
Integrin β1 Promotes Pancreatic Tumor Growth by Upregulating Kindlin-2 and TGF-β Receptor-2

Abstract
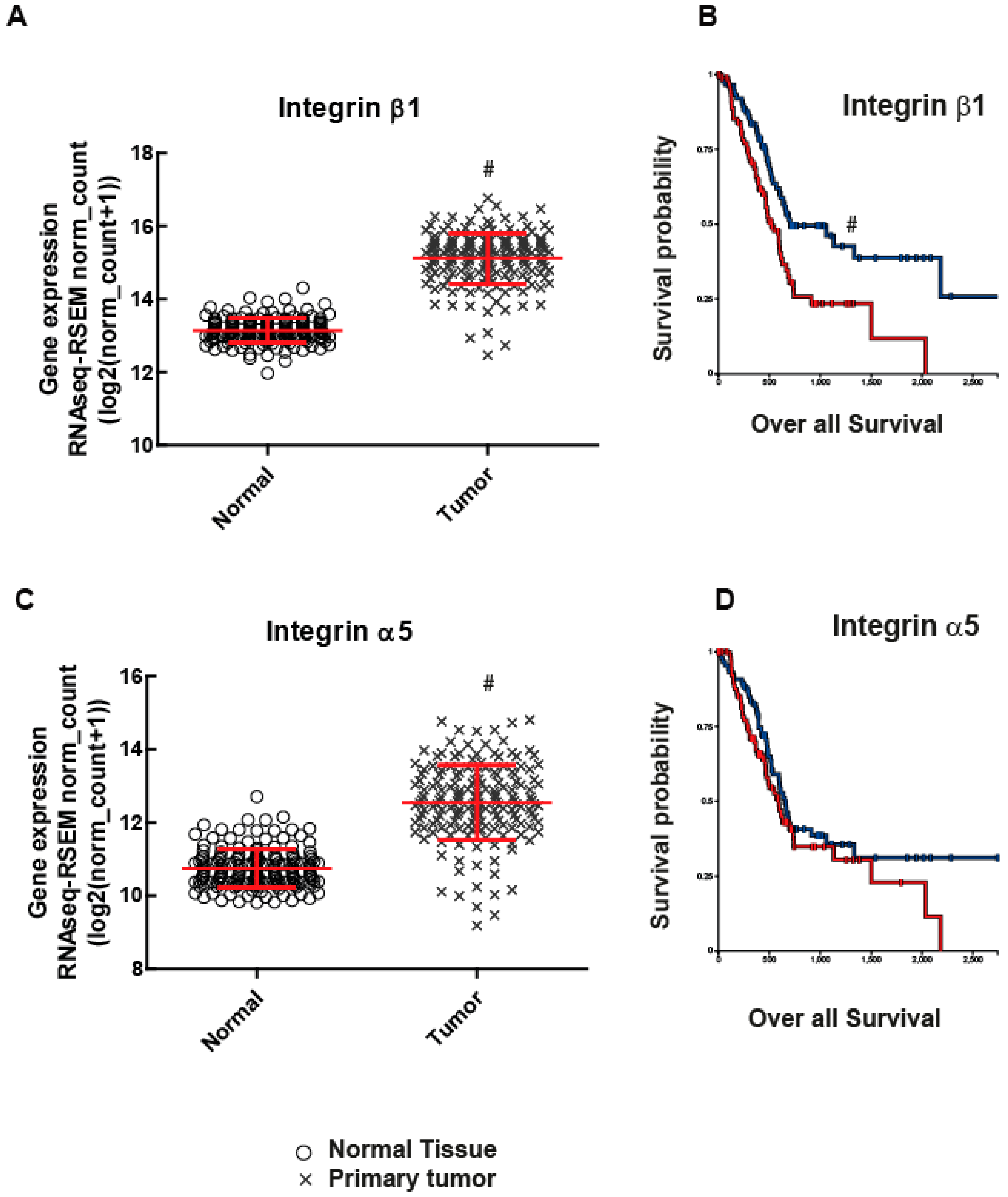
:1. Introduction
2. Results
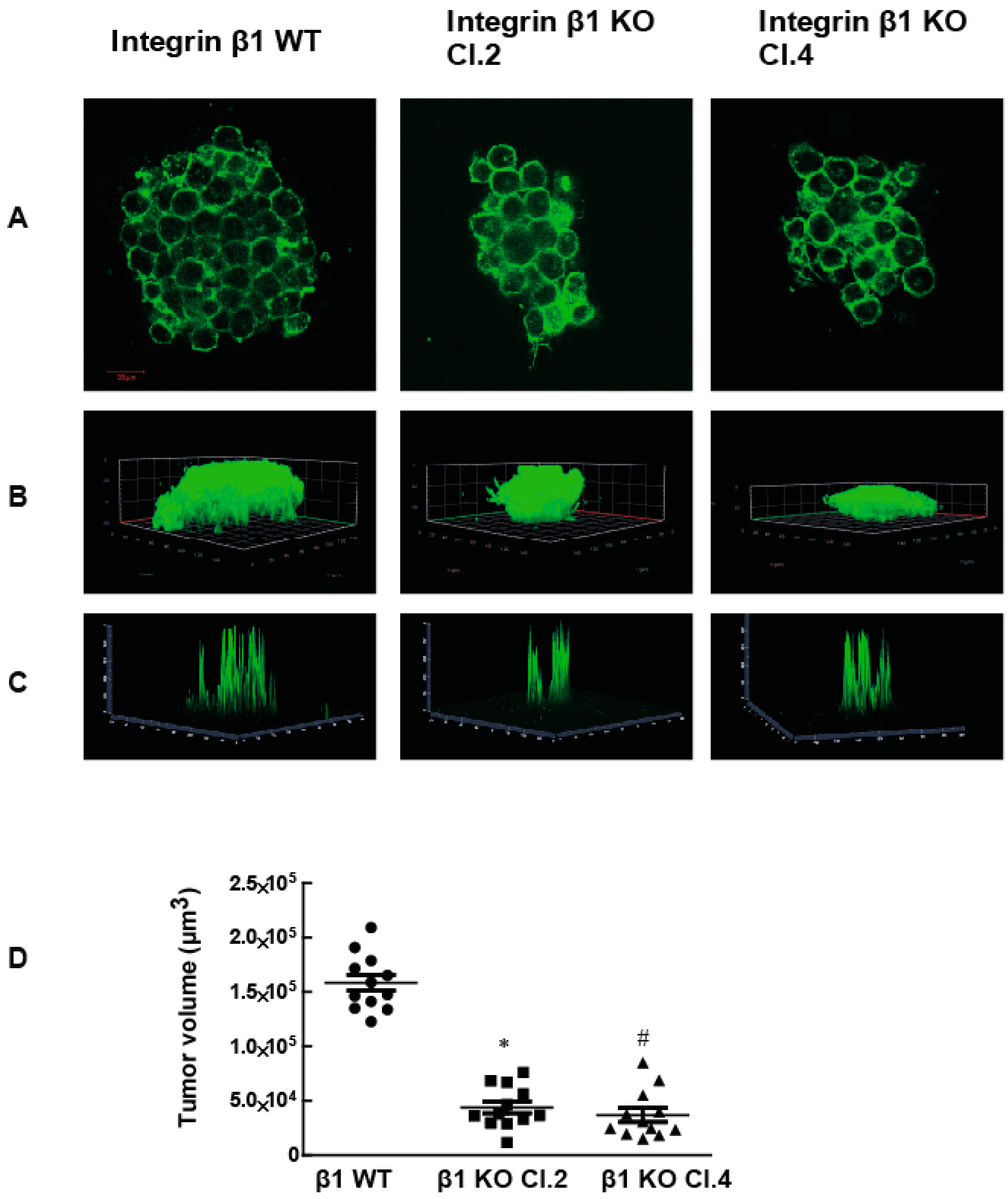
2.1. Integrin β1 Promotes Pancreatic Tumor Growth
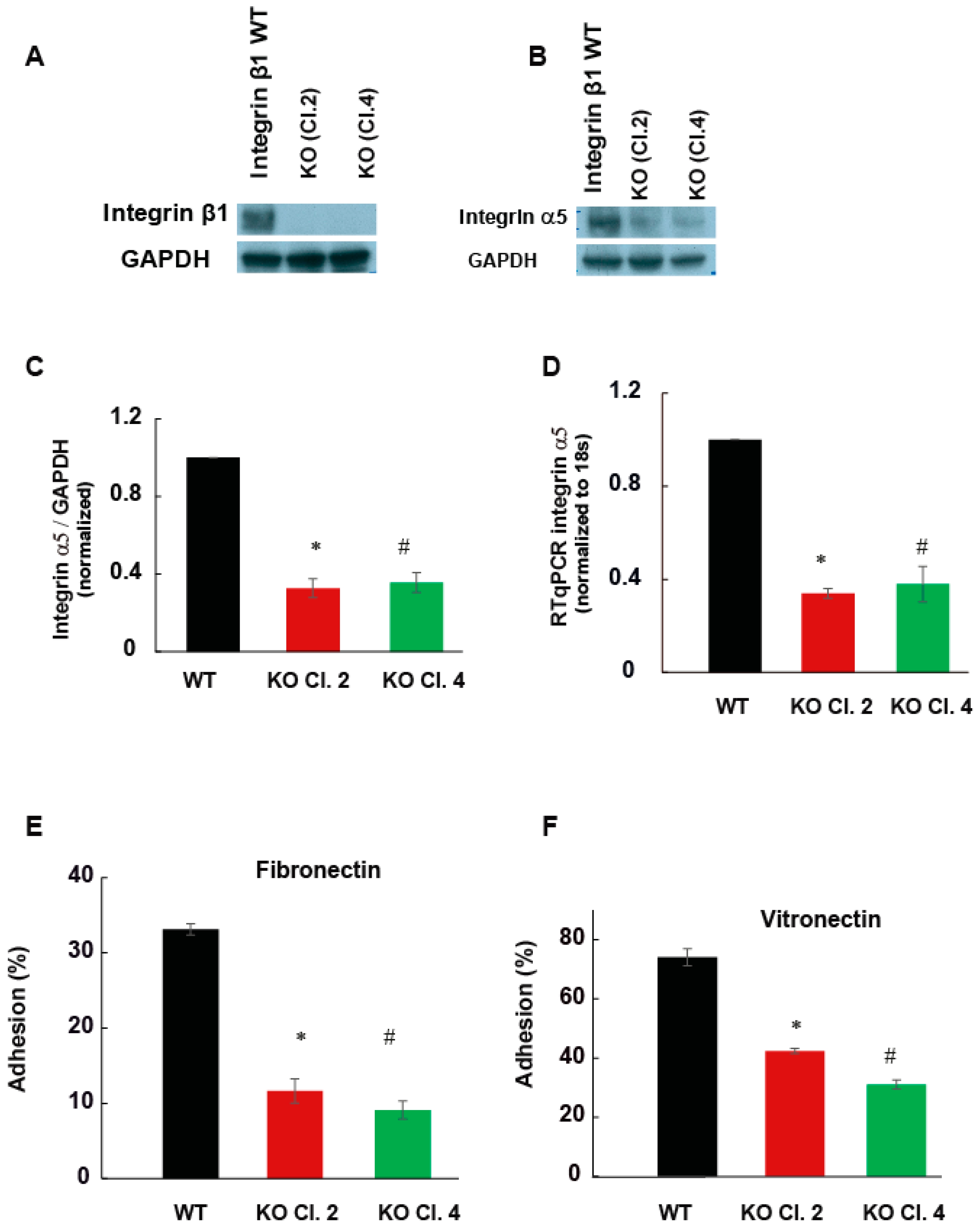
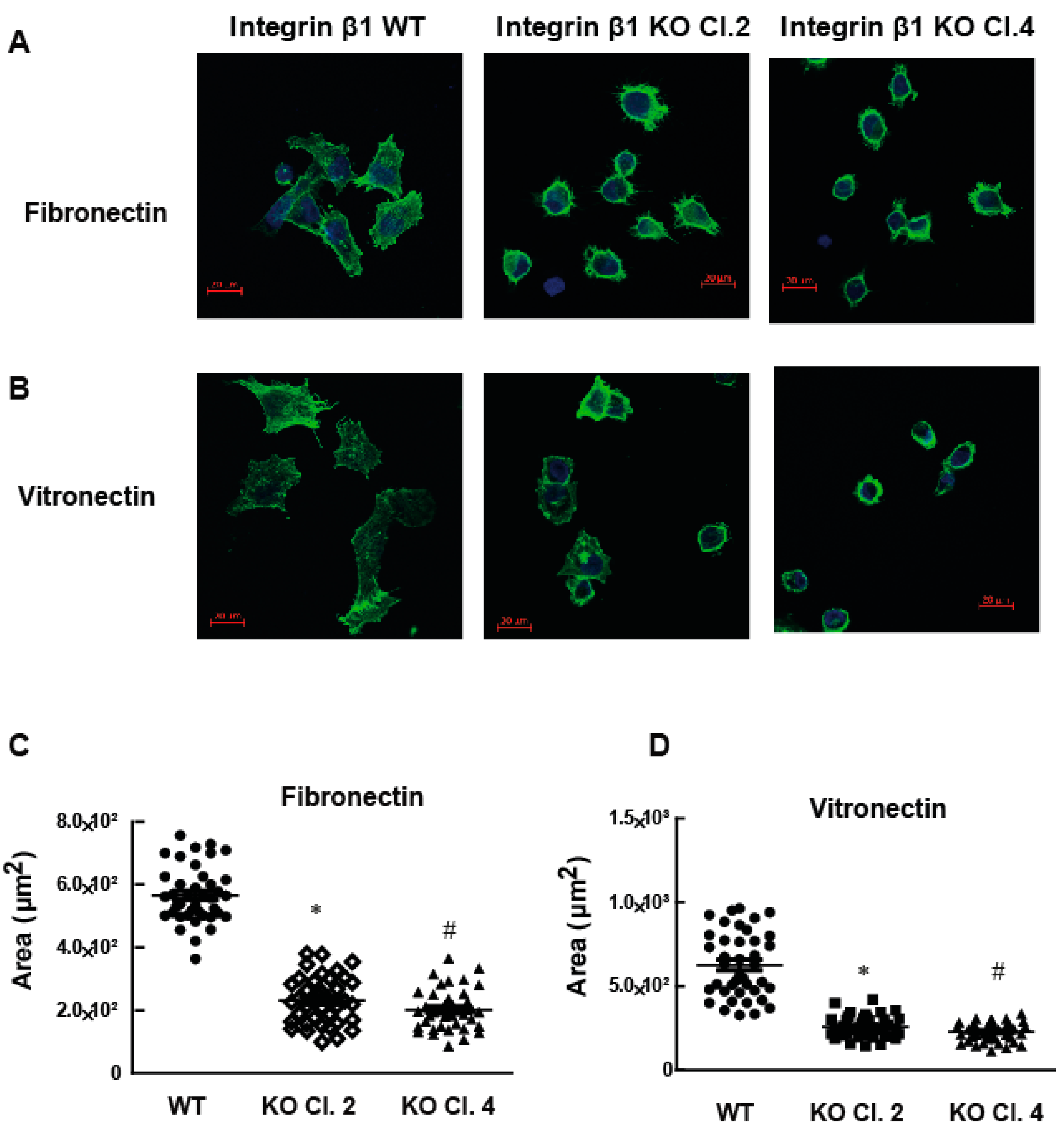
2.2. Adhesion of MIA PaCa-2 Cells to Vitronectin and Fibronectin Is Decreased by the Deletion of Integrin β1
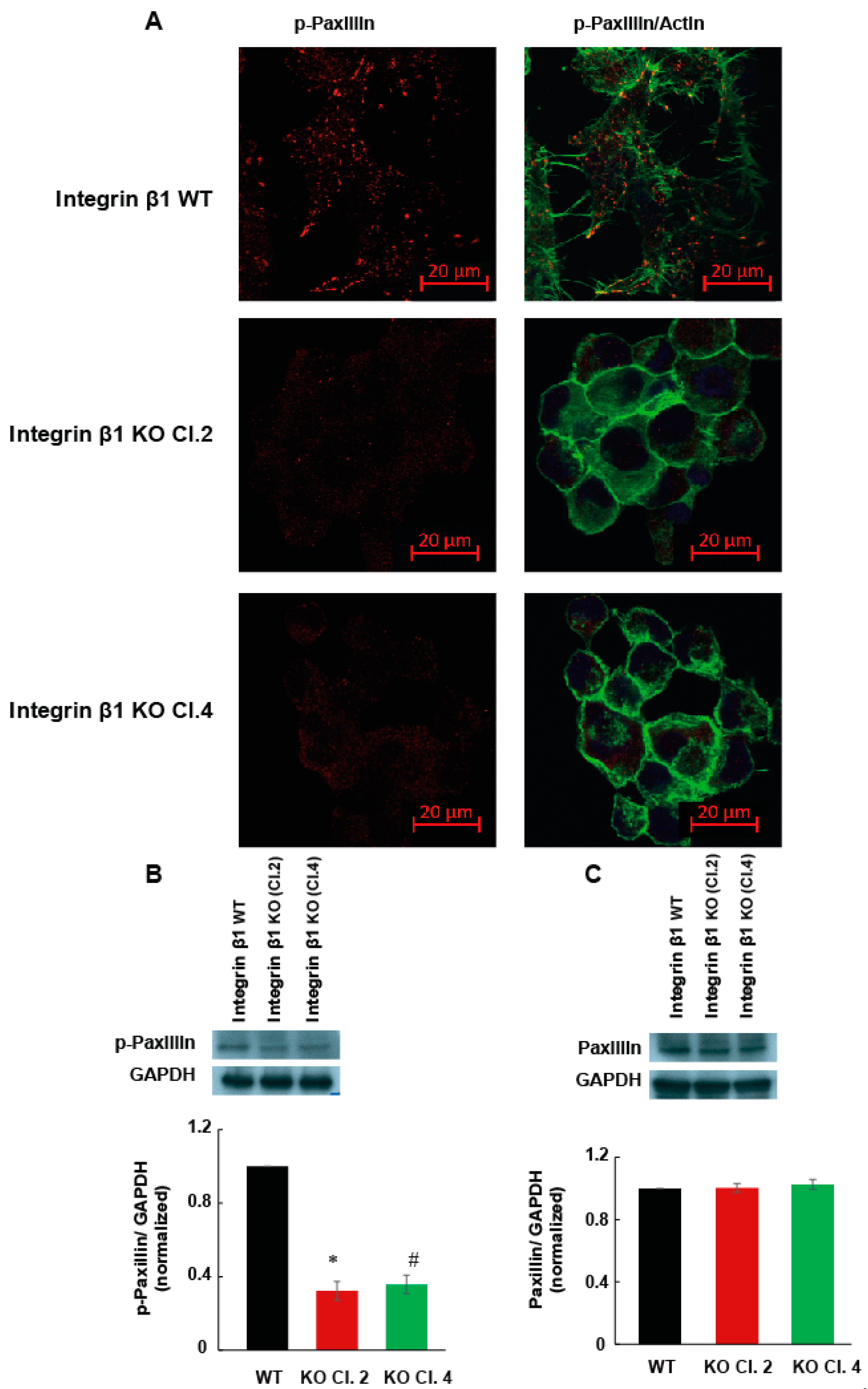
2.3. Integrin β1 Deletion Impaired the Cell Spreading and Focal Adhesions of MIA PaCa-2 Cells
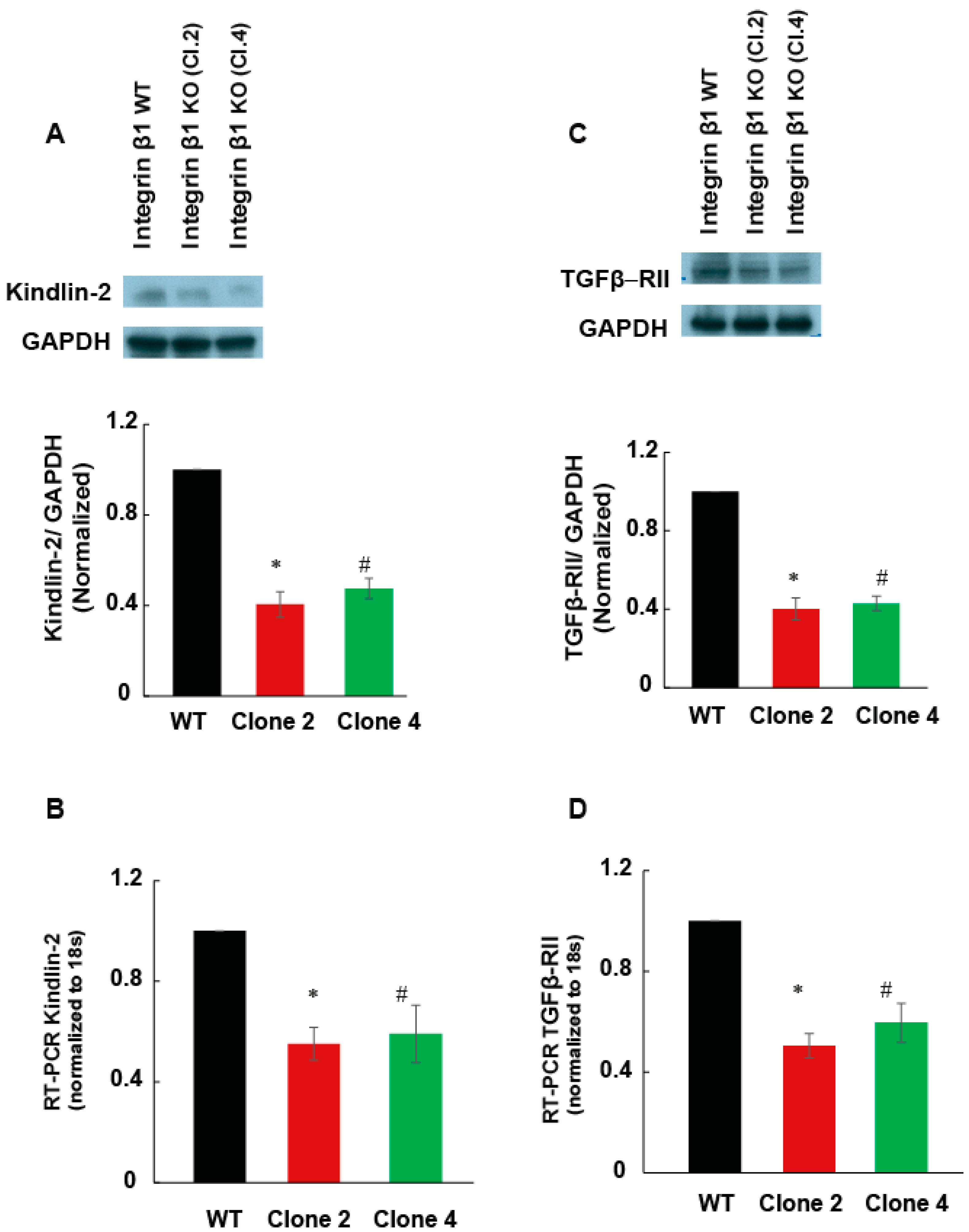
2.4. Integrin β1 Regulates the Expression of Kindlin-2
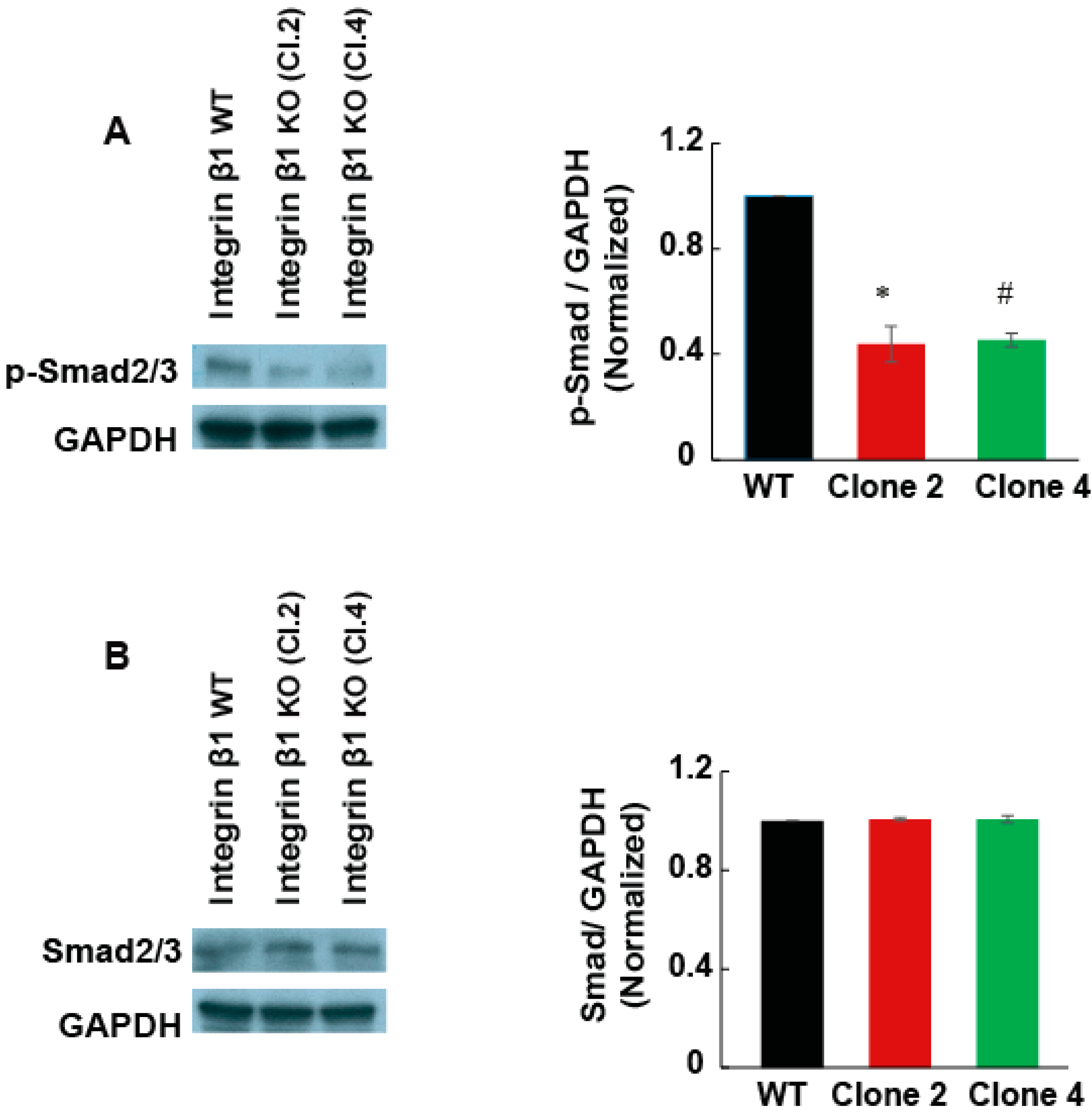
2.5. Integrin β1 Regulates the Expression of TGF-β Receptor 2 and TGF-β Cell Signaling in MIA PaCa-2 Cells
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Cell Culture
4.3. Generation of Knockout Cells
4.4. Immunoblotting
4.5. RNA Isolation and RT PCR
4.6. Immunocytochemistry for Focal Adhesion Assay
4.7. Cell Spreading Assay
4.8. Cell Adhesion Assay
4.9. 3D Matrix Cell Proliferation and Colony Growth
- // ImageJ Macro Code
- // Measure Volume of Thresholded Pixels in an Image Stack
- //
- //
- macro "Measure Stack" {
- run("Clear Results"); // First, clear the results table
- // loop through each slice in the stack. Start at n=1 (the first slice),
- // keep going while n <= nSlices (nSlices is the total number of slices in the stack)
- // and increment n by one after each loop (n++)
- for (n=1; n<=nSlices; n++) {
- setSlice(n); // set the stack’s current slice to n
- run("Measure"); // Run the "Measure" function in ImageJ
- }
- // Create a variable that we will use to store the area measured in each slice
- totalArea = 0;
- // Loop through each result from 0 (the first result on the table) to nResult (the total number of results on the table)
- for (n=0; n < nResults; n++)
- {
- totalArea += getResult("Area",n); // Add the area of the current result to the total
- }
- // Get the calibration information from ImageJ and store into width, height, depth, and unit variables.
- // We will only be using depth and unit
- getVoxelSize (width, height, depth, unit);
- // Calculate the volume by multiplying the sum of area of each slice by the depth
- volume = totalArea*depth;
- // Print the result of the volume calculation to the log
- print(volume + " " + unit + "^3");
- }
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- ACS, Cancer Statistics, American Cancer Society. 2020. Available online: https://www.cancer.org/research/cancer-facts-statistics/all-cancer-facts-figures/cancer-facts-figures-2020.html (accessed on 26 September 2021).
- Zhuang, H.; Zhou, Z.; Ma, Z.; Li, Z.; Liu, C.; Huang, S.; Zhang, C.; Hou, B. Characterization of the prognostic and oncologic values of ITGB superfamily members in pancreatic cancer. J. Cell Mol. Med. 2020, 24, 13481–13493. [Google Scholar] [CrossRef] [PubMed]
- Grzesiak, J.J.; Cao, H.S.T.; Burton, D.W.; Kaushal, S.; Vargas, F.; Clopton, P.; Snyder, C.S.; Deftos, L.J.; Hoffman, R.M.; Bouvet, M. Knockdown of the beta(1) integrin subunit reduces primary tumor growth and inhibits pancreatic cancer metastasis. Int. J. Cancer 2011, 129, 2905–2915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walsh, N.; Clynes, M.; Crown, J.; O’Donovan, N. Alterations in integrin expression modulates invasion of pancreatic cancer cells. J. Exp. Clin. Cancer Res. 2009, 28, 140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sebaï, N.E.; Trojjet, S.; Khaled, A.; Zekri, S.; Fazaa, B.; Jaafoura, M.H.; Kamoun, M.R. Kindler syndrome: Three cases reports in three siblings. Ann. Derm. Venereol. 2007, 134, 774–778. [Google Scholar]
- Kloeker, S.; Major, M.B.; Calderwood, D.A.; Ginsberg, M.H.; Jones, D.A.; Beckerle, M.C. The Kindler syndrome protein is regulated by transforming growth factor-beta and involved in integrin-mediated adhesion. J. Biol. Chem. 2004, 279, 6824–6833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Has, C.; Yordanova, I.; Balabanova, M.; Kazandjieva, J.; Herz, C.; Kohlhase, J.; Bruckner-Tuderman, L. A novel large FERMT1 (KIND1) gene deletion in Kindler syndrome. J. Dermatol. Sci. 2008, 52, 209–212. [Google Scholar] [CrossRef]
- Böttcher, R.T.; Veelders, M.; Rombaut, P.; Faix, J.; Theodosiou, M.; Stradal, T.E.; Rottner, K.; Zent, R.; Herzog, F.; Fässler, R. Kindlin-2 recruits paxillin and Arp2/3 to promote membrane protrusions during initial cell spreading. J. Cell Biol. 2017, 216, 3785–3798. [Google Scholar] [CrossRef] [Green Version]
- Theodosiou, M.; Widmaier, M.; Böttcher, R.T.; Rognoni, E.; Veelders, M.; Bharadwaj, M.; Lambacher, A.; Austen, K.; Müller, D.J.; Zent, R.; et al. Kindlin-2 cooperates with talin to activate integrins and induces cell spreading by directly binding paxillin. eLife 2016, 5, e10130. [Google Scholar] [CrossRef]
- Kadry, Y.A.; Huet-Calderwood, C.; Simon, B.; Calderwood, D.A. Calderwood, Kindlin-2 interacts with a highly conserved surface of ILK to regulate focal adhesion localization and cell spreading. J. Cell Sci. 2018, 131, jcs221184. [Google Scholar] [CrossRef] [Green Version]
- Mahawithitwong, P.; Ohuchida, K.; Ikenaga, N.; Fujita, H.; Zhao, M.; Kozono, S.; Shindo, K.; Ohtsuka, T.; Aishima, S.; Mizumoto, K.; et al. Kindlin-1 expression is involved in migration and invasion of pancreatic cancer. Int. J. Oncol. 2013, 42, 1360–1366. [Google Scholar] [CrossRef] [Green Version]
- Mahawithitwong, P.; Ohuchida, K.; Ikenaga, N.; Fujita, H.; Zhao, M.; Kozono, S.; Shindo, K.; Ohtsuka, T.; Mizumoto, K.; Tanaka, M. Kindlin-2 expression in peritumoral stroma is associated with poor prognosis in pancreatic ductal adenocarcinoma. Pancreas 2013, 42, 663–669. [Google Scholar] [CrossRef]
- Binkley, C.E.; Zhang, L.; Greenson, J.K.; Giordano, T.J.; Kuick, R.; Misek, D.; Hanash, S.; Logsdon, C.D.; Simeone, D.M. The molecular basis of pancreatic fibrosis: Common stromal gene expression in chronic pancreatitis and pancreatic adenocarcinoma. Pancreas 2004, 29, 254–263. [Google Scholar] [CrossRef]
- Nishimura, S.L. Integrin-mediated transforming growth factor-beta activation, a potential therapeutic target in fibrogenic disorders. Am. J. Pathol. 2009, 175, 1362–1370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munger, J.S.; Sheppard, D. Cross talk among TGF-beta signaling pathways, integrins, and the extracellular matrix. Cold Spring Harb. Perspect. Biol. 2011, 3, a005017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhan, J.; Song, J.; Wang, P.; Chi, X.; Wang, Y.; Guo, Y.; Fang, W.; Zhang, H. Kindlin-2 induced by TGF-beta signaling promotes pancreatic ductal adenocarcinoma progression through downregulation of transcriptional factor HOXB9. Cancer Lett. 2015, 361, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Gradiz, R.; Silva, H.C.; Carvalho, L.; Botelho, M.F.; Mota-Pinto, A. MIA PaCa-2 and PANC-1—Pancreas ductal adenocarcinoma cell lines with neuroendocrine differentiation and somatostatin receptors. Sci. Rep. 2016, 6, 21648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brannon, A.; Drouillard, D.; Steele, N.; Schoettle, S.; Abel, E.V.; Crawford, H.C.; di Magliano, M.P. Beta 1 integrin signaling mediates pancreatic ductal adenocarcinoma resistance to MEK inhibition. Sci. Rep. 2020, 10, 11133. [Google Scholar] [CrossRef]
- Sossey-Alaoui, K.; Pluskota, E.; Bialkowska, K.; Szpak, D.; Parker, Y.; Morrison, C.D.; Lindner, D.J.; Schiemann, W.P.; Plow, E.F. Kindlin-2 Regulates the Growth of Breast Cancer Tumors by Activating CSF-1Mediated Macrophage Infiltration. Cancer Res. 2017, 77, 5129–5141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.; Jiao, H.; Lai, Y.; Zheng, W.; Chen, K.; Qu, H.; Deng, W.; Song, P.; Zhu, K.; Cao, H.; et al. Kindlin-2 controls TGF-beta signalling and Sox9 expression to regulate chondrogenesis. Nat. Commun. 2015, 6, 7531. [Google Scholar] [CrossRef] [Green Version]
- Wei, X.; Xia, Y.; Li, F.; Tang, Y.; Nie, J.; Liu, Y.; Zhou, Z.; Zhang, H.; Hou, F.F. Kindlin-2 mediates activation of TGF-beta/Smad signaling and renal fibrosis. J. Am. Soc. Nephrol. 2013, 24, 1387–1398. [Google Scholar] [CrossRef] [Green Version]
- Hamidi, H.; Pietila, M.; Ivaska, J. The complexity of integrins in cancer and new scopes for therapeutic targeting. Br. J. Cancer 2016, 115, 1017–1023. [Google Scholar] [CrossRef] [Green Version]
- Su, C.Y.; Li, J.Q.; Zhang, L.L.; Wang, H.; Wang, F.H.; Tao, Y.W.; Wang, Y.Q.; Guo, Q.R.; Li, J.J.; Liu, Y.; et al. The Biological Functions and Clinical Applications of Integrins in Cancers. Front. Pharmacol. 2020, 11, 579068. [Google Scholar] [CrossRef]
- Raab-Westphal, S.; Marshall, J.F.; Goodman, S.L. Integrins as Therapeutic Targets: Successes and Cancers. Cancers 2017, 9, 110. [Google Scholar] [CrossRef]
- Maziveyi, M.; Alahari, S.K. Cell matrix adhesions in cancer: The proteins that form the glue. Oncotarget 2017, 8, 48471–48487. [Google Scholar] [CrossRef] [Green Version]
- López-Colomé, A.M.; Lee-Rivera, I.; Benavides-Hidalgo, R.; López, E. Paxillin: A crossroad in pathological cell migration. J. Hematol. Oncol. 2017, 10, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivaska, J. Unanchoring integrins in focal adhesions. Nat. Cell Biol. 2012, 14, 981–983. [Google Scholar] [CrossRef] [PubMed]
- Fischer, L.S.; Klingner, C.; Schlichthaerle, T.; Strauss, M.T.; Böttcher, R.; Fässler, R.; Jungmann, R.; Grashoff, C. Quantitative single-protein imaging reveals molecular complex formation of integrin, talin, and kindlin during cell adhesion. Nat. Commun. 2021, 12, 919. [Google Scholar] [CrossRef]
- Harburger, D.S.; Bouaouina, M.; Calderwood, D.A. Kindlin-1 and -2 directly bind the C-terminal region of beta integrin cytoplasmic tails and exert integrin-specific activation effects. J. Biol. Chem. 2009, 284, 11485–11497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.; Hu, Y.; Gao, Y.; Li, Q.; Zeng, Z.; Li, Y.; Chen, H. Kindlin-2 regulates hepatic stellate cells activation and liver fibrogenesis. Cell Death Discov. 2018, 4, 34. [Google Scholar] [CrossRef]
- Zhang, H.F.; Zhang, K.; Liao, L.D.; Li, L.Y.; Du, Z.P.; Wu, B.L.; Wu, J.Y.; Xu, X.E.; Zeng, F.M.; Chen, B.; et al. miR-200b suppresses invasiveness and modulates the cytoskeletal and adhesive machinery in esophageal squamous cell carcinoma cells via targeting Kindlin-2. Carcinogenesis 2014, 35, 292–301. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, N.; Masamune, A.; Hamada, S.; Kikuta, K.; Takikawa, T.; Motoi, F.; Unno, M.; Shimosegawa, T. Kindlin-2 in pancreatic stellate cells promotes the progression of pancreatic cancer. Cancer Lett. 2017, 390, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Bhowmick, N.A.; Zent, R.; Ghiassi, M.; McDonnell, M.; Moses, H.L. Integrin beta 1 signaling is necessary for transforming growth factor-beta activation of p38MAPK and epithelial plasticity. J. Biol. Chem. 2001, 276, 4670746713. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer Sequence | Reverse Primer Sequence |
|---|---|---|
| ITGα5 | 5′-TGCAGTGTGAGGCTGTGTACA-3′ | 5′-GTGGCCACCTGACGCTCT-3′ |
| Kindlin 2 | 5′-GTCCCCGCTATCTAAAAAAGT-3′ | 5′-GATGGGCCTCCAAGATTCT-3′ |
| TGFβR2 | 5′-ATGACATCTCGCTGTAATGC-3′ | 5′-GGATGCCCTGGTGGTTGA-3′ |
| 18S | 5′-GTAACCCGTTGAACCCCATT-3′ | 5′-CCATCCAATCGGTAGTAGCG-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mia, M.S.; Jarajapu, Y.; Rao, R.; Mathew, S. Integrin β1 Promotes Pancreatic Tumor Growth by Upregulating Kindlin-2 and TGF-β Receptor-2. Int. J. Mol. Sci. 2021, 22, 10599. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910599
Mia MS, Jarajapu Y, Rao R, Mathew S. Integrin β1 Promotes Pancreatic Tumor Growth by Upregulating Kindlin-2 and TGF-β Receptor-2. International Journal of Molecular Sciences. 2021; 22(19):10599. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910599
Chicago/Turabian StyleMia, Md Saimon, Yagna Jarajapu, Reena Rao, and Sijo Mathew. 2021. "Integrin β1 Promotes Pancreatic Tumor Growth by Upregulating Kindlin-2 and TGF-β Receptor-2" International Journal of Molecular Sciences 22, no. 19: 10599. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910599