
Twist1 Influences the Expression of Leading Members of the IL-17 Signaling Pathway in HER2-Positive Breast Cancer Cells

 , , and
, , and
Abstract
:1. Introduction
2. Results
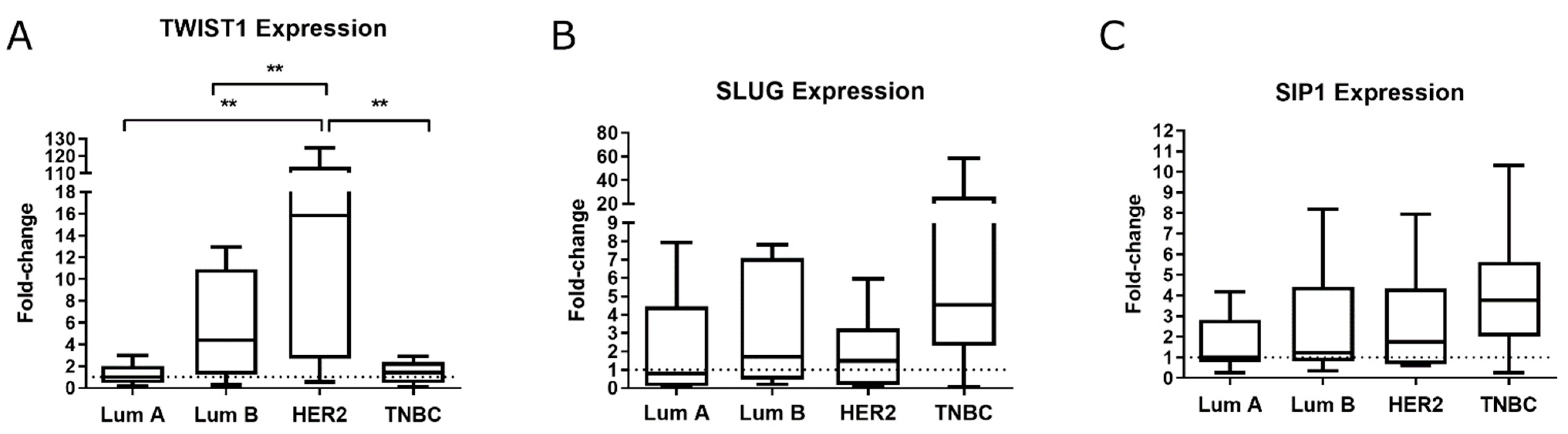
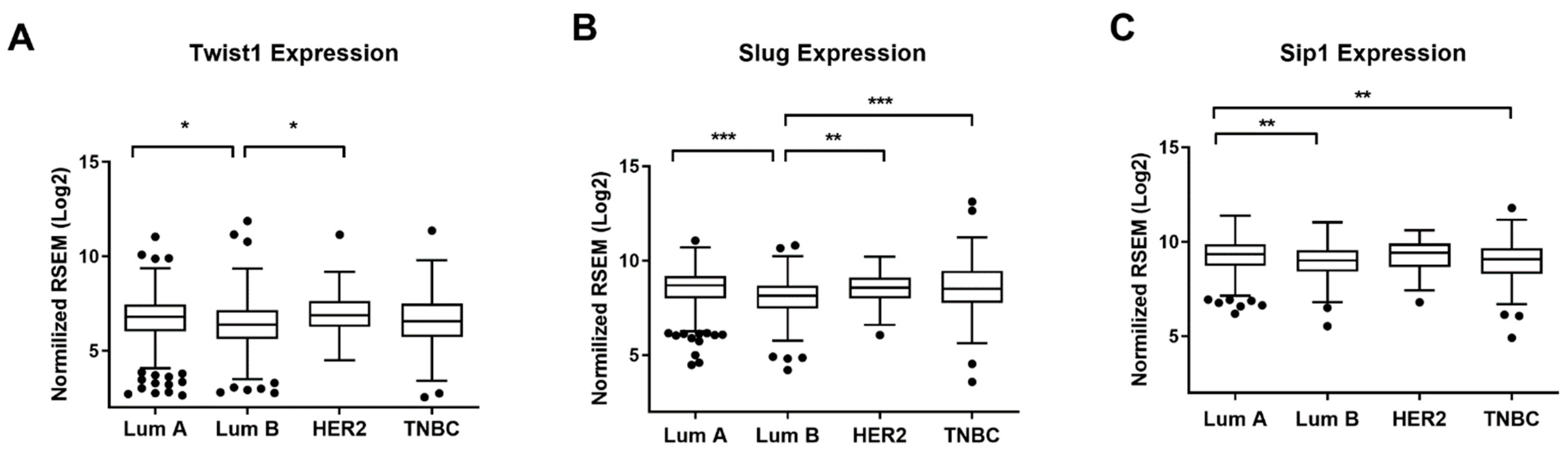
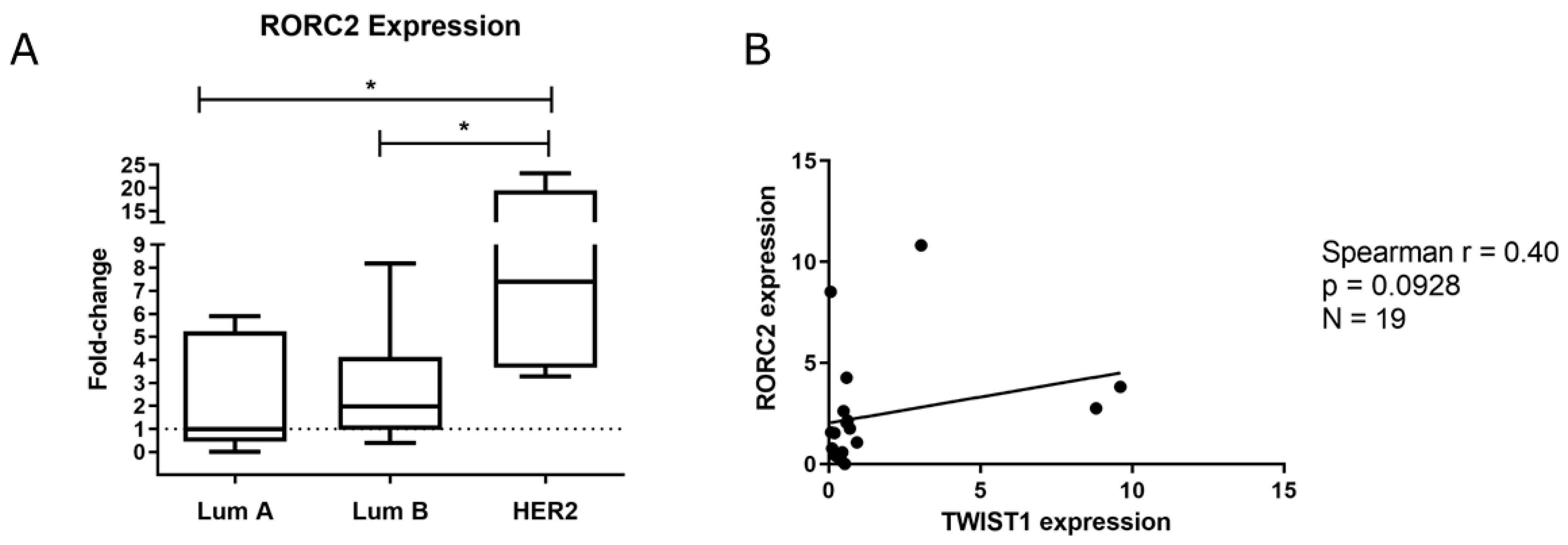
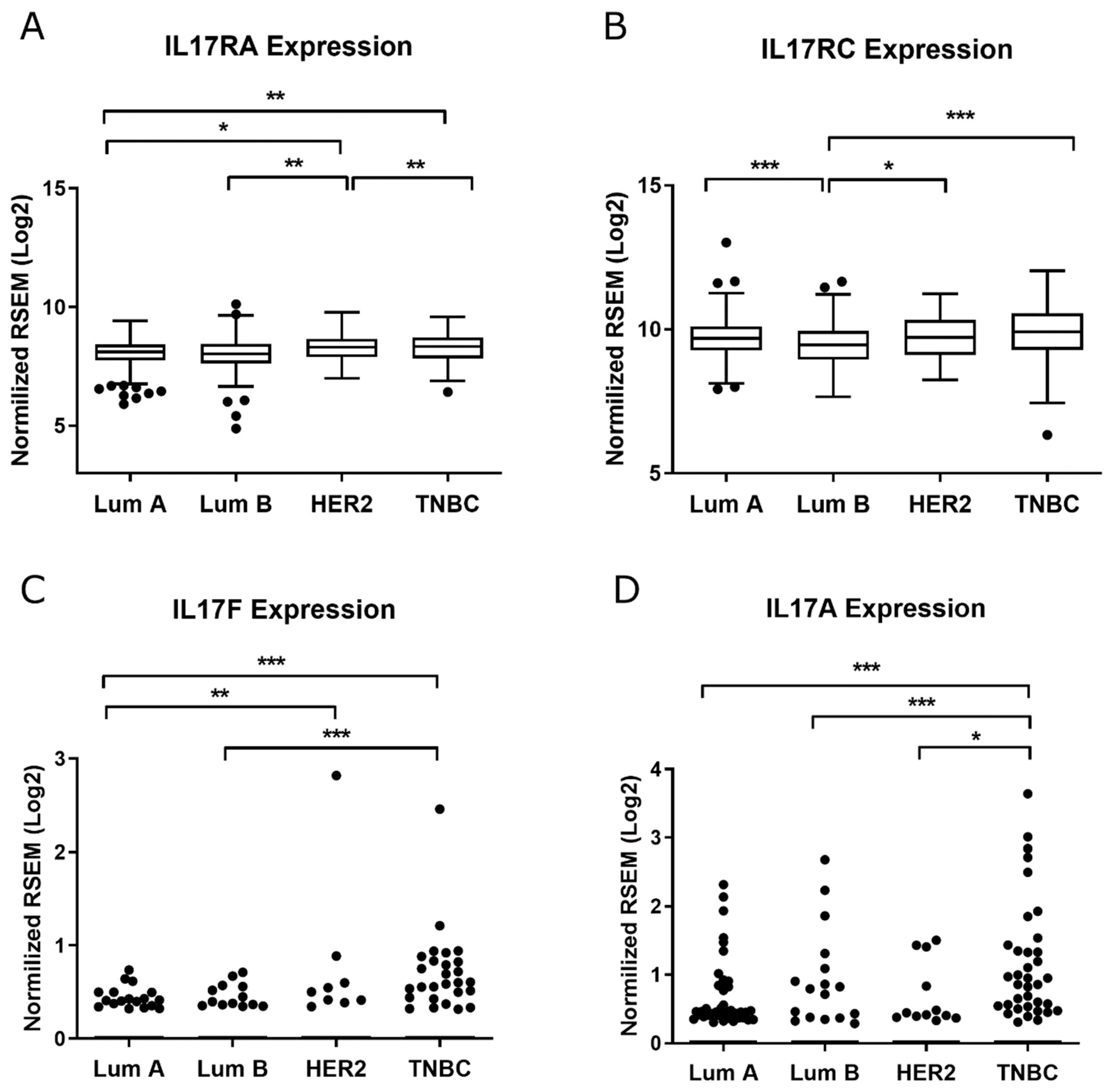
2.1. TWIST1 Is Overexpressed in the HER2+ BC Group
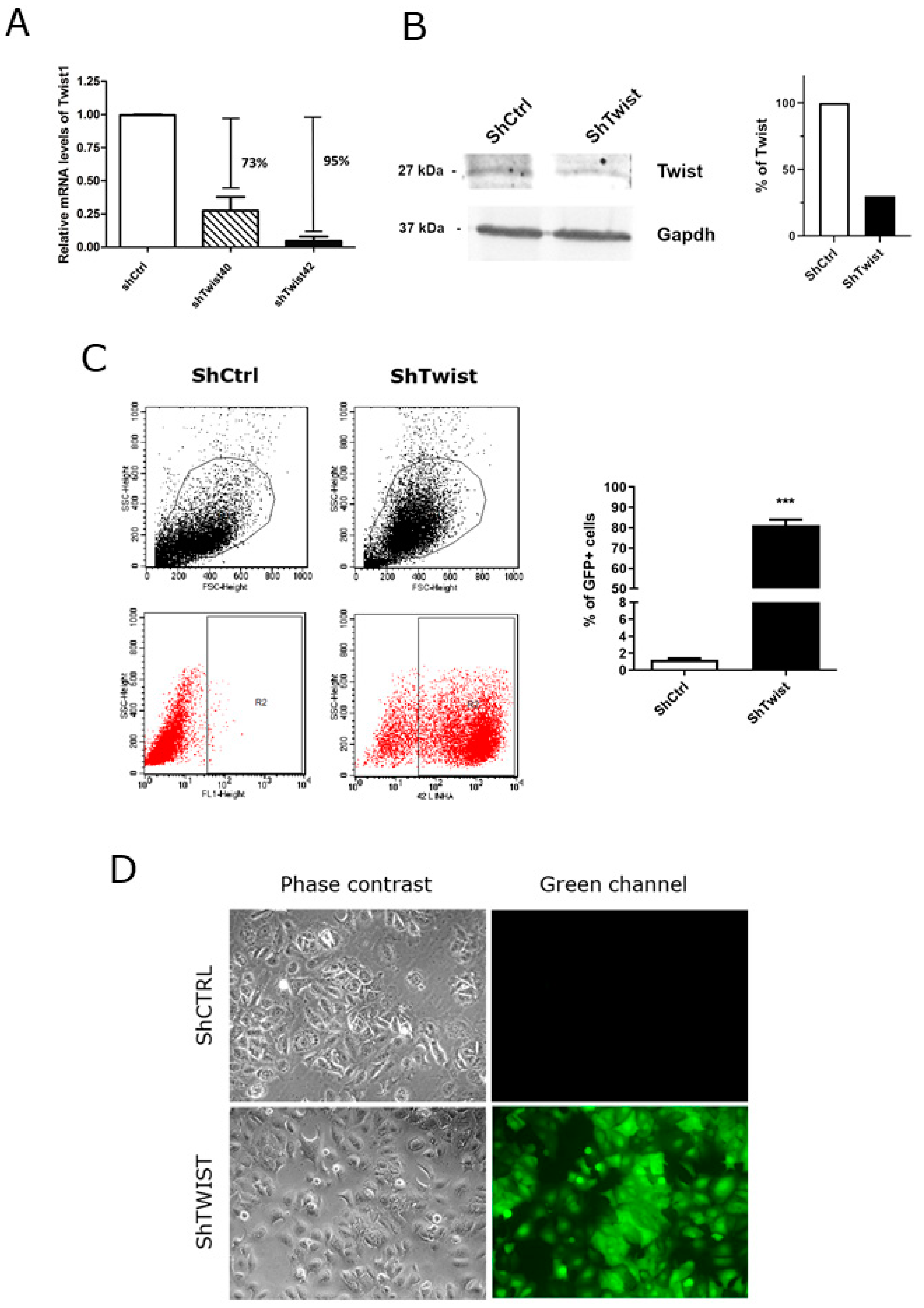
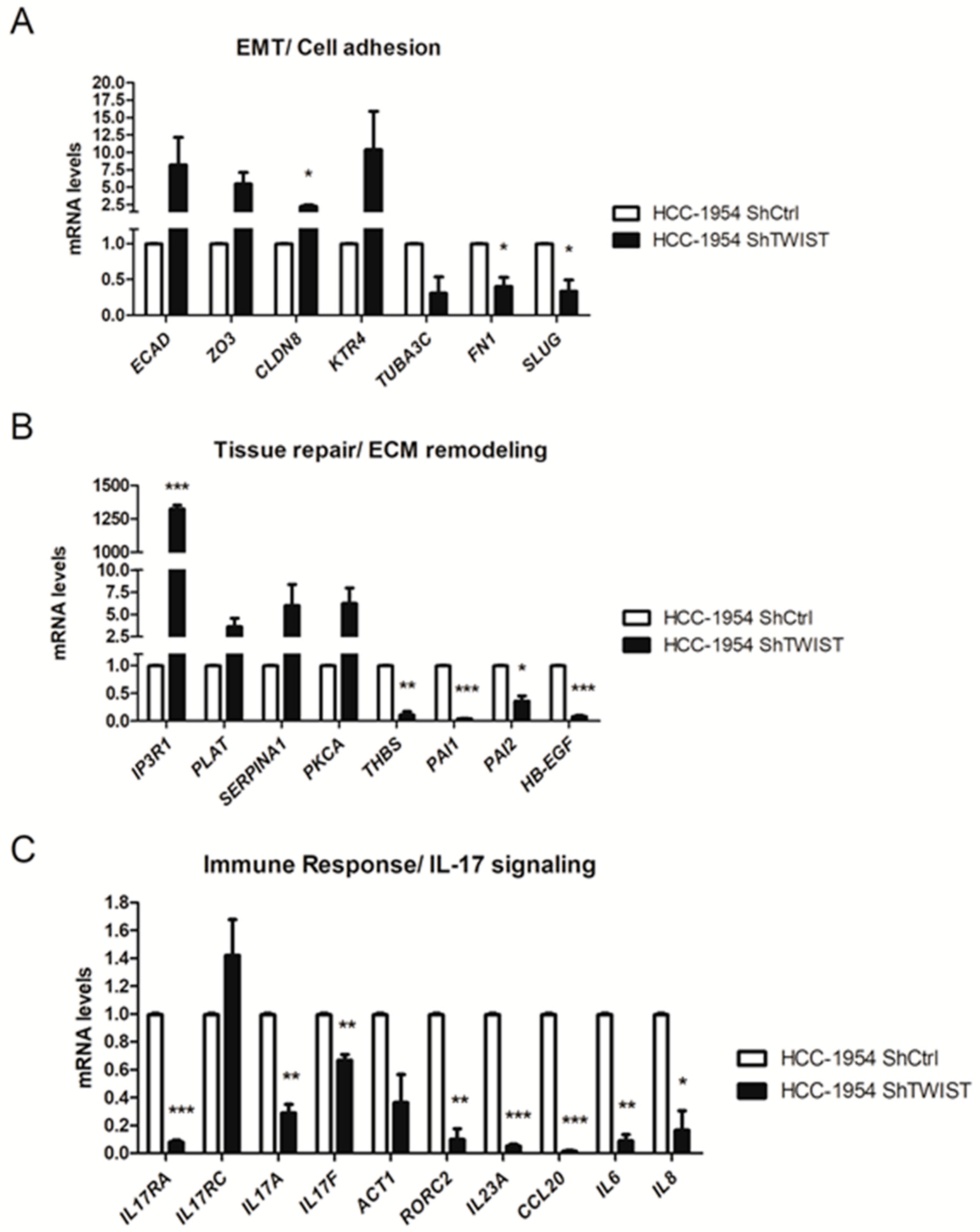
2.2. TWIST1 Knockdown Altered the Gene Expression Profile of Signaling Pathways Relevant to BC
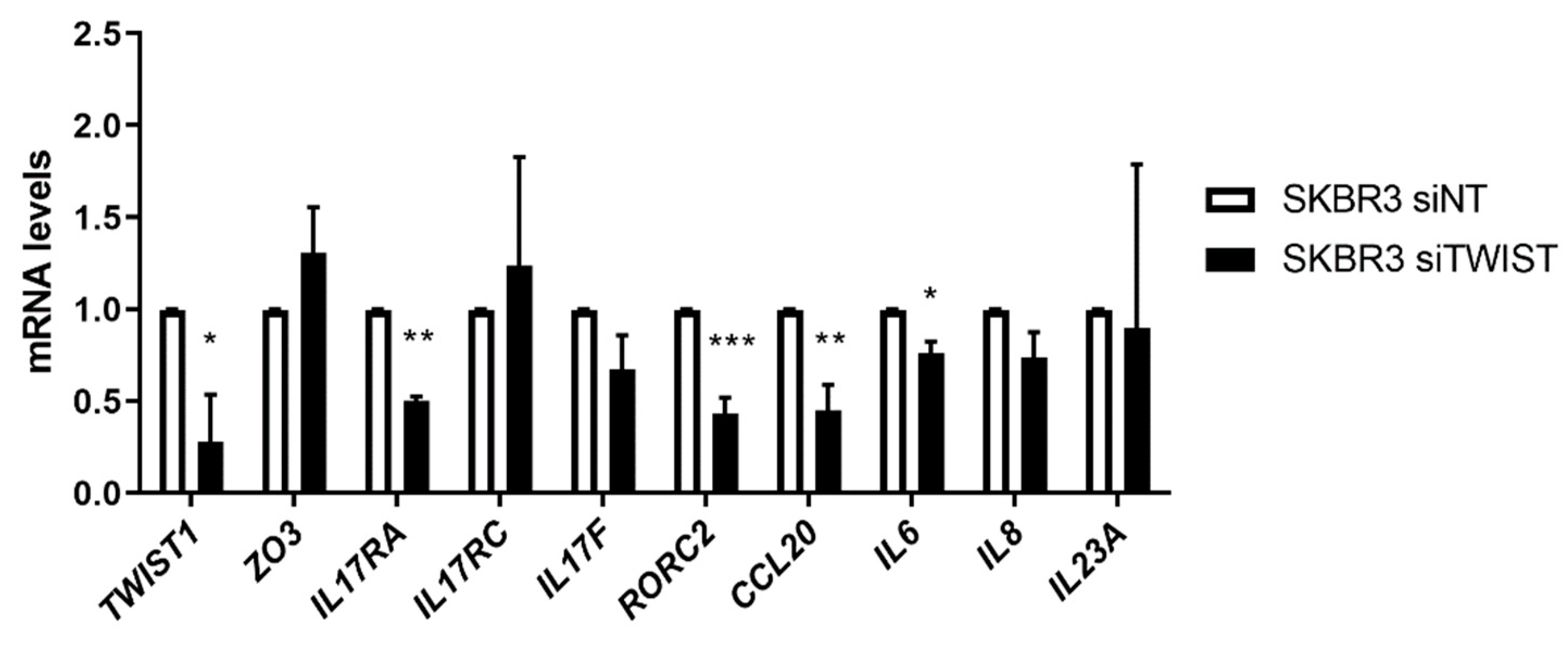
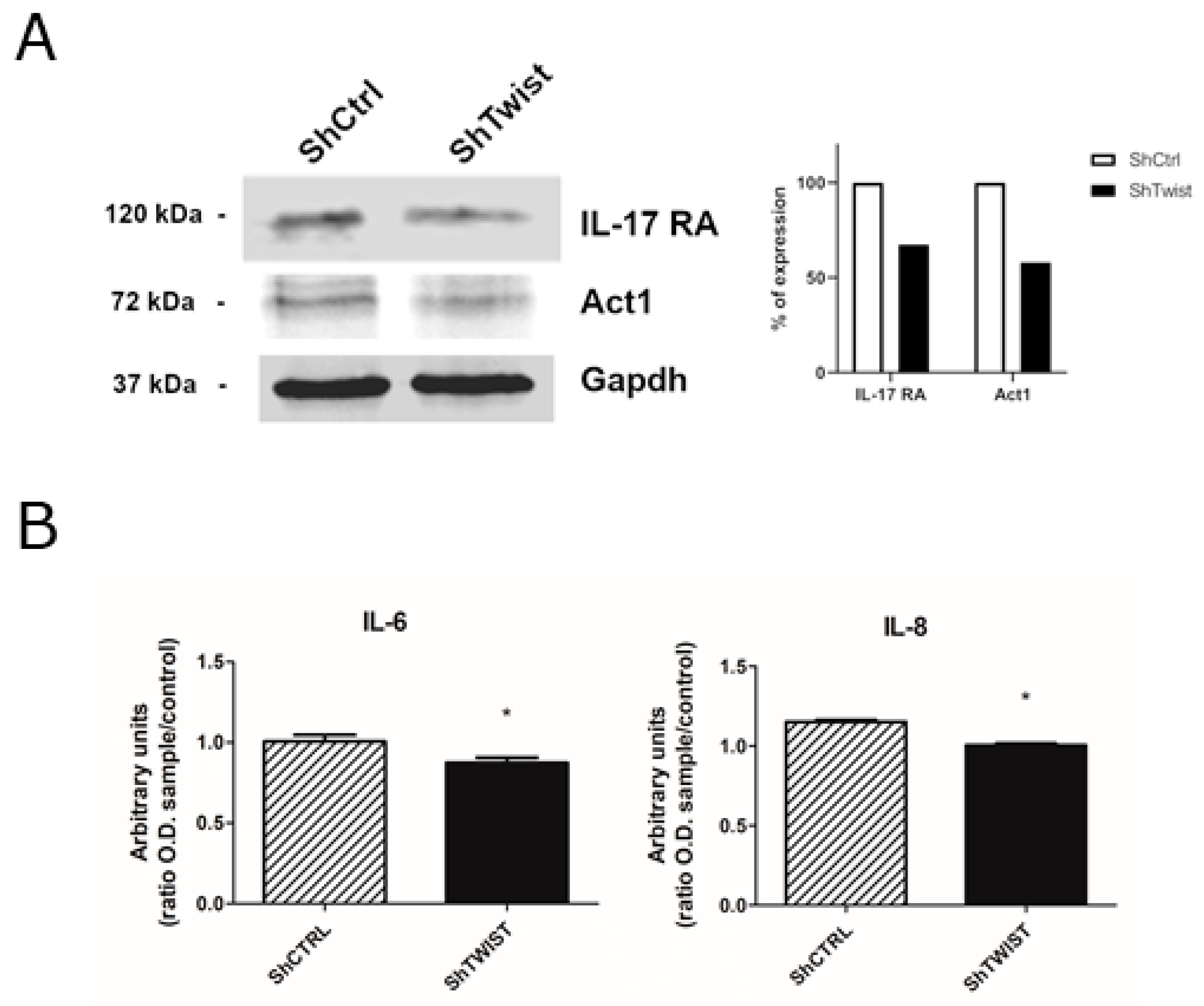
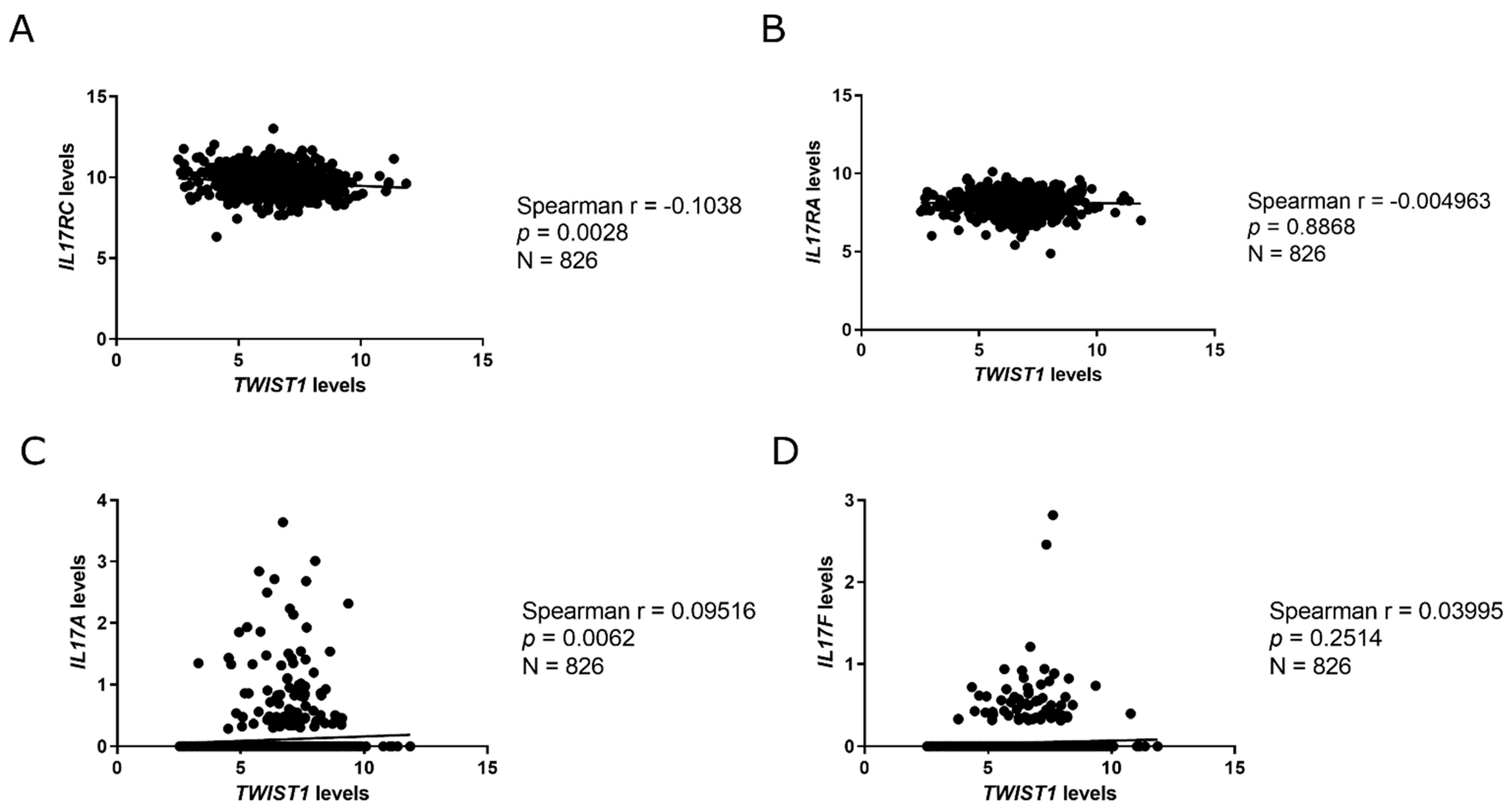
2.3. Twist1 Influences Leading Members of the IL-17 Signaling Pathway
3. Discussion
4. Materials and Methods
4.1. Patient Samples
4.2. Cell Culture and Lentiviral Transduction
4.3. Expression Chip Array Data Analysis
4.4. Real-Time Reverse Transcription Polymerase Chain Reaction (RT-qPCR)
4.5. Immunoblotting
4.6. Flow Cytometry
4.7. Light Phase Contrast and Fluorescence Microscopy
4.8. Enzyme-Linked Immunosorbent Assay (ELISA)
4.9. TCGA Analysis
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2021. CA A Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Voduc, K.D.; Cheang, M.C.; Tyldesley, S.; Gelmon, K.; Nielsen, T.O.; Kennecke, H. Breast cancer subtypes and the risk of local and regional relapse. J. Clin. Oncol. 2010, 28, 1684–1691. [Google Scholar] [CrossRef] [Green Version]
- Polyak, K.; Metzger Filho, O. SnapShot: Breast cancer. Cancer cell 2012, 22, 562. [Google Scholar] [CrossRef]
- Loberg, R.D.; Bradley, D.A.; Tomlins, S.A.; Chinnaiyan, A.M.; Pienta, K.J. The lethal phenotype of cancer: The molecular basis of death due to malignancy. CA A Cancer J. Clin. 2007, 57, 225–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huber, M.A.; Kraut, N.; Beug, H. Molecular requirements for epithelial–mesenchymal transition during tumor progression. Curr. Opin. Cell Biol. 2005, 17, 548–558. [Google Scholar] [CrossRef] [PubMed]
- Kalluri, R.; Weinberg, R.A. The basics of epithelial-mesenchymal transition. J. Clin. Investig. 2009, 119, 1420–1428. [Google Scholar] [CrossRef] [Green Version]
- Zeisberg, M.; Neilson, E.G. Biomarkers for epithelial-mesenchymal transitions. J. Clin. Investig. 2009, 119, 1429–1437. [Google Scholar] [CrossRef] [Green Version]
- Lamouille, S.; Xu, J.; Derynck, R. Molecular mechanisms of epithelial–mesenchymal transition. Nat. Rev. Mol. Cell Biol. 2014, 15, 178–196. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Mani, S.A.; Donaher, J.L.; Ramaswamy, S.; Itzykson, R.A.; Come, C.; Savagner, P.; Gitelman, I.; Richardson, A.; Weinberg, R.A. Twist, a master regulator of morphogenesis, plays an essential role in tumor metastasis. Cell 2004, 117, 927–939. [Google Scholar] [CrossRef] [Green Version]
- Bulzico, D.; de Faria, P.A.S.; Maia, C.B.; de Paula, M.P.; Torres, D.C.; Ferreira, G.M.; Pires, B.R.B.; Hassan, R.; Abdelhay, E.; Vaisman, M. Is there a role for epithelial-mesenchymal transition in adrenocortical tumors? Endocrine 2017, 58, 276–288. [Google Scholar] [CrossRef]
- Kong, Y.H.; Syed Zanaruddin, S.N.; Lau, S.H.; Ramanathan, A.; Kallarakkal, T.G.; Vincent-Chong, V.K.; Wan Mustafa, W.M.; Abraham, M.T.; Abdul Rahman, Z.A.; Zain, R.B. Co-expression of TWIST1 and ZEB2 in oral squamous cell carcinoma is associated with poor survival. PLoS ONE 2015, 10, e0134045. [Google Scholar] [CrossRef] [Green Version]
- Martin, T.A.; Goyal, A.; Watkins, G.; Jiang, W.G. Expression of the transcription factors snail, slug, and twist and their clinical significance in human breast cancer. Ann. Surg. Oncol. 2005, 12, 488–496. [Google Scholar] [CrossRef] [PubMed]
- Soini, Y.; Tuhkanen, H.; Sironen, R.; Virtanen, I.; Kataja, V.; Auvinen, P.; Mannermaa, A.; Kosma, V.-M. Transcription factors zeb1, twist and snai1 in breast carcinoma. BMC Cancer 2011, 11, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Hu, B.; Qin, L.; Zhao, L.; Wang, Q.; Wang, Q.; Xu, Y.; Jiang, J. SRC-1 and Twist1 expression positively correlates with a poor prognosis in human breast cancer. Int. J. Biol. Sci. 2014, 10, 396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, M.H.; Chen, C.L.; Chau, G.Y.; Chiou, S.H.; Su, C.W.; Chou, T.Y.; Peng, W.L.; Wu, J.C. Comprehensive analysis of the independent effect of twist and snail in promoting metastasis of hepatocellular carcinoma. Hepatology 2009, 50, 1464–1474. [Google Scholar] [CrossRef] [PubMed]
- Yan-Qi, Z.; Xue-Yan, G.; Shuang, H.; Yu, C.; Fu-Lin, G.; Fei-Hu, B.; Shi-Ren, S.; Xu-Feng, W.; Jie, D.; Dai-Ming, F. Expression and significance of TWIST basic helix-loop-helix protein over-expression in gastric cancer. Pathology 2007, 39, 470–475. [Google Scholar] [CrossRef] [PubMed]
- Pires, B.R.; Mencalha, A.L.; Ferreira, G.M.; de Souza, W.F.; Morgado-Diaz, J.A.; Maia, A.M.; Correa, S.; Abdelhay, E.S. NF-kappaB Is involved in the regulation of EMT genes in breast cancer cells. PLoS ONE 2017, 12, e0169622. [Google Scholar] [CrossRef] [Green Version]
- Unutmaz, D. RORC2: The Master of Human Th17 Cell Programming. Eur. J. Immunol. 2009, 39, 1452–1455. [Google Scholar] [CrossRef]
- Thisse, B.; Messal, M.E.; Perrin-Schmitt, F. The twist gene: Isolation of a drosophila zygotle gene necessary for the establishment of dorsoventral pattern. Nucleic Acids Res. 1987, 15, 3439–3453. [Google Scholar] [CrossRef]
- El Ghouzzi, V.; Le Merrer, M.; Perrin-Schmitt, F.; Lajeunie, E.; Benit, P.; Renier, D.; Bourgeois, P.; Bolcato-Bellemin, A.-L.; Munnich, A.; Bonaventure, J. Mutations of the TWIST gene in the Saethre-Chotzene syndrome. Nat. Genet. 1997, 15, 42–46. [Google Scholar] [CrossRef]
- Taube, J.H.; Herschkowitz, J.I.; Komurov, K.; Zhou, A.Y.; Gupta, S.; Yang, J.; Hartwell, K.; Onder, T.T.; Gupta, P.B.; Evans, K.W. Core epithelial-to-mesenchymal transition interactome gene-expression signature is associated with claudin-low and metaplastic breast cancer subtypes. Proc. Natl. Acad. Sci. USA 2010, 107, 15449–15454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiao, W.; Jia, Z.; Liu, H.; Liu, Q.; Zhang, T.; Guo, W.; Li, P.; Deng, M.; Li, S. Prognostic and clinicopathological value of Twist expression in breast cancer: A meta-analysis. PLoS ONE 2017, 12, e0186191. [Google Scholar]
- Byun, J.S.; Singhal, S.K.; Park, S.; Yi, D.I.; Yan, T.; Caban, A.; Jones, A.; Mukhopadhyay, P.; Gil, S.M.; Hewitt, S.M. Racial differences in the association between luminal master regulator gene expression levels and breast cancer survival. Clin. Cancer Res. 2020, 26, 1905–1914. [Google Scholar] [CrossRef] [Green Version]
- Romero-Cordoba, S.L.; Salido-Guadarrama, I.; Rebollar-Vega, R.; Bautista-Piña, V.; Dominguez-Reyes, C.; Tenorio-Torres, A.; Villegas-Carlos, F.; Fernández-López, J.C.; Uribe-Figueroa, L.; Alfaro-Ruiz, L. Comprehensive omic characterization of breast cancer in Mexican-Hispanic women. Nat. Commun. 2021, 12, 2245. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-W.; Lee, N.K.; Kim, J.H.; Kang, M.S.; Yoo, H.Y.; Kim, H.H.; Um, S.H.; Kim, S.-H. Twist1 causes the transcriptional repression of claudin-4 with prognostic significance in esophageal cancer. Biochem. Biophys. Res. Commun. 2012, 423, 454–460. [Google Scholar] [CrossRef] [PubMed]
- Vesuna, F.; Lisok, A.; Kimble, B.; Domek, J.; Kato, Y.; van der Groep, P.; Artemov, D.; Kowalski, J.; Carraway, H.; van Diest, P. Twist contributes to hormone resistance in breast cancer by downregulating estrogen receptor-α. Oncogene 2012, 31, 3223–3234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexander, N.R.; Tran, N.L.; Rekapally, H.; Summers, C.E.; Glackin, C.; Heimark, R.L. N-cadherin gene expression in prostate carcinoma is modulated by integrin-dependent nuclear translocation of Twist1. Cancer Res. 2006, 66, 3365–3369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, J.; Chen, S.; Han, J.-X.; Qian, B.; Wang, X.-R.; Zhong, W.-L.; Qin, Y.; Zhang, H.; Gao, W.-F.; Lei, Y.-Y. Twist1 regulates vimentin through Cul2 circular RNA to promote EMT in hepatocellular carcinoma. Cancer Res. 2018, 78, 4150–4162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z.; Zhang, X.; Gang, H.; Li, X.; Li, Z.; Wang, T.; Han, J.; Luo, T.; Wen, F.; Wu, X. Up-regulation of gastric cancer cell invasion by Twist is accompanied by N-cadherin and fibronectin expression. Biochem. Biophys. Res. Commun. 2007, 358, 925–930. [Google Scholar] [CrossRef] [PubMed]
- Casas, E.; Kim, J.; Bendesky, A.; Ohno-Machado, L.; Wolfe, C.J.; Yang, J. Snail2 is an essential mediator of Twist1-induced epithelial mesenchymal transition and metastasis. Cancer Res. 2011, 71, 245–254. [Google Scholar] [CrossRef] [Green Version]
- Hu, L.; Roth, J.M.; Brooks, P.; Ibrahim, S.; Karpatkin, S. Twist is required for thrombin-induced tumor angiogenesis and growth. Cancer Res. 2008, 68, 4296–4302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mironchik, Y.; Winnard, P.T.; Vesuna, F.; Kato, Y.; Wildes, F.; Pathak, A.P.; Kominsky, S.; Artemov, D.; Bhujwalla, Z.; Van Diest, P. Twist overexpression induces in vivo angiogenesis and correlates with chromosomal instability in breast cancer. Cancer Res. 2005, 65, 10801–10809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okamura, H.; Yoshida, K.; Haneji, T. Negative regulation of TIMP1 is mediated by transcription factor TWIST1. Int. J. Oncol. 2009, 35, 181–186. [Google Scholar] [PubMed] [Green Version]
- Merkhofer, E.C.; Cogswell, P.; Baldwin, A.S. Her2 activates NF-κB and induces invasion through the canonical pathway involving IKKα. Oncogene 2010, 29, 1238–1248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabre, J.; Giustiniani, J.; Garbar, C.; Antonicelli, F.; Merrouche, Y.; Bensussan, A.; Bagot, M.; Al-Dacak, R. Targeting the tumor microenvironment: The protumor effects of IL-17 related to cancer type. Int. J. Mol. Sci. 2016, 17, 1433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaffen, S.L. An overview of IL-17 function and signaling. Cytokine 2008, 43, 402–407. [Google Scholar] [CrossRef] [Green Version]
- Jin, W.; Dong, C. IL-17 cytokines in immunity and inflammation. Emerg. Microbes Infect. 2013, 2, 1–5. [Google Scholar] [CrossRef]
- Chang, S.H.; Park, H.; Dong, C. Act1 adaptor protein is an immediate and essential signaling component of interleukin-17 receptor. J. Biol. Chem. 2006, 281, 35603–35607. [Google Scholar] [CrossRef] [Green Version]
- Pham, D.; Walline, C.C.; Hollister, K.; Dent, A.L.; Blum, J.S.; Firulli, A.B.; Kaplan, M.H. The transcription factor Twist1 limits T helper 17 and T follicular helper cell development by repressing the gene encoding the interleukin-6 receptor α chain. J. Biol. Chem. 2013, 288, 27423–27433. [Google Scholar] [CrossRef] [Green Version]
- Huang, Q.; Han, J.; Fan, J.; Duan, L.; Guo, M.; Lv, Z.; Hu, G.; Chen, L.; Wu, F.; Tao, X. IL-17 induces EMT via Stat3 in lung adenocarcinoma. Am. J. Cancer Res. 2016, 6, 440. [Google Scholar]
- Cao, J.; Wang, X.; Dai, T.; Wu, Y.; Zhang, M.; Cao, R.; Zhang, R.; Wang, G.; Jiang, R.; Zhou, B.P. Twist promotes tumor metastasis in basal-like breast cancer by transcriptionally upregulating ROR1. Theranostics 2018, 8, 2739. [Google Scholar] [CrossRef]
- Klemm, F.; Bleckmann, A.; Siam, L.; Chuang, H.; Rietkötter, E.; Behme, D.; Schulz, M.; Schaffrinski, M.; Schindler, S.; Trümper, L. β-catenin-independent WNT signaling in basal-like breast cancer and brain metastasis. Carcinogenesis 2011, 32, 434–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaffen, S.L. The role of interleukin-17 in the pathogenesis of rheumatoid arthritis. Curr. Rheumatol. Rep. 2009, 11, 365. [Google Scholar] [CrossRef] [PubMed]
- Krueger, J.G.; Fretzin, S.; Suárez-Fariñas, M.; Haslett, P.A.; Phipps, K.M.; Cameron, G.S.; McColm, J.; Katcherian, A.; Cueto, I.; White, T. IL-17A is essential for cell activation and inflammatory gene circuits in subjects with psoriasis. J. Allergy Clin. Immunol. 2012, 130, 145–154. [Google Scholar] [CrossRef] [Green Version]
- Pan, B.; Shen, J.; Cao, J.; Zhou, Y.; Shang, L.; Jin, S.; Cao, S.; Che, D.; Liu, F.; Yu, Y. Interleukin-17 promotes angiogenesis by stimulating VEGF production of cancer cells via the STAT3/GIV signaling pathway in non-small-cell lung cancer. Sci. Rep. 2015, 5, 16053. [Google Scholar] [CrossRef] [Green Version]
- Welte, T.; Zhang, X.H.-F. Interleukin-17 could promote breast cancer progression at several stages of the disease. Mediat. Inflamm. 2015, 2015, 804347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joerger, M.; Finn, S.P.; Cuffe, S.; Byrne, A.T.; Gray, S.G. The IL-17-Th1/Th17 pathway: An attractive target for lung cancer therapy? Expert Opin. Ther. Targets 2016, 20, 1339–1356. [Google Scholar] [CrossRef] [PubMed]
- Giuliano, A.E.; Edge, S.B.; Hortobagyi, G.N. of the AJCC cancer staging manual: Breast cancer. Ann. Surg. Oncol. 2018, 25, 1783–1785. [Google Scholar] [CrossRef] [PubMed]
- Hammond, M.E.H.; Hayes, D.F.; Dowsett, M.; Allred, D.C.; Hagerty, K.L.; Badve, S.; Fitzgibbons, P.L.; Francis, G.; Goldstein, N.S.; Hayes, M. American society of clinical oncology/college of American pathologists guideline recommendations for immunohistochemical testing of estrogen and progesterone receptors in breast cancer (unabridged version). Arch. Pathol. Lab. Med. 2010, 134, e48–e72. [Google Scholar] [CrossRef]
- Wolff, A.C.; Hammond, M.E.H.; Hicks, D.G.; Dowsett, M.; McShane, L.M.; Allison, K.H.; Allred, D.C.; Bartlett, J.M.; Bilous, M.; Fitzgibbons, P. Recommendations for human epidermal growth factor receptor 2 testing in breast cancer: American society of clinical oncology/college of american pathologists clinical practice guideline update. Arch. Pathol. Lab. Med. 2014, 138, 241–256. [Google Scholar] [CrossRef] [Green Version]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Functional Enrichment Analysis * | Gene # | |
|---|---|---|
| Upregulated | Downregulated | |
| EMT/Cell adhesion | ZO3, CLDN8, KTR4 | TUBA3C, FN1, SLUG, HMGA2 |
| ECM remodeling/Blood coagulation | IP3R1, PLAT, SERPINA1, PKCA, ITGB3, GNAI2 | HBEGF, PAI1, PAI2 |
| Immune response/Th17-derived cytokines | IL23A, IL6, IL8, CCL20, CSF2 | |
| Clinical Characteristic | n = 46 | % = 100 |
|---|---|---|
| Age, yr | ||
| Early onset (20–50) Late onset (51–80) | 17 29 | 37.0 63.0 |
| Female, n | 46 | 100 |
| IHC-based molecular classification * | ||
| Luminal A Luminal B HER2-enriched Triple Negative | 16 11 8 11 | 34.8 23.9 17.4 23.9 |
| Tumor histological type | ||
| Invasive ductal carcinoma | 46 | 100 |
| Histological grade | ||
| Grade 1 Grade 2 Grade 3 | 3 8 35 | 6.5 17.4 76.1 |
| TNM staging | ||
| I/II III/IV | 31 15 | 67.4 32.6 |
| Tumor largest diameter, cm | ||
| <2 2–5 >5 | 9 35 2 | 19.6 76.1 4.3 |
| Lymph node status, n | ||
| Stage 1 (negative) Stage 2 (1–3 LN) Stage 3 (>3 LN) | 17 20 9 | 37.0 43.5 19.6 |
| Distant metastasis, n | ||
| Presence Absence | 10 36 | 21.7 78.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pires, B.R.B.; Binato, R.; Ferreira, G.M.; Corrêa, S.; Du Rocher, B.; Bulzico, D.; Crocamo, S.; dos Santos, E.C.; Lima, L.G.; Abdelhay, E. Twist1 Influences the Expression of Leading Members of the IL-17 Signaling Pathway in HER2-Positive Breast Cancer Cells. Int. J. Mol. Sci. 2021, 22, 12144. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222212144
Pires BRB, Binato R, Ferreira GM, Corrêa S, Du Rocher B, Bulzico D, Crocamo S, dos Santos EC, Lima LG, Abdelhay E. Twist1 Influences the Expression of Leading Members of the IL-17 Signaling Pathway in HER2-Positive Breast Cancer Cells. International Journal of Molecular Sciences. 2021; 22(22):12144. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222212144
Chicago/Turabian StylePires, Bruno R. B., Renata Binato, Gerson M. Ferreira, Stephany Corrêa, Bárbara Du Rocher, Daniel Bulzico, Susanne Crocamo, Everton Cruz dos Santos, Luize G. Lima, and Eliana Abdelhay. 2021. "Twist1 Influences the Expression of Leading Members of the IL-17 Signaling Pathway in HER2-Positive Breast Cancer Cells" International Journal of Molecular Sciences 22, no. 22: 12144. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222212144