditions. Biochim. Biophys. Acta 2007, 1773, 954–969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Upadhya, R.; Zingg, W.; Shetty, S.; Shetty, A.K. Astrocyte-derived extracellular vesicles: Neuroreparative properties and role in the pathogenesis of neurodegenerative disorders. J. Control. Release 2020, 323, 225–239. [Google Scholar] [CrossRef]
- Kjeldsen, L.; Johnsen, A.H.; Sengelov, H.; Borregaard, N. Isolation and primary structure of NGAL, a novel protein associated with human neutrophil gelatinase. J. Biol. Chem. 1993, 268, 10425–10432. [Google Scholar] [CrossRef]
- Goetz, D.H.; Holmes, M.A.; Borregaard, N.; Bluhm, M.E.; Raymond, K.N.; Strong, R.K. The neutrophil lipocalin NGAL is a bacteriostatic agent that interferes with siderophore-mediated iron acquisition. Mol. Cell 2002, 10, 1033–1043. [Google Scholar] [CrossRef]
- Cavalcante, C.; Cavalcante, M.B.; Castello Branco, K.M.P.; Chan, T.; Maia, I.C.L.; Pompeu, R.G.; de Oliveira Telles, A.C.; Brito, A.K.M.; Liborio, A.B. Biomarkers of acute kidney injury in pediatric cardiac surgery. Pediatr. Nephrol. 2021. [Google Scholar] [CrossRef]
- Cowland, J.B.; Borregaard, N. Molecular characterization and pattern of tissue expression of the gene for neutrophil gelatinase-associated lipocalin from humans. Genomics 1997, 45, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Kjeldsen, L.; Cowland, J.B.; Borregaard, N. Human neutrophil gelatinase-associated lipocalin and homologous proteins in rat and mouse. Biochim. Biophys. Acta 2000, 1482, 272–283. [Google Scholar] [CrossRef]
- Thogersen, M.; Frydland, M.; Lerche Helgestad, O.K.; Okkels Jensen, L.; Josiassen, J.; Goetze, J.P.; Moller, J.E.; Hassager, C. Admission biomarkers among patients with acute myocardial-infarction related cardiogenic shock with or without out-of-hospital cardiac arrest An exploratory study. Biomarkers 2021, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Frydland, M.; Mogelvang, R.; Moller, J.E.; Helgestad, O.K.L.; Holmvang, L.; Mark, P.D.; Jensen, L.O.; Hassager, C. Neutrophil Gelatinase-Associated Lipocalin (NGAL) Measured at Admission is Associated with Development of Late Cardiogenic Shock and Mortality in Patients With ST-Segment Elevation Myocardial Infarction. Shock 2021, 56, 255–259. [Google Scholar] [CrossRef]
- Mostafa, E.A.; Elelwany, S.E.; Shahin, K.M.; El Midany, A.A.H.; Hassaballa, A.S.; El-Sokkary, I.N.; Gamal, M.A.; Elsaid, M.E.; ElBarbary, M.G.; Khorshid, R. Validation of Cardiac Surgery-Associated Neutrophil Gelatinase-Associated Lipocalin Score for Prediction of Cardiac Surgery-Associated Acute Kidney Injury. Heart Lung Circ. 2021. [Google Scholar] [CrossRef]
- Wang, Y.; Lam, K.S.; Kraegen, E.W.; Sweeney, G.; Zhang, J.; Tso, A.W.; Chow, W.S.; Wat, N.M.; Xu, J.Y.; Hoo, R.L.; et al. Lipocalin-2 is an inflammatory marker closely associated with obesity, insulin resistance, and hyperglycemia in humans. Clin. Chem. 2007, 53, 34–41. [Google Scholar] [CrossRef] [Green Version]
- Catalan, V.; Gomez-Ambrosi, J.; Rodriguez, A.; Ramirez, B.; Silva, C.; Rotellar, F.; Gil, M.J.; Cienfuegos, J.A.; Salvador, J.; Fruhbeck, G. Increased adipose tissue expression of lipocalin-2 in obesity is related to inflammation and matrix metalloproteinase-2 and metalloproteinase-9 activities in humans. J. Mol. Med. 2009, 87, 803–813. [Google Scholar] [CrossRef]
- Auguet, T.; Quintero, Y.; Terra, X.; Martinez, S.; Lucas, A.; Pellitero, S.; Aguilar, C.; Hernandez, M.; del Castillo, D.; Richart, C. Upregulation of lipocalin 2 in adipose tissues of severely obese women: Positive relationship with proinflammatory cytokines. Obesity 2011, 19, 2295–2300. [Google Scholar] [CrossRef]
- Takaya, J.; Tanabe, Y.; Kaneko, K. Increased lipocalin 2 levels in adolescents with type 2 diabetes mellitus. J. Pediatr. Endocrinol. Metab. 2021. [Google Scholar] [CrossRef] [PubMed]
- Petropoulou, P.I.; Mosialou, I.; Shikhel, S.; Hao, L.; Panitsas, K.; Bisikirska, B.; Luo, N.; Bahna, F.; Kim, J.; Carberry, P.; et al. Lipocalin-2 is an anorexigenic signal in primates. Elife 2020, 9, e58949. [Google Scholar] [CrossRef] [PubMed]
- Principi, E.; Buschiazzo, A.; Papait, A.; Castagnola, P.; Costa, D.; Pece, R.; Maric, I.; Scussolini, M.; Marini, C.; Sambuceti, G.; et al. Anthropometric and glucometabolic changes in an aged mouse model of lipocalin-2 overexpression. Int. J. Obes. 2019, 43, 189–201. [Google Scholar] [CrossRef]
- Lin, Y.; Rajala, M.W.; Berger, J.P.; Moller, D.E.; Barzilai, N.; Scherer, P.E. Hyperglycemia-induced production of acute phase reactants in adipose tissue. J. Biol. Chem. 2001, 276, 42077–42083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Law, I.K.; Xu, A.; Lam, K.S.; Berger, T.; Mak, T.W.; Vanhoutte, P.M.; Liu, J.T.; Sweeney, G.; Zhou, M.; Yang, B.; et al. Lipocalin-2 deficiency attenuates insulin resistance associated with aging and obesity. Diabetes 2010, 59, 872–882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, H.; Jin, D.; Zhang, Y.; Wright, W.; Bazuine, M.; Brockman, D.A.; Bernlohr, D.A.; Chen, X. Lipocalin-2 deficiency impairs thermogenesis and potentiates diet-induced insulin resistance in mice. Diabetes 2010, 59, 1376–1385. [Google Scholar] [CrossRef] [Green Version]
- Mosialou, I.; Shikhel, S.; Luo, N.; Petropoulou, P.I.; Panitsas, K.; Bisikirska, B.; Rothman, N.J.; Tenta, R.; Cariou, B.; Wargny, M.; et al. Lipocalin-2 counteracts metabolic dysregulation in obesity and diabetes. J. Exp. Med. 2020, 217. [Google Scholar] [CrossRef]
- Deis, J.A.; Guo, H.; Wu, Y.; Liu, C.; Bernlohr, D.A.; Chen, X. Adipose Lipocalin 2 overexpression protects against age-related decline in thermogenic function of adipose tissue and metabolic deterioration. Mol. Metab. 2019, 24, 18–29. [Google Scholar] [CrossRef]
- Ponzetti, M.; Aielli, F.; Ucci, A.; Cappariello, A.; Lombardi, G.; Teti, A.; Rucci, N. Lipocalin 2 increases after high-intensity exercise in humans and influences muscle gene expression and differentiation in mice. J. Cell Physiol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Guo, H.; Deis, J.A.; Mashek, M.G.; Zhao, M.; Ariyakumar, D.; Armien, A.G.; Bernlohr, D.A.; Mashek, D.G.; Chen, X. Lipocalin 2 regulates brown fat activation via a nonadrenergic activation mechanism. J. Biol. Chem. 2014, 289, 22063–22077. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.T.; Huang, Z.; Pan, K.; Zhang, H.J.; Woo, C.W.; Xu, A.; Wong, C.M. Adipose-derived lipocalin 14 alleviates hyperglycaemia by suppressing both adipocyte glycerol efflux and hepatic gluconeogenesis in mice. Diabetologia 2016, 59, 604–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Briand, L.; Eloit, C.; Nespoulous, C.; Bezirard, V.; Huet, J.C.; Henry, C.; Blon, F.; Trotier, D.; Pernollet, J.C. Evidence of an odorant-binding protein in the human olfactory mucus: Location, structural characterization, and odorant-binding properties. Biochemistry 2002, 41, 7241–7252. [Google Scholar] [CrossRef]
- Lacazette, E.; Gachon, A.M.; Pitiot, G. A novel human odorant-binding protein gene family resulting from genomic duplicons at 9q34: Differential expression in the oral and genital spheres. Hum. Mol. Genet. 2000, 9, 289–301. [Google Scholar] [CrossRef] [Green Version]
- Schiefner, A.; Freier, R.; Eichinger, A.; Skerra, A. Crystal structure of the human odorant binding protein, OBPIIa. Proteins 2015, 83, 1180–1184. [Google Scholar] [CrossRef]
- Kobayashi, T.; Kurano, M.; Nanya, M.; Shimizu, T.; Ohkawa, R.; Tozuka, M.; Yatomi, Y. Glycation of HDL Polymerizes Apolipoprotein M and Attenuates Its Capacity to Bind to Sphingosine 1-Phosphate. J. Atheroscler. Thromb. 2021, 28, 730–741. [Google Scholar] [CrossRef] [PubMed]
- Sevvana, M.; Kassler, K.; Ahnstrom, J.; Weiler, S.; Dahlback, B.; Sticht, H.; Muller, Y.A. Mouse ApoM displays an unprecedented seven-stranded lipocalin fold: Folding decoy or alternative native fold? J. Mol. Biol. 2010, 404, 363–371. [Google Scholar] [CrossRef] [Green Version]
- Kurano, M.; Tsukamoto, K.; Shimizu, T.; Kassai, H.; Nakao, K.; Aiba, A.; Hara, M.; Yatomi, Y. Protection Against Insulin Resistance by Apolipoprotein M/Sphingosine-1-Phosphate. Diabetes 2020, 69, 867–881. [Google Scholar] [CrossRef]
- Kurano, M.; Yatomi, Y. Sphingosine 1-Phosphate and Atherosclerosis. J. Atheroscler. Thromb. 2018, 25, 16–26. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Pan, J.M.; Pei, X.; Li, J.S. Interaction Between Apolipoprotein M Gene Single-Nucleotide Polymorphisms and Obesity and its Effect on Type 2 Diabetes Mellitus Susceptibility. Sci. Rep. 2020, 10, 7859. [Google Scholar] [CrossRef]
- Sramkova, V.; Berend, S.; Siklova, M.; Caspar-Bauguil, S.; Carayol, J.; Bonnel, S.; Marques, M.; Decaunes, P.; Kolditz, C.I.; Dahlman, I.; et al. Apolipoprotein M: A novel adipokine decreasing with obesity and upregulated by calorie restriction. Am. J. Clin. Nutr. 2019, 109, 1499–1510. [Google Scholar] [CrossRef] [Green Version]
- Dullaart, R.P.; Plomgaard, P.; de Vries, R.; Dahlback, B.; Nielsen, L.B. Plasma apolipoprotein M is reduced in metabolic syndrome but does not predict intima media thickness. Clin. Chim. Acta 2009, 406, 129–133. [Google Scholar] [CrossRef] [PubMed]
- Frej, C.; Mendez, A.J.; Ruiz, M.; Castillo, M.; Hughes, T.A.; Dahlback, B.; Goldberg, R.B. A Shift in ApoM/S1P Between HDL-Particles in Women With Type 1 Diabetes Mellitus Is Associated With Impaired Anti-Inflammatory Effects of the ApoM/S1P Complex. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 1194–1205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denimal, D.; Pais de Barros, J.P.; Petit, J.M.; Bouillet, B.; Verges, B.; Duvillard, L. Significant abnormalities of the HDL phosphosphingolipidome in type 1 diabetes despite normal HDL cholesterol concentration. Atherosclerosis 2015, 241, 752–760. [Google Scholar] [CrossRef]
- Yu, Y.; Zhang, J.; Yao, S.; Pan, L.; Luo, G.; Xu, N. Apolipoprotein M overexpression through adeno-associated virus gene transfer improves insulin secretion and insulin sensitivity in Goto-Kakizaki rats. J. Diabetes Investig. 2020, 11, 1150–1158. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.; Liu, H.; Liu, H.; Yu, Y.; Zhang, J.; Li, Y.; Luo, G.; Zhang, X.; Xu, N. Increased expression levels of inflammatory cytokines and adhesion molecules in lipopolysaccharideinduced acute inflammatory apoM/ mice. Mol. Med. Rep. 2020, 22, 3117–3126. [Google Scholar] [CrossRef] [PubMed]
- Yao, S.; Luo, G.; Liu, H.; Zhang, J.; Zhan, Y.; Xu, N.; Zhang, X.; Zheng, L. Apolipoprotein M promotes the anti-inflammatory effect of high-density lipoprotein by binding to scavenger receptor BI. Ann. Transl. Med. 2020, 8, 1676. [Google Scholar] [CrossRef] [PubMed]
- Yao, S.; Zheng, F.; Yu, Y.; Zhan, Y.; Xu, N.; Luo, G.; Zheng, L. Apolipoprotein M promotes cholesterol uptake and efflux from mouse macrophages. FEBS Open Bio 2021, 11, 1607–1620. [Google Scholar] [CrossRef]
- Del Gaudio, I.; Rubinelli, L.; Sasset, L.; Wadsack, C.; Hla, T.; Di Lorenzo, A. Endothelial Spns2 and ApoM Regulation of Vascular Tone and Hypertension Via Sphingosine-1-Phosphate. J. Am. Heart Assoc. 2021, 10, e021261. [Google Scholar] [CrossRef]
- Christoffersen, C.; Federspiel, C.K.; Borup, A.; Christensen, P.M.; Madsen, A.N.; Heine, M.; Nielsen, C.H.; Kjaer, A.; Holst, B.; Heeren, J.; et al. The Apolipoprotein M/S1P Axis Controls Triglyceride Metabolism and Brown Fat Activity. Cell Rep. 2018, 22, 175–188. [Google Scholar] [CrossRef] [Green Version]
- Kitada, Y.; Kajita, K.; Taguchi, K.; Mori, I.; Yamauchi, M.; Ikeda, T.; Kawashima, M.; Asano, M.; Kajita, T.; Ishizuka, T.; et al. Blockade of Sphingosine 1-Phosphate Receptor 2 Signaling Attenuates High-Fat Diet-Induced Adipocyte Hypertrophy and Systemic Glucose Intolerance in Mice. Endocrinology 2016, 157, 1839–1851. [Google Scholar] [CrossRef]
- Torabi, R.; Ghourchian, H.; Amanlou, M.; Pasalar, P. Aptamer-Conjugated Calcium Phosphate Nanoparticles for Reducing Diabetes Risk via Retinol Binding Protein 4 Inhibition. Can. J. Diabetes 2017, 41, 305–311. [Google Scholar] [CrossRef]
- Shaheer, A.; Kumar, A.; Menon, P.; Jallo, M.; Basha, S. Effect of Add-On Therapy of Sodium-Glucose Cotransporter 2 Inhibitors and Dipeptidyl Peptidase 4 Inhibitors on Adipokines in Type 2 Diabetes Mellitus. J. Clin. Med. Res. 2021, 13, 355–362. [Google Scholar] [CrossRef]
- Gokulakrishnan, K.; Pandey, G.K.; Sathishkumar, C.; Sundararajan, S.; Durairaj, P.; Manickam, N.; Mohan, V.; Balasubramanyam, M. Augmentation of RBP4/STRA6 signaling leads to insulin resistance and inflammation and the plausible therapeutic role of vildagliptin and metformin. Mol. Biol. Rep. 2021, 48, 4093–4106. [Google Scholar] [CrossRef]
- Furuhashi, M.; Mita, T.; Moniwa, N.; Hoshina, K.; Ishimura, S.; Fuseya, T.; Watanabe, Y.; Yoshida, H.; Shimamoto, K.; Miura, T. Angiotensin II receptor blockers decrease serum concentration of fatty acid-binding protein 4 in patients with hypertension. Hypertens. Res. 2015, 38, 252–259. [Google Scholar] [CrossRef] [PubMed]
- Karpisek, M.; Stejskal, D.; Kotolova, H.; Kollar, P.; Janoutova, G.; Ochmanova, R.; Cizek, L.; Horakova, D.; Yahia, R.B.; Lichnovska, R.; et al. Treatment with atorvastatin reduces serum adipocyte-fatty acid binding protein value in patients with hyperlipidaemia. Eur. J. Clin. Investig. 2007, 37, 637–642. [Google Scholar] [CrossRef] [PubMed]
- Furuhashi, M.; Hiramitsu, S.; Mita, T.; Omori, A.; Fuseya, T.; Ishimura, S.; Watanabe, Y.; Hoshina, K.; Matsumoto, M.; Tanaka, M.; et al. Reduction of circulating FABP4 level by treatment with omega-3 fatty acid ethyl esters. Lipids Health Dis. 2016, 15, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salehpour, A.; Shidfar, F.; Hedayati, M.; Farshad, A.A.; Tehrani, A.N.; Mohammadi, S. Molecular mechanisms of vitamin D plus Bisphenol A effects on adipogenesis in human adipose-derived mesenchymal stem cells. Diabetol. Metab. Syndr. 2021, 13, 41. [Google Scholar] [CrossRef] [PubMed]
- Burak, M.F.; Inouye, K.E.; White, A.; Lee, A.; Tuncman, G.; Calay, E.S.; Sekiya, M.; Tirosh, A.; Eguchi, K.; Birrane, G.; et al. Development of a therapeutic monoclonal antibody that targets secreted fatty acid-binding protein aP2 to treat type 2 diabetes. Sci. Transl. Med. 2015, 7, 319ra205. [Google Scholar] [CrossRef] [PubMed]
- Miao, X.; Wang, Y.; Wang, W.; Lv, X.; Wang, M.; Yin, H. The mAb against adipocyte fatty acid-binding protein 2E4 attenuates the inflammation in the mouse model of high-fat diet-induced obesity via toll-like receptor 4 pathway. Mol. Cell. Endocrinol. 2015, 403, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Sulsky, R.; Magnin, D.R.; Huang, Y.; Simpkins, L.; Taunk, P.; Patel, M.; Zhu, Y.; Stouch, T.R.; Bassolino-Klimas, D.; Parker, R.; et al. Potent and selective biphenyl azole inhibitors of adipocyte fatty acid binding protein (aFABP). Bioorg. Med. Chem. Lett. 2007, 17, 3511–3515. [Google Scholar] [CrossRef] [PubMed]
- Hertzel, A.V.; Hellberg, K.; Reynolds, J.M.; Kruse, A.C.; Juhlmann, B.E.; Smith, A.J.; Sanders, M.A.; Ohlendorf, D.H.; Suttles, J.; Bernlohr, D.A. Identification and characterization of a small molecule inhibitor of Fatty Acid binding proteins. J. Med. Chem. 2009, 52, 6024–6031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Schematic structures of the lipocalin and fatty acid binding protein family (adapted from [4]). (A) Lipocalin structure. The β-strands forming the β-barrel are represented as arrows labeled A–H and the loops as circular lines labeled L1–L7. The short N-terminus 310-helix and the C-terminus α-helix and short β-strand are also represented. Grey background represents hydrogen bonds between strands. Dotted squares surround the three structurally conserved regions of kernel lipocalins. (B) Fatty acid binding protein structure. The β-strands are represented as arrows labeled A–J and the loops as circular lines labeled L1–L9. Two conserved α-helixes in loop L1 are shown as α1 and α2. The N-terminus 310-helix and the C-terminus are also represented. Grey background represents hydrogen bonds between strands.
Figure 1.
Schematic structures of the lipocalin and fatty acid binding protein family (adapted from [4]). (A) Lipocalin structure. The β-strands forming the β-barrel are represented as arrows labeled A–H and the loops as circular lines labeled L1–L7. The short N-terminus 310-helix and the C-terminus α-helix and short β-strand are also represented. Grey background represents hydrogen bonds between strands. Dotted squares surround the three structurally conserved regions of kernel lipocalins. (B) Fatty acid binding protein structure. The β-strands are represented as arrows labeled A–J and the loops as circular lines labeled L1–L9. Two conserved α-helixes in loop L1 are shown as α1 and α2. The N-terminus 310-helix and the C-terminus are also represented. Grey background represents hydrogen bonds between strands.


{kind=link}
Table 1.
Adipose-derived lipid-binding proteins features.
| Gene Symbol/Aliases | Gene Location | Tissue Expression | Ligands | Biological Functions | Review |
|---|---|---|---|---|---|
| RBP4, Retinol binding protein | HSA 10q23.33 MMU 19 | Adipose tissue, liver | Retinol retinoic acid | Vitamin A transport Inflammation Glucose homeostasis | [8,9] |
| FABP4/aP2, A-FABP, ALBP | HSA 8q21.13 MMU 3 | Adipose tissue, immune cells (macrophages) | Fatty acids | Lipogenesis/Lipolysis Inflammation | [10] |
| APOD/GCDFP-24 | HSA 3q29 MMU 16 | Brain, mammary gland, lacrimal gland, pancreas, kidney, heart, adipose tissue, liver, intestine | Arachidonic acid Sphingomyelin Progesterone Pregnenolone Retinoic acid Bilirubin | Immunity/Inflammation Neuronal function Embryonic development Sexual development Antioxidant Cholesterol transport Glucose homeostasis | [11] |
| LCN2/NGAL, siderocalin, murine 24p3 protein, rat α2-microglobulin related protein | HSA 9q34.11 MMU 2 | Kidney, bone marrow, liver, adipose tissue, immune cells, salivary gland, stomach, intestine, trachea, lung, heart, uterus, prostate | Iron, siderophores Leukotrienes Retinoic acid Unsaturated fatty acids | Immunity/Inflammation Renal and cardiac function Glucose homeostasis | [12] |
| OBP2a/2b/LCN14 | HSA 9q34.2 MMU 2 | Nasopharynx, lachrymal gland, prostate, mammary gland, adipose tissue (mouse) | Odorant molecules (e.g., aldehydes, carboxylic acids) | Odors detection Pheromones signaling Glucose homeostasis | - |
| APOM/G3a, NG20 | HSA 6p21.33 MMU 17 | Liver, kidney, adipose tissue, brain (blood–brain barrier) | S1P Saturated fatty acids Retinol, retinoic acid Oxidized phospholipids | Glucose homeostasis Inflammation | [13,14] |
RBP4: Retinol-binding protein 4; FABP4: Fatty acid-binding protein 4; aP2: adipocyte protein 2; A-FABP: adipocyte fatty acid binding protein; ALBP: adipocyte lipid-binding protein; APOD: Apolipoprotein D; GCDFP-24: Gross cystic disease fluid protein 24; LCN2: Lipocalin-2; NGAL: Neutrophil gelatinase-associated lipocalin; LCN14: Lipocalin-14; OBP: Odorant binding protein; APOM: Apolipoprotein M; Chr.: chromosome; S1P: sphingosine-1-phosphate. Regular type indicates adipokines associated to unhealthy phenotype, italics indicate adipokines described as detrimental or beneficial depending on the report, and bold type indicates adipokines reported as associated to healthy phenotype.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Frances, L.; Tavernier, G.; Viguerie, N. Adipose-Derived Lipid-Binding Proteins: The Good, the Bad and the Metabolic Diseases. Int. J. Mol. Sci. 2021, 22, 10460. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910460
AMA Style
Frances L, Tavernier G, Viguerie N. Adipose-Derived Lipid-Binding Proteins: The Good, the Bad and the Metabolic Diseases. International Journal of Molecular Sciences. 2021; 22(19):10460. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910460
Chicago/Turabian StyleFrances, Laurie, Geneviève Tavernier, and Nathalie Viguerie. 2021. "Adipose-Derived Lipid-Binding Proteins: The Good, the Bad and the Metabolic Diseases" International Journal of Molecular Sciences 22, no. 19: 10460. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910460
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.