
Knockdown NRPC2, 3, 8, NRPABC1 and NRPABC2 Affects RNAPIII Activity and Disrupts Seed Development in Arabidopsis


Abstract
:1. Introduction
2. Results
2.1. Subunits of RNAPIII Are Encoded by 23 Genes in Arabidopsis
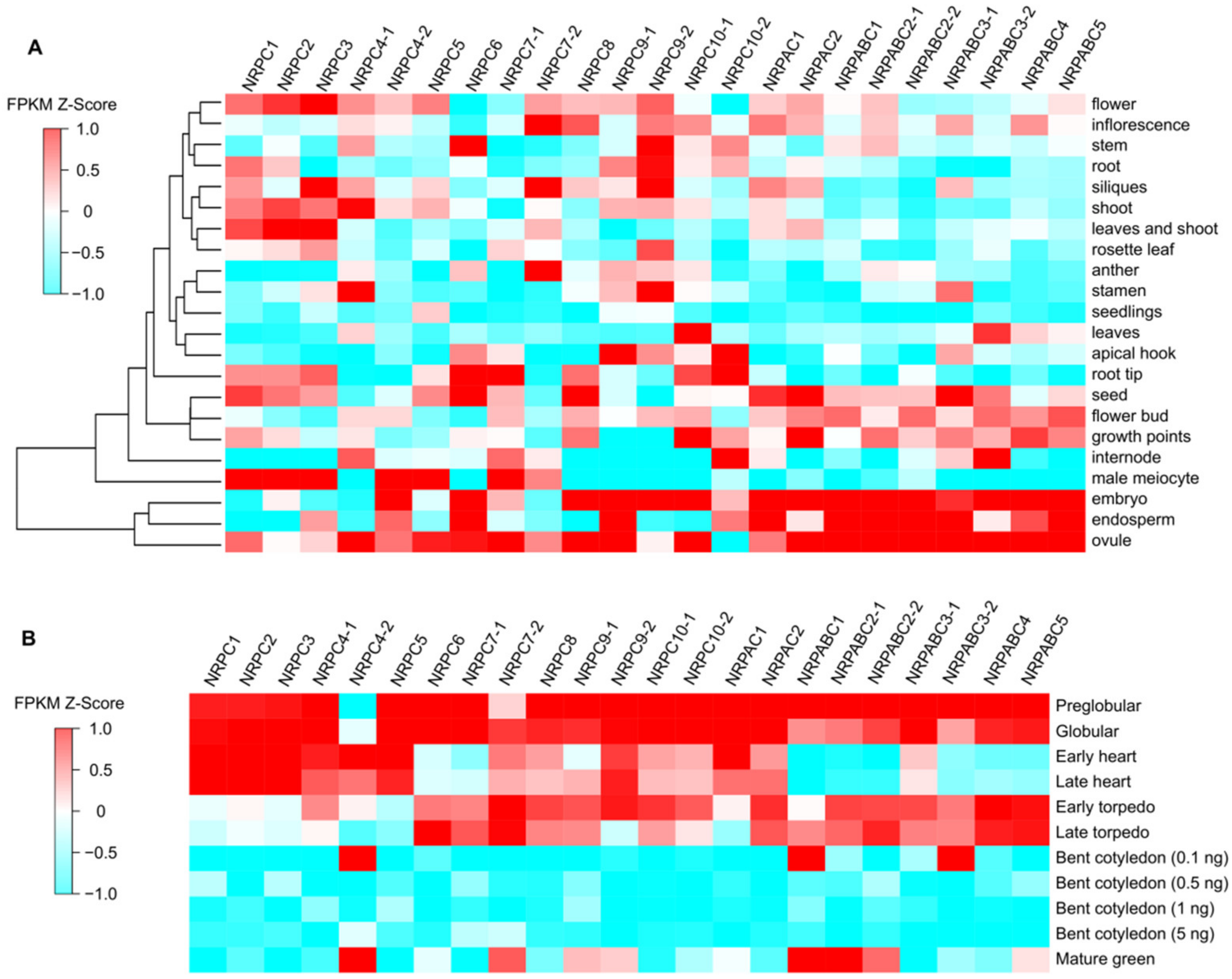
2.2. RNAPIII Subunits Have Different Expression Patterns during Seed Development
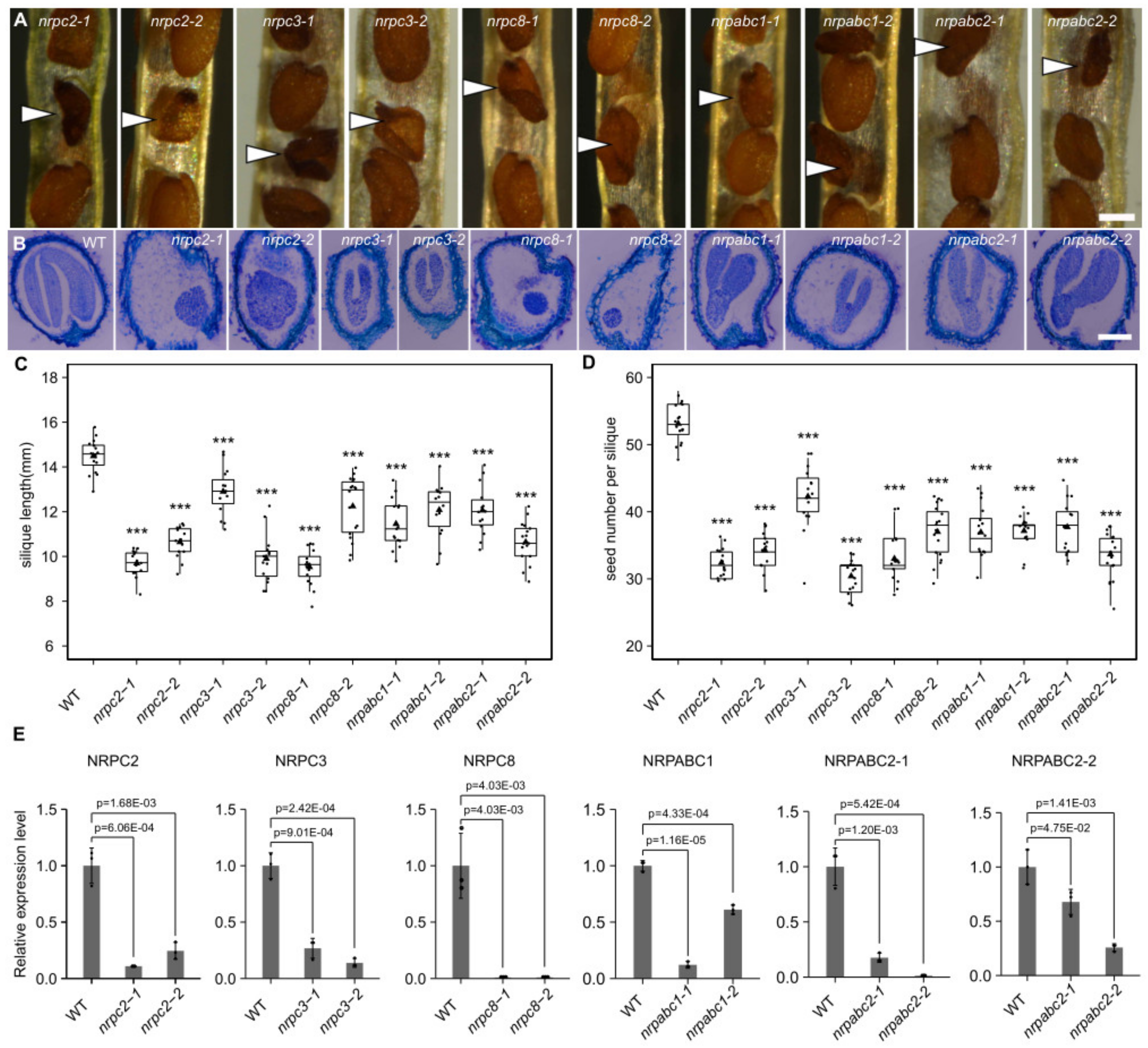
2.3. Knockdown of Five RNAPIII Subunits Leads to Seed Development Arrest at Different Stage in Arabidopsis
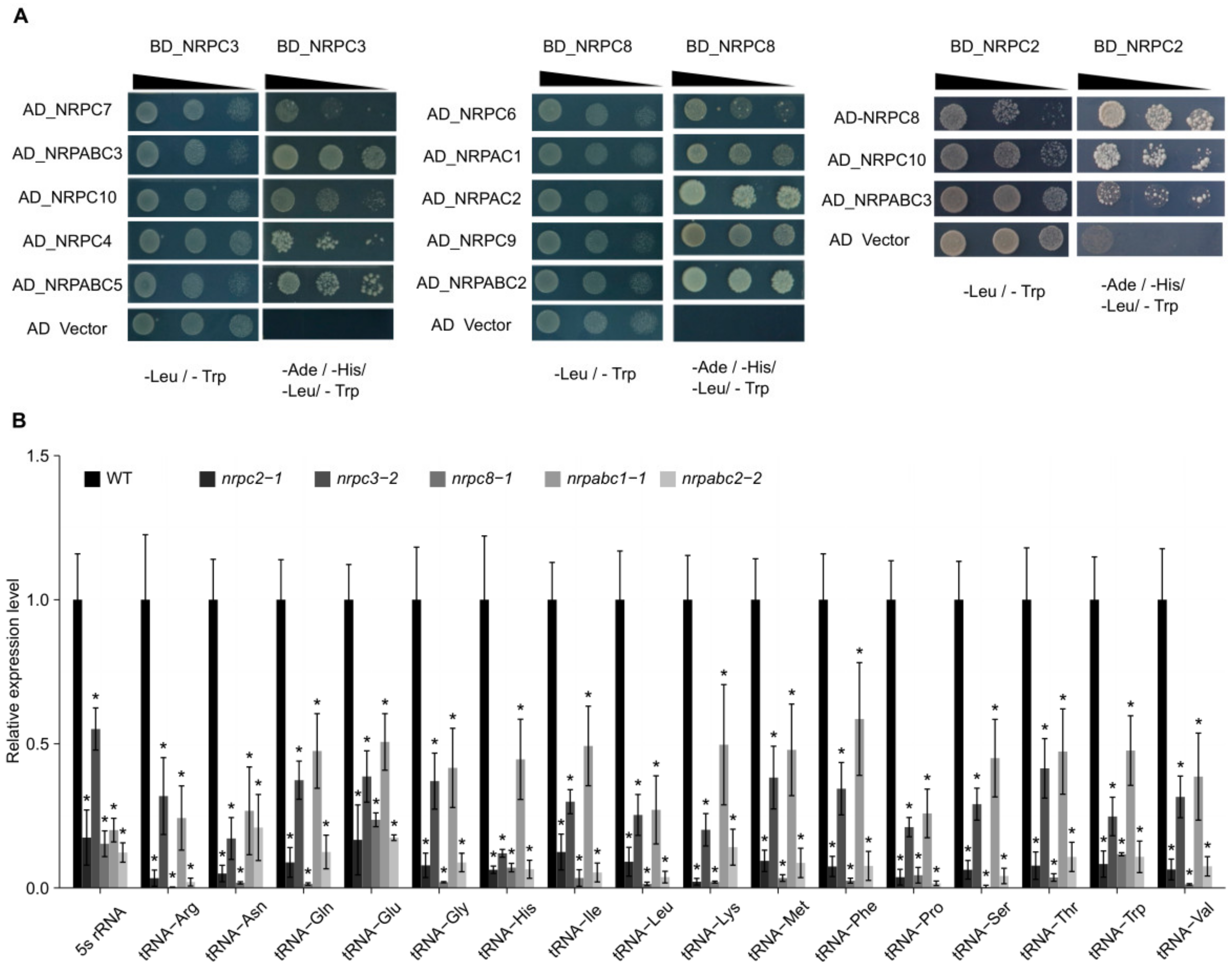
2.4. NRPC2, NRPC3, NRPC8, and NRPABC2 Interact with Other Subunits of the RNAPIII Complex
2.5. Expression of tRNAs and 5S rRNA Is Disrupted in nrpc2, nrpc3, nrpc8, nrpabc1, and nrpabc2 Mutant Seeds
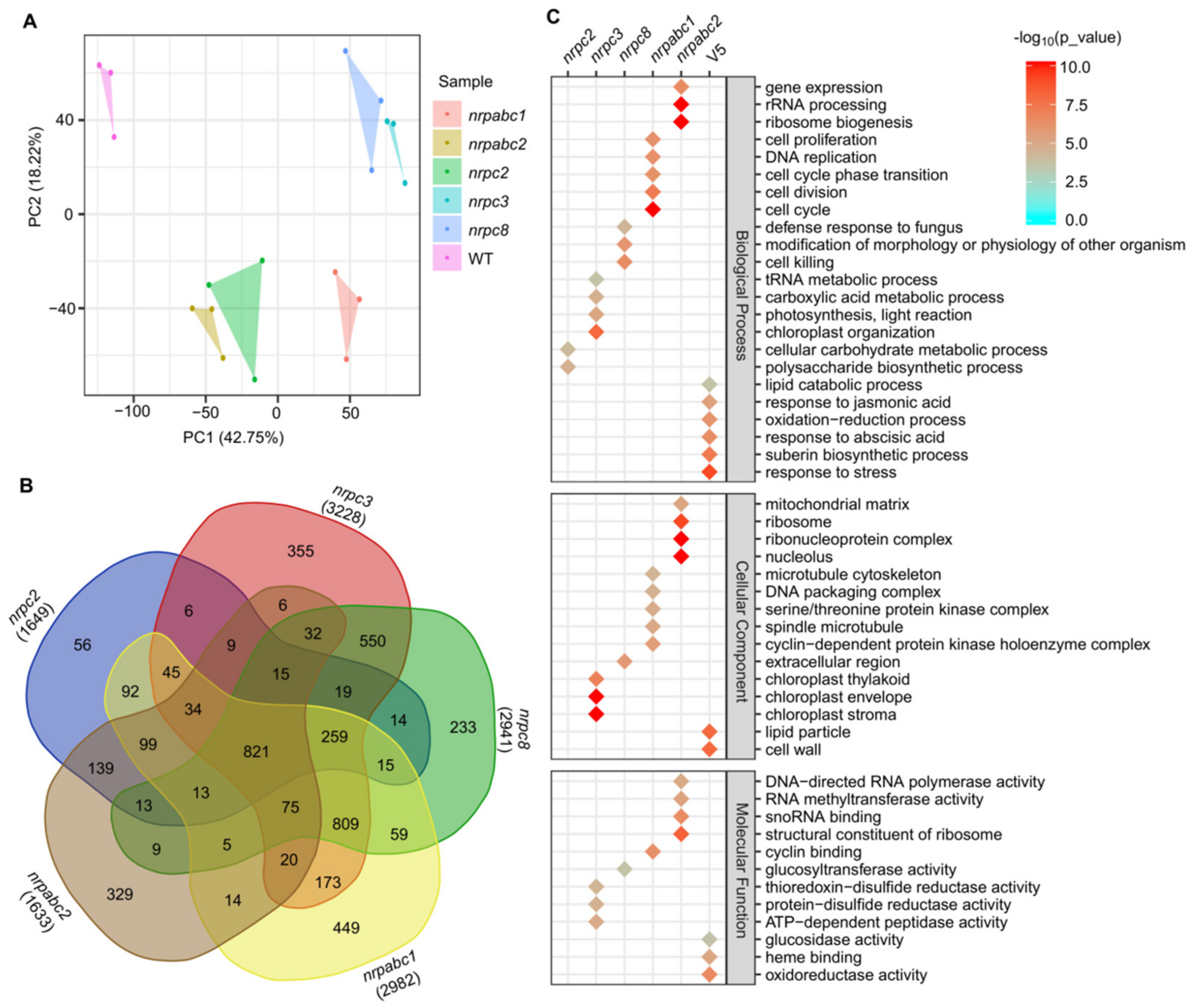
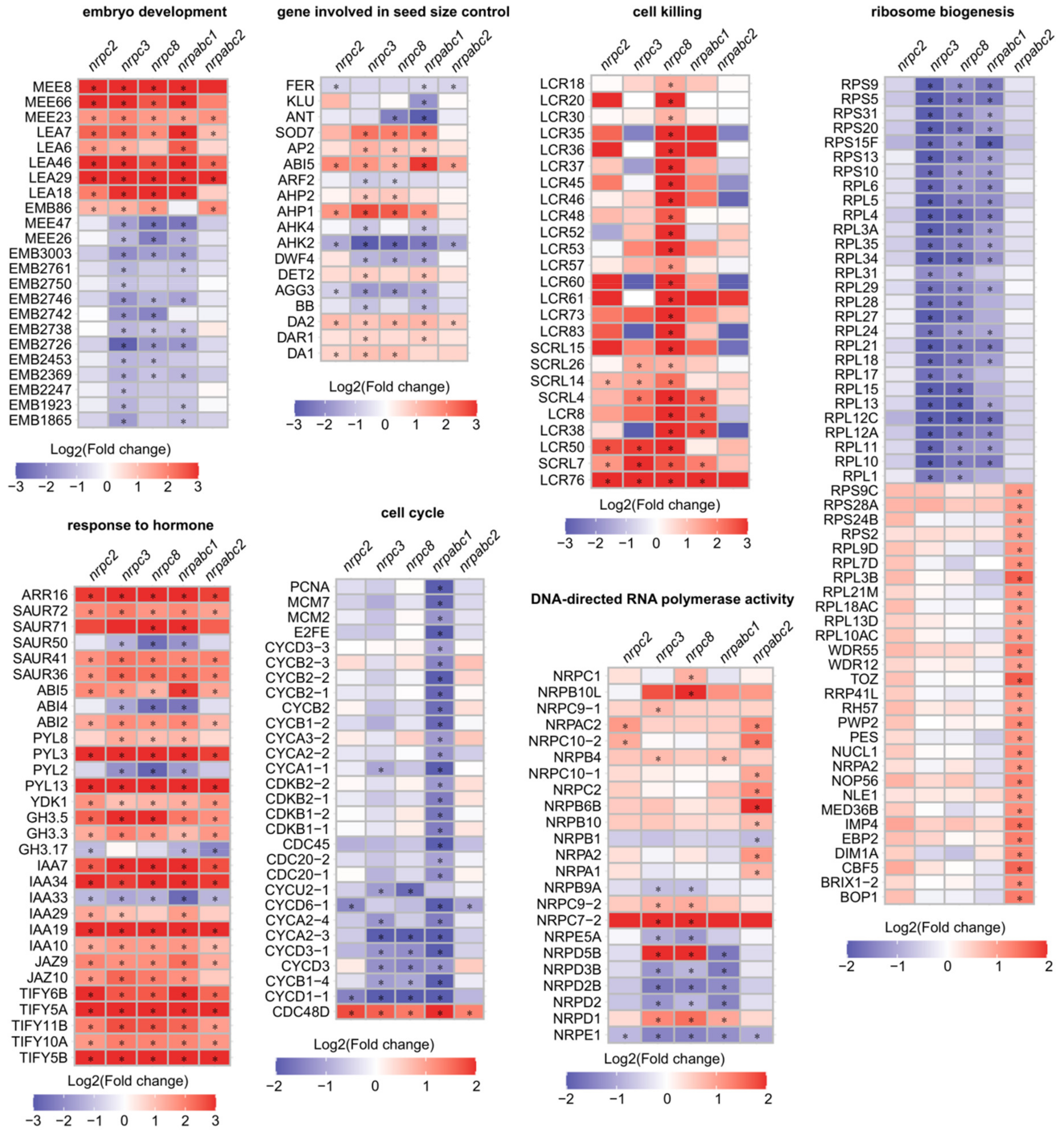
2.6. nrpc2, nrpc3, nrpc8, nrpabc1, and nrpabc2 Mutant Seeds Show Transcriptomic Alterations
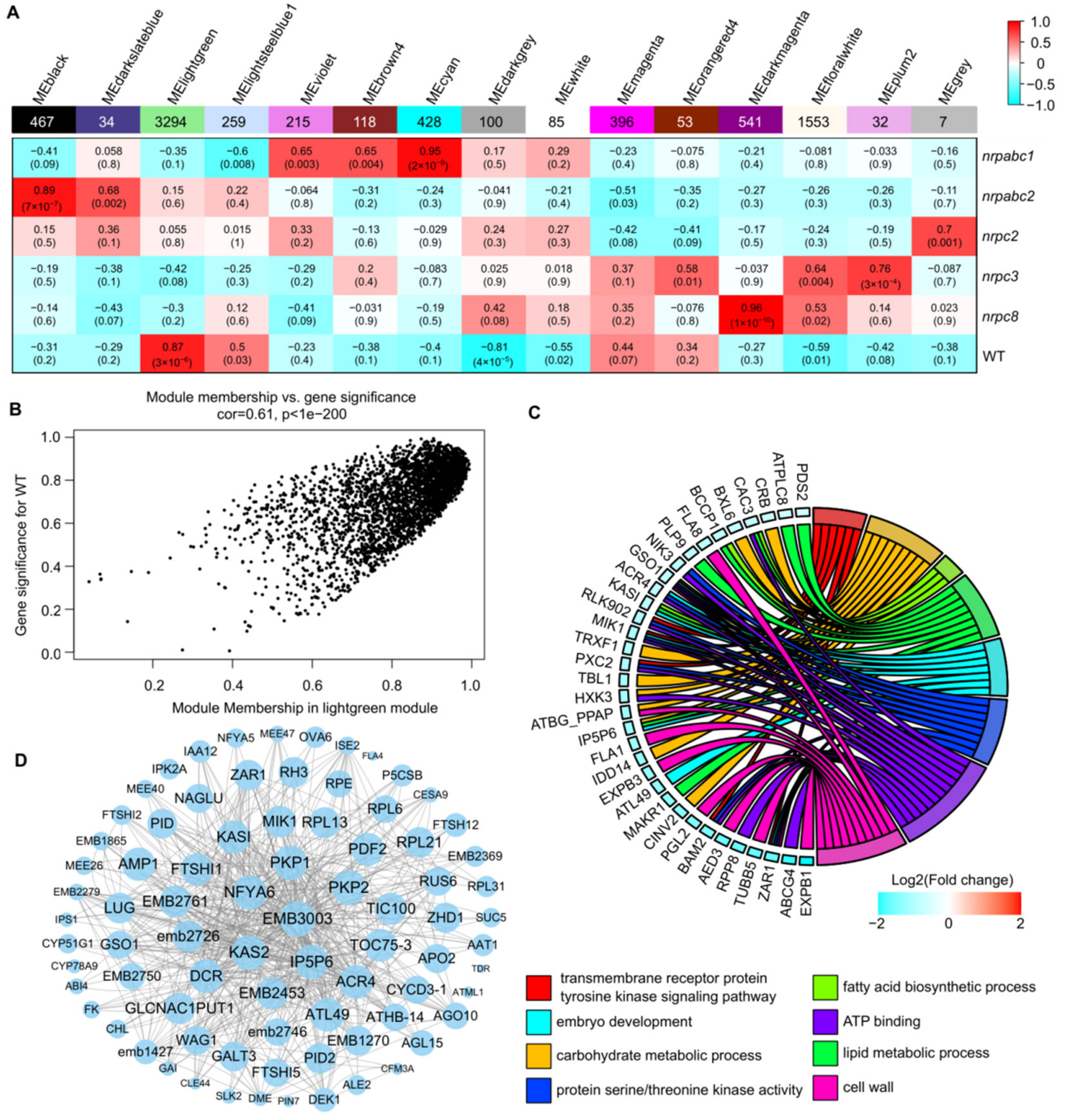
2.7. Co-Expression Network Analysis Identified Genes Related to Seed Development
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Cytological Observations
4.3. Yeast Two-Hybrid Assay
4.4. RNA Extraction and qRT-PCR
4.5. RNA-Seq Analysis
4.6. Weighted Gene Co-Expression Network Analysis (WGCNA)
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, N.; Li, Y. Signaling pathways of seed size control in plants. Curr. Opin. Plant Biol. 2016, 33, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Xu, R.; Li, Y. Molecular Networks of Seed Size Control in Plants. Annu. Rev. Plant Biol. 2019, 70, 435–463. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Shantharaj, D.; Kang, X.; Ni, M. Transcriptional and hormonal signaling control of Arabidopsis seed development. Curr. Opin. Plant Biol. 2010, 13, 611–620. [Google Scholar] [CrossRef] [PubMed]
- Abascal-Palacios, G.; Ramsay, E.P.; Beuron, F.; Morris, E.; Vannini, A. Structural basis of RNA polymerase III transcription initiation. Nature 2018, 553, 301–306. [Google Scholar] [CrossRef]
- Hoffmann, N.A.; Jakobi, A.J.; Moreno-Morcillo, M.; Glatt, S.; Kosinski, J.; Hagen, W.J.; Sachse, C.; Muller, C.W. Molecular structures of unbound and transcribing RNA polymerase III. Nature 2015, 528, 231–236. [Google Scholar] [CrossRef] [Green Version]
- Vannini, A.; Cramer, P. Conservation between the RNA polymerase I, II, and III transcription initiation machineries. Mol. Cell 2012, 45, 439–446. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.C.; Herzog, F.; Jennebach, S.; Lin, Y.C.; Pai, C.Y.; Aebersold, R.; Cramer, P.; Chen, H.T. RNA polymerase III subunit architecture and implications for open promoter complex formation. Proc. Natl. Acad. Sci. USA 2012, 109, 19232–19237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khoo, S.K.; Wu, C.C.; Lin, Y.C.; Lee, J.C.; Chen, H.T. Mapping the protein interaction network for TFIIB-related factor Brf1 in the RNA polymerase III preinitiation complex. Mol. Cell. Biol. 2014, 34, 551–559. [Google Scholar] [CrossRef] [Green Version]
- Thuillier, V.; Stettler, S.; Sentenac, A.; Thuriaux, P.; Werner, M. A mutation in the C31 subunit of Saccharomyces cerevisiae RNA polymerase III affects transcription initiation. EMBO J. 1995, 14, 351–359. [Google Scholar] [CrossRef]
- Khoo, S.K.; Wu, C.C.; Lin, Y.C.; Chen, H.T. The TFIIE-related Rpc82 subunit of RNA polymerase III interacts with the TFIIB-related transcription factor Brf1 and the polymerase cleft for transcription initiation. Nucleic Acids Res. 2018, 46, 1157–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, Y.Y.; Chen, H.T. Functions of the TFIIE-Related Tandem Winged-Helix Domain of Rpc34 in RNA Polymerase III Initiation and Elongation. Mol. Cell. Biol. 2018, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez-Tornero, C.; Bottcher, B.; Riva, M.; Carles, C.; Steuerwald, U.; Ruigrok, R.W.; Sentenac, A.; Muller, C.W.; Schoehn, G. Insights into transcription initiation and termination from the electron microscopy structure of yeast RNA polymerase III. Mol. Cell 2007, 25, 813–823. [Google Scholar] [CrossRef] [PubMed]
- Kassavetis, G.A.; Prakash, P.; Shim, E. The C53/C37 subcomplex of RNA polymerase III lies near the active site and participates in promoter opening. J. Biol. Chem. 2010, 285, 2695–2706. [Google Scholar] [CrossRef] [Green Version]
- Landrieux, E.; Alic, N.; Ducrot, C.; Acker, J.; Riva, M.; Carles, C. A subcomplex of RNA polymerase III subunits involved in transcription termination and reinitiation. EMBO J. 2006, 25, 118–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jasiak, A.J.; Armache, K.J.; Martens, B.; Jansen, R.P.; Cramer, P. Structural biology of RNA polymerase III: Subcomplex C17/25 X-ray structure and 11 subunit enzyme model. Mol. Cell 2006, 23, 71–81. [Google Scholar] [CrossRef]
- Coulombe, B.; Derksen, A.; La Piana, R.; Brais, B.; Gauthier, M.S.; Bernard, G. POLR3-related leukodystrophy: How do mutations affecting RNA polymerase III subunits cause hypomyelination? Fac. Rev. 2021, 10, 12. [Google Scholar] [CrossRef]
- Furukawa, S.; Kunii, M.; Doi, H.; Kondo, N.; Ogura, A.; Hirabuki, K.; Itoh, T.; Matsumoto, N.; Tanaka, F.; Katsuno, M.; et al. Case Report: Severe Osteoporosis and Preventive Therapy in RNA Polymerase III-Related Leukodystrophy. Front. Neurol. 2021, 12, 622355. [Google Scholar] [CrossRef]
- Ramanathan, A.; Weintraub, M.; Orlovetskie, N.; Serruya, R.; Mani, D.; Marcu, O.; Stepensky, P.; Weisblum, Y.; Djian, E.; Shaag, A.; et al. A mutation in POLR3E impairs antiviral immune response and RNA polymerase III. Proc. Natl. Acad. Sci. USA 2020, 117, 22113–22121. [Google Scholar] [CrossRef]
- Mann, C.; Micouin, J.Y.; Chiannilkulchai, N.; Treich, I.; Buhler, J.M.; Sentenac, A. RPC53 encodes a subunit of Saccharomyces cerevisiae RNA polymerase C (III) whose inactivation leads to a predominantly G1 arrest. Mol. Cell. Biol. 1992, 12, 4314–4326. [Google Scholar] [CrossRef] [Green Version]
- Yee, N.S.; Gong, W.; Huang, Y.; Lorent, K.; Dolan, A.C.; Maraia, R.J.; Pack, M. Mutation of RNA Pol III subunit rpc2/polr3b Leads to Deficiency of Subunit Rpc11 and disrupts zebrafish digestive development. PLoS Biol. 2007, 5, e312. [Google Scholar] [CrossRef] [Green Version]
- Kieckhaefer, J.; Lukovac, S.; Ye, D.Z.; Lee, D.; Beetler, D.J.; Pack, M.; Kaestner, K.H. The RNA polymerase III subunit Polr3b is required for the maintenance of small intestinal crypts in mice. Cell Mol. Gastroenterol. Hepatol. 2016, 2, 783–795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, H.; Qin, Y.; Xiao, Z.; Li, Q.; Yang, N.; Pan, Z.; Gong, D.; Sun, Q.; Yang, F.; Zhang, Z.; et al. Loss of Function of an RNA Polymerase III Subunit Leads to Impaired Maize Kernel Development. Plant Physiol. 2020, 184, 359–373. [Google Scholar] [CrossRef] [PubMed]
- Ream, T.S.; Haag, J.R.; Pontvianne, F.; Nicora, C.D.; Norbeck, A.D.; Pasa-Tolic, L.; Pikaard, C.S. Subunit compositions of Arabidopsis RNA polymerases I and III reveal Pol I- and Pol III-specific forms of the AC40 subunit and alternative forms of the C53 subunit. Nucleic Acids Res. 2015, 43, 4163–4178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Zhang, F.; Yu, Y.; Feng, L.; Jia, J.; Liu, B.; Li, B.; Guo, H.; Zhai, J. A Comprehensive Online Database for Exploring approximately 20,000 Public Arabidopsis RNA-Seq Libraries. Mol. Plant 2020, 13, 1231–1233. [Google Scholar] [CrossRef]
- Arimbasseri, A.G.; Maraia, R.J. Mechanism of Transcription Termination by RNA Polymerase III Utilizes a Non-template Strand Sequence-Specific Signal Element. Mol. Cell 2015, 58, 1124–1132. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Lunn, D.; Wallis, J.G.; Browse, J. Phosphatidylglycerol Composition Is Central to Chilling Damage in the Arabidopsis fab1 Mutant. Plant Physiol. 2020, 184, 1717–1730. [Google Scholar] [CrossRef]
- Baud, S.; Wuilleme, S.; Dubreucq, B.; de Almeida, A.; Vuagnat, C.; Lepiniec, L.; Miquel, M.; Rochat, C. Function of plastidial pyruvate kinases in seeds of Arabidopsis thaliana. Plant J. 2007, 52, 405–419. [Google Scholar] [CrossRef]
- Wang, A.; Hou, Q.; Si, L.; Huang, X.; Luo, J.; Lu, D.; Zhu, J.; Shangguan, Y.; Miao, J.; Xie, Y.; et al. The PLATZ Transcription Factor GL6 Affects Grain Length and Number in Rice. Plant Physiol. 2019, 180, 2077–2090. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Wang, J.; Ye, J.; Zheng, X.; Xiang, X.; Li, C.; Fu, M.; Wang, Q.; Zhang, Z.; Wu, Y. The Maize Imprinted Gene Floury3 Encodes a PLATZ Protein Required for tRNA and 5S rRNA Transcription through Interaction with RNA Polymerase III. Plant Cell 2017, 29, 2661–2675. [Google Scholar] [CrossRef] [Green Version]
- Disch, S.; Anastasiou, E.; Sharma, V.K.; Laux, T.; Fletcher, J.C.; Lenhard, M. The E3 ubiquitin ligase BIG BROTHER controls arabidopsis organ size in a dosage-dependent manner. Curr. Biol. 2006, 16, 272–279. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zheng, L.; Corke, F.; Smith, C.; Bevan, M.W. Control of final seed and organ size by the DA1 gene family in Arabidopsis thaliana. Genes Dev. 2008, 22, 1331–1336. [Google Scholar] [CrossRef] [Green Version]
- Xia, T.; Li, N.; Dumenil, J.; Li, J.; Kamenski, A.; Bevan, M.W.; Gao, F.; Li, Y. The ubiquitin receptor DA1 interacts with the E3 ubiquitin ligase DA2 to regulate seed and organ size in Arabidopsis. Plant Cell 2013, 25, 3347–3359. [Google Scholar] [CrossRef] [Green Version]
- Chakravorty, D.; Trusov, Y.; Zhang, W.; Acharya, B.R.; Sheahan, M.B.; McCurdy, D.W.; Assmann, S.M.; Botella, J.R. An atypical heterotrimeric G-protein gamma-subunit is involved in guard cell K(+)-channel regulation and morphological development in Arabidopsis thaliana. Plant J. 2011, 67, 840–851. [Google Scholar] [CrossRef]
- Li, S.; Liu, Y.; Zheng, L.; Chen, L.; Li, N.; Corke, F.; Lu, Y.; Fu, X.; Zhu, Z.; Bevan, M.W.; et al. The plant-specific G protein gamma subunit AGG3 influences organ size and shape in Arabidopsis thaliana. New Phytol. 2012, 194, 690–703. [Google Scholar] [CrossRef]
- Roy Choudhury, S.; Riesselman, A.J.; Pandey, S. Constitutive or seed-specific overexpression of Arabidopsis G-protein gamma subunit 3 (AGG3) results in increased seed and oil production and improved stress tolerance in Camelina sativa. Plant Biotechnol. J. 2014, 12, 49–59. [Google Scholar] [CrossRef]
- Li, N.; Li, Y. Maternal control of seed size in plants. J. Exp. Bot. 2015, 66, 1087–1097. [Google Scholar] [CrossRef] [Green Version]
- Schruff, M.C.; Spielman, M.; Tiwari, S.; Adams, S.; Fenby, N.; Scott, R.J. The AUXIN RESPONSE FACTOR 2 gene of Arabidopsis links auxin signalling, cell division, and the size of seeds and other organs. Development 2006, 133, 251–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riefler, M.; Novak, O.; Strnad, M.; Schmulling, T. Arabidopsis cytokinin receptor mutants reveal functions in shoot growth, leaf senescence, seed size, germination, root development, and cytokinin metabolism. Plant Cell 2006, 18, 40–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choe, S.; Fujioka, S.; Noguchi, T.; Takatsuto, S.; Yoshida, S.; Feldmann, K.A. Overexpression of DWARF4 in the brassinosteroid biosynthetic pathway results in increased vegetative growth and seed yield in Arabidopsis. Plant J. 2001, 26, 573–582. [Google Scholar] [CrossRef] [PubMed]
- Mizukami, Y.; Fischer, R.L. Plant organ size control: AINTEGUMENTA regulates growth and cell numbers during organogenesis. Proc. Natl. Acad. Sci. USA 2000, 97, 942–947. [Google Scholar] [CrossRef] [Green Version]
- Hutchison, C.E.; Li, J.; Argueso, C.; Gonzalez, M.; Lee, E.; Lewis, M.W.; Maxwell, B.B.; Perdue, T.D.; Schaller, G.E.; Alonso, J.M.; et al. The Arabidopsis histidine phosphotransfer proteins are redundant positive regulators of cytokinin signaling. Plant Cell 2006, 18, 3073–3087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuwamoto, R.; Fukuoka, H.; Takahata, Y. GASSHO1 and GASSHO2 encoding a putative leucine-rich repeat transmembrane-type receptor kinase are essential for the normal development of the epidermal surface in Arabidopsis embryos. Plant J. 2008, 54, 30–42. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.Y.; Shi, D.Q.; Jia, P.F.; Tang, J.; Li, H.J.; Liu, J.; Yang, W.C. The Arabidopsis Receptor Kinase ZAR1 Is Required for Zygote Asymmetric Division and Its Daughter Cell Fate. PLoS Genet. 2016, 12, e1005933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, G.Z.; Xue, H.W. Arabidopsis beta-ketoacyl-[acyl carrier protein] synthase i is crucial for fatty acid synthesis and plays a role in chloroplast division and embryo development. Plant Cell 2010, 22, 3726–3744. [Google Scholar] [CrossRef] [Green Version]
- Mu, J.; Tan, H.; Hong, S.; Liang, Y.; Zuo, J. Arabidopsis transcription factor genes NF-YA1, 5, 6, and 9 play redundant roles in male gametogenesis, embryogenesis, and seed development. Mol. Plant 2013, 6, 188–201. [Google Scholar] [CrossRef] [Green Version]
- Vidaurre, D.P.; Ploense, S.; Krogan, N.T.; Berleth, T. AMP1 and MP antagonistically regulate embryo and meristem development in Arabidopsis. Development 2007, 134, 2561–2567. [Google Scholar] [CrossRef] [Green Version]
- Ogawa, E.; Yamada, Y.; Sezaki, N.; Kosaka, S.; Kondo, H.; Kamata, N.; Abe, M.; Komeda, Y.; Takahashi, T. ATML1 and PDF2 Play a Redundant and Essential Role in Arabidopsis Embryo Development. Plant Cell Physiol. 2015, 56, 1183–1192. [Google Scholar] [CrossRef] [Green Version]
- Chymkowitch, P.; Nguea, P.A.; Aanes, H.; Robertson, J.; Klungland, A.; Enserink, J.M. TORC1-dependent sumoylation of Rpc82 promotes RNA polymerase III assembly and activity. Proc. Natl. Acad. Sci. USA 2017, 114, 1039–1044. [Google Scholar] [CrossRef] [Green Version]
- Vernekar, D.V.; Bhargava, P. Yeast Bud27 modulates the biogenesis of Rpc128 and Rpc160 subunits and the assembly of RNA polymerase III. Biochim. Biophys. Acta 2015, 1849, 1340–1353. [Google Scholar] [CrossRef]
- Johnson, K.C.; Yu, Y.; Gao, L.; Eng, R.C.; Wasteneys, G.O.; Chen, X.; Li, X. A partial loss-of-function mutation in an Arabidopsis RNA polymerase III subunit leads to pleiotropic defects. J. Exp. Bot. 2016, 67, 2219–2230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, S.L. Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 2016, 11, 1650–1667. [Google Scholar] [CrossRef]
- Tian, T.; Liu, Y.; Yan, H.; You, Q.; Yi, X.; Du, Z.; Xu, W.; Su, Z. agriGO v2.0: A GO analysis toolkit for the agricultural community, 2017 update. Nucleic Acids Res. 2017, 45, W122–W129. [Google Scholar] [CrossRef]
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stuart, J.M.; Segal, E.; Koller, D.; Kim, S.K. A gene-coexpression network for global discovery of conserved genetic modules. Science 2003, 302, 249–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. cerevisiae | Accession Number | H. sapiens | Accession Number | Arabidopsis thaliana | Gene ID |
|---|---|---|---|---|---|
| RNA Pol III Subunits | RNA Pol III Subunits | RNA Pol III Subunits | |||
| C160 (β-like) | P04051 | HsNRPC1 | AAB86536 | AtNRPC1 | AT5G60040 |
| C128 (β-like) | AAB59324 | HsNRPC2 | AY092084 | AtNRPC2 | AT5G45140 |
| C82 | CAA45072 | HsNRPC3 | NP_006459/ | AtNRPC3 | AT3G49000 |
| XP_034604 | |||||
| C53 | P25441 | HsNRPC4 | AY092086 | AtNRPC4-1 | AT5G09380 |
| AtNRPC4-2 | AT4G25180 | ||||
| C37 | NP_012950 | HsNRPC5 | AY092085 | AtNRPC5 | AT5G49530 |
| C34 | P32910 | HsNRPC6 | NP_006457/ | AtNRPC6 | AT5G23710 |
| XP_009639 | |||||
| C31 | P17890 | HsNRPC7 | AAB63676/ | AtNRPC7-1 | AT4G01590 |
| XP_036456 | AtNRPC7-2 | AT4G35685 | |||
| C25 | P35718 | HsNRPC8 | AY092087 | AtNRPC8 | AT1G06790 |
| C17 | P47076 | HsNRPC9 | AAC25992 | AtNRPC9-1 | AT5G62950 |
| AtNRPC9-2 | AT3G28956 | ||||
| C11 | AAD12060 | HsNRPC10 | NP_057394 | AtNRPC10-1 | AT1G01210 |
| AtNRPC10-2 | AT4G07950 | ||||
| AC40 (α-like) | P07703 | HsNRPAC1 | NP_004866 | AtNRPAC1 | AT1G60620 |
| AC19 (α-like) | P28000 | HsNRPAC2 | NP_057056 | AtNRPAC2 | AT2G29540 |
| ABC27 | P20434 | HsNRPABC1 | P19388 | AtNRPABC1 | AT3G22320 |
| ABC23 (ω-like) | AAA34989 | HsNRPABC2 | P41584 | AtNRPABC2-1 | AT5G51940 |
| AtNRPABC2-2 | AT2G04630 | ||||
| ABC14.5 | CAA37383 | HsNRPABC3 | P52434 | AtNRPABC3-1 | AT1G54250 |
| AtNRPABC3-2 | AT3G59600 | ||||
| ABC10α | AAA64417 | HsNRPABC4 | P53803 | AtNRPABC4 | AT5G41010 |
| ABC10β | P22139 | HsNRPABC5 | P52436 | AtNRPABC5 | AT1G11475 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, H.; Qin, Y.; Xiao, Z.; Liang, K.; Gong, D.; Sun, Q.; Qiu, F. Knockdown NRPC2, 3, 8, NRPABC1 and NRPABC2 Affects RNAPIII Activity and Disrupts Seed Development in Arabidopsis. Int. J. Mol. Sci. 2021, 22, 11314. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222111314
Zhao H, Qin Y, Xiao Z, Liang K, Gong D, Sun Q, Qiu F. Knockdown NRPC2, 3, 8, NRPABC1 and NRPABC2 Affects RNAPIII Activity and Disrupts Seed Development in Arabidopsis. International Journal of Molecular Sciences. 2021; 22(21):11314. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222111314
Chicago/Turabian StyleZhao, Hailiang, Yao Qin, Ziyi Xiao, Kun Liang, Dianming Gong, Qin Sun, and Fazhan Qiu. 2021. "Knockdown NRPC2, 3, 8, NRPABC1 and NRPABC2 Affects RNAPIII Activity and Disrupts Seed Development in Arabidopsis" International Journal of Molecular Sciences 22, no. 21: 11314. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222111314