
Hydro-Electro Hybrid Priming Promotes Carrot (Daucus carota L.) Seed Germination by Activating Lipid Utilization and Respiratory Metabolism


Abstract
:1. Introduction
2. Results
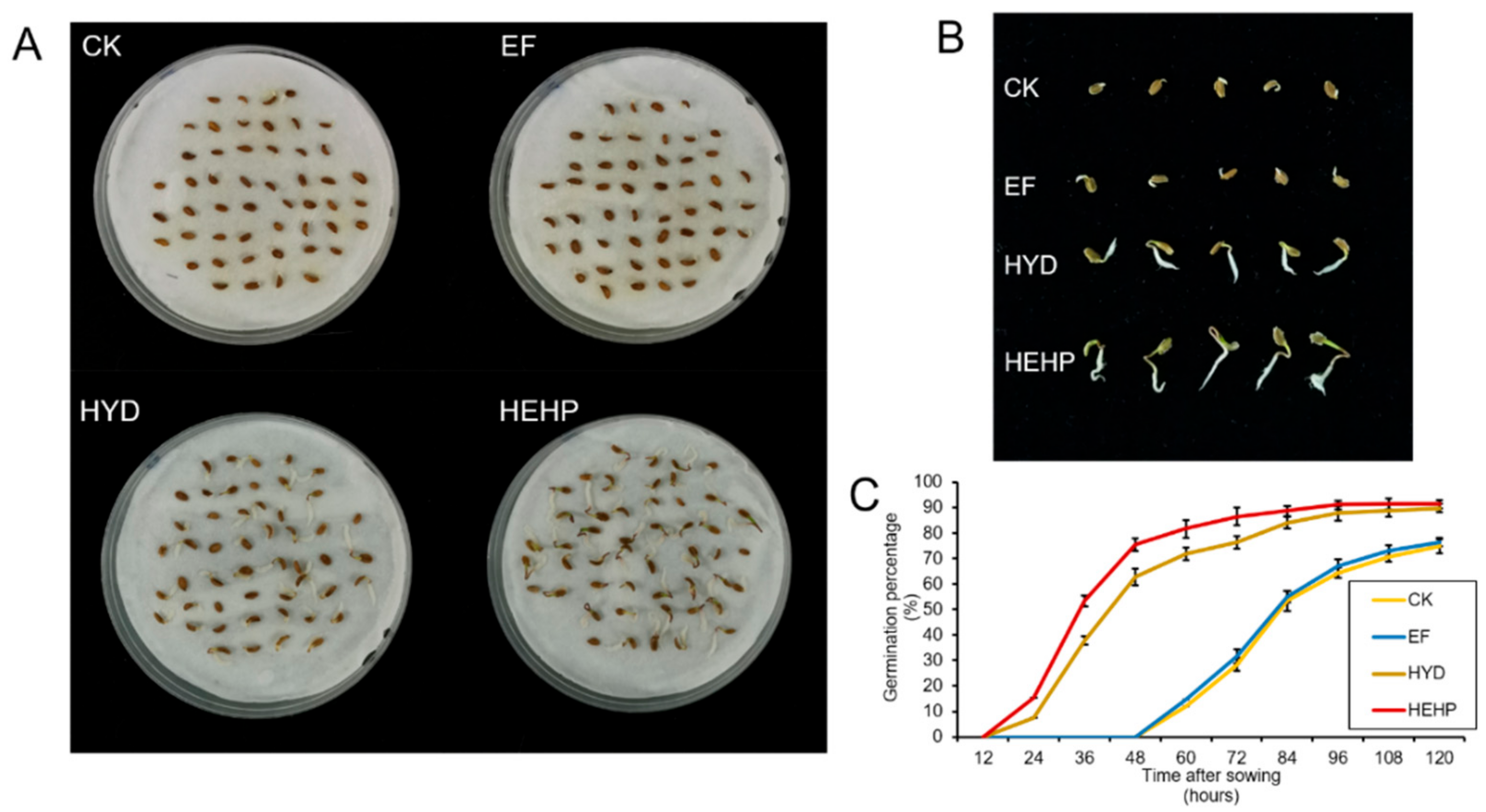
2.1. Induction of Carrot Seed Germination by Different Treatments
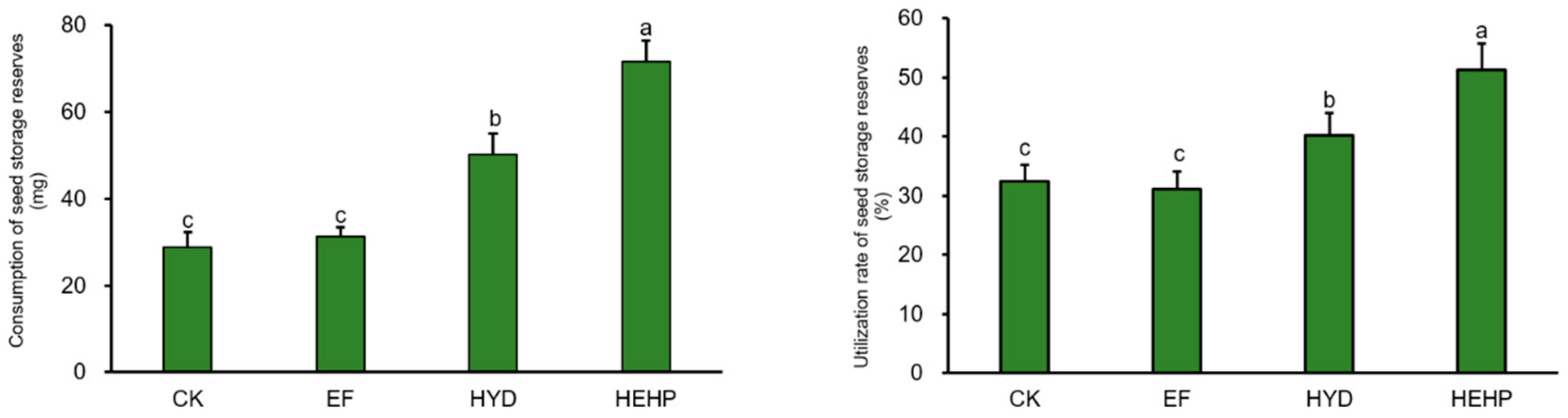
2.2. Effects of Different Treatments on the Consumption and Utilization Rate of Seed Storage Reserves
2.3. Effect of Different Treatments on Subcellular Structure of Endosperm
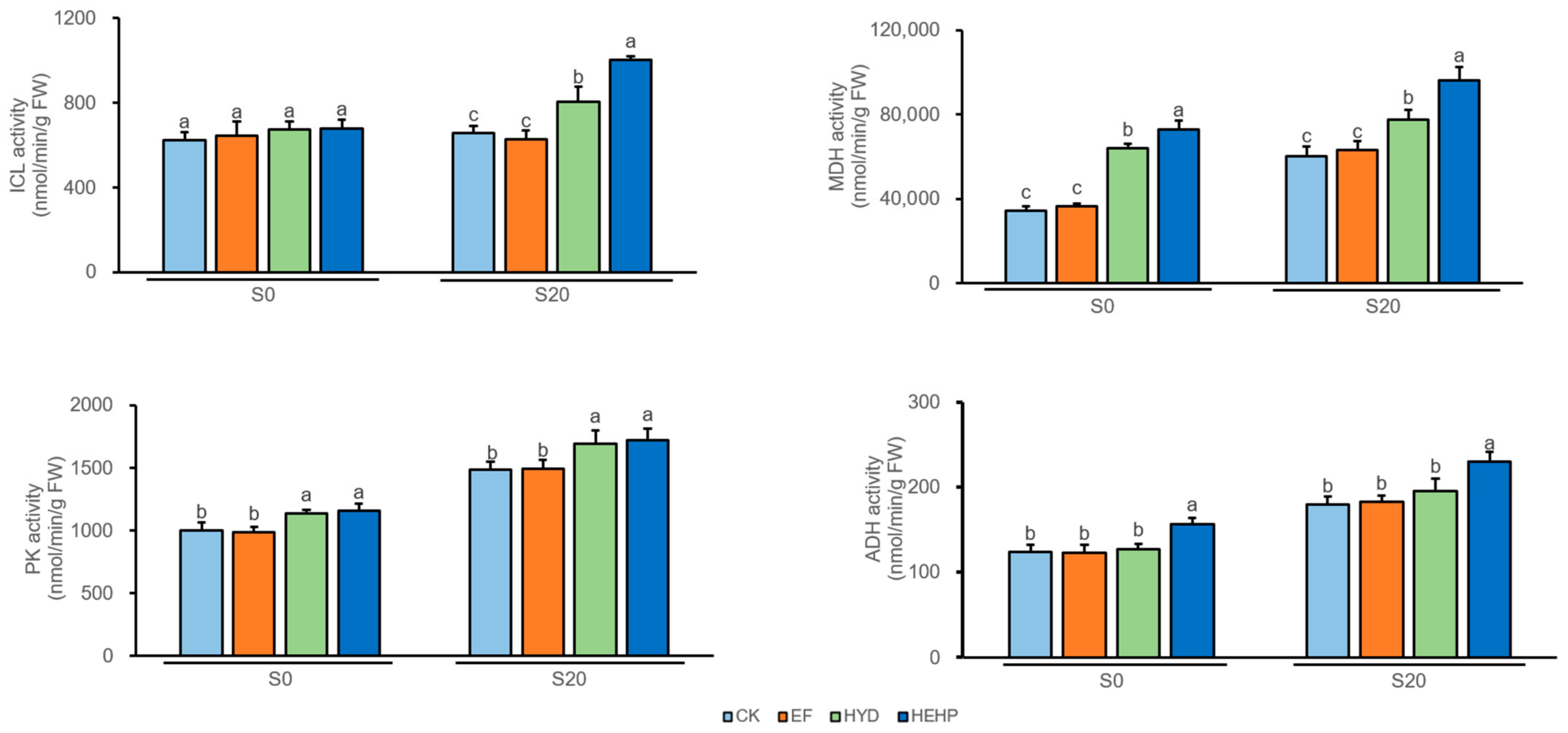
2.4. Analysis of Key Enzyme Activities in the Glyoxylate Cycle and Respiratory Metabolism
2.5. Transcriptome Sequencing Data Statistics, Transcriptional Assembly and Functional Annotation
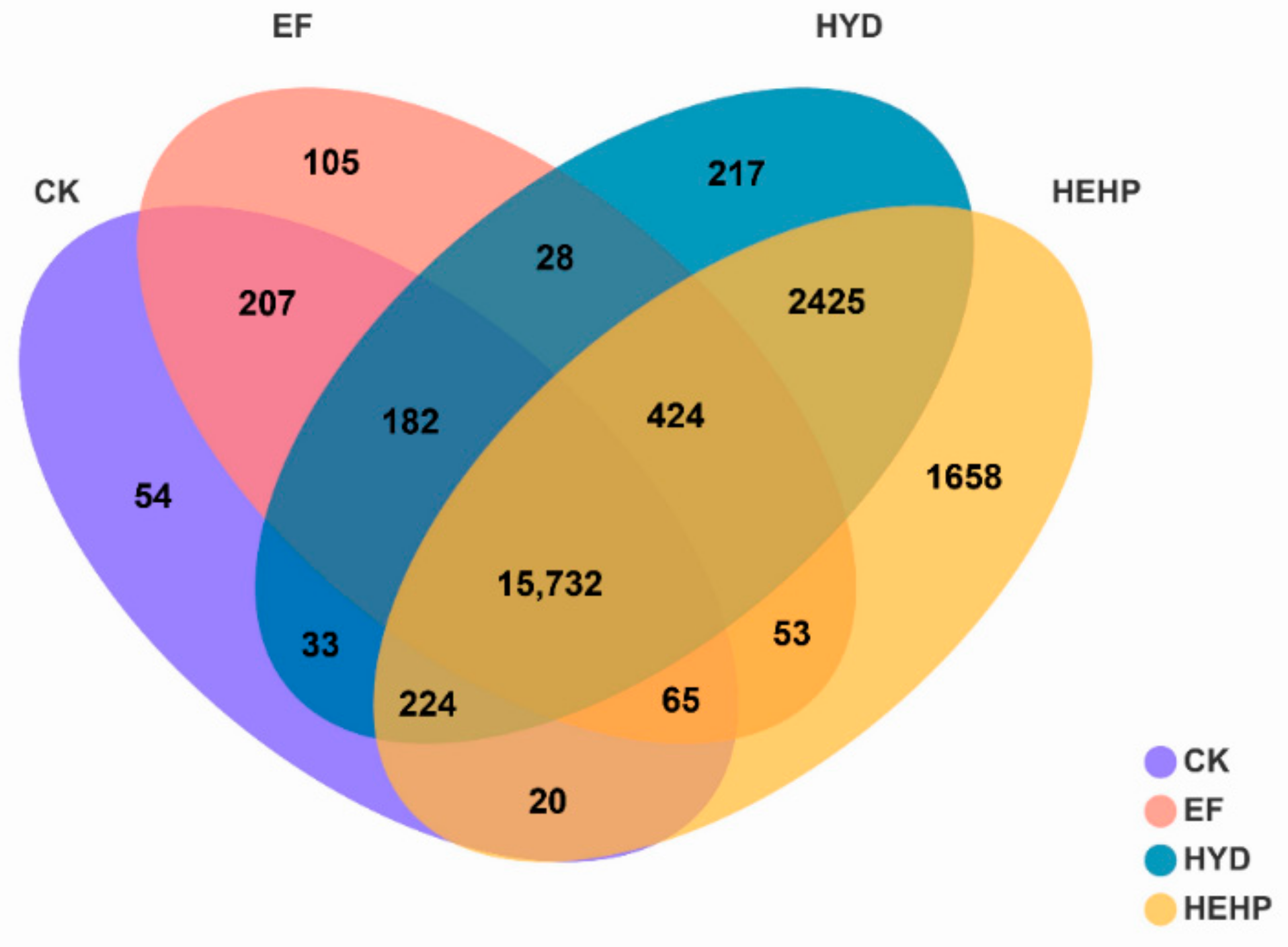
2.6. DEGs Statistics
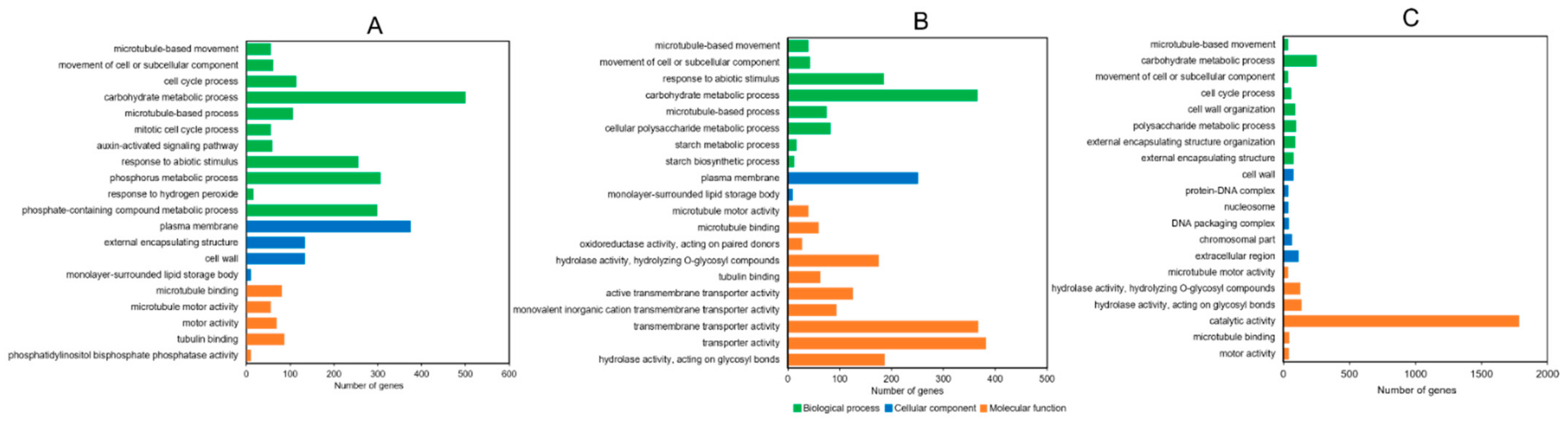
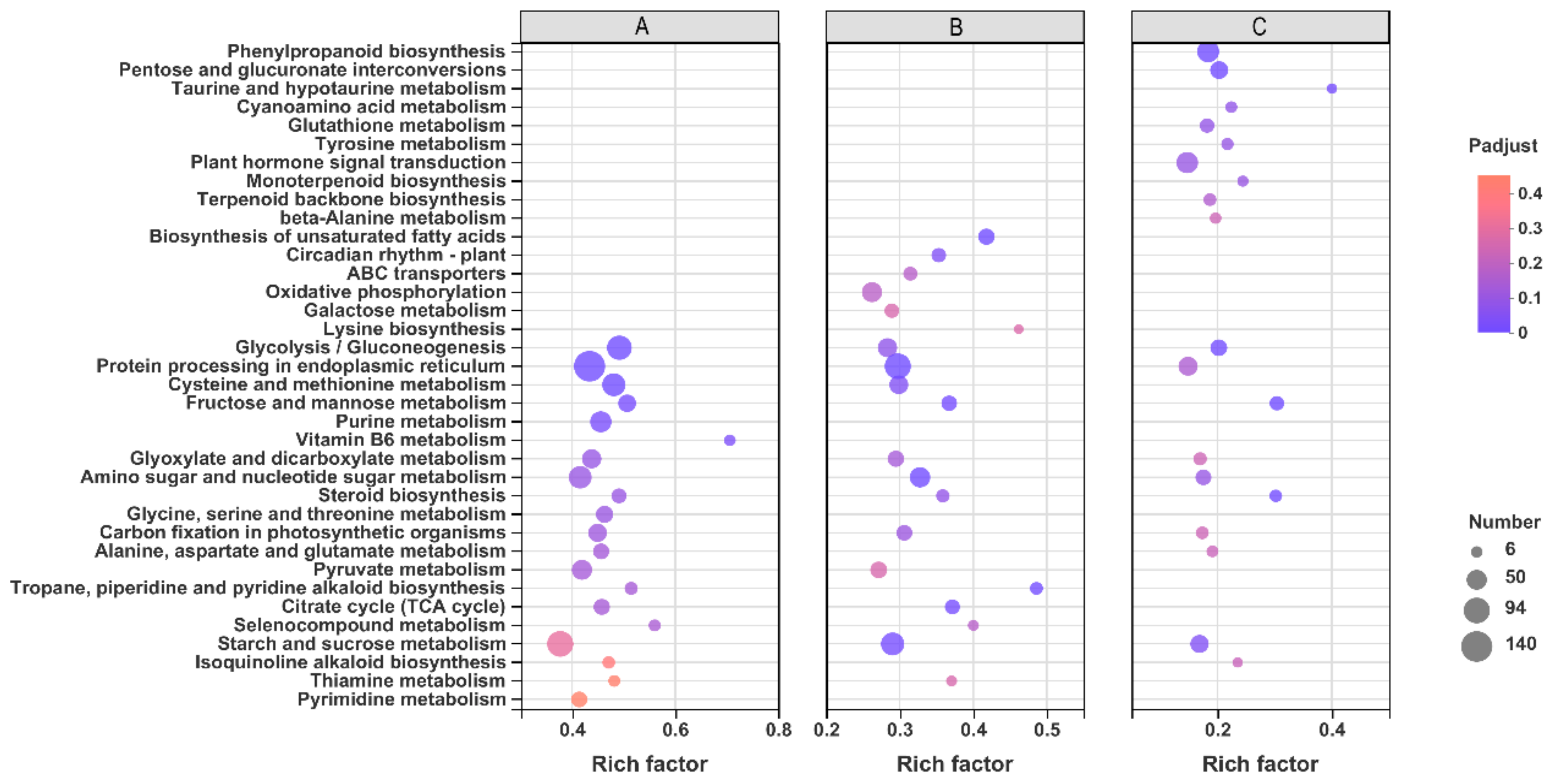
2.7. Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) Enrichment Analysis of DEGs
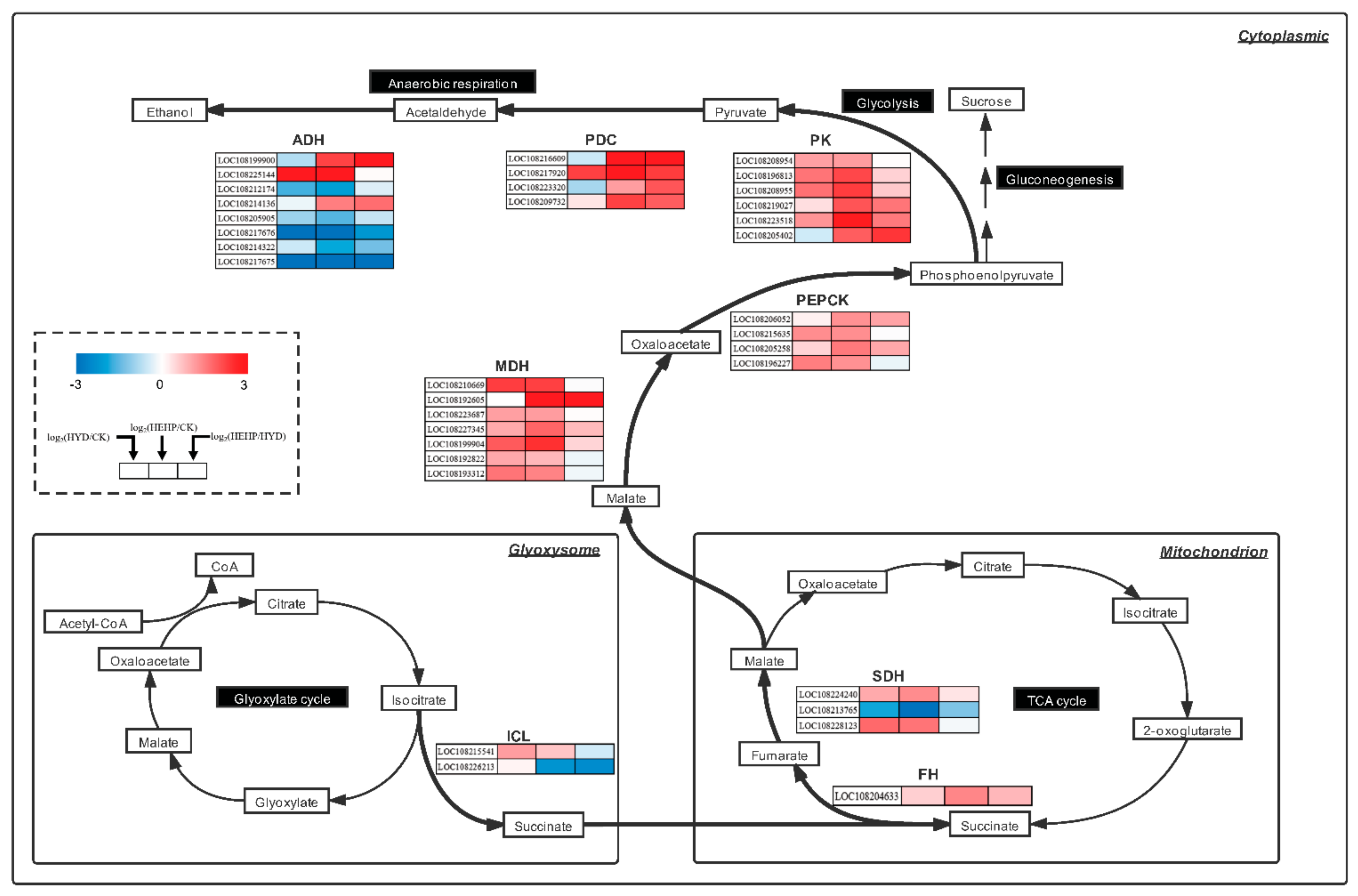
2.8. DEGs Involved in the Glyoxylate Cycle and Respiratory Metabolism
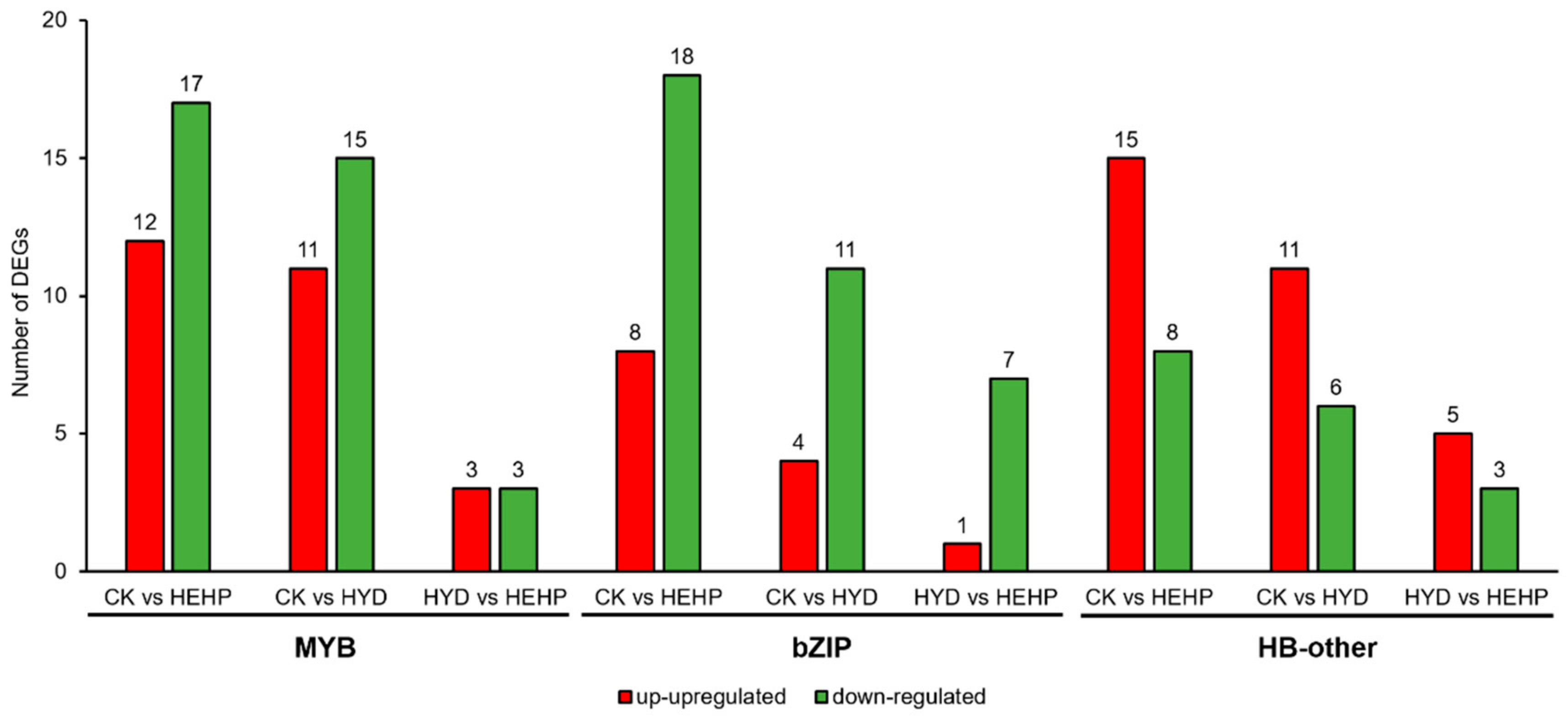
2.9. Analysis of DEGs Identified as Transcription Factors (TFs)
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Treatment Protocols
4.2. Seed Germination Experiments
4.3. Analysis of the Consumption and Utilization Rate of Seed Storage Reserves
4.4. TEM Observation of Endosperm
4.5. Determination of Key Enzyme Activities in the Glyoxylate Cycle and Respiratory Metabolism
4.6. RNA Extraction, Library Preparation and RNA Sequencing Analysis
4.6.1. Extraction of Total RNA
4.6.2. Illumina Sequencing
4.6.3. Gene Expression Statistics
4.6.4. Screening and Analysis of Differentially Expressed Genes (DEGs)
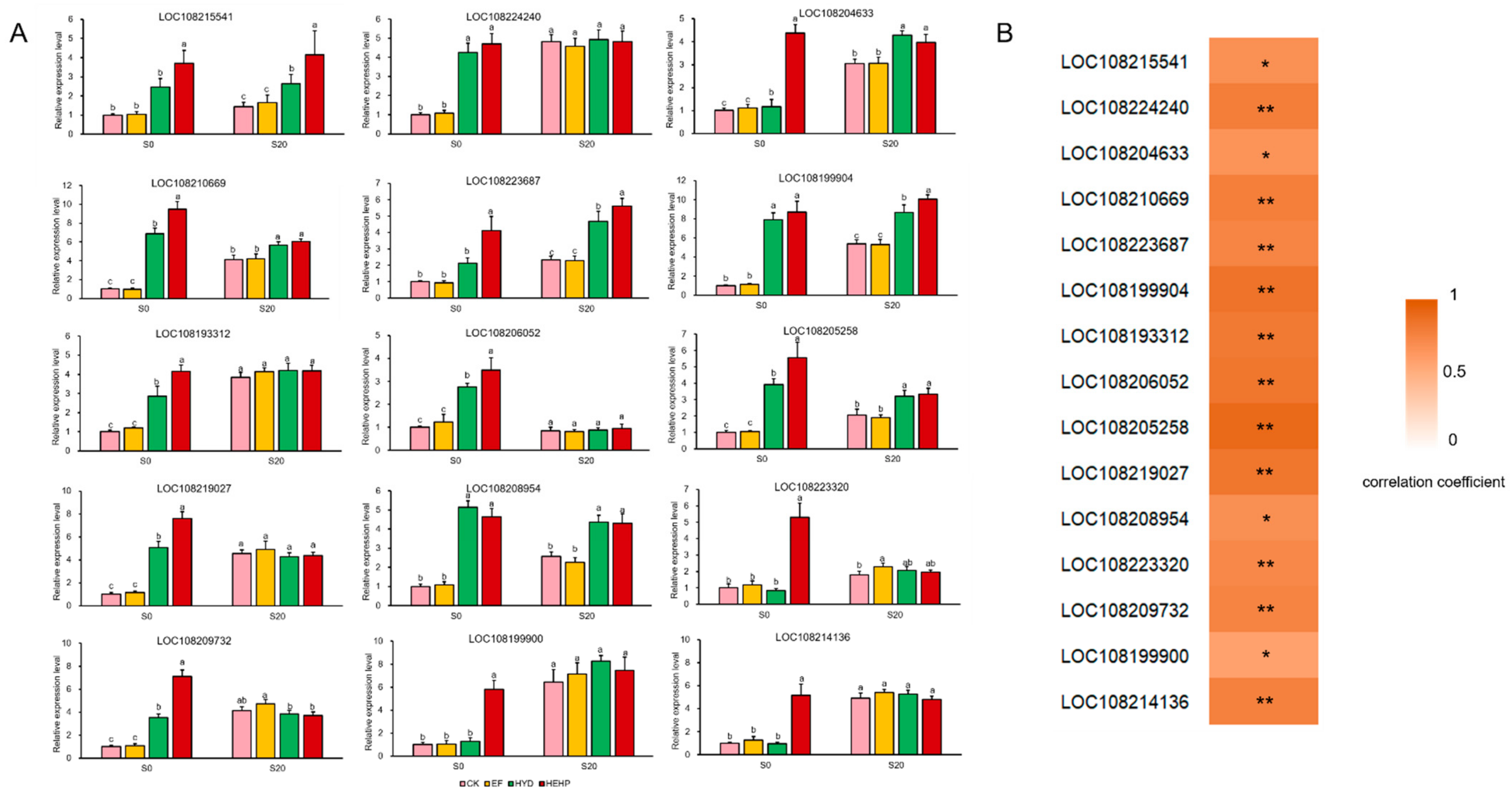
4.7. qRT-PCR Analysis
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Que, F.; Hou, X.L.; Wang, G.L.; Xu, Z.S.; Xiong, A.S. Advances in research on the carrot, an important root vegetable in the Apiaceae family. Hortic. Res. 2019, 6, 69. [Google Scholar] [CrossRef] [Green Version]
- Luby, C.H.; Maeda, H.A.; Goldman, I.L. Genetic and phenological variation of tocochromanol (vitamin E) content in wild (Daucus carota L. var. carota) and domesticated carrot (D. carota L. var. sativa). Hortic. Res. 2014, 1, 15. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, R.T.; Ning, H.L.; Li, W.X.; Bai, Y.L.; Li, Y.G. Evaluation and management of fungal-infected carrot seeds. Sci. Rep. 2020, 10, 8. [Google Scholar] [CrossRef] [PubMed]
- Bewley, J.D. Seed Germination and Dormancy. Plant Cell 1997, 9, 1055–1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szopinska, D.; Jarosz, M.; Slawinska, B. The effect of hydrogen peroxide on seed quality and emergence of carrot (Daucus carota L.). Acta Sci. Pol. Hortorum Cultus 2017, 16, 21–33. [Google Scholar]
- Nascimento, W.M.; Silva, J.B.C.; Santos, P.E.; Carmona, R. Germination of carrot seed primed and pelleted with several ingredients. Photonics West 2009, 27, 12–16. [Google Scholar] [CrossRef]
- Paparella, S.; Araujo, S.S.; Rossi, G.; Wijayasinghe, M.; Carbonera, D.; Balestrazzi, A. Seed priming: State of the art and new perspectives. Plant Cell Rep. 2015, 34, 1281–1293. [Google Scholar] [CrossRef]
- Yan, H.F.; Mao, P.S. Comparative Time-Course Physiological Responses and Proteomic Analysis of Melatonin Priming on Promoting Germination in Aged Oat (Avena sativa L.) Seeds. Int. J. Mol. Sci. 2021, 22, 811. [Google Scholar] [CrossRef]
- Zhang, N.; Zhang, H.J.; Zhao, B.; Sun, Q.Q.; Cao, Y.Y.; Li, R.; Wu, X.X.; Weeda, S.; Li, L.; Ren, S.X.; et al. The RNA-seq approach to discriminate gene expression profiles in response to melatonin on cucumber lateral root formation. J. Pineal Res. 2014, 56, 39–50. [Google Scholar] [CrossRef]
- Rai-Kalal, P.; Jajoo, A. Priming with zinc oxide nanoparticles improve germination and photosynthetic performance in wheat. Plant Physiol. Biochem. 2021, 160, 341–351. [Google Scholar] [CrossRef]
- Khan, M.N.; Zhang, J.; Luo, T.; Liu, J.; Rizwan, M.; Fahad, S.; Xu, Z.; Hu, L. Seed priming with melatonin coping drought stress in rapeseed by regulating reactive oxygen species detoxification: Antioxidant defense system, osmotic adjustment, stomatal traits and chloroplast ultrastructure perseveration. Ind. Crop. Prod. 2019, 140, 111597. [Google Scholar] [CrossRef]
- Nakao, Y.; Sone, C.; Sakagami, J.I. Genetic Diversity of Hydro Priming Effects on Rice Seed Emergence and Subsequent Growth under Different Moisture Conditions. Genes 2020, 11, 994. [Google Scholar] [CrossRef]
- Yan, M. Hydro-priming increases seed germination and early seedling growth in two cultivars of Napa cabbage (Brassica rapa subsp pekinensis) grown under salt stress. J. Horticult. Sci. Biotechnol. 2016, 91, 421–426. [Google Scholar] [CrossRef]
- Singh, H.; Jassal, R.K.; Kang, J.S.; Sandhu, S.S.; Grewal, K. Seed priming techniques in field crops—A review. Agric. Rev. 2015, 36, 251–264. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Huang, J.; Gao, W.; Lu, J.; Li, J.; Liao, R.; Jaleel, C.A. The effect of high-voltage electrostatic field (HVEF) on aged rice (Oryza sativa L.) seeds vigor and lipid peroxidation of seedlings. J. Electrost. 2009, 67, 759–764. [Google Scholar] [CrossRef]
- Cramariuc, R.; Donescu, V.; Popa, M.; Cramariuc, B. The biological effect of the electrical field treatment on the potato seed: Agronomic evaluation. J. Electrost. 2005, 63, 837–846. [Google Scholar] [CrossRef]
- Moon, J.D.; Chung, H.S. Acceleration of germination of tomato seed by applying AC electric and magnetic fields. J. Electrost. 2000, 48, 103–114. [Google Scholar] [CrossRef]
- Guo, Q.; Wang, Y.; Zhang, H.; Qu, G.; Wang, T.; Sun, Q.; Liang, D. Alleviation of adverse effects of drought stress on wheat seed germination using atmospheric dielectric barrier discharge plasma treatment. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Bao, S.-Q.; Yang, T.-Q.; Ma, Z.-X.; Ao, D.-G. Aging Property of the Effect on Wheat Seeds in High Voltage Electric Field. High Volt. Eng. 2010, 36, 467–473. [Google Scholar]
- Liu, X.; Yang, T.; Zhu, H.; Hid, Y.; Wang, Z.; Wang, W. Effective Time of the Effects of High Electric Field Treatment on Drought Resistance of Caragana korshinskii Seeds During its Germination and Seedling Growth. Seed 2014, 33, 8–11. [Google Scholar]
- Zhao, Y.; Hu, M.; Gao, Z.; Chen, X.; Huang, D. Biological mechanisms of a novel hydro-electro hybrid priming recovers potential vigor of onion seeds. Environ. Exp. Bot. 2018, 150, 260–271. [Google Scholar] [CrossRef]
- Dong, K.; Zhen, S.; Cheng, Z.; Cao, H.; Ge, P.; Yan, Y. Proteomic Analysis Reveals Key Proteins and Phosphoproteins upon Seed Germination of Wheat (Triticum aestivum L.). Front. Plant Sci. 2015, 6, 1017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, S.; Kumari, A.; Shree, M.; Kumar, V.; Singh, P.; Bharadwaj, C.; Loake, G.J.; Parida, S.K.; Masakapalli, S.K.; Gupta, K.J. Nitric oxide accelerates germination via the regulation of respiration in chickpea. J. Exp. Bot. 2019, 70, 4539–4555. [Google Scholar] [CrossRef] [PubMed]
- Ngo-Duy, C.C.; Destaillats, F.; Keskitalo, M.; Arul, J.; Angers, P. Triacylglycerols of Apiaceae seed oils: Composition and regiodistribution of fatty acids. Eur. J. Lipid Sci. Technol. 2009, 111, 164–169. [Google Scholar] [CrossRef]
- Footitt, S.; Slocombe, S.P.; Larner, V.; Kurup, S.; Wu, Y.S.; Larson, T.; Graham, I.; Baker, A.; Holdsworth, M. Control of germination and lipid mobilization by COMATOSE, the Arabidopsis homologue of human ALDP. Embo J. 2002, 21, 2912–2922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bello, P.; Bradford, K.J. Single-seed oxygen consumption measurements and population-based threshold models link respiration and germination rates under diverse conditions. Seed Sci. Res. 2016, 26, 199–221. [Google Scholar] [CrossRef] [Green Version]
- Plazek, A.; Dubert, F.; Kopec, P.; Dziurka, M.; Kalandyk, A.; Pastuszak, J.; Wolko, B. Seed Hydropriming and Smoke Water Significantly Improve Low-Temperature Germination of Lupinus angustifolius L. Int. J. Mol. Sci. 2018, 19, 992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosental, L.; Nonogaki, H.; Fait, A. Activation and regulation of primary metabolism during seed germination. Seed Sci. Res. 2014, 24, 1–15. [Google Scholar] [CrossRef]
- Bewley, J.D.; Bradford, K.J.; Hilhorst, H.W.M.; Nonogaki, H. Seeds: Physiology of Development, Germination and Dormancy, 3rd ed.; Springer: New York, NY, USA, 2013; ISBN 978-1-4614-4693-4. [Google Scholar]
- Weitbrecht, K.; Mueller, K.; Leubner-Metzger, G. First off the mark: Early seed germination. J. Exp. Bot. 2011, 62, 3289–3309. [Google Scholar] [CrossRef] [Green Version]
- Eastmond, P.J.; Graham, I.A. Re-examining the role of the glyoxylate cycle in oilseeds. Trends Plant Sci. 2001, 6, 72–77. [Google Scholar] [CrossRef]
- Nascimento, W.M.; Huber, D.J.; Cantliffe, D.J. Carrot seed germination and respiration at high temperature in response to seed maturity and priming. Seed Sci. Technol. 2013, 41, 164–169. [Google Scholar] [CrossRef]
- Singh, P.K.; Pandita, V.K.; Tomar, B.S.; Seth, R. Standardization of priming treatments for enhancement of seed germination and field emergence in carrot. Indian J. Hortic. 2015, 72, 306–309. [Google Scholar] [CrossRef]
- Tylkowska, K.; Bulk, R.W.V.D. Effects of osmo-and hydropriming on fungal infestation levels and germination of carrot (Daucus carota L.) seeds contaminated with Alternaria spp. Seed Sci. Technol. 2001, 29, 365–375. [Google Scholar]
- Rajjou, L.; Duval, M.; Gallardo, K.; Catusse, J.; Job, D. Seed Germination and Vigor. Annu. Rev. Plant Biol. 2012, 63, 507–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finch-Savage, W.E.; Bassel, G.W. Seed vigour and crop establishment: Extending performance beyond adaptation. J. Exp. Bot. 2016, 67, 567–591. [Google Scholar] [CrossRef] [Green Version]
- Ayuso, M.; Ramil-Rego, P.; Landin, M.; Gallego, P.P.; Barreal, M.E. Computer-Assisted Recovery of Threatened Plants: Keys for Breaking Seed Dormancy of Eryngium viviparum. Front. Plant Sci. 2017, 8, 10. [Google Scholar] [CrossRef] [Green Version]
- Hassell, R.L.; Kretchman, D.W. The effects of umbel order, soaking, and scarification on germination inhibiting substances in Petroselinum crispum L. and other Apiaceae seeds. Hortscience 1997, 32, 1227–1230. [Google Scholar] [CrossRef] [Green Version]
- Walck, J.L.; Baskin, C.C.; Hidayati, S.N.; Baskin, J.M. Comparison of the seed germination of native and non-native winter annual Apiaceae in North America, with particular focus on Cyclospermum leptophyllum naturalized from South America. Plant Species Biol. 2008, 23, 33–42. [Google Scholar] [CrossRef]
- Chen, J.; Hu, J.; Du, H.; Wang, H.; Wang, Y. Response of endogenous hormone in germination of millet seeds to optimal high voltage electrostatic field. Trans. Chin. Soc. Agric. Eng. 2016, 32, 286–292. [Google Scholar]
- Morar, R.; Munteanu, R.; Simion, E.; Munteanu, I.; Dascalescu, L. Electrostatic treatment of bean seeds. IEEE Trans. Ind. Appl. 1995, 2, 1335–1337. [Google Scholar] [CrossRef]
- Patwardhan, M.S. High voltage electric field effects on the germination rate of tomato seeds. Acta Agrophys. 2013, 20, 403–413. [Google Scholar]
- Nonogaki, H.; Bassel, G.W.; Bewley, J.D. Germination-Still a mystery. Plant Sci. 2010, 179, 574–581. [Google Scholar] [CrossRef]
- Banilas, G.; Karampelias, M.; Makariti, I.; Kourti, A.; Hatzopoulos, P. The olive DGAT2 gene is developmentally regulated and shares overlapping but distinct expression patterns with DGAT1. J. Exp. Bot. 2011, 62, 521–532. [Google Scholar] [CrossRef] [Green Version]
- Huang, A.H.C. Oleosins and Oil Bodies in Seeds and Other Organs. Plant Physiol. 1996, 110, 1055–1061. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.-L.; Hsiao, Y.-Y.; Lu, H.-C.; Liang, C.-K.; Fu, C.-H.; Huang, T.-H.; Chuang, M.-H.; Chen, L.-J.; Liu, Z.-J.; Tsai, W.-C. Expression regulation of MALATE SYNTHASE involved in glyoxylate cycle during protocorm development in Phalaenopsis aphrodite (Orchidaceae). Sci. Rep. 2020, 10, 1–16. [Google Scholar] [CrossRef]
- Bewley, J.D. Seed Germination and Reserve Mobilization; Nature Publishing Group: London, UK, 2001; ISBN 0470016175. [Google Scholar]
- Ma, Z.; Marsolais, F.; Bernards, M.A.; Sumarah, M.W.; Bykova, N.V.; Igamberdiev, A.U. Glyoxylate cycle and metabolism of organic acids in the scutellum of barley seeds during germination. Plant Sci. 2016, 248, 37–44. [Google Scholar] [CrossRef]
- Su, C.; Yang, C.; Gong, M.; Ke, Y.; Yuan, P.; Wang, X.; Li, M.; Zheng, X.; Feng, W. Antidiabetic Activity and Potential Mechanism of Amentoflavone in Diabetic Mice. Molecules 2019, 24, 2184. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Ye, Z.; Ying, Q.; Ma, Y.; Zang, Y.; Wang, H.; Yu, Y.; Zhu, Z. Glyoxylate cycle and reactive oxygen species metabolism are involved in the improvement of seed vigor in watermelon by exogenous GA(3). Sci. Hortic. 2019, 247, 184–194. [Google Scholar] [CrossRef]
- Qu, C.; Zhang, S.; Zhao, H.; Chen, J.; Zuo, Z.; Sun, X.; Cheng, Y.; Xu, Z.; Liu, G. Analysis of the energy source at the early stage of poplar seed germination: Verification of Perl’s pathway. 3 Biotech 2020, 10, 1–16. [Google Scholar] [CrossRef]
- Tougou, M.; Hashiguchi, A.; Yukawa, K.; Nanjo, Y.; Hiraga, S.; Nakamura, T.; Nishizawa, K.; Komatsu, S. Responses to flooding stress in soybean seedlings with the alcohol dehydrogenase transgene. Plant Biotechnol. 2012, 29, 301–305. [Google Scholar] [CrossRef] [Green Version]
- Vijayan, J.; Senapati, S.; Ray, S.; Chakraborty, K.; Molla, K.A.; Basak, N.; Pradhan, B.; Yeasmin, L.; Chattopadhyay, K.; Sarkar, R.K. Transcriptomic and physiological studies identify cues for germination stage oxygen deficiency tolerance in rice. Environ. Exp. Bot. 2018, 147, 234–248. [Google Scholar] [CrossRef]
- Mondal, S.; Khan, M.I.R.; Entila, F.; Dixit, S.; Cruz, P.C.S.; Ali, M.P.; Pittendrigh, B.; Septiningsih, E.M.; Ismail, A.M. Responses of AG1 and AG2 QTL introgression lines and seed pre-treatment on growth and physiological processes during anaerobic germination of rice under flooding. Sci. Rep. 2020, 10, 15. [Google Scholar] [CrossRef]
- Bellieny-Rabelo, D.; Gamosa de Oliveira, E.A.; Ribeiro, E.d.S.; Costa, E.P.; Amancio Oliveira, A.E.; Venancio, T.M. Transcriptome analysis uncovers key regulatory and metabolic aspects of soybean embryonic axes during germination. Sci. Rep. 2016, 6, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.H.; Chu, P.; Zhou, Y.L.; Li, Y.; Liu, J.; Ding, Y.; Tsang, E.W.T.; Jiang, L.W.; Wu, K.Q.; Huang, S.Z. Overexpression of AtOGG1, a DNA glycosylase/AP lyase, enhances seed longevity and abiotic stress tolerance in Arabidopsis. J. Exp. Bot. 2012, 63, 4107–4121. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Qin, F.; Tran, L.S.P. Contribution of Genomics to Gene Discovery in Plant Abiotic Stress Responses. Mol. Plant. 2012, 5, 1176–1178. [Google Scholar] [CrossRef]
- Abid, M.; Hakeem, A.; Shao, Y.H.; Liu, Y.; Zahoor, R.; Fan, Y.H.; Jiang, S.Y.; Ata-Ul-Karim, S.T.; Tian, Z.W.; Jiang, D.; et al. Seed osmopriming invokes stress memory against post-germinative drought stress in wheat (Triticum aestivum L.). Environ. Exp. Bot. 2018, 145, 12–20. [Google Scholar] [CrossRef]
- Chen, K.; Arora, R. Priming memory invokes seed stress-tolerance. Environ. Exp. Bot. 2013, 94, 33–45. [Google Scholar] [CrossRef]
- Shu, K.; Liu, X.D.; Xie, Q.; He, Z.H. Two Faces of One Seed: Hormonal Regulation of Dormancy and Germination. Mol. Plant. 2016, 9, 34–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piskurewicz, U.; Jikumaru, Y.; Kinoshita, N.; Nambara, E.; Kamiya, Y.; Lopez-Molina, L. The Gibberellic Acid Signaling Repressor RGL2 Inhibits Arabidopsis Seed Germination by Stimulating Abscisic Acid Synthesis and ABI5 Activity. Plant Cell 2008, 20, 2729–2745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.; Hong, G.L.; Yoon, S.; Kim, H.U.; Seo, P.J. The Arabidopsis MYB96 Transcription Factor Is a Positive Regulator of ABSCISIC ACID-INSENSITIVE4 in the Control of Seed Germination. Plant Physiol. 2015, 168, 677–689. [Google Scholar] [CrossRef] [Green Version]
- Gao, F.; Yao, H.P.; Zhao, H.X.; Zhou, J.; Luo, X.P.; Huang, Y.J.; Li, C.L.; Chen, H.; Wu, Q. Tartary buckwheat FtMYB10 encodes an R2R3-MYB transcription factor that acts as a novel negative regulator of salt and drought response in transgenic Arabidopsis. Plant Physiol. Biochem. 2016, 109, 387–396. [Google Scholar] [CrossRef]
- Kim, J.H.; Hyun, W.Y.; Nguyen, H.N.; Jeong, C.Y.; Xiong, L.; Hong, S.W.; Lee, H. AtMyb7, a subgroup 4 R2R3 Myb, negatively regulates ABA-induced inhibition of seed germination by blocking the expression of the bZIP transcription factor ABI5. Plant Cell Environ. 2015, 38, 559–571. [Google Scholar] [CrossRef]
- Penfield, S.; Li, Y.; Gilday, A.D.; Graham, S.; Ian, A. Arabidopsis ABA INSENSITIVE4 Regulates Lipid Mobilization in the Embryo and Reveals Repression of Seed Germination by the Endosperm. Plant Cell 2006, 18, 1887–1899. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Zhang, X.; Gong, Z.; Yang, S.; Shi, Y. ABI4 represses the expression of type-A ARRs to inhibit seed germination in Arabidopsis. Plant J. 2017, 89, 354–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ariel, F.D.; Manavella, P.A.; Dezar, C.A.; Chan, R.L. The true story of the HD-Zip family. Trends Plant Sci. 2007, 12, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Barrero, J.M.; Millar, A.A.; Griffiths, J.; Czechowski, T.; Scheible, W.R.; Udvardi, M.; Reid, J.B.; Ross, J.J.; Jacobsen, J.V.; Gubler, F. Gene expression profiling identifies two regulatory genes controlling dormancy and ABA sensitivity in Arabidopsis seeds. Plant J. 2010, 61, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Rombola-Caldentey, B.; Rueda-Romero, P.; Iglesias-Fernandez, R.; Carbonero, P.; Onate-Sanchez, L. Arabidopsis DELLA and Two HD-ZIP Transcription Factors Regulate GA Signaling in the Epidermis through the L1 Box cis-Element. Plant Cell 2014, 26, 2905–2919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baima, S.; Forte, V.; Possenti, M.; Penalosa, A.; Leoni, G.; Salvi, S.; Felici, B.; Ruberti, I.; Morelli, G. Negative Feedback Regulation of Auxin Signaling by ATHB8/ACL5-BUD2 Transcription Module. Mol. Plant. 2014, 7, 1006–1025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capella, M.; Ribone, P.A.; Arce, A.L.; Chan, R.L. Arabidopsis thaliana HomeoBox 1 (AtHB1), a Homedomain-Leucine Zipper I (HD-Zip I) transcription factor, is regulated by PHYTOCHROME-INTERACTING FACTOR 1 to promote hypocotyl elongation. New Phytol. 2015, 207, 669–682. [Google Scholar] [CrossRef]
- Soltani, A.; Gholipoor, A.; Zeinali, E. Seed reserve utilization and seedling growth of wheat as affected by drought and salinity. Environ. Exp. Bot. 2006, 55, 195–200. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | GP/% | GR/% | RL/mm | FW/g | GI | VI | MGT/d |
|---|---|---|---|---|---|---|---|
| CK | 27.0 ± 2.4c | 74.7 ± 3.0b | 6.84 ± 0.40c | 0.058 ± 0.004d | 20.06 ± 0.48c | 1.16 ± 0.09c | 4.29 ± 0.02a |
| EF | 29.3 ± 1.6c | 76.3 ± 2.3b | 7.47 ± 0.26c | 0.062 ± 0.003c | 20.90 ± 0.15c | 1.29 ± 0.05c | 4.28 ± 0.02a |
| HYD | 76.3 ± 2.3b | 89.7 ± 1.5a | 17.01 ± 1.48b | 0.120 ± 0.002b | 51.98 ± 1.16b | 6.23 ± 0.02b | 3.59 ± 0.02b |
| HEHP | 86.3 ± 2.0a | 91.3 ± 1.6a | 22.96 ± 1.09a | 0.129 ± 0.003a | 61.40 ± 1.85a | 7.93 ± 0.35a | 3.47 ± 0.03c |
| Group | Total DEGs | Up | Down |
|---|---|---|---|
| CK vs. EF | 36 | 31 | 5 |
| CK vs. HYD | 6808 | 4234 | 2574 |
| CK vs. HEHP | 10,986 | 6655 | 4331 |
| EF vs. HYD | 6612 | 4053 | 2559 |
| EF vs. HEHP | 10,721 | 6531 | 4190 |
| HYD vs. HEHP | 3853 | 2940 | 913 |
| Treatment | Soaking | Exposure in Electrostatic Field | Imbibition in Climate Chamber | Desiccation |
|---|---|---|---|---|
| CK | × | × | × | × |
| EF | × | √ | × | × |
| HYD | √ | × | √ | √ |
| HEHP | √ | √ | √ | √ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, S.; Garcia, D.; Zhao, Y.; Huang, D. Hydro-Electro Hybrid Priming Promotes Carrot (Daucus carota L.) Seed Germination by Activating Lipid Utilization and Respiratory Metabolism. Int. J. Mol. Sci. 2021, 22, 11090. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222011090
Zhao S, Garcia D, Zhao Y, Huang D. Hydro-Electro Hybrid Priming Promotes Carrot (Daucus carota L.) Seed Germination by Activating Lipid Utilization and Respiratory Metabolism. International Journal of Molecular Sciences. 2021; 22(20):11090. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222011090
Chicago/Turabian StyleZhao, Shuo, Daniel Garcia, Yinglei Zhao, and Danfeng Huang. 2021. "Hydro-Electro Hybrid Priming Promotes Carrot (Daucus carota L.) Seed Germination by Activating Lipid Utilization and Respiratory Metabolism" International Journal of Molecular Sciences 22, no. 20: 11090. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222011090