
The Effects of SRT1720 Treatment on Endothelial Cells Derived from the Lung and Bone Marrow of Young and Aged, Male and Female Mice

 , ,
, , 
Abstract
:1. Introduction
2. Results
2.1. Characterization of Lung and Bone Marrow Endothelial Cells
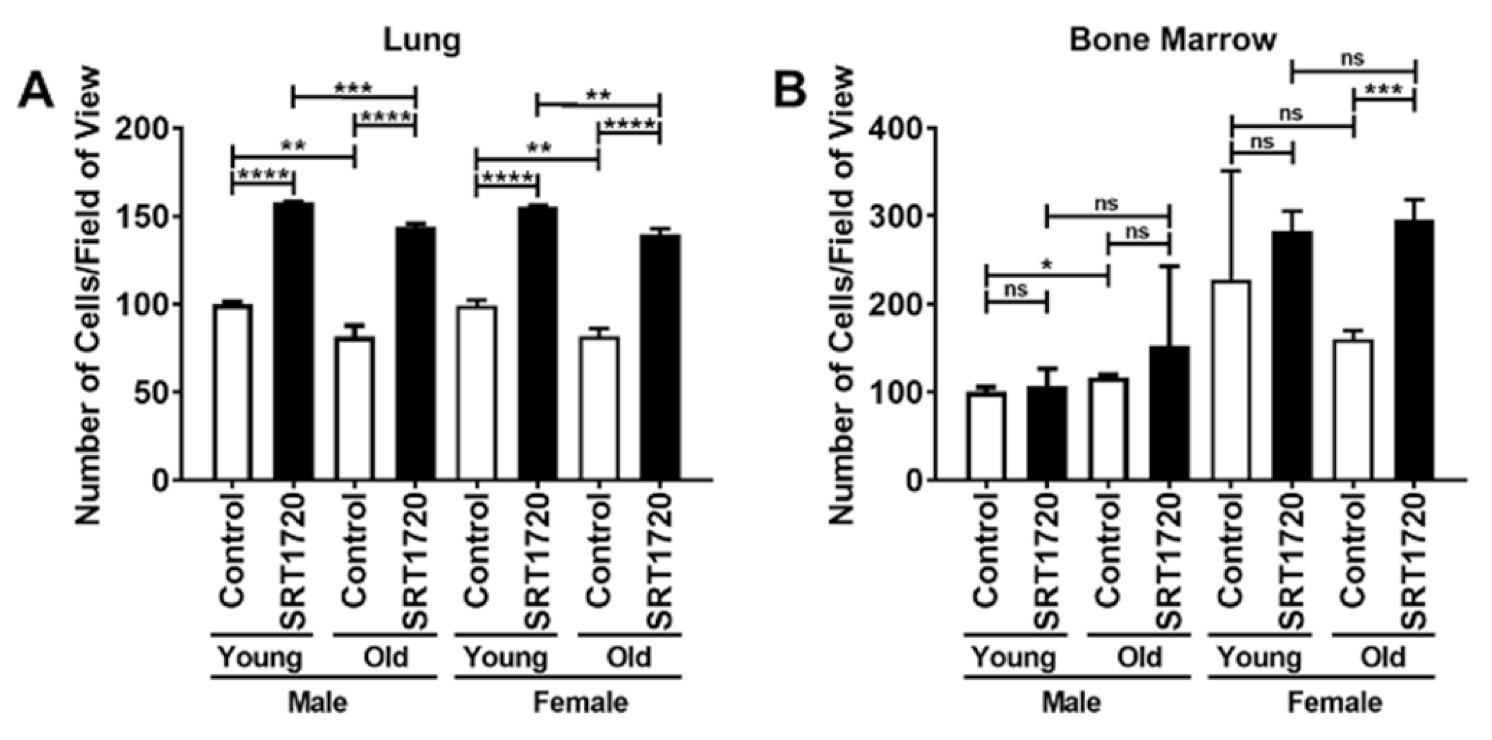
2.2. Proliferation of Lung and Bone Marrow Endothelial Cells
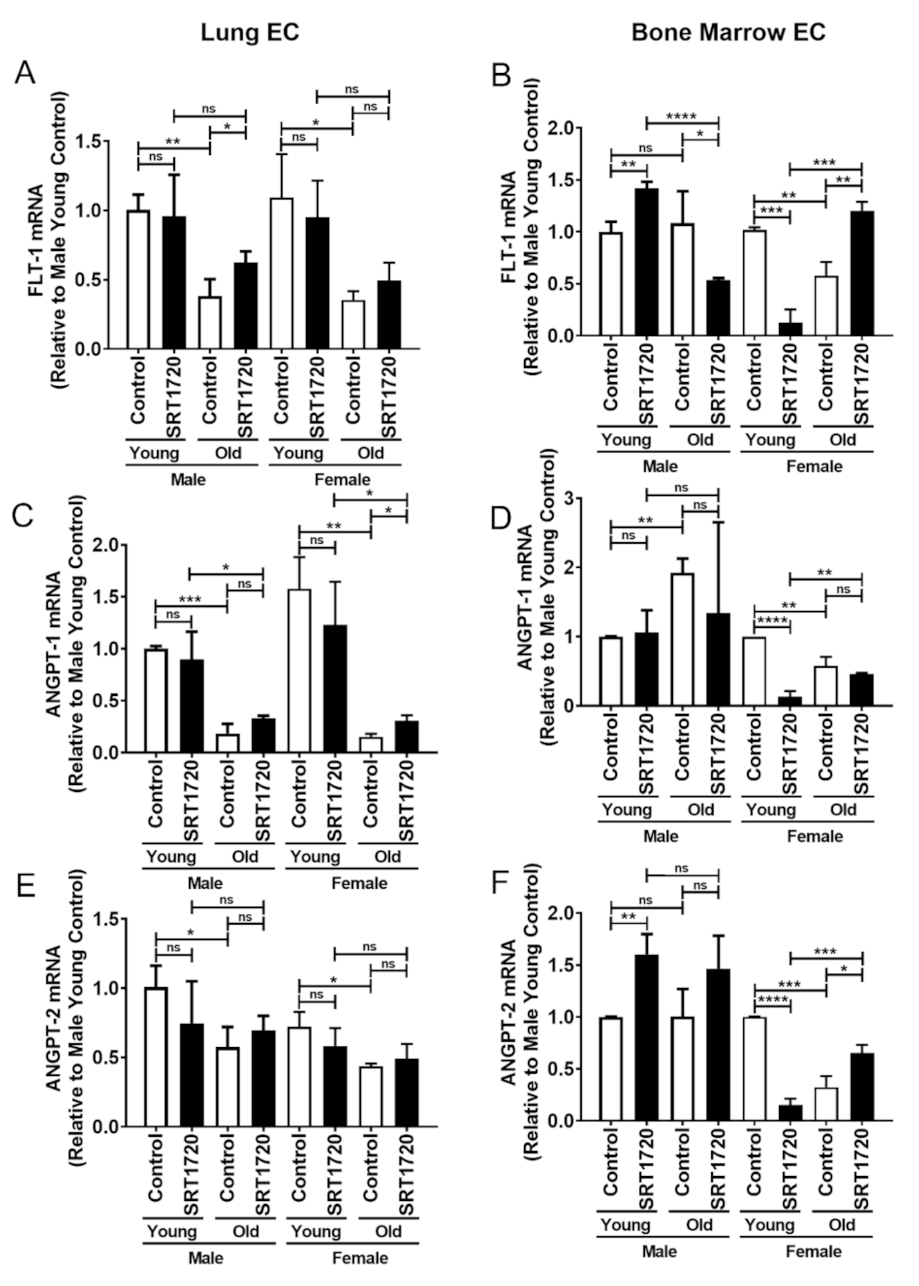
2.3. Gene Expression in Endothelial Cells
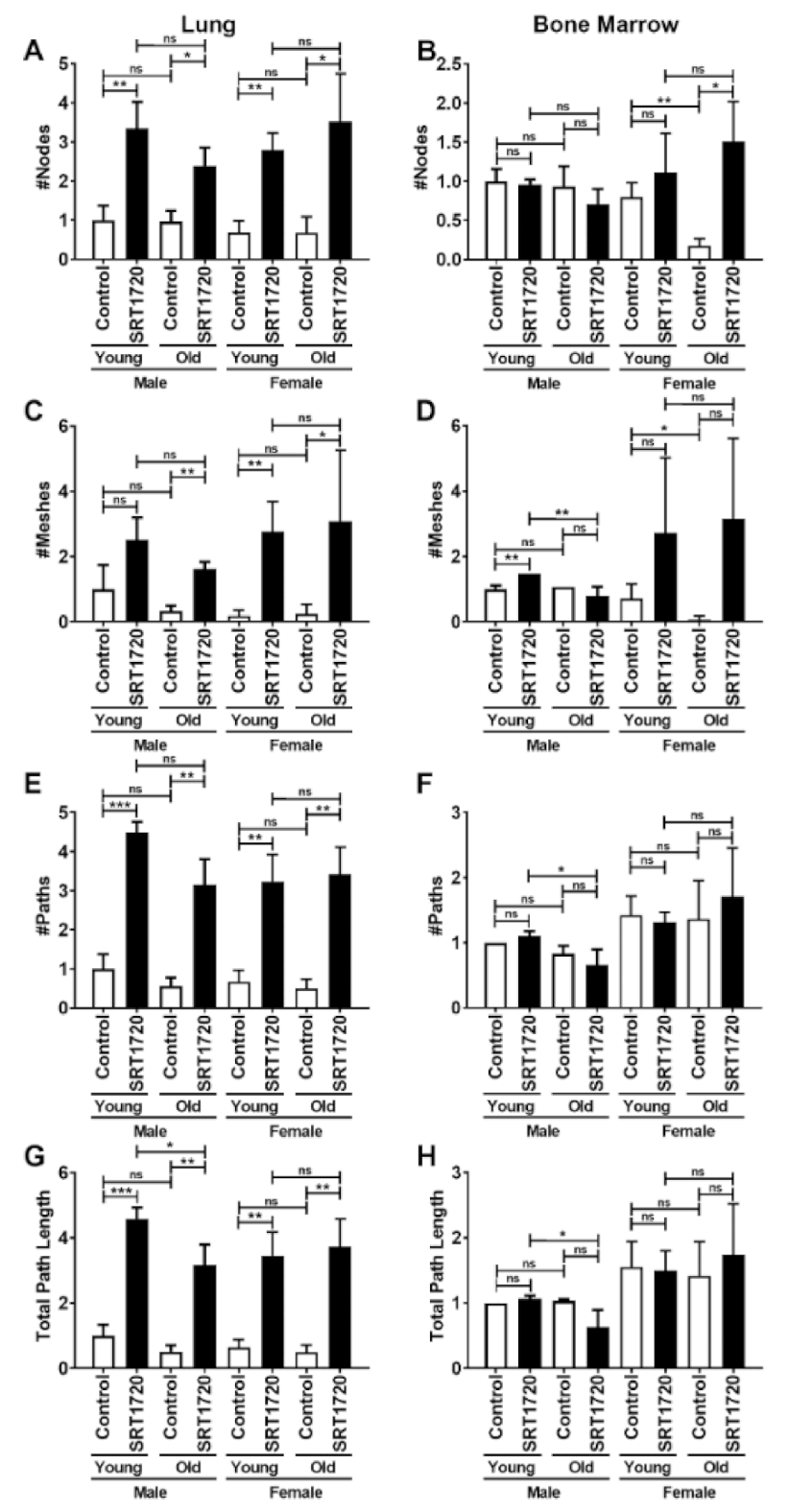
2.4. Angiogenic Potential of Endothelial Cells by Vessel-like Tube Formation
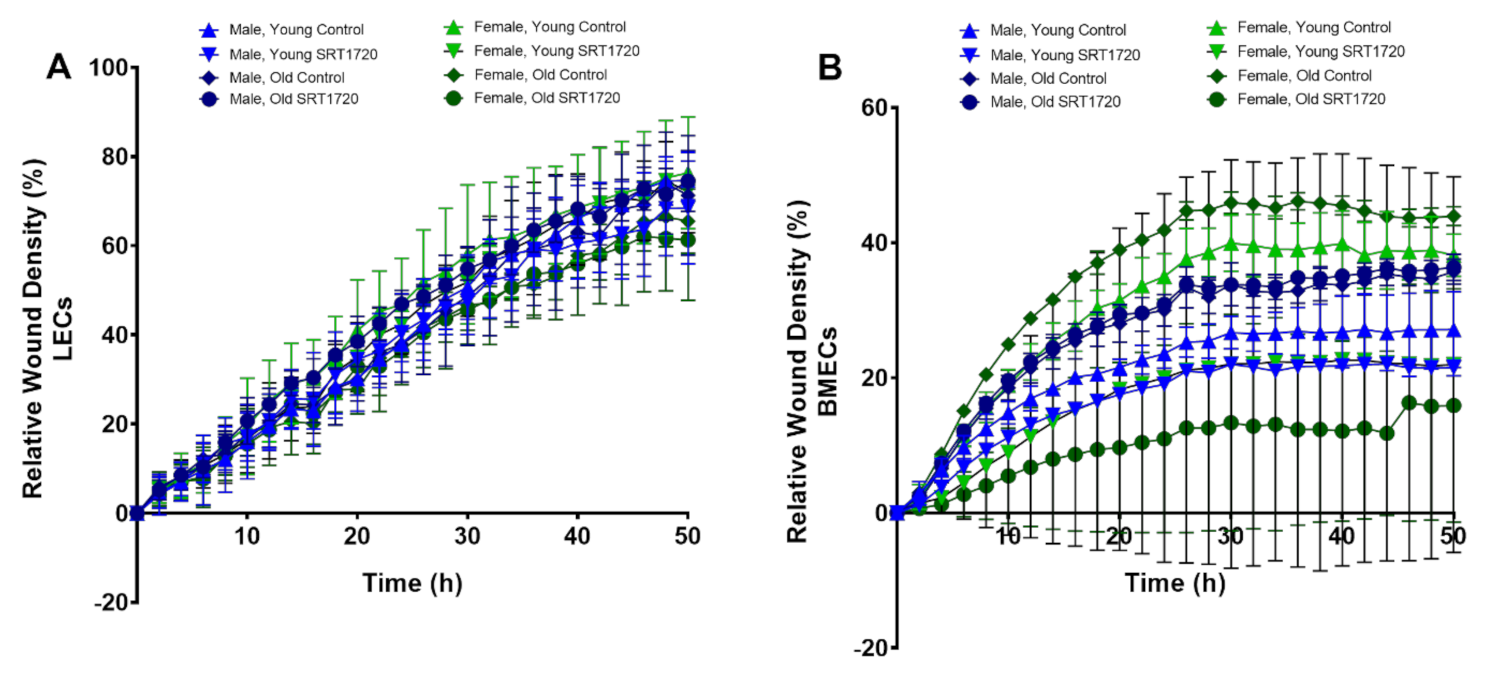
2.5. Migration of Endothelial Cells
3. Discussion
4. Materials and Methods
4.1. Mouse Model
4.2. Isolation of Mouse Primary Endothelial Cells
4.3. In Vitro SRT1720 Treatment
4.4. Proliferation Assay
4.5. Gene Expression Analysis
4.6. Vessel-like Structure Formation Assay
4.7. Wound Migration Assay
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wright, N.C.; Looker, A.C.; Saag, K.G.; Curtis, J.R.; Delzell, E.S.; Randall, S.; Dawson-Hughes, B. The recent prevalence of osteoporosis and low bone mass in the United States based on bone mineral density at the femoral neck or lumbar spine. J. Bone Miner. Res. 2014, 29, 2520–2526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burge, R.; Dawson-Hughes, B.; Solomon, D.H.; Wong, J.B.; King, A.; Tosteson, A. Incidence and economic burden of osteoporosis-related fractures in the United States, 2005-2025. J. Bone Miner. Res. 2007, 22, 465–475. [Google Scholar] [CrossRef]
- Mears, S.C.; Kates, S.L. A Guide to Improving the Care of Patients with Fragility Fractures, Edition 2. Geriatr. Orthop. Surg. Rehabil. 2015, 6, 58–120. [Google Scholar] [CrossRef]
- Iorio, R.; Healy, W.L.; Lemos, D.W.; Appleby, D.; Lucchesi, C.A.; Saleh, K.J. Displaced femoral neck fractures in the elderly: Outcomes and cost effectiveness. Clin. Orthop. Relat. Res. 2001, 383, 229–242. [Google Scholar] [CrossRef] [PubMed]
- Kos, N.; Burger, H.; Vidmar, G. Mobility and functional outcomes after femoral neck fracture surgery in elderly patients: A comparison between hemiarthroplasty and internal fixation. Disabil. Rehabil. 2011, 33, 2264–2271. [Google Scholar] [CrossRef] [PubMed]
- Puvanesarajah, V.; Amin, R.; Qureshi, R.; Shafiq, B.; Stein, B.; Hassanzadeh, H.; Yarboro, S. Outcomes following surgical management of femoral neck fractures in elderly dialysis-dependent patients. Arch. Orthop. Trauma Surg. 2018, 138, 757–764. [Google Scholar] [CrossRef] [PubMed]
- Wagner, D.R.; Karnik, S.; Gunderson, Z.J.; Nielsen, J.J.; Fennimore, A.; Promer, H.J.; Lowery, J.W.; Loghmani, M.T.; Low, P.S.; McKinley, T.O.; et al. Dysfunctional stem and progenitor cells impair fracture healing with age. World J. Stem Cells 2019, 11, 281–296. [Google Scholar] [CrossRef] [PubMed]
- Burkhardt, R.; Kettner, G.; Bohm, W.; Schmidmeier, M.; Schlag, R.; Frisch, B.; Mallmann, B.; Eisenmenger, W.; Gilg, T. Changes in trabecular bone, hematopoiesis and bone marrow vessels in aplastic anemia, primary osteoporosis, and old age: A comparative histomorphometric study. Bone 1987, 8, 157–164. [Google Scholar] [CrossRef] [Green Version]
- Lu, C.; Hansen, E.; Sapozhnikova, A.; Hu, D.; Miclau, T.; Marcucio, R.S. Effect of age on vascularization during fracture repair. J. Orthop. Res. 2008, 26, 1384–1389. [Google Scholar] [CrossRef] [Green Version]
- Lu, C.; Miclau, T.; Hu, D.; Hansen, E.; Tsui, K.; Puttlitz, C.; Marcucio, R.S. Cellular basis for age-related changes in fracture repair. J. Orthop. Res. 2005, 23, 1300–1307. [Google Scholar] [CrossRef] [PubMed]
- Watson, E.C.; Adams, R.H. Biology of Bone: The Vasculature of the Skeletal System. Cold Spring Harb. Perspect. Med. 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Townsley, M.I. Structure and composition of pulmonary arteries, capillaries, and veins. Compr. Physiol. 2012, 2, 675–709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, H.F.; Zhai, C.; Yan, X.L.; Zhao, D.D.; Wang, J.X.; Zeng, Q.; Chen, L.; Nan, X.; He, L.J.; Li, S.T.; et al. SIRT1 is required for long-term growth of human mesenchymal stem cells. J. Mol. Med. 2012, 90, 389–400. [Google Scholar] [CrossRef] [PubMed]
- Dryden, S.C.; Nahhas, F.A.; Nowak, J.E.; Goustin, A.S.; Tainsky, M.A. Role for human SIRT2 NAD-dependent deacetylase activity in control of mitotic exit in the cell cycle. Mol. Cell Biol. 2003, 23, 3173–3185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaeberlein, M.; McVey, M.; Guarente, L. The SIR2/3/4 complex and SIR2 alone promote longevity in Saccharomyces cerevisiae by two different mechanisms. Genes Dev. 1999, 13, 2570–2580. [Google Scholar] [CrossRef] [Green Version]
- Son, M.J.; Kwon, Y.; Son, T.; Cho, Y.S. Restoration of Mitochondrial NAD(+) Levels Delays Stem Cell Senescence and Facilitates Reprogramming of Aged Somatic Cells. Stem Cells 2016, 34, 2840–2851. [Google Scholar] [CrossRef]
- Anwar, T.; Khosla, S.; Ramakrishna, G. Increased expression of SIRT2 is a novel marker of cellular senescence and is dependent on wild type p53 status. Cell Cycle 2016, 15, 1883–1897. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Xie, J.-J.; Jin, M.-Y.; Gu, Y.-T.; Wu, C.-C.; Guo, W.-J.; Yan, Y.-Z.; Zhang, Z.-J.; Wang, J.-L.; Zhang, X.-L.; et al. Sirt6 overexpression suppresses senescence and apoptosis of nucleus pulposus cells by inducing autophagy in a model of intervertebral disc degeneration. Cell Death Dis. 2018, 9, 56. [Google Scholar] [CrossRef]
- Potente, M.; Ghaeni, L.; Baldessari, D.; Mostoslavsky, R.; Rossig, L.; Dequiedt, F.; Haendeler, J.; Mione, M.; Dejana, E.; Alt, F.W.; et al. SIRT1 controls endothelial angiogenic functions during vascular growth. Genes Dev. 2007, 21, 2644–2658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donato, A.J.; Magerko, K.A.; Lawson, B.R.; Durrant, J.R.; Lesniewski, L.A.; Seals, D.R. SIRT-1 and vascular endothelial dysfunction with ageing in mice and humans. J. Physiol. 2011, 589, 4545–4554. [Google Scholar] [CrossRef]
- Gano, L.B.; Donato, A.J.; Pasha, H.M.; Hearon, C.M., Jr.; Sindler, A.L.; Seals, D.R. The SIRT1 activator SRT1720 reverses vascular endothelial dysfunction, excessive superoxide production, and inflammation with aging in mice. Am. J. Physiol. Heart Circ. Physiol. 2014, 307, H1754–H1763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Onofrio, N.; Servillo, L.; Giovane, A.; Casale, R.; Vitiello, M.; Marfella, R.; Paolisso, G.; Balestrieri, M.L. Ergothioneine oxidation in the protection against high-glucose induced endothelial senescence: Involvement of SIRT1 and SIRT6. Free Radic. Biol. Med. 2016, 96, 211–222. [Google Scholar] [CrossRef]
- Zarzuelo, M.J.; Lopez-Sepulveda, R.; Sanchez, M.; Romero, M.; Gomez-Guzman, M.; Ungvary, Z.; Perez-Vizcaino, F.; Jimenez, R.; Duarte, J. SIRT1 inhibits NADPH oxidase activation and protects endothelial function in the rat aorta: Implications for vascular aging. Biochem. Pharmacol. 2013, 85, 1288–1296. [Google Scholar] [CrossRef] [PubMed]
- Cohen-Kfir, E.; Artsi, H.; Levin, A.; Abramowitz, E.; Bajayo, A.; Gurt, I.; Zhong, L.; D’Urso, A.; Toiber, D.; Mostoslavsky, R.; et al. Sirt1 is a regulator of bone mass and a repressor of Sost encoding for sclerostin, a bone formation inhibitor. Endocrinology 2011, 152, 4514–4524. [Google Scholar] [CrossRef] [PubMed]
- Edwards, J.R.; Perrien, D.S.; Fleming, N.; Nyman, J.S.; Ono, K.; Connelly, L.; Moore, M.M.; Lwin, S.T.; Yull, F.E.; Mundy, G.R.; et al. Silent information regulator (Sir)T1 inhibits NF-kappaB signaling to maintain normal skeletal remodeling. J. Bone Miner. Res. 2013, 28, 960–969. [Google Scholar] [CrossRef]
- Herranz, D.; Munoz-Martin, M.; Canamero, M.; Mulero, F.; Martinez-Pastor, B.; Fernandez-Capetillo, O.; Serrano, M. Sirt1 improves healthy ageing and protects from metabolic syndrome-associated cancer. Nat. Commun. 2010, 1, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mercken, E.M.; Mitchell, S.J.; Martin-Montalvo, A.; Minor, R.K.; Almeida, M.; Gomes, A.P.; Scheibye-Knudsen, M.; Palacios, H.H.; Licata, J.J.; Zhang, Y.; et al. SRT2104 extends survival of male mice on a standard diet and preserves bone and muscle mass. Aging Cell 2014, 13, 787–796. [Google Scholar] [CrossRef] [PubMed]
- Pearson, K.J.; Baur, J.A.; Lewis, K.N.; Peshkin, L.; Price, N.L.; Labinskyy, N.; Swindell, W.R.; Kamara, D.; Minor, R.K.; Perez, E.; et al. Resveratrol delays age-related deterioration and mimics transcriptional aspects of dietary restriction without extending life span. Cell Metab. 2008, 8, 157–168. [Google Scholar] [CrossRef] [Green Version]
- Elseweidy, M.M.; El-Swefy, S.E.; Shaheen, M.A.; Baraka, N.M.; Hammad, S.K. Effect of resveratrol and mesenchymal stem cell monotherapy and combined treatment in management of osteoporosis in ovariectomized rats: Role of SIRT1/FOXO3a and Wnt/beta-catenin pathways. Arch. Biochem. Biophys. 2021, 703, 108856. [Google Scholar] [CrossRef]
- Hong, W.; Wei, Z.; Qiu, Z.; Li, Z.; Fu, C.; Ye, Z.; Xu, X. Atorvastatin promotes bone formation in aged apoE(-/-) mice through the Sirt1-Runx2 axis. J. Orthop. Surg. Res. 2020, 15, 303. [Google Scholar] [CrossRef] [PubMed]
- Ameen, O.; Yassien, R.I.; Naguib, Y.M. Activation of FoxO1/SIRT1/RANKL/OPG pathway may underlie the therapeutic effects of resveratrol on aging-dependent male osteoporosis. BMC Musculoskelet Disord. 2020, 21, 375. [Google Scholar] [CrossRef] [PubMed]
- Zainabadi, K.; Liu, C.J.; Caldwell, A.L.M.; Guarente, L. SIRT1 is a positive regulator of in vivo bone mass and a therapeutic target for osteoporosis. PLoS ONE 2017, 12, e0185236. [Google Scholar] [CrossRef]
- El-Haj, M.; Gurt, I.; Cohen-Kfir, E.; Dixit, V.; Artsi, H.; Kandel, L.; Yakubovsky, O.; Safran, O.; Dresner-Pollak, R. Reduced Sirtuin1 expression at the femoral neck in women who sustained an osteoporotic hip fracture. Osteoporos. Int. 2016, 27, 2373–2378. [Google Scholar] [CrossRef] [PubMed]
- Nesmith, J.E.; Chappell, J.C.; Cluceru, J.G.; Bautch, V.L. Blood vessel anastomosis is spatially regulated by Flt1 during angiogenesis. Development 2017, 144, 889–896. [Google Scholar] [CrossRef] [Green Version]
- Ferrara, N.; Gerber, H.P.; LeCouter, J. The biology of VEGF and its receptors. Nat. Med. 2003, 9, 669–676. [Google Scholar] [CrossRef] [PubMed]
- Jeansson, M.; Gawlik, A.; Anderson, G.; Li, C.; Kerjaschki, D.; Henkelman, M.; Quaggin, S.E. Angiopoietin-1 is essential in mouse vasculature during development and in response to injury. J. Clin. Invest. 2011, 121, 2278–2289. [Google Scholar] [CrossRef] [Green Version]
- Maisonpierre, P.C.; Suri, C.; Jones, P.F.; Bartunkova, S.; Wiegand, S.J.; Radziejewski, C.; Compton, D.; McClain, J.; Aldrich, T.H.; Papadopoulos, N.; et al. Angiopoietin-2, a natural antagonist for Tie2 that disrupts in vivo angiogenesis. Science 1997, 277, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Ungvari, Z.; Tarantini, S.; Kiss, T.; Wren, J.D.; Giles, C.B.; Griffin, C.T.; Murfee, W.L.; Pacher, P.; Csiszar, A. Endothelial dysfunction and angiogenesis impairment in the ageing vasculature. Nat. Rev. Cardiol. 2018, 15, 555–565. [Google Scholar] [CrossRef] [PubMed]
- Lahteenvuo, J.; Rosenzweig, A. Effects of aging on angiogenesis. Circ. Res. 2012, 110, 1252–1264. [Google Scholar] [CrossRef] [Green Version]
- ClinicalTrials.gov [Internet]. Bethesda (MD): National Library of Medicine (US). Available online: https://clinicaltrials.gov/ct2/results?cond=&term=SIRT1&cntry=&state=&city=&dist= (accessed on 10 September 2021).
- Dai, H.; Sinclair, D.A.; Ellis, J.L.; Steegborn, C. Sirtuin activators and inhibitors: Promises, achievements, and challenges. Pharmacol. Ther. 2018, 188, 140–154. [Google Scholar] [CrossRef] [PubMed]
- Elbaz, A.; Rivas, D.; Duque, G. Effect of estrogens on bone marrow adipogenesis and Sirt1 in aging C57BL/6J mice. Biogerontology 2009, 10, 747–755. [Google Scholar] [CrossRef] [PubMed]
- Artsi, H.; Cohen-Kfir, E.; Gurt, I.; Shahar, R.; Bajayo, A.; Kalish, N.; Bellido, T.M.; Gabet, Y.; Dresner-Pollak, R. The Sirtuin1 activator SRT3025 down-regulates sclerostin and rescues ovariectomy-induced bone loss and biomechanical deterioration in female mice. Endocrinology 2014, 155, 3508–3515. [Google Scholar] [CrossRef]
- Lipphardt, M.; Dihazi, H.; Muller, G.A.; Goligorsky, M.S. Fibrogenic Secretome of Sirtuin 1-Deficient Endothelial Cells: Wnt, Notch and Glycocalyx Rheostat. Front. Physiol. 2018, 9, 1325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mulcrone, P.L.; Campbell, J.P.; Clement-Demange, L.; Anbinder, A.L.; Merkel, A.R.; Brekken, R.A.; Sterling, J.A.; Elefteriou, F. Skeletal Colonization by Breast Cancer Cells Is Stimulated by an Osteoblast and beta2AR-Dependent Neo-Angiogenic Switch. J. Bone Miner. Res. 2017, 32, 1442–1454. [Google Scholar] [CrossRef]
- Sekiguchi, H.; Ii, M.; Jujo, K.; Yokoyama, A.; Hagiwara, N.; Asahara, T. Improved culture-based isolation of differentiating endothelial progenitor cells from mouse bone marrow mononuclear cells. PLoS ONE 2011, 6, e28639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dadwal, U.C.; Bhatti, F.U.R.; Awosanya, O.D.; Nagaraj, R.U.; Perugini, A.J., 3rd; Sun, S.; Valuch, C.R.; de Andrade Staut, C.; Mendenhall, S.K.; Tewari, N.P.; et al. The effects of bone morphogenetic protein 2 and thrombopoietin treatment on angiogenic properties of endothelial cells derived from the lung and bone marrow of young and aged, male and female mice. FASEB J. 2021, 35, e21840. [Google Scholar] [CrossRef] [PubMed]
- Dadwal, U.C.; Staut, C.A.; Tewari, N.P.; Awosanya, O.D.; Mendenhall, S.K.; Valuch, C.R.; Nagaraj, R.U.; Blosser, R.J.; Li, J.; Kacena, M.A. Effects of diet, BMP-2 treatment, and femoral skeletal injury on endothelial cells derived from the ipsilateral and contralateral limbs. J. Orthop. Res. 2021. online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Bhatti, F.U.R.; Dadwal, U.C.; Valuch, C.R.; Tewari, N.P.; Awosanya, O.D.; de Andrade Staut, C.; Sun, S.; Mendenhall, S.K.; Perugini, A.J., 3rd; Nagaraj, R.U.; et al. The effects of high fat diet, bone healing, and BMP-2 treatment on endothelial cell growth and function. Bone 2021, 146, 115883. [Google Scholar] [CrossRef]
- Longair, M.H.; Baker, D.A.; Armstrong, J.D. Simple Neurite Tracer: Open source software for reconstruction, visualization and analysis of neuronal processes. Bioinformatics 2011, 27, 2453–2454. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Niu, N.; Xu, S.; Jin, Z.G. A simple protocol for isolating mouse lung endothelial cells. Sci. Rep. 2019, 9, 1458. [Google Scholar] [CrossRef] [Green Version]
- Rueden, C.T.; Schindelin, J.; Hiner, M.C.; DeZonia, B.E.; Walter, A.E.; Arena, E.T.; Eliceiri, K.W. ImageJ2: ImageJ for the next generation of scientific image data. BMC Bioinform. 2017, 18, 529. [Google Scholar] [CrossRef] [PubMed]
- Pearson, H.B.; Mason, D.E.; Kegelman, C.D.; Zhao, L.; Dawahare, J.H.; Kacena, M.A.; Boerckel, J.D. Effects of Bone Morphogenetic Protein-2 on Neovascularization During Large Bone Defect Regeneration. Tissue Eng Part. A 2019, 25, 1623–1634. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Outcome Measure | Results | Age | Treatment | |||||
|---|---|---|---|---|---|---|---|---|
| Male | Female | Control | SRT1720 | |||||
| Young | Old | Young | Old | |||||
| Proliferation | ▲ | X | X | X | X | X | ||
| RT PCR | FLT-1 | ▼ | X | X | X | |||
| ▲ | X | X | ||||||
| ANGPT1 | ▼ | X | X | X | ||||
| ▲ | X | X | ||||||
| ANGPT2 | ▼ | X | X | X | ||||
| Vessel-like Structure Formation | Number of nodes | ▲ | X | X | X | X | X | |
| Number of meshes | ▲ | X | X | X | X | X | ||
| Number of paths | ▲ | X | X | X | X | X | ||
| total path length | ▲ | X | X | X | X | X | ||
| Migration | ⁃⁃⁃ | |||||||
| Outcome Measure | Results | Age | Treatment | |||||
|---|---|---|---|---|---|---|---|---|
| Male | Female | Control | SRT1720 | |||||
| Young | Old | Young | Old | |||||
| Proliferation | ▲ | X | X | |||||
| ▲ | X | X | ||||||
| RT PCR | FLT-1 | ▼ | X | X | ||||
| ▲ | X | X | X | |||||
| ▼ | X | X | X | |||||
| ANGPT1 | ▲ | X | X | |||||
| ▼ | X | X | ||||||
| ▼ | X | X | ||||||
| ANGPT2 | ▼ | X | X | |||||
| ▲ | X | X | X | |||||
| ▼ | X | X | ||||||
| Vessel-like Structure Formation | Number of nodes | ▼ | X | X | ||||
| ▲ | X | X | ||||||
| Number of meshes | ▼ | X | X | |||||
| ▲ | X | X | ||||||
| Number of paths | ⁃⁃⁃ | |||||||
| total path length | ⁃⁃⁃ | |||||||
| Migration | ▲ | X | X | X | ||||
| ▼ | X | X | X | |||||
| Target | 5′-3′ Sequence |
|---|---|
| GAPDH | F: CGTGGGGCTGCCCAGAACAT R: TCTCCAGGCGGCACGTCAGA |
| CD31 | F: ACGCTGGTGCTCTATGCAAG R: TCAGTTGCTGCCCATTCATCA |
| FLT-1 | F: CCACCTCTCTATCCGCTGG R: ACCAATGTGCTAACCGTCTTATT |
| ANGPT1 | F: CACATAGGGTGCAGCAACCA R: CGTCGTGTTCTGGAAGAATGA |
| ANGPT2 | F: CCTCGACTACGACGACTCAGT R: TCTGCACCACATTCTGTTGGA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dadwal, U.C.; Bhatti, F.U.R.; Awosanya, O.D.; Staut, C.d.A.; Nagaraj, R.U.; Perugini, A.J., III; Tewari, N.P.; Valuch, C.R.; Sun, S.; Mendenhall, S.K.; et al. The Effects of SRT1720 Treatment on Endothelial Cells Derived from the Lung and Bone Marrow of Young and Aged, Male and Female Mice. Int. J. Mol. Sci. 2021, 22, 11097. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222011097
Dadwal UC, Bhatti FUR, Awosanya OD, Staut CdA, Nagaraj RU, Perugini AJ III, Tewari NP, Valuch CR, Sun S, Mendenhall SK, et al. The Effects of SRT1720 Treatment on Endothelial Cells Derived from the Lung and Bone Marrow of Young and Aged, Male and Female Mice. International Journal of Molecular Sciences. 2021; 22(20):11097. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222011097
Chicago/Turabian StyleDadwal, Ushashi Chand, Fazal Ur Rehman Bhatti, Olatundun Dupe Awosanya, Caio de Andrade Staut, Rohit U. Nagaraj, Anthony Joseph Perugini, III, Nikhil Prasad Tewari, Conner Riley Valuch, Seungyup Sun, Stephen Kyle Mendenhall, and et al. 2021. "The Effects of SRT1720 Treatment on Endothelial Cells Derived from the Lung and Bone Marrow of Young and Aged, Male and Female Mice" International Journal of Molecular Sciences 22, no. 20: 11097. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222011097