
Acid Stripping after Infection Improves the Detection of Viral HLA Class I Natural Ligands Identified by Mass Spectrometry


 ,
,
Abstract
:1. Introduction
2. Results
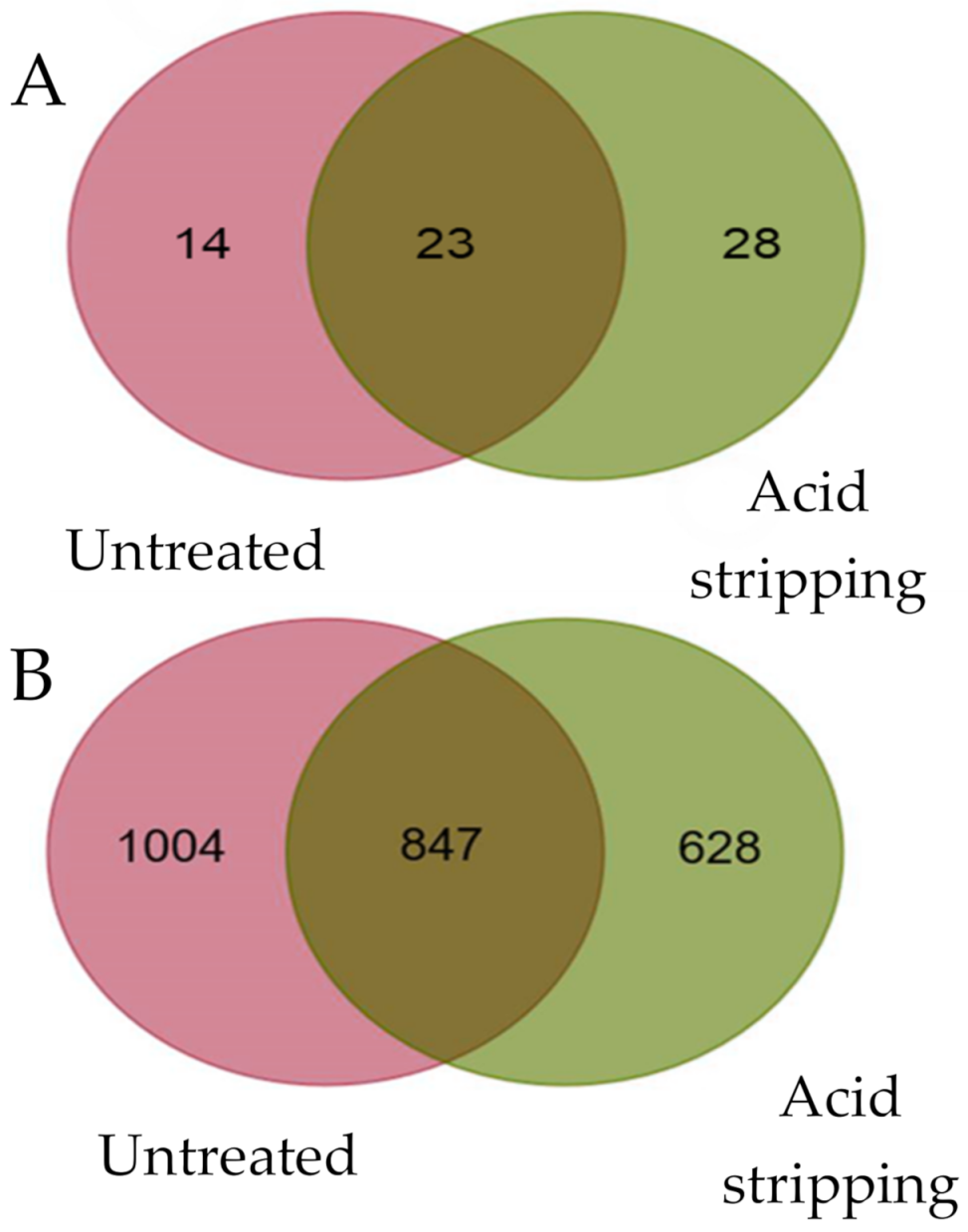
2.1. Physiological Processing Generated Different Viral HLA-B*27:05 Ligands in Human VACV-Infected Cells
2.2. Diverse Viral HLA-B*27:05 Ligands Were Physiological Generated in Human VACV-Infected Cells Treated with Acid Stripping
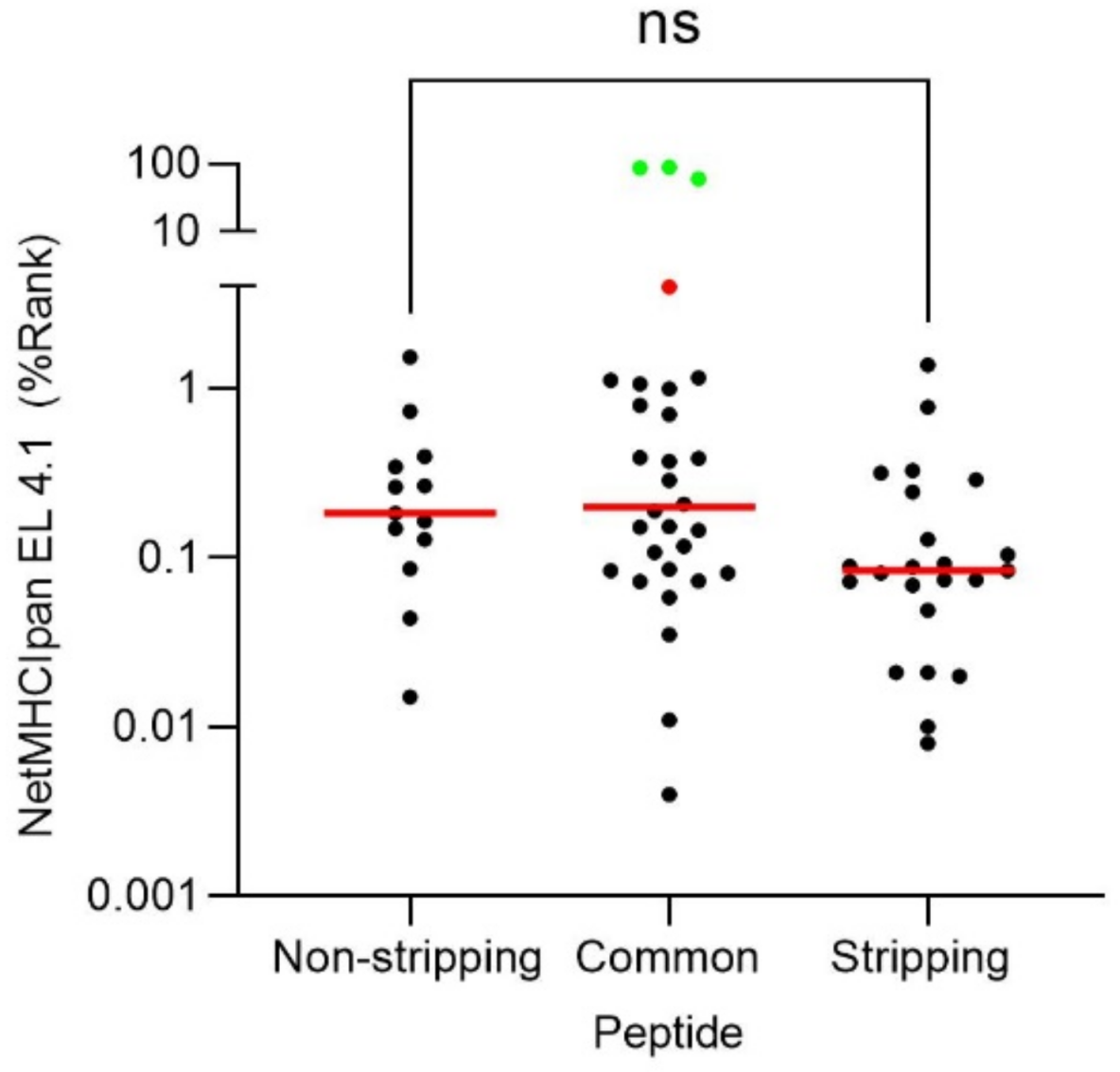
2.3. Most of Viral HLA-B*27:05 Ligands Were Independent of Acid Stripping Treatment
2.4. Theoretical Binding Affinity of the VACV HLA-B*27:05 Ligands
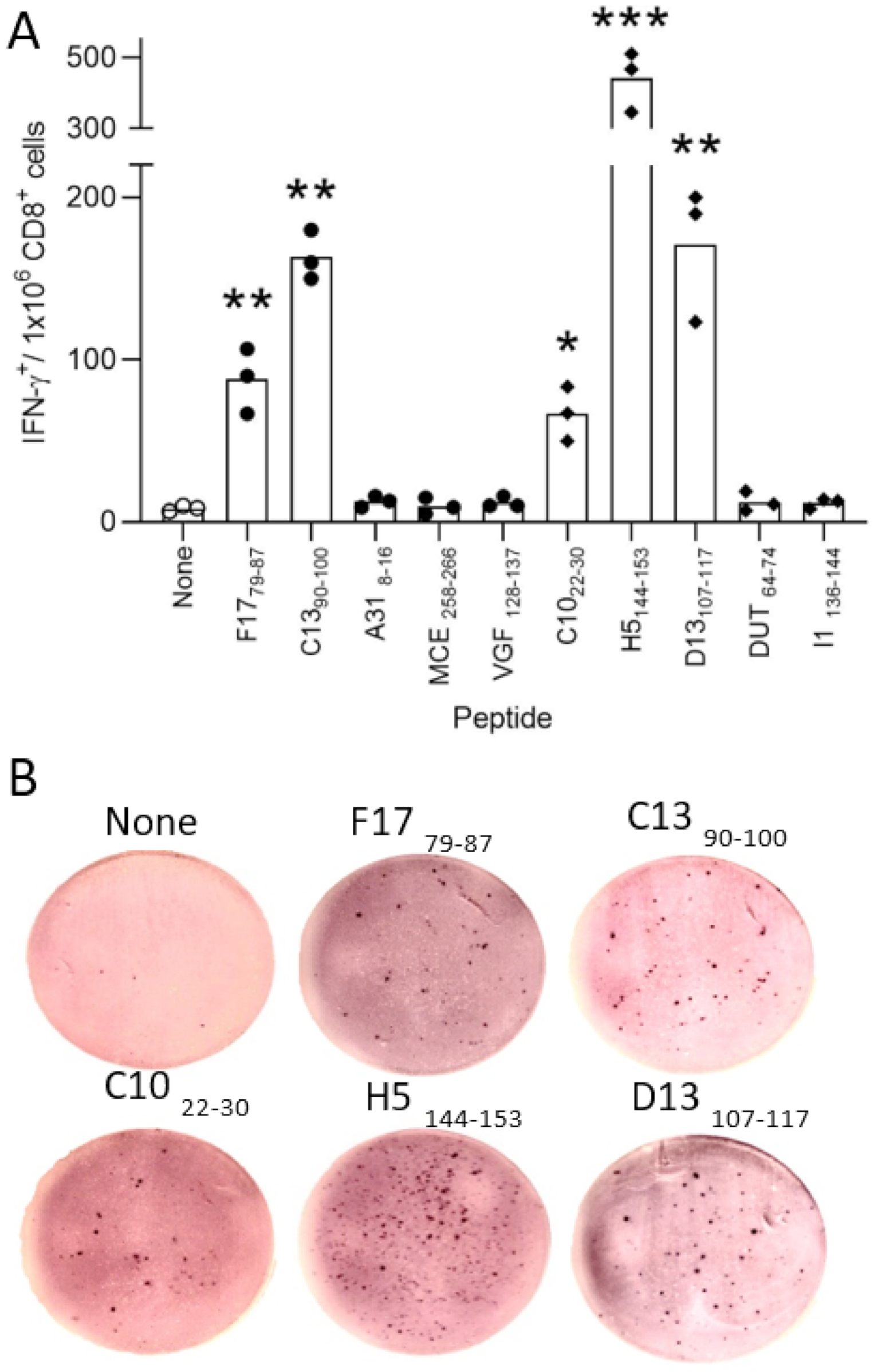
2.5. Immunogenicity of VACV-Derived Peptides Obtained after Acid Stripping in HLA-B*27:05 Transgenic Mice
3. Discussion
4. Materials and Methods
4.1. Cells, Antibodies and Synthetic Peptides
4.2. Mice
4.3. Acid Stripping and HLA Class I Re-Expression
4.4. HLA Class I-Bound Peptide Isolation
4.5. LC-MS/MS Analysis
4.6. MS/MS Ion Search and Peptide Identification
4.7. In Silico Binding Prediction of HLA-B*27:05 Class I Ligands
4.8. IFN-γ-Secreting CD8+ Cell Detection by ELISPOT
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Shastri, N.; Schwab, S.; Serwold, T. Producing nature’s gene-chips: The generation of peptides for display by MHC class I molecules. Annu. Rev. Immunol. 2002, 20, 463–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bjorkman, P.J.; Saper, M.A.; Samraoui, B.; Bennett, W.S.; Strominger, J.L.; Wiley, D.C. Structure of the human class I histocompatibility antigen, HLA-A2. Nature 1987, 329, 506–512. [Google Scholar] [CrossRef]
- Parker, K.C.; Bednarek, M.A.; Coligan, J.E. Scheme for ranking potential HLA-A2 binding peptides based on independent binding of individual peptide side-chains. J. Immunol. 1994, 152, 163–175. [Google Scholar] [PubMed]
- Rammensee, H.G.; Bachmann, J.; Emmerich, N.P.N.; Bachor, O.A.; Stevanovic, S. SYFPEITHI: Database for MHC ligands and peptide motifs. Immunogenetics 1999, 50, 213–219. [Google Scholar] [CrossRef]
- Saric, T.; Chang, S.C.; Hattori, A.; York, I.A.; Markant, S.; Rock, K.L.; Tsujimoto, M.; Goldberg, A.L. An IFN-γ-induced aminopeptidase in the ER, ERAP1, trims precursors to MHC class I-presented peptides. Nat. Immunol. 2002, 3, 1169–1176. [Google Scholar] [CrossRef] [PubMed]
- Tanioka, T.; Hattori, A.; Masuda, S.; Nomura, Y.; Nakayama, H.; Mizutani, S.; Tsujimoto, M. Human leukocyte-derived arginine aminopeptidase—The third member of the oxytocinase subfamily of aminopeptidases. J. Biol. Chem. 2003, 278, 32275–32283. [Google Scholar] [CrossRef] [Green Version]
- Hattori, A.; Matsumoto, H.; Mizutani, S.; Tsujimoto, M. Molecular cloning of adipocyte-derived leucine aminopeptidase highly related to placental leucine aminopeptidase/oxytocinase. J. Biochem. 1999, 125, 931–938. [Google Scholar] [CrossRef] [PubMed]
- York, I.A.; Goldberg, A.L.; Mo, X.Y.; Rock, K.L. Proteolysis and class I major histocompatibility complex antigen presentation. Immunol. Rev. 1999, 172, 49–66. [Google Scholar] [CrossRef]
- Ovsyannikova, I.G.; Johnson, K.L.; Bergen, H.R., III; Poland, G.A. Mass spectrometry and peptide-based vaccine development. Clin. Pharmacol. Ther. 2007, 82, 644–652. [Google Scholar] [CrossRef]
- Le Bouteiller, P.P.; Mishal, Z.; Lemonnier, F.A.; Kourilsky, F.M. Quantification by flow cytofluorimetry of HLA class I molecules at the surface of murine cells transformed by cloned HLA genes. J. Immunol. Methods 1983, 61, 301–315. [Google Scholar] [CrossRef]
- Vanderburg, S.H.; Ras, E.; Drijfhout, J.W.; Benckhuijsen, W.E.; Bremers, A.J.A.; Melief, C.J.M.; Kast, W.M. An HLA class I peptide-binding assay based on competition for binding to class I molecules on intact human B cells—Identification of conserved HIV-1 polymerase peptides binding to HLA-A*0301. Hum. Immunol. 1995, 44, 189–198. [Google Scholar] [CrossRef]
- Schulze zur, W.J.; Lauer, G.M.; Day, C.L.; Kim, A.Y.; Ouchi, K.; Duncan, J.E.; Wurcel, A.G.; Timm, J.; Jones, A.M.; Mothe, B.; et al. Broad repertoire of the CD4+ Th cell response in spontaneously controlled hepatitis C virus infection includes dominant and highly promiscuous epitopes. J. Immunol. 2005, 175, 3603–3613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thaker, S.K.; Chng, J.; Christofk, H.R. Viral hijacking of cellular metabolism. BMC Biol. 2019, 17, 59. [Google Scholar] [CrossRef] [PubMed]
- Ternette, N.; Block, P.D.; Sanchez-Bernabeu, A.; Borthwick, N.; Pappalardo, E.; Abdul-Jawad, S.; Ondondo, B.; Charles, P.D.; Dorrell, L.; Kessler, B.M.; et al. Early Kinetics of the HLA Class I-Associated Peptidome of MVA.HIVconsv-Infected Cells. J. Virol. 2015, 89, 5760–5771. [Google Scholar] [CrossRef] [Green Version]
- Gilchuk, P.; Spencer, C.T.; Conant, S.B.; Hill, T.; Gray, J.J.; Niu, X.; Zheng, M.; Erickson, J.J.; Boyd, K.L.; McAfee, K.J.; et al. Discovering naturally processed antigenic determinants that confer protective T cell immunity. J. Clin. Investig. 2013, 123, 1976–1987. [Google Scholar] [CrossRef] [Green Version]
- Johnson, K.L.; Ovsyannikova, I.G.; Mason, C.J.; Bergen, H.R., III; Poland, G.A. Discovery of naturally processed and HLA-presented class I peptides from vaccinia virus infection using mass spectrometry for vaccine development. Vaccine 2009, 28, 38–47. [Google Scholar] [CrossRef] [Green Version]
- Lorente, E.; Martin-Galiano, A.J.; Barnea, E.; Barriga, A.; Palomo, C.; Garcia-Arriaza, J.; Mir, C.; Lauzurica, P.; Esteban, M.; Admon, A.; et al. Proteomics analysis reveals that structural proteins of the virion core and involved in gene expression are the main source for HLA class II ligands in vaccinia virus-infected cells. J. Proteom. Res. 2019, 18, 900–911. [Google Scholar] [CrossRef]
- Mestas, J.; Hughes, C.C. Of mice and not men: Differences between mouse and human immunology. J. Immunol. 2004, 172, 2731–2738. [Google Scholar] [CrossRef] [Green Version]
- Neefjes, J.; Gottfried, E.; Roelse, J.; Grommé, M.; Obst, R.; Hämmerling, G.J.; Momburg, F. Analysis of the fine specificity of rat, mouse and human TAP peptide transporters. Eur. J. Immunol. 1995, 25, 1133–1136. [Google Scholar] [CrossRef]
- Alvarez, I.; Sesma, L.; Marcilla, M.; Ramos, M.; Marti, M.; Camafeita, E.; de Castro, J.A. Identification of novel HLA-B27 ligands derived from polymorphic regions of its own or other class I molecules based on direct generation by 20 S proteasome. J. Biol. Chem. 2001, 276, 32729–32737. [Google Scholar] [CrossRef] [Green Version]
- Ben Dror, L.; Barnea, E.; Beer, I.; Mann, M.; Admon, A. The HLA-B*2705 peptidome. Arthritis Rheum. 2010, 62, 420–429. [Google Scholar] [PubMed]
- Infantes, S.; Lorente, E.; Barnea, E.; Beer, I.; Barriga, A.; Lasala, F.; Jimenez, M.; Admon, A.; Lopez, D. Natural HLA-B*2705 protein ligands with glutamine as anchor motif: Implications for HLA-B27 association with spondyloarthropathy. J. Biol. Chem. 2013, 288, 10882–10889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samino, Y.; López, D.; Guil, S.; Saveanu, L.; van Endert, P.M.; Del Val, M. A long N-terminal-extended nested set of abundant and antigenic major histocompatibility complex class I natural ligands from HIV envelope protein. J. Biol. Chem. 2006, 281, 6358–6365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calvo, V.; Rojo, S.; López, D.; Galocha, B.; Lopez de Castro, J.A. Structure and diversity of HLA-B27-specific T cell epitopes. Analysis with site-directed mutants mimicking HLA-B27 subtype polymorphism. J. Immunol. 1990, 144, 4038–4045. [Google Scholar]
- Barnstable, C.J.; Bodmer, W.F.; Brown, G.; Galfre, G.; Milstein, C.; Williams, A.F.; Ziegler, A. Production of monoclonal antibodies to group A erythrocytes, HLA and other human cell surface antigens-new tools for genetic analysis. Cell 1978, 14, 9–20. [Google Scholar] [CrossRef]
- Khare, S.D.; Luthra, H.S.; David, C.S. Spontaneous inflammatory arthritis in HLA-B27 transgenic mice lacking beta 2-microglobulin: A model of human spondyloarthropathies. J. Exp. Med. 1995, 182, 1153–1158. [Google Scholar] [CrossRef] [Green Version]
- Luckey, C.J.; Marto, J.A.; Partridge, M.; Hall, E.; White, F.M.; Lippolis, J.D.; Shabanowitz, J.; Hunt, D.F.; Engelhard, V.H. Differences in the expression of human class I MHC alleles and their associated peptides in the presence of proteasome inhibitors. J. Immunol. 2001, 167, 1212–1221. [Google Scholar] [CrossRef] [Green Version]
- Infantes, S.; Lorente, E.; Barnea, E.; Beer, I.; Cragnolini, J.J.; García, R.; Lasala, F.; Jiménez, M.; Admon, A.; López, D. Multiple, non-conserved, internal viral ligands naturally presented by HLA-B27 in human respiratory syncytial virus-infected cells. Mol. Cell. Proteom. 2010, 9, 1533–1539. [Google Scholar] [CrossRef] [Green Version]
- Johnstone, C.; Lorente, E.; Barriga, A.; Barnea, E.; Infantes, S.; Lemonnier, F.A.; David, C.S.; Admon, A.; Lopez, D. The Viral Transcription Group Determines the HLA Class I Cellular Immune Response Against Human Respiratory Syncytial Virus. Mol. Cell Proteom. 2015, 14, 893–904. [Google Scholar] [CrossRef] [Green Version]
- Tapia, E.; Perez-Jimenez, E.; Lopez-Fuertes, L.; Gonzalo, R.; Gherardi, M.M.; Esteban, M. The combination of DNA vectors expressing IL-12 + IL-18 elicits high protective immune response against cutaneous leishmaniasis after priming with DNA-p36/LACK and the cytokines, followed by a booster with a vaccinia virus recombinant expressing p36/LACK. Microbes Infect. 2003, 5, 73–84. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Peptide | Experimental Mass | Length | Protein | Position | Accession | Untreated Cells a | Stripping Condition a |
|---|---|---|---|---|---|---|---|
| GRYIVKLLI | 1073.696 | 9 | 3BHS | 14–22 | P26670 | 1 | 1 |
| KRNDEVLFR | 1175.6411 | 9 | A27 | 99–107 | P11258 | ND | 2 |
| LRFLEKTSF b | 1139.6338 | 9 | A31 | 8–16 | P24760 | 2 | ND |
| YRHVPILGR | 1109.6458 | 9 | A32 | 206–214 | P68615 | ND | 2 |
| GRIDNINMSI | 1147.5656 | 10 | A37 | 254–263 | P24762 | ND | 2 |
| GRLFNEIKKF | 1250.7135 | 10 | A52 | 77–86 | Q01220 | ND | 2 |
| YRFLVINR | 1079.624 | 8 | B1 | 103–110 | P16913 | 1 | 1 |
| YRFLVINRL | 1192.708 | 9 | B1 | 103–111 | P16913 | 1 | 1 |
| RRGDLETLGY | 1178.6044 | 10 | B1 | 210–219 | P16913 | ND | 2 |
| IRWLGGILPWTK | 1438.8448 | 12 | B1 | 229–240 | P16913 | 2 | 2 |
| VRIKNDIVVSR | 1297.783 | 11 | B18 | 232–242 | P25213 | 2 | 3 |
| TRFNPSVLK | 1060.6029 | 9 | B25 | 111–119 | Q805H0 | 3 | 3 |
| TRFNPSVLKILLH | 1536.9139 | 13 | B25 | 111–123 | Q805H0 | 3 | 3 |
| IRNDIRELF | 1174.6458 | 9 | C10 | 22–30 | P03296 | ND | 2 |
| TRFYFNMPK | 1202.5906 | 9 | C10 | 323–331 | P03296 | 3 | 3 |
| SRFTIGRALF | 1166.656 | 10 | C13 | 90–99 | P17365 | 1 | 2 |
| SRFTIGRALFK | 1294.751 | 11 | C13 | 90–100 | P17365 | 2 | ND |
| ARFDNKSIYR | 1268.6625 | 10 | D2 | 63–72 | P04300 | ND | 3 |
| ARFDNKSIYRI | 1381.7466 | 11 | D2 | 63–73 | P04300 | 1 | ND |
| FRVSTKLLRF | 1265.7607 | 10 | D9 | 69–78 | P04311 | ND | 2 |
| GRFGYVPYVGY | 1276.624 | 11 | D13 | 107–117 | P68440 | ND | 2 |
| GRIAPRSGL | 925.5457 | 9 | DUT | 64–72 | P17374 | ND | 2 |
| GRIAPRSGLSL | 1125.6617 | 11 | DUT | 64–74 | P17374 | ND | 2 |
| SRNPSKMVY | 1096.5336 | 9 | E5 | 130–138 | P21606 | ND | 3 |
| IRILVEERF | 1173.6869 | 9 | E5 | 179–187 | P21606 | 3 | 3 |
| IRILVEERFY | 1336.7502 | 10 | E5 | 179–188 | P21606 | 3 | 3 |
| WRIIGTQVDK | 1214.6771 | 10 | E5 | 197–206 | P21606 | 3 | 3 |
| ARYNLKPMYR | 1310.6918 | 10 | E5 | 219–228 | P21606 | 2 | 2 |
| YRYDDDVENGF | 1391.563 | 11 | E5 | 274–284 | P21606 | 3 | 3 |
| YRYDDDVENGFIGL | 1674.7526 | 14 | E5 | 274–287 | P21606 | 3 | 2 |
| KRFDELDINNSY | 1512.7208 | 12 | E6 | 372–383 | P21607 | ND | 2 |
| SRGLSRPLM | 1015.5597 | 9 | E8 | 27–35 | P23372 | 2 | ND |
| VRTIIDENR | 1114.6095 | 9 | ETF1 | 31–39 | P04308 | ND | 2 |
| SRFKKVYIL | 1152.7019 | 9 | ETF1 | 63–71 | P04308 | ND | 1 |
| GRSIRKFSY | 1112.609 | 9 | ETF1 | 453–461 | P04308 | ND | 2 |
| YRQQLELAY | 1182.6033 | 9 | F11 | 247–255 | Q80HX7 | 2 | ND |
| TRTIILVGY | 1034.6124 | 9 | F12 | 275–283 | Q80HX6 | 3 | 3 |
| MRTDMLQNM | 1138.4934 | 9 | F17 | 79–87 | P07396 | 2 | ND |
| AANRDNVASRLLN | 1412.7484 | 13 | F17 | 89–101 | P07396 | ND | 2 |
| FAANRDNVASRLLN | 1559.8168 | 14 | F17 | 88–101 | P07396 | ND | 1 |
| ARVEFGPLYM | 1181.5903 | 10 | F7 | 57–66 | P24359 | 2 | ND |
| YRHTIESVYF | 1313.6404 | 10 | F7 | 83–92 | P24359 | ND | 3 |
| KRFTHTTAFF | 1254.6509 | 10 | G7 | 99–108 | P68716 | ND | 3 |
| PVIDRLPSETFPNVH | 1719.8944 | 15 | H3 | 12–26 | P07240 | 3 | ND |
| ARIENEMKINR | 1372.7245 | 11 | H3 | 217–227 | P07240 | 2 | ND |
| KRYPGVMYAF | 1230.6219 | 10 | H3 | 266–275 | P07240 | ND | 2 |
| ARSDLSDLKV | 1102.5981 | 10 | H5 | 144–153 | P07242 | ND | 2 |
| VRIPVDLVK | 1037.6597 | 9 | I1 | 51–59 | P16714 | 2 | 3 |
| KRSATQFNF | 1097.5618 | 9 | I1 | 136–144 | P16714 | ND | 3 |
| TRLYDYFTR | 1233.6141 | 9 | I1 | 210–218 | P16714 | ND | 3 |
| SQFDDKGNTAL | 1194.5516 | 11 | K1 | 88–98 | P04297 | 1 | 1 |
| VRNKVVVNF | 1073.6345 | 9 | MCE | 258–266 | P07617 | 2 | ND |
| GRVPSVNEY | 1019.5036 | 9 | PAP1 | 23–31 | P23371 | ND | 2 |
| ARDEPVFVK | 1059.5713 | 9 | RAP94 | 292–300 | P68438 | ND | 3 |
| GRLPLVSEF | 1016.5654 | 9 | RP19 | 96–104 | P68611 | 3 | 3 |
| ARDPYAVINR | 1173.6254 | 10 | RP22 | 20–29 | P68609 | 1 | 3 |
| HRFDMTKVDVELFIK | 1876.9869 | 15 | RP35 | 275–289 | P24757 | 1 | 1 |
| SRVSLEFIR | 1105.6244 | 9 | RP132 | 241–249 | Q76ZP7 | 2 | 1 |
| SRVSLEFIRR | 1261.7255 | 10 | RP132 | 241–250 | Q76ZP7 | ND | 2 |
| KRITESITDF | 1208.64 | 10 | RP132 | 272–281 | Q76ZP7 | 1 | 2 |
| GRYSAVFKDSFLR | 1544.8099 | 13 | SPI2 | 70–82 | P15059 | ND | 2 |
| RRTKLPIQDM | 1256.7023 | 10 | VGF | 128–137 | P01136 | 2 | ND |
| ARTIFNFHLI | 1230.6873 | 10 | VITF3 | 16–25 | Q80HV2 | 3 | ND |
| SRTVEIFER | 1135.5985 | 9 | VITF3 | 106–114 | Q80HV2 | 3 | ND |
| IRIKIDKLR | 1153.7659 | 9 | VLTF1 | 18–26 | P68613 | 1 | ND |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lorente, E.; Marcilla, M.; de la Sota, P.G.; Quijada-Freire, A.; Mir, C.; López, D. Acid Stripping after Infection Improves the Detection of Viral HLA Class I Natural Ligands Identified by Mass Spectrometry. Int. J. Mol. Sci. 2021, 22, 10503. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910503
Lorente E, Marcilla M, de la Sota PG, Quijada-Freire A, Mir C, López D. Acid Stripping after Infection Improves the Detection of Viral HLA Class I Natural Ligands Identified by Mass Spectrometry. International Journal of Molecular Sciences. 2021; 22(19):10503. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910503
Chicago/Turabian StyleLorente, Elena, Miguel Marcilla, Patricia G. de la Sota, Adriana Quijada-Freire, Carmen Mir, and Daniel López. 2021. "Acid Stripping after Infection Improves the Detection of Viral HLA Class I Natural Ligands Identified by Mass Spectrometry" International Journal of Molecular Sciences 22, no. 19: 10503. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910503